

Abstract
The systematic investigation of susceptibility-induced contrast in MRI is important to better interpret the influence of microvascular and microcellular morphology on DSC-MRI derived perfusion data. Recently, a novel computational approach called the Finite Perturber Method (FPM), which enables the study of susceptibility-induced contrast in MRI arising from arbitrary microvascular morphologies in 3D has been developed. However, the FPM has lower efficiency in simulating water diffusion especially for complex tissues. In this work, an improved computational approach that combines the FPM with a matrix-based finite difference method (FDM), which we call the Finite Perturber the Finite Difference Method (FPFDM), has been developed in order to efficiently investigate the influence of vascular and extravascular morphological features on susceptibility-induced transverse relaxation. The current work provides a framework for better interpreting how DSC-MRI data depend on various phenomena, including contrast agent leakage in cancerous tissues and water diffusion rates. In addition, we illustrate using simulated and micro-CT extracted tissue structures the improved FPFDM along with its potential applications and limitations.
Introduction
The passage of paramagnetic or superparamagnetic contrast agents (CA) through brain tissue induces a measurable drop in T2- or T2 *-weighted MR signal [1] that forms the basis for dynamic susceptibility contrast (DSC) MRI. When combined with appropriate kinetic models, DSC-MRI can be used to measure hemodynamic parameters quantitatively, such as blood flow, blood volume and mean transit time [2]. This imaging approach relies upon MR signal relaxation enhancement created by CA-induced susceptibility differences between tissue compartments, such as blood vessels and the surrounding extravascular space. The assessment of tumor perfusion parameters using DSC-MRI has proven to be useful for characterizing tumor grade [3]–[9] and treatment response [10]–[14]. Despite its increased use in brain tumor and stroke patients, accurate calculation of perfusion parameters using DSC-MRI relies on two assumptions: 1) a linear relationship, with a spatially uniform rate constant termed the vascular susceptibility calibration factor (kp), exists between CA concentration and the measured transverse relaxation rate change [15]; and 2) the blood-brain barrier (BBB) is intact, so that contrast agent remains intravascular and can be treated as a nondiffusible tracer [2]. However, heterogeneous distributions of blood vessels within tissue and the dependence of susceptibility field gradients on vascular geometry may yield spatially variant kp values across tissue. Furthermore, leakage of contrast agent in tumors with BBB disruption causes additional T1 and T2 * shortening with subsequent distortion of DSC-MRI signal profiles [16]–[20]. Improved characterization of these potential confounding factors could shed new insights into the biophysical basis of DSC-MRI signals and direct future improvements in acquisition and post-processing strategies.
In order to better understand susceptibility-based image contrast, several theoretical [21]–[25] and computational models using fixed perturber geometry (e.g., cylinders or spheres) [25]–[32] have been proposed. To address the limited ability of these computational models to represent the complex vascular morphologies in both normal brain and tumors, Pathak et al introduced the Finite Perturber Method (FPM) for simulating susceptibility-based contrast for arbitrary microvessel geometries [33] and evaluating differences in kp for normal brain and tumor [34]. The FPM uses estimated magnetic field perturbations to calculate MR signal by simulating proton diffusion and phase accumulation using conventional time consuming Monte Carlo methods.
For realistic complex tissues, the MC method needs to track the diffusion of a large number of spins to capture complex structural features, which in turn can increase the computation time. As an alternative, the Bloch-Torrey partial differential equation describing the transverse magnetization can be directly solved using finite difference methods (FDM). This approach has been previously shown to improve the computational efficiency of such simulations [35], [36] and used to explore water diffusion in MRI and to aid the interpretation of diffusion-weighted imaging measures and their dependence on the morphology of biological structures such as those found in tumors.
In this study, we propose to evaluate the combination of the finite pertuber and finite difference methods, termed the FPFDM, as a tool for modeling susceptibility based contrast mechanisms. Such an approach leverages the strengths of the FPM, for computing magnetic field perturbations for arbitrarily shaped structures, and the FDM, for efficiently computing the resulting MRI signal evolution. The accuracy of the FPFDM is validated by comparison to traditional Monte Carlo methods. The potential of the FPFDM to compute DSC-MRI signals arising from realistic three-dimensional cellular and vascular models as well as micro-CT based renal angiograms is demonstrated. Going forward, the FPFDM provides a useful tool with which to investigate the influence of vascular morphology, contrast agent kinetics and extravasation on DSC-MRI signals.
Methods
In this section, we first describe a new approach for generating more realistic, three-dimensional tissue structures that can be used for the systematic investigation of DSC-MRI signals arising from heterogeneous tissues. We then describe the computation of the appropriate magnetic field perturbations and the associated MR signal evolution, including the influence of water diffusion, using the FPFDM.
1. Tissue Structures
Tissue structures consisting of cells, vessels or both were simulated in a 3D volume sampled with N3 voxels. The motivation for exploring whether cellular properties influence DSC-MRI data originates from previous reports demonstrating that contrast agent extravasation and compartmentalization around cells can induce measurable and dynamic changes in gradient echo acquired signals [16]–[18], [37]. While spheres have been used extensively for evaluating susceptibility-based contrast mechanisms they poorly represent in vivo cellular distribution and shape. In particular, packed spheres intrinsically provide no means for modeling orientation heterogeneity and are unable to achieve cellular densities that approximate those found in vivo. To overcome these limitations we explore here the use of randomly packed ellipsoids [38]. Modeling cells as ellipsoids enables the systematic investigation of several features relevant to DSC-MRI including ellipsoid orientation heterogeneity, volume, aspect ratio and higher packing fractions. For completeness, we compare results from randomly distributed spheres, closely-packed spheres on a face centered cubic (FCC) grid and randomly packed ellipsoids.
Typically, randomly oriented cylinders are used to explore susceptibility contrast mechanisms [25]–[32]. More recently, several groups have employed the use of microvascular angiograms in order to better model in vivo conditions [33], [39], [40]. In order to provide a framework that mimics in vivo conditions but also enables the systematic investigation of the influence of vascular features on DSC-MRI data we explored the use of fractal tree based vascular networks [41], [42]. Starting with an initial cylindrical segment representing an arterial vessel, the vascular tree was created using bifurcation at each junction into smaller daughter segments and a target vascular volume fraction (2%) was used to terminate the fractal tree development. At each junction the diameter of each daughter vessels was calculated using Murray’s law [43] and given some degree of randomness along with the branching angles to create tumor-like heterogeneous structures.
All simulated tissue structures were represented by a binary function V(x,y,z) indicating whether a given point within the simulation volume lies inside or outside the simulated compartments, i.e.:
| (1) |
To further illustrate the versatility of the FPFDM, in addition to the simulated structures, micro-CT was used to create a three-dimensional rendering of a murine kidney vasculature perfused with Microfil (MV–122, Flow Tech). Following perfusion and fixation in 10% neutral buffered formalin, the kidney was scanned in a microCT50 (Scanco Medical AG, Brüttisellen Switzerland). Cross-sectional images of the entire kidney were acquired with an isotropic voxel size of 5.0 µm using an energy of 55 kVp, 200 µA intensity, 700 msec sample time, and 1000 projections per rotation using the manufacturers 1200 mg HA/ccm beam hardening correction algorithm in a 10.24 mm field of view. Using the manufacturer’s software, we assembled individual slices into a z-stack and contrast-filled vessels were segmented from soft tissue by applying a threshold of 260 mg HA/ccm (determined by calibration against a hydroxyapatite phantom) and a three-dimensional Gaussian noise filter with sigma 2.3 and support of 4. The resulting binary three-dimensional reconstruction of the vasculature was then subdivided into MR voxel size sections using in-house Matlab codes (Mathworks, Natick, MA) and used as an input for the FPFDM simulation.
2. Computation of Magnetic Field Perturbations
The magnetic field perturbations induced by susceptibility variations within the simulated volume were computed using the FPM [33]. To calculate the magnetic field shift at a given point, the FPM breaks the structure into numerous small cubic perturbers and the contribution to the magnetic field shift due to each perturber is calculated independently. The total magnetic field shift is then evaluated as the sum of the magnetic field shifts from all of the perturbers. As computational input the FPM requires a tabular listing of the vascular/cellular structure V(x,y,z), the B0 field components, and the susceptibility difference (Δχ) between tissue compartments. The magnetic field shift computed by the FPM is:
| (2) |
where V(x,y,z) is a binary tissue structure, 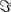 represents the Fourier transform and
represents the Fourier transform and 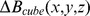 is the magnetic field change arising from a single finite perturber approximated by the magnetic field shift of an embedded sphere within the cube weighted by a volume factor 6/π, and is described by Eq. [3]:
is the magnetic field change arising from a single finite perturber approximated by the magnetic field shift of an embedded sphere within the cube weighted by a volume factor 6/π, and is described by Eq. [3]:
| (3) |
where R is one-half the side length of the cube, Δχ is the susceptibility difference between tissue compartments, and r and θ indicate the distance from the center of the cube and the angle with respect to B0 of the magnetic field calculation point, respectively. The accuracy of this method has been tested using simple geometries with known theoretical field perturbations [33].
3. Computation of MR Signal
Estimation of the MR signal relaxation induced by the inhomogeneous field gradients requires simulation of proton diffusion. To track the Brownian motion of thousands of protons over a large number of time steps and calculate their phase accumulation, a Monte Carlo (MC) simulation is frequently used [25]–[31], [33]. The MC method is potentially time consuming for complex tissue structures because in order to accurately calculate the phase distribution it must track a large number of spins that encounter tissue boundaries during their random walks. An alternative approach is to directly solve the Bloch-Torrey partial differential equation using the FDM [35]. The FDM discretizes the tissue sample to a spatial grid and updates the magnetization at each grid point over a series of time steps. To increase the computational efficiency and eliminate edge effects encountered with traditional FD methods we previously developed a matrix-based FDM with a revised periodic boundary condition [36]. For tissue structures sampled with N 3 voxels the discretized solution of the Bloch-Torrey equation for transverse magnetization (M) using the matrix-based FDM is described by:
| (4) |
A is an N 3×N 3 diffusion transition matrix containing the tissue structural information given in terms of the jump probabilities (probability that a spin starts at one grid point and diffuses to another grid point after a time interval Δt), I is an identity matrix with the same size of A, and ∶ represents element-by-element vector multiplication. The Φ(t) term is a 1×N 3 vector containing the phase accumulation and relaxation for each voxel at each time step and is given by:
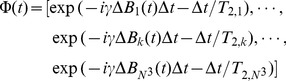 |
(5) |
where γ is the proton gyromagnetic ratio (267.5×106 rad s−1 T−1), Δt is the simulation time step, the subscript k indicates a spatial index, 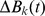 is the field perturbation at point k
, and
T2,k is the transverse relaxation time at location k within the simulation grid. When a GE sequence is used T2,k represent the intrinsic tissue T2*, and in the case of SE it represents the intrinsic T2. In general, the jump probability from simulation grid a to b is described by:
is the field perturbation at point k
, and
T2,k is the transverse relaxation time at location k within the simulation grid. When a GE sequence is used T2,k represent the intrinsic tissue T2*, and in the case of SE it represents the intrinsic T2. In general, the jump probability from simulation grid a to b is described by:
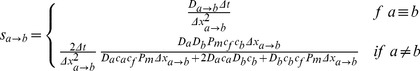 |
(6) |
where 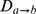 is the diffusion coefficient if a and b are within the same compartment,
is the diffusion coefficient if a and b are within the same compartment, 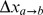 is the distance between simulation grid a and b, Pm is the permeability of the membrane when a and b are in different compartments, cf is the free concentration of water. The explicit form of the 1D transition matrix can be found in [36]. The MR signal normalized to the initial magnetization M0 is estimated by summing the magnetizations over all grid points at a particular t and is given by:
is the distance between simulation grid a and b, Pm is the permeability of the membrane when a and b are in different compartments, cf is the free concentration of water. The explicit form of the 1D transition matrix can be found in [36]. The MR signal normalized to the initial magnetization M0 is estimated by summing the magnetizations over all grid points at a particular t and is given by:
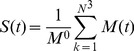 |
(7) |
The associated spin echo and gradient echo change in transverse relaxation rates are calculated at a particular echo time t = TE using:
| (8) |
For spin echo imaging, the phase was inverted at t = TE/2. This model is designed to handle cases where the three tissue compartments within a voxel can have different intrinsic transverse relaxation times. By updating T2,k in equation 5, for each grid point at each simulation time step, the total transverse relaxation, which includes the microscopic and mesoscopic relaxation effects, can also be calculated. The decay of signal from large static perturbers is known not to be exponential (e.g. diffusion in a static linear field gradient) but a simple exponential fit is a good approximation for realistic cases, and other functions can be easily fit. All simulations were performed in the Matlab environment (Mathworks, Natick, MA) running on Intel Core 2 Duo at 2.66 GHz and 4 GB of RAM processors. For clarity, the computational steps involved to obtain the final results are illustrated in Figure 1.
Figure 1. Computational steps involved in the FPFDM.

This figure illustrates the steps involved in computing the susceptibility induced transverse relaxation rates for a 3D tissue structure using the FPFDM: (a) The tissue structure is V(x,y,z). (b) The 3D Fourier transform of (a). (c) The magnetic field from the cubic finite perturber. (d) The 3D Fourier transform of 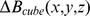 . (e) The point-wise multiplication of (b) and (d) in the Fourier domain. (f) The magnetic field shift due to the vascular structure computed as the 3D inverse Fourier transform of (e) or the convolution of (a) and (c). (g), (h) and (i) are the phase accumulation, the magnetization and the diffusion transition matrix, respectively. These are used to compute the magnetization in (j). (k) The computed MR signal. (l) The transverse relaxation rates associated with an arbitrarily shaped tissue structure.
. (e) The point-wise multiplication of (b) and (d) in the Fourier domain. (f) The magnetic field shift due to the vascular structure computed as the 3D inverse Fourier transform of (e) or the convolution of (a) and (c). (g), (h) and (i) are the phase accumulation, the magnetization and the diffusion transition matrix, respectively. These are used to compute the magnetization in (j). (k) The computed MR signal. (l) The transverse relaxation rates associated with an arbitrarily shaped tissue structure.
4. Contrast Agent Kinetics
To demonstrate the potential of the FPFDM to simulate DSC-MRI signals arising from the dynamic passage of contrast agent through the vascular and extravascular spaces, such as would occur in brain tumors with a breakdown of the blood brain barrier, we used tissue structures composed of ellipsoids packed around fractal based vascular network. Concentration time curves were sampled using 150 time points for a total of 9 minutes. The arterial input function (AIF) was generated as previously described [44]. The plasma and extravascular extracellular concentration time curves were computed using the pharmacokinetic two compartmental model described by Brix et al [45]. The relevant input physiological, pulse sequence and physical parameters (e.g. blood flow, blood volume, contrast agent transfer coefficient, T1, T2, etc) were selected from values measured in previous MRI, PET and CT brain tumor studies as previously described [16]. To investigate the influence of extravascular features on DSC-MRI, the signal is computed for two cellular structures with a similar cell volume fraction (∼60%) but different ellipsoid radii (5 and 15 µm). The ellipsoids were packed around a fixed vascular tree with a 4% volume fraction.
Results
1. Validation of FPFDM
For validation, FPFDM and Monte Carlo based MRI signals were computed and compared for models consisting of randomly orientated cylinders and packed spheres. The dependence of gradient-echo (ΔR2 *) and spin-echo (ΔR2) relaxivity on perturber (vessel) size has previously been characterized using Monte Carlo techniques [27]. To replicate these findings we created 10 different structures composed of approximately 40 randomly distributed cylinders for each vessel radius between 1 µm and 100 µm, each with total cylinder volume fraction equal to 2% of the simulation cube. Using the previously reported simulation parameters [27], [33] (susceptibility difference Δχ = 10−7 , cylinder volume fraction (Vp) = 2%, B0 = 1.5T, water diffusion coefficient D = 10−5 cm2/s, simulation time step Δt = 0.2 ms, GE TE = 60 ms and SE TE = 100 ms), we computed the vessel size dependence of ΔR2 * and ΔR2 averaged over all cylinder arrangements. The computed ΔR2 * and ΔR2 values show negligible changes as the number of averaged structures increases beyond 10. As shown in Fig. 2a, there is excellent agreement between the FPFDM results and those obtained in the Monte Carlo-based comparison studies, which used analytical expressions [27] and FPM [33] for field perturbation calculations.
Figure 2. Validation of the FPFDM.

(a) FPFDM replicates the characteristic vessel size dependence of ΔR2 *and ΔR2 as has been previously shown with MC methods. (b) A comparison of computed ΔR2 * values as a function of sphere volume fraction and packing arrangement using MC (filled symbols) and FPFDM (open symbols) techniques, with excellent agreement between the two methods. (c) The computed ΔR2 * percentage difference between MC and FPFDM decreases as the number of FPFDM structures used for averaging increases.
To compare the computational efficiency of the FPFDM with that of the MC method, we computed ΔR2 * values using both techniques. For each technique ΔR2 * values were computed for 18 radii using a TE = 60 ms and Δt = 0.2 ms. The computation time for the FPFDM was approximately 140 seconds per structure. Using 1000 randomly distributed spins, the computation time for the MC method was approximately 220 seconds per structure. Table 1 summarizes the simulation parameters used in the MC and FPFDM along with the respective computational times to generate ΔR2 * values for 18 cylinder radii.
Table 1. Parameters used in MC and FPFDM simulations along with total computing times to calculate ΔR2 * values for 18 cylinder radii.
| Parameters | Meaning | Value |
| TE | Echo time | 60 ms |
| Δt | Simulation time step | 0.2 ms |
| Δχ | Susceptibility difference | 10−7 |
| B0 | Static magnetic field | 1.5 T |
| Vp | Cylinder volume fraction | 2% |
| D | Water diffusion coefficient | 10−5 cm2/s |
| Ns | Number of spins used in MC method | 1000 |
| TimeMC | Computing time for MC method | 220 s |
| TimeFPFDM | Computing time for FPFDM | 140 s |
To further validate the accuracy of the FPFDM we also computed ΔR2 * for simulated 3D cellular models consisting of packed spheres. Two packing conditions were considered: randomly distributed spheres and sphere packing on FCC gird. For each model, the sphere size was fixed at 9 µm radius corresponding to an approximate pertuber size where the SE relaxivity peaks and the GE relaxivity reaches plateau [27]. The ΔR2 * dependence on cell (sphere) volume fraction for the FPFDM was compared to that for the MC method [27] using similar simulation parameters. The MC method was carried out on a different computer system using the approach described previously [27]. Fig. 2b compares the volume fraction dependence of ΔR2 * for each of the two sphere packing techniques computed by both the MC and FPFDM, using Δχ = 5×10−8, B0 = 1.5T, D = 10−5 cm2/s, Δt = 0.2 ms, GE TE = 40 ms, and simulation universe size = (0.5 mm)3. The FPFDM results were obtained by averaging the MR signal for 5 different sphere distributions for each packing and cell volume fraction using a simulation grid size of 1283. In contrast, the MC method involves tracking 15,000 random walks for each cell volume fraction and the redistribution of the spheres prior to each random walk. The FPFDM results are in excellent agreement to those produced from the MC methodology.
To investigate the convergence of the FPFDM for randomly distributed structures such as those used above, ΔR2 * values obtained from [27] for vessel sizes of 10 µm and 15 µm were compared to the FPFDM results as a function of the number of structures used for averaging. Fig. 2c shows the percentage difference between the MC and FPFDM derived ΔR2 * values. For both vessel sizes the computed FPFDM ΔR2 * values converge to the corresponding reported values [27], [33] to within 7% with only five structure averages. This percentage difference decreases to 0.8% as the number of averaged structures increases to 30.
2. Modeling the Effects of Contrast Agent Extravastion on DSC-MRI
To demonstrate the potential of the FPFDM for investigating the complex susceptibility effects that occur when contrast agent extravasates and compartmentalizes around cells, we computed the volume fraction dependence of ΔR2 *and ΔR2 for 3D cellular models consisting of packed spheres or ellipsoids (9 µm radius). To model contrast agent leakage effects relevant contrast agent levels corresponding to a Δχ value that is half the peak value of single dose Gd-DTPA injection was assumed. Fig. 3a illustrates the packed ellipsoid model with a representative 2D slice through the computed magnetic field perturbations. Fig. 3b and Fig. 3c demonstrates the influence of packing arrangements on the computed ΔR2 * and ΔR2 values for Δχ = 5×10−8, B0 = 1.5T, D = 10−5 cm2/s, simulation time step Δt = 0.2 ms, GE TE = 40 ms and SE TE = 80 ms.
Figure 3. Dependence of ΔR2 * and ΔR2 on cellular shape and packing arrangement.

(a) Example of a cellular model using ellipsoid packing (left) and a 2D slice through the associated magnetic field perturbation for B0 = 1.5T and Δχ = 5×10−8 (right). (b,c) The computed ΔR2 * and ΔR2 dependence on cell volume fraction and packing arrangement. For all packing arrangements, the relaxivity increases and then decreases with cell volume fraction. Ellipsoid packing yields greater relaxivity than spheres. ΔR2 exhibits qualitatively similar behavior to ΔR2 * yet with a reduced magnitude.
The highly ordered FCC packing of spheres resulted in the smallest relaxivity, reflecting the more homogeneous magnetic field perturbations and proton phase distributions. Randomly distributed spheres yielded slightly greater relaxivities with a non-linear relationship with packing fraction. Finally, the packed ellipsoids, which better approximate cell shape in vivo, enable higher random non-overlapping packing fractions (>65%), are less ordered and also yielded a non-linear relationship between relaxivity and cell volume fraction. For all cell volume fractions, the ΔR2 * and ΔR2 values associated with the ellipsoid-based structures were greater in magnitude than those found with spheres.
3. Modeling the Effects of Vascular Tree Heterogeneity on DSC-MRI
To illustrate the potential of the FPFDM for modeling the complex geometries of the microvasculature, we used fractal-based branching networks as input to the FPFDM. Fig. 4 illustrates the effect of branching angle heterogeneity (Δθ) on the concentration dependence of ΔR2 and ΔR2 * for typical DSC-MRI contrast agent concentrations. For these simulations we generated three different vascular networks within a 1 mm3 volume containing 1283 voxels. Fig. 4a–4c shows sample vascular trees with homogenous rotation (φ) angle and bifurcation index (α), which measures the relative diameter of daughter branches at each branching node, with increasing branching heterogeneity (θ). The model for normal vasculature is shown in Fig. 4a, with branching angles ranging from 25°–40°. To represent the tortuous and chaotically organized morphology of tumor vessels, the range of branching angle heterogeneity were increased to 25°–80° (Fig. 4b) and 25°–140° (Fig. 4c). Fig. 4d shows three orthogonal 2D slices through the body center of the magnetic field perturbations computed using the FPM for the vascular structure in Fig. 4c. Fig. 4e and 4f plot the concentration dependence of ΔR2 * and ΔR2 for the three θs considered. For these simulations, Δχ = χm. CA, where χm is the CA molar susceptibility (0.027×10−6 mM−1) [46], B0 = 4.7T, D = 10−5 cm2/s, Δt = 0.2 ms, GE TE = 40 ms and SE TE = 80 ms. All simulated vascular structures incorporate vessel radii ranging from 12 µm to 80 µm with a 2% target vascular volume fraction (v p). The computed relaxation rates were averaged over five different orientations for each simulated vascular network. Using the slope of the ΔR2 * dose response curves, kp values ranging from 100–295 (mM–sec) −1 were obtained. For this low vascular volume fraction, the kp values for the more tumor-like vascular trees (higher Δθ) were higher than those in normal trees, up to three fold for this simplified simulation. Similar dependency on branching angle with a reduced susceptibility effect was observed for ΔR2 dose response curves.
Figure 4. The influence of vascular morphology on ΔR2 * and ΔR2.

(a–c) Sample microvascular networks simulated using a fractal tree model with increasing branching angle heterogeneity. (d) Three orthogonal slices through the magnetic field perturbation at the body center for the vascular network in (c). (e–f) Effect of branching angle heterogeneity on the concentration dependence of ΔR2 * and ΔR2 computed with FPFDM (B0 = 4.7T, Δχ = 1×10−7, 2% target vascular volume fraction). Both ΔR2 and ΔR2 * increase with branching angle heterogeneity.
4. Modeling First-pass DSC-MRI Data in Brain Tumors
To demonstrate the feasibility of using the FPFDM as a tool to simulate DSC-MRI signals in the presence of contrast agent extravasation, we used two sample tissue structures composed of 60% cells and 4% blood vessels. The two tissue structures were constructed by packing ellipsoids with radii of 5 µm and 15 µm around a fixed fractal-based vascular network. Fig. 5a shows a sample 3D volume rendering of such a tissue structure, which contains ellipsoids of 15 µm average radius and vascular network with vessel size ranging from 5 µm to 45 µm. The simulated vascular (Cp) and extravascular (Ce) contrast agent concentration time curves are shown in (Fig. 5b). Fig. 5c shows a representative 2D slice through the computed magnetic field perturbations at a particular time. The standard deviation of the field perturbation (std ΔB) for all simulated time points is shown in (Fig. 5d). The simulated Cp and Ce curves along with model tissue structure, in Fig. 5, were used as an input to compute the dynamic DSC-MRI signal. Fig. 6 shows the GE post-contrast to pre-contrast DSC-MRI signal ratio time curves (S/S0), both in the presence (K Trans = 0.2 min−1) and absence (K Trans = 0 min−1) of contrast agent extravasation. Fig. 6a-c show the time series for the tissue structure composed of ellipsoids with a 5 µm mean radii, at pre-contrast longitudinal relaxation times values of T 10 = 500 ms, T 10 = 1000 ms and T 10 = 1500 ms, respectively. The corresponding time series for the tissue structures modeled with 15 µm cellular radii are shown in (Fig. 6d–6f). The following input parameters were used to compute the DSC-MRI signal: B0 = 3T, D = 1.3×10−5 cm2/s, Δt = 0.2 ms, TE = 50 ms, TR = 1500 ms, flip angle α = 90°, pre-contrast transverse relaxation time T20 * = 50 ms. The CA T1 and T2 relaxivity values, r1 and r2, were set to 3.9 and 5.3 mM−1s−1, respectively [47]. The compartmental membrane water permeability values were set at P m = 0, to model restricted water diffusion. For a fixed cell volume fraction, the simulated time series demonstrate a marked cell size dependency. In general, for both tissue structures, as T10 increases from 500 ms to 1500 ms the influence of T1 leakage effects becomes more substantial, as indicated by the increased signal recovery. For the small cell size structure, the T1 leakage effects eventually result in a signal overshot from baseline (e.g. Fig. 6c). However, the structure constructed with larger cell sizes is dominated by T2 * leakage effects (as apparent from the low signal recovery well after the CA’s first pass) even at T10 = 1500 ms (Fig. 6f). The simulation time to compute the signal for 150 time points, for 3 T10 values, 2 contrast agent leakage conditions and 2 tissue structures was approximately 240 mins.
Figure 5. Example simulation with realistic tissue structure and contrast agent extravasation.
(a) Sample tissue structure composed of ellipsoids packed around fractal tree based vascular network. (b) Simulated C p and C e curves derived using 2-compartment model. (c) Example 2D map through the magnetic field perturbation computed at time t = 300 sec. (d). The time evolution of the standard deviation of the field perturbation (std ΔB) computed using B0 = 3T, C p and C e for the given sample structure.
Figure 6. Dependence of DSC-MRI signals on cellular features in the presence of CA leakage.

The GE post-contrast to pre-contrast DSC-MRI signal ratio (S/S0), both in the presence (K Trans = 0.2 min−1) and absence (K Trans = 0 min−1) of CA leakage at pre-contrast T 1 values of T 10 = 500 ms, T 10 = 1000 ms and T 10 = 1500 ms, for tissue structures constructed using ellipsoids with mean radii of 5 µm (a–c) and 15 µm (d–f), respectively. The (S/S0) values were computed using input parameters of B0 = 3T, D = 1.3×10−5 cm2/s, Δt = 0.2 ms, TE = 50 ms TR = 1500 ms, α = 90°, T20 * = 50 ms, r1 = 3.9 mM−1s−1, r2 = 5.3 mM−1s−1 and Pm = 0.
5. Application to Image-based Tissue Structures
To further illustrate the versatility of the FPFDM, micro-CT images of perfused mouse kidney vasculature (1,242 slices with 1428×1012 matrix size and 5 µm isotropic voxels) were used as source data for multiple 1 mm3 vascular models with 2003 finite cubic perturbers, each 5 µm in size. Fig. 7 shows the entire extracted kidney vascular structure, with sample MR voxel-sized sub-structures and their respective vascular volume fractions. Fig. 8a and 8b shows the FPFDM derived SE and GE kp values obtained from the slope of the ΔR2 and ΔR2 * dose response curves, respectively. These results are normalized to the vascular fractional volumes and were computed using B0 = 4.7T, D = 10−5 cm2/s, Δt = 0.2 ms, GE TE = 40 ms, SE TE = 80 ms, and a clinically relevant range of Δχ values ranging from 0 to 9.4×10−8, corresponding to a Gd-DTPA concentration ranging from 0 to 3.5 mM. In general, the SE and GE kp values were highest for low vascular volume fractions and tended to decrease as the vascular volume fraction increased, with SE and GE kp values ranging from 3.6–27.8 and 53.8–174.3 (mM–sec) −1, respectively.
Figure 7. Kidney vascular structure extracted from micro-CT.

Kidney vasculature extracted from micro-CT along with representative MR voxel-sized (1 mm3) microvascular models taken from different sections of the kidney vasculature with their respective vascular volume fractions. The existence of the bubble-like structures demonstrates the filling of glomeruli with Microfil but a higher resolution would be required to differentiate the individual capillaries.
Figure 8. Computed kp values for vascular structure extracted from micro-CT.
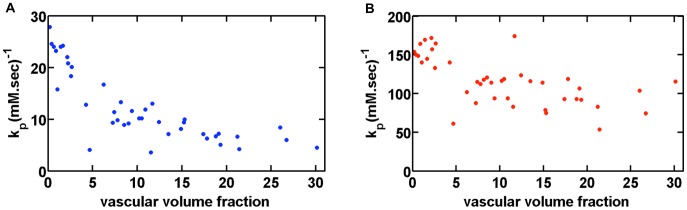
(a) SE and (b) GE kp values as a function of vascular volume fraction computed using the FPFDM for the kidney microvascular models (with vascular volume fractions >0.1%) shown in Fig. 7. SE kp values ranged from 3.6–27.8 (mM–sec) −1, and GE kp values ranged from 53.8–174.3 (mM–sec) −1. Above 5% volume fraction, the GE kp values were relatively constant with a mean value of 103.3(mM–sec) −1.
Discussion
The FPFDM is a novel efficient computational tool combining features of the FP and FD techniques for calculating susceptibility-induced relaxivity changes for realistic simulated or imaging-based 3D vascular and cellular geometries that might be observed in vivo. The FPM can compute the induced magnetic fields around arbitrary microvasculature structures without necessitating any assumptions about the underlying vessel geometry [33]. Although the Fast Fourier transform (FFT) improves the computational efficiency of the FPM for computing magnetic field perturbations, the application of MC techniques for tracking proton diffusion through the tissue in order to derive the resulting relaxivity change reduces its computational efficiency. The replacement of the MC component of FPM with matrix-based FDM can increase the computational efficiency by computing the evolution of the discretized magnetization simultaneously [36]. The transition matrix A is either invariant or requires only partial updating for most tissues under consideration, further increasing the computing efficiency of matrix-based FDM that also benefit from optimized MATLAB packages for computations involving large matrices [36].
A Gaussian diffusion kernel convolution can also be used to model CA and water diffusion [39], [40], [48]. This approach is computationally more efficient than MC approaches, but limited to unrestricted water diffusion. Although non-Gaussian diffusion, a consequence of tissue structure that creates diffusion barriers and compartments, could be modeled by adding a kurtosis term to the kernel, it is not clear how this will affect the slower diffusion process observed in the restricted CA diffusion model [40]. Modeling restricted water diffusion using the MC method [33] or the Gaussian diffusion kernel approach [39], [40], [48] requires either incorporation of elastic collisions at membrane boundaries or neglecting proton diffusion steps that involve membrane crossing. Unlike the case of unrestricted water diffusion, using these later methods to model restricted water diffusion and/or water diffusion in complex tissue with different compartmental diffusion coefficients will require additional computations, thereby increasing the overall processing time. Given compartmental diffusion coefficients and membrane permeability values, the FPFDM can be used to model restricted water and CA diffusion and water exchange across compartments. For the FPFDM, including these additional structural features requires the computation of multiple versions of the diffusion transition matrix, A. Since A can be determined at the start of the simulation, a library of diffusion transition matrices, for a range of tissue structures, can be established to increase the computing efficiency. For example, computing a dynamic signal for the same structure only requires loading the transition matrix corresponding to the structure once from the library of diffusion transition matrices.
We validated the FPFDM in two ways. First, we replicated the vessel size dependence of ΔR2 * and ΔR2 (Fig. 2a) using identical simulation parameters to previously described MC and FP techniques [27], [33]. Next, we found excellent agreement for relaxivity from packed spheres across a range of packing densities and packing strategies using traditional MC technique versus FPFDM (Fig. 2b). The agreement between MC and FPFDM converges as the number of structures included in the average for the FPFDM increases (Fig. 2c). Unlike MC simulation, which tracks a large number of particles in the simulation or, equivalently, runs the same simulation many times to obtain an accurate average result, the FPFDM converges to the average result with only a few simulation runs. This behavior can be explained in the following way. In the MC simulation, a population of particles distributes in the whole system and the particles that encounter membranes within the complex tissue are only a small portion of all the particles such that the echo signal does not contain sufficient enough information about the tissue features that restrict diffusion. Hence, to solve this problem, more particles are considered in the simulation or, equivalently, the same simulation is run many times to obtain an accurate average result. In contrast, the FDFDM determines the diffusion transition matrix at the start of the simulation, which already contains the tissue structural information and results in a faster convergence of the average signal.
For a simplistic structure containing randomly oriented cylinders with a total of 18 different radii, the FPFDM, as compared to MC, reduced the computation time to calculate ΔR2 * values from 220 s to 140 s. For complex tissue structures, and under conditions of restricted water diffusion, the increase in computational efficiency afforded by the FPFDM will improve even further. In such cases, the MC method requires a larger number of spins and additional computation steps in order to converge and capture sufficient information about the tissue structure [49]. In contrast, for these more complex structures, the FPFDM does not require additional computing time and is not limited by restricted water diffusion [36].
The FPFDM has the potential for modeling nonstandard geometries that may better simulate cells and microvasculature in vivo. We computed relaxivities for simulated 3D cellular models consisting of packed spheres and ellipsoids (Fig. 3), and found greater relaxivity for packed ellipsoids over all volume fractions compared to the sphere packing. This suggests that the additional degree of freedom in spatial orientation for ellipsoids increases field perturbation heterogeneity. The greater orientational heterogeneity and packing density afforded by ellipsoids compared to spheres would seem to make ellipsoids better suited for simulating susceptibility-based contrast mechanisms of cellular structures.
Although simulations in this study are based on a simple 2-compartment model, at the expense of computational time, the same approach to model water diffusion and exchange can be used to model CA diffusion and transport across compartments, yielding a more realistic heterogeneous CA distribution within a voxel. This can be achieved by updating CA concentration for each voxel at each simulation time step using a CA diffusion transition matrix (ACA) calculated using appropriate CA diffusion coefficients and permeability across membranes from literature [50], [51]. For the purposes of this study, we assumed that the contrast agent equilibrates within each compartment over the time it takes to acquire each DSC-MRI image (1–2 seconds). Such an assumption is traditionally employed and consistent with current DSC-MRI analysis techniques.
We also computed the contrast agent concentration dependence of transverse relaxation rates for vascular trees. Traditionally, randomly oriented cylinders were used to investigate the influence of vascular properties (e.g. vessel size, vessel volume fraction) on relaxation rates. Fractal-based vascular trees better approximate the microvascular network in vivo, but this complex geometry with variable vessel rotation, size distribution, branching angles, and diameters of daughter vessels is very difficult to model and require high resolution to achieve structural details. Our results demonstrate the feasibility of using FPFDM for complex geometries, and suggest that although the generally accepted linear relationship between relaxation rate and CA concentration is reasonable, the proportionality constant kp depends upon the microvascular geometry, a finding that is consistent with previous studies [27], [33], [52]. The higher relaxation rate for vascular structures with greater range of branching angles is most likely due to the greater heterogeneity of vessel branch orientation with respect to B0 and their larger space occupancy which impacts the frequency offset of a larger volume compared to narrow branching angles that pack vessels in a small region. These preliminary computational results show marked kp heterogeneity across vascular networks, suggesting that further work is needed to better characterize the influence of vascular heterogeneity on DSC-MRI derived perfusion parameters in brain tumors. More systematic studies are underway for wide range of morphological, physiological and pulse sequence parameters to investigate these findings at the higher vascular volume fractions more typically encountered in DSC-MRI experiments, such as in grey matter or brain tumors.
The systematic evaluation of fractal-based vascular trees using the FPFDM could shed new insights into the relationship between DSC-MRI relaxation rate and vascular geometry. Furthermore, the use of fractal trees enables the application of well-established flow models [53]–[55] such that contrast agent kinetics and the associated DSC-MRI time series can be considered for each vascular structure. This would enable a more rigorous investigation of DSC-MRI-based voxel-wise measures of vessel size, transit time and flow distributions and oxygen extraction. Realistic 3D vascular and flow models could then be expanded to incorporate the extravasation of contrast agent and its subsequent diffusion around cells in the extravascular space. Such expansions would create a powerful framework with which to investigate DSC-MRI and susceptibility-based imaging methodologies in brain, tumors and other organs of the body.
The FPFDM also provides a computationally reasonable approach for simulating DSC-MRI derived transverse relaxation rates both in the presence and absence of CA extravasation, and restricted water diffusion induced by membrane permeability (Fig. 5 and Fig. 6). The results shown in Fig. 6 demonstrate that contrast agent leakage can lead competing T1 and T2 * effects as the CA traverses the extravascular extracellular space. For a given T10, the structure with smaller sized cells exhibited higher signal intensity recoveries as compared to that with larger sized cells. The compartmentalization of CA around the larger cells creates stronger magnetic field perturbations and greater relaxation rate changes (T2 * effects). In general, as T10 increases, T1 leakage effects will be more pronounced and may dominate any T2 * leakage effects, as is the case for the smaller-sized cells. In such cases, the characteristic signal overshoot may be observed (Fig. 6c). For the tissue structure with larger perturber sizes, the signal intensity exhibits less recovery due to the presence of substantial T2 * leakage effects (Fig. 6d–6f). As shown in Fig. 3b, cell density may also influence the shape of DSC-MRI signals, with the magnitude of T2 * leakage effects decreasing (and T1 leakage effects increasing) as the cell density increases. Consequently, DSC-MRI data from tumors with tightly packed, smaller-sized cells would likely present with pronounced T1 leakage effects (e.g. signal overshoot). Given the clinical importance of DSC-MRI signal recovery characteristics to help differentiate among tumor types [37], [56], a systematic in silico study of DSC-MRI signal recovery and its dependence on physiological, pulse sequence and physical parameters is currently under investigation.
Prior studies have shown the potential and value of incorporating image-based vascular structures into susceptibility simulations [39]. Similar to these previous studies we sought to demonstrate the versatility of the FPFDM by determining the dose-response of relaxation rates for vascular structures derived from ex vivo micro-CT scans of perfused kidney vasculature. The dose-response curves from MRI voxel-sized regions of the kidney vasculature were used to determine the distribution of vascular susceptibility calibration factors, kp, within the kidney. For vascular volume fractions up to 30%, kp values were very heterogeneous (Fig. 8), with decreased heterogeneity for vascular volume fractions greater than 5%. The kp decreased over vascular volume fractions between 0 and 5% with a slower decrease above 5%, consistent with a previous study in rodent brain that found grey matter kp to be nearly twice that of tumor [34]. It should be noted that the kidney microvascular structure presented in this study is limited by the spatial resolution of the micro-CT data. With a 5 mm, resolution individual capillaries could not be resolved and capillary dense regions, such as in the glomeruli, present as a single large perturber. The differentiation and inclusion of these capillaries will likely influence the overall kp heterogeneity across voxels for both SE and GE computations. For the purposes of this study, this example illustrates the ability of the FPFDM to explore susceptibility contrast in tissue structures acquired using ex vivo imaging modalities. As the FPFDM only requires that structures consist of a digital format it could accept structural input from any imaging modality (e.g. optical, CT, electron microscopy, MRI).
One of the limitations of the FPM is the use of FFT to calculate the spatial convolution of the vascular structure with the finite perturber magnetic field perturbation. As demonstrated in [33] the resulting field perturbation is equivalent to the field perturbation from a periodic array of the tissue structure under consideration. Although realistic tissue structures extend beyond the boundary of the simulation space, which introduces a “boundary problem”, we used zero-padding of the tissue structure to avoid additional field perturbation at the boundaries from the neighboring array. The padding size to eliminate boundary field effects depends on the perturber size and the tissue structure. Here we used a zero-pad size of one-tenth of the simulation box, since the field perturbation changes we observed by using higher zero-pad sizes were negligible.
The FPM was designed to compute the magnetic field changes from a single finite perturber convolved with a digitized tissue structure array, and hence this approach cannot be used for arbitrary magnetic susceptibility distributions. While methods capable of computing arbitrary susceptibility distributions are more comprehensive and should be explored [57], [58], it is typically assumed that contrast agent instantaneously distributes within each tissue compartment (e.g. intravascular and extravascular extracellular space) at each imaging time point. Accordingly, the FPFDM is a practical approach to compute field perturbations arising from tissue structure with only a few susceptibility compartments, such as the intravascular, intracellular, and extravascular extracellular spaces.
The sampling of tissue structures at higher resolution increases the computational accuracy of the FPM but it comes at the expense of computational time. Such increases in resolution would also add to computational time needed to compute the MR signal using the FDM. This is particularly true if a need arises to reduce the simulation time step (Δt) due to increased resolution or decreased perturber size (Δx), in order to satisfy the constraint that the jump probability (see Eq. 6) should be ≤1/6. This is because when the number of spins leaving a given node exceeds the number that was present, the FDM becomes unstable [35]. With the parallel high-performance computing techniques we previously developed [36], we are exploring ways to increase the computational efficiency of the FPFDM at higher resolutions so that we can more accurately characterize fine tissue microstructure across a broader range of structural dimensions (e.g. a few microns up to a hundreds of microns).
Conclusion
The FPFDM is an alternative computational tool for efficiently modeling susceptibility induced MR signal relaxation from complex perturber geometries. In general, the proposed FPFDM could be used to investigate the influence of realistic tissue microstructure on any susceptibility based contrast mechanism such as vessel size imaging, BOLD contrast, single cell imaging, and quantitative susceptibility mapping. Currently, the proposed method is being utilized to assess the influence of geometrical, morphological and physiological parameters of microvessels and cells on susceptibility induced MR relaxation rate changes. Such studies should shed new insights into DSC-MRI contrast mechanisms and enable the systematic evaluation of how acquisition and analysis methods influence the measurement of reliable perfusion parameters in brain and tumor tissue.
Acknowledgments
We thank Dr. Daniel Perrien for his help with the kidney CT imaging.
Funding Statement
This research was supported by the National Institutes of Health through NIH/NCI K99/R00 CA127599, NIH/NCI R01 CA158079, P30 CA068485, NCI U24 CA126588. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Rosen BR, Belliveau JW, Vevea JM, Brady TJ (1990) Perfusion imaging with NMR contrast agents. Magn Reson Med 14: 249–265. [DOI] [PubMed] [Google Scholar]
- 2. Meier P, Zierler KL (1954) On the theory of the indicator-dilution method for measurement of blood flow and volume. J Appl Physiol 6: 731–744. [DOI] [PubMed] [Google Scholar]
- 3. Aronen HJ, Gazit IE, Louis DN, Buchbinder BR, Pardo FS, et al. (1994) Cerebral blood volume maps of gliomas: comparison with tumor grade and histologic findings. Radiology 191: 41–51. [DOI] [PubMed] [Google Scholar]
- 4. Aronen HJ, Pardo FS, Kennedy DN, Belliveau JW, Packard SD, et al. (2000) High microvascular blood volume is associated with high glucose uptake and tumor angiogenesis in human gliomas. Clin Cancer Res 6: 2189–2200. [PubMed] [Google Scholar]
- 5. Batchelor TT, Sorensen AG, di Tomaso E, Zhang WT, Duda DG, et al. (2007) AZD2171, a pan-VEGF receptor tyrosine kinase inhibitor, normalizes tumor vasculature and alleviates edema in glioblastoma patients. Cancer Cell 11: 83–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Bruening R, Kwong KK, Vevea MJ, Hochberg FH, Cher L, et al. (1996) Echo-planar MR determination of relative cerebral blood volume in human brain tumors: T1 versus T2 weighting. AJNR Am J Neuroradiol 17: 831–840. [PMC free article] [PubMed] [Google Scholar]
- 7. Donahue KM, Krouwer HG, Rand SD, Pathak AP, Marszalkowski CS, et al. (2000) Utility of simultaneously acquired gradient-echo and spin-echo cerebral blood volume and morphology maps in brain tumor patients. Magn Reson Med 43: 845–853. [DOI] [PubMed] [Google Scholar]
- 8. Maeda M, Itoh S, Kimura H, Iwasaki T, Hayashi N, et al. (1993) Tumor vascularity in the brain: evaluation with dynamic susceptibility-contrast MR imaging. Radiology 189: 233–238. [DOI] [PubMed] [Google Scholar]
- 9. Sugahara T, Korogi Y, Kochi M, Ushio Y, Takahashi M (2001) Perfusion-sensitive MR imaging of gliomas: comparison between gradient-echo and spin-echo echo-planar imaging techniques. AJNR Am J Neuroradiol 22: 1306–1315. [PMC free article] [PubMed] [Google Scholar]
- 10. Lemasson B, Serduc R, Maisin C, Bouchet A, Coquery N, et al. (2010) Monitoring blood-brain barrier status in a rat model of glioma receiving therapy: dual injection of low-molecular-weight and macromolecular MR contrast media. Radiology 257: 342–352. [DOI] [PubMed] [Google Scholar]
- 11. Siegal T, Rubinstein R, Tzuk-Shina T, Gomori JM (1997) Utility of relative cerebral blood volume mapping derived from perfusion magnetic resonance imaging in the routine follow up of brain tumors. J Neurosurg 86: 22–27. [DOI] [PubMed] [Google Scholar]
- 12. Sugahara T, Korogi Y, Tomiguchi S, Shigematsu Y, Ikushima I, et al. (2000) Posttherapeutic intraaxial brain tumor: the value of perfusion-sensitive contrast-enhanced MR imaging for differentiating tumor recurrence from nonneoplastic contrast-enhancing tissue. AJNR Am J Neuroradiol 21: 901–909. [PMC free article] [PubMed] [Google Scholar]
- 13. Sorensen AG, Emblem KE, Polaskova P, Jennings D, Kim H, et al. (2012) Increased survival of glioblastoma patients who respond to antiangiogenic therapy with elevated blood perfusion. Cancer Res 72: 402–407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Kamoun WS, Ley CD, Farrar CT, Duyverman AM, Lahdenranta J, et al. (2009) Edema control by cediranib, a vascular endothelial growth factor receptor-targeted kinase inhibitor, prolongs survival despite persistent brain tumor growth in mice. J Clin Oncol 27: 2542–2552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Rosen BR, Belliveau JW, Buchbinder BR, McKinstry RC, Porkka LM, et al. (1991) Contrast agents and cerebral hemodynamics. Magn Reson Med 19: 285–292. [DOI] [PubMed] [Google Scholar]
- 16. Quarles CC, Gochberg DF, Gore JC, Yankeelov TE (2009) A theoretical framework to model DSC-MRI data acquired in the presence of contrast agent extravasation. Phys Med Biol 54: 5749–5766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Sourbron S, Heilmann M, Biffar A, Walczak C, Vautier J, et al. (2009) Bolus-tracking MRI with a simultaneous T1- and T2*-measurement. Magn Reson Med 62: 672–681. [DOI] [PubMed] [Google Scholar]
- 18. Sourbron S, Heilmann M, Walczak C, Vautier J, Schad LR, et al. (2013) T2*-relaxivity contrast imaging: first results. Magn Reson Med 69: 1430–1437. [DOI] [PubMed] [Google Scholar]
- 19. Bjornerud A, Sorensen AG, Mouridsen K, Emblem KE (2011) T1- and T2*-dominant extravasation correction in DSC-MRI: part I–theoretical considerations and implications for assessment of tumor hemodynamic properties. J Cereb Blood Flow Metab 31: 2041–2053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Paulson ES, Schmainda KM (2008) Comparison of dynamic susceptibility-weighted contrast-enhanced MR methods: recommendations for measuring relative cerebral blood volume in brain tumors. Radiology 249: 601–613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Frohlich AF, Ostergaard L, Kiselev VG (2005) Theory of susceptibility-induced transverse relaxation in the capillary network in the diffusion narrowing regime. Magn Reson Med 53: 564–573. [DOI] [PubMed] [Google Scholar]
- 22. Kiselev VG (2001) On the theoretical basis of perfusion measurements by dynamic susceptibility contrast MRI. Magn Reson Med 46: 1113–1122. [DOI] [PubMed] [Google Scholar]
- 23. Kiselev VG, Posse S (1999) Analytical model of susceptibility-induced MR signal dephasing: effect of diffusion in a microvascular network. Magn Reson Med 41: 499–509. [DOI] [PubMed] [Google Scholar]
- 24. Kjolby BF, Ostergaard L, Kiselev VG (2006) Theoretical model of intravascular paramagnetic tracers effect on tissue relaxation. Magn Reson Med 56: 187–197. [DOI] [PubMed] [Google Scholar]
- 25. Weisskoff RM, Zuo CS, Boxerman JL, Rosen BR (1994) Microscopic susceptibility variation and transverse relaxation: theory and experiment. Magn Reson Med 31: 601–610. [DOI] [PubMed] [Google Scholar]
- 26. Boxerman JL, Bandettini PA, Kwong KK, Baker JR, Davis TL, et al. (1995a) The intravascular contribution to fMRI signal change: Monte Carlo modeling and diffusion-weighted studies in vivo. Magn Reson Med 34: 4–10. [DOI] [PubMed] [Google Scholar]
- 27. Boxerman JL, Hamberg LM, Rosen BR, Weisskoff RM (1995b) MR contrast due to intravascular magnetic susceptibility perturbations. Magn Reson Med 34: 555–566. [DOI] [PubMed] [Google Scholar]
- 28. Fisel CR, Ackerman JL, Buxton RB, Garrido L, Belliveau JW, et al. (1991) MR contrast due to microscopically heterogeneous magnetic susceptibility: numerical simulations and applications to cerebral physiology. Magn Reson Med 17: 336–347. [DOI] [PubMed] [Google Scholar]
- 29. Kennan RP, Zhong J, Gore JC (1994) Intravascular susceptibility contrast mechanisms in tissues. Magn Reson Med 31: 9–21. [DOI] [PubMed] [Google Scholar]
- 30. Ogawa S, Lee TM (1990) Magnetic resonance imaging of blood vessels at high fields: in vivo and in vitro measurements and image simulation. Magn Reson Med 16: 9–18. [DOI] [PubMed] [Google Scholar]
- 31. Yablonskiy DA, Haacke EM (1994) Theory of NMR signal behavior in magnetically inhomogeneous tissues: the static dephasing regime. Magn Reson Med 32: 749–763. [DOI] [PubMed] [Google Scholar]
- 32. Stables LA, Kennan RP, Gore JC (1998) Asymmetric spin-echo imaging of magnetically inhomogeneous systems: theory, experiment, and numerical studies. Magn Reson Med 40: 432–442. [DOI] [PubMed] [Google Scholar]
- 33. Pathak AP, Ward BD, Schmainda KM (2008) A novel technique for modeling susceptibility-based contrast mechanisms for arbitrary microvascular geometries: the finite perturber method. Neuroimage 40: 1130–1143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Pathak AP, Rand SD, Schmainda KM (2003) The effect of brain tumor angiogenesis on the in vivo relationship between the gradient-echo relaxation rate change (DeltaR2*) and contrast agent (MION) dose. J Magn Reson Imaging 18: 397–403. [DOI] [PubMed] [Google Scholar]
- 35. Hwang SN, Chin CL, Wehrli FW, Hackney DB (2003) An image-based finite difference model for simulating restricted diffusion. Magn Reson Med 50: 373–382. [DOI] [PubMed] [Google Scholar]
- 36. Xu J, Does MD, Gore JC (2007) Numerical study of water diffusion in biological tissues using an improved finite difference method. Phys Med Biol 52: N111–126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Mangla R, Kolar B, Zhu T, Zhong J, Almast J, et al. (2011) Percentage signal recovery derived from MR dynamic susceptibility contrast imaging is useful to differentiate common enhancing malignant lesions of the brain. AJNR Am J Neuroradiol 32: 1004–1010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Delaney GW, Cleary PW (2010) The packing properties of superellipsoids. Europhys Lett 89: 34002. [Google Scholar]
- 39. Christen T, Zaharchuk G, Pannetier N, Serduc R, Joudiou N, et al. (2012) Quantitative MR estimates of blood oxygenation based on T2*: a numerical study of the impact of model assumptions. Magn Reson Med 67: 1458–1468. [DOI] [PubMed] [Google Scholar]
- 40. Pannetier NA, Debacker CS, Mauconduit F, Christen T, Barbier EL (2013) A simulation tool for dynamic contrast enhanced MRI. PLoS One 8: e57636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Rybaczuk M, Kedzia A, Paradowski L (2002) Fractal characteristics of brain vessel microangioarchitecture during the fetal period. Med Sci Monit 8: MT145–152. [PubMed] [Google Scholar]
- 42. Schreiner W, Neumann F, Neumann M, End A, Muller MR (1996) Structural quantification and bifurcation symmetry in arterial tree models generated by constrained constructive optimization. J Theor Biol 180: 161–174. [DOI] [PubMed] [Google Scholar]
- 43. Murray CD (1926) The Physiological Principle of Minimum Work: I. The Vascular System and the Cost of Blood Volume. Proc Natl Acad Sci U S A 12: 207–214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Simpson NE, He Z, Evelhoch JL (1999) Deuterium NMR tissue perfusion measurements using the tracer uptake approach: I. Optimization of methods. Magn Reson Med 42: 42–52. [DOI] [PubMed] [Google Scholar]
- 45. Brix G, Bahner ML, Hoffmann U, Horvath A, Schreiber W (1999) Regional blood flow, capillary permeability, and compartmental volumes: measurement with dynamic CT–initial experience. Radiology 210: 269–276. [DOI] [PubMed] [Google Scholar]
- 46. Weisskoff RM, Kiihne S (1992) MRI susceptometry: image-based measurement of absolute susceptibility of MR contrast agents and human blood. Magn Reson Med 24: 375–383. [DOI] [PubMed] [Google Scholar]
- 47. Pintaske J, Martirosian P, Graf H, Erb G, Lodemann KP, et al. (2006) Relaxivity of Gadopentetate Dimeglumine (Magnevist), Gadobutrol (Gadovist), and Gadobenate Dimeglumine (MultiHance) in human blood plasma at 0.2, 1.5, and 3 Tesla. Invest Radiol 41: 213–221. [DOI] [PubMed] [Google Scholar]
- 48. Klassen LM, Menon RS (2007) NMR simulation analysis of statistical effects on quantifying cerebrovascular parameters. Biophys J 92: 1014–1021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Hall MG, Alexander DC (2009) Convergence and parameter choice for Monte-Carlo simulations of diffusion MRI. IEEE Trans Med Imaging 28: 1354–1364. [DOI] [PubMed] [Google Scholar]
- 50. Beaumont M, Lemasson B, Farion R, Segebarth C, Remy C, et al. (2009) Characterization of tumor angiogenesis in rat brain using iron-based vessel size index MRI in combination with gadolinium-based dynamic contrast-enhanced MRI. J Cereb Blood Flow Metab 29: 1714–1726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Marty B, Djemai B, Robic C, Port M, Robert P, et al. (2013) Hindered diffusion of MRI contrast agents in rat brain extracellular micro-environment assessed by acquisition of dynamic T1 and T2 maps. Contrast Media Mol Imaging 8: 12–19. [DOI] [PubMed] [Google Scholar]
- 52. Newman GC, Hospod FE, Patlak CS, Fain SE, Pulfer KA, et al. (2006) Experimental estimates of the constants relating signal change to contrast concentration for cerebral blood volume by T2* MRI. J Cereb Blood Flow Metab 26: 760–770. [DOI] [PubMed] [Google Scholar]
- 53. Gabrys E, Rybaczuk M, Kedzia A (2005) Fractal models of circulatory system. Symmetrical and asymmetrical approach comparison. Chaos Solitons & Fractals 24(3): 707–715. [Google Scholar]
- 54. Gabrys E, Rybaczuk M, Kedzia A (2006) Blood flow simulation through fractal models of circulatory system. Chaos Solitons & Fractals 27(1): 1–7. [Google Scholar]
- 55. Karshafian R, Burns PN, Henkelman MR (2003) Transit time kinetics in ordered and disordered vascular trees. Phys Med Biol 48: 3225–3237. [DOI] [PubMed] [Google Scholar]
- 56.Boxerman JL, Paulson ES, Prah MA, Schmainda KM (2013) The Effect of Pulse Sequence Parameters and Contrast Agent Dose on Percentage Signal Recovery in DSC-MRI: Implications for Clinical Applications. AJNR Am J Neuroradiol. [DOI] [PMC free article] [PubMed]
- 57. Marques JP, Bowtell R (2005) Application of a fourier-based method for rapid calculation of field inhomogeneity due to spatial variation of magnetic susceptibility. Concepts in Magnetic Resonance Part B-Magnetic Resonance Engineering 25B(1): 65–78. [Google Scholar]
- 58. Salomir R, Senneville BDD, Moonen CTW (2003) A fast calculation method for magnetic field inhomogeneity due to an arbitrary distribution of bulk susceptibility. Concepts in Magnetic Resonance Part B-Magnetic Resonance Engineering 19B(1): 26–34. [Google Scholar]