

Abstract
AIM: To explore the epithelial-mesenchymal transition (EMT) in tissue from patients with Lynch syndrome, and to interpret biological behaviour of Lynch syndrome.
METHODS: Sixty-eight formalin-fixed and paraffin embedded tissue blocks were analyzed in this study, including tissues from Lynch syndrome (n = 30), sporadic colorectal carcinoma (CRC) (n = 30), and tumor-adjacent tissues (n = 8). Tissue sections were stained for human mutS homolog 2 (hMSH2), human mutL homolog 1 (hMLH1), transforming growth factor-β type II receptor (TGFβRII), E-cadherin, β-catenin, matrix metalloproteinase-7 (MMP-7) and tissue inhibitor of metalloproteinase-2 (TIMP-2) by immunohistochemical staining. Furthermore, clinical data such as age, gender and tumor-node-metastasis stage were also collected retrospectively.
RESULTS: The positive expression rates of hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, MMP-7 and TIMP-2 were significantly related to the depth of invasion and lymph node metastasis, but not to sex or tumour size or location. The differences in the positive expression rates of hMSH2, hMLH1, TGFβRII, E-cadherin, cytomembrane β-catenin, cytoplasmic β-catenin, MMP-7 and TIMP-2 were significant between sporadic CRC and Lynch syndrome. The expression of hMSH2 had a positive correlation with that of hMLH1 in Lynch syndrome and sporadic CRC. The expression of TGFβRII had a positive correlation with that of hMSH2, hMLH1 and MMP-7, and a negative correlation with that of TIMP-2. The expression of MMP-7 had a negative correlation with that of TIMP-2 in Lynch syndrome and sporadic CRC. The expression of E-cadherin was positively correlated with that of cytomembrane β-catenin. However, the expression of cytomembrane β-catenin was negatively correlated with that of cytoplasmic β-catenin, and the expression of cytoplasmic β-catenin was positively correlated with that of MMP-7.
CONCLUSION: EMT may play an important role in the development and progression of Lynch syndrome. Lynch syndrome was caused by the mutations of mismatch repair genes, mainly hMSH2 and hMLH1, which also beget the mutational inactivation of TGFβRII. Therefore, the colorectal cancer of Lynch syndrome can escape the inhibitory effect of TGFβ1. However, TGFβ1 can up-regulate the expression of MMP-7 and down-regulate the expression of TIMP-2 in tumors by disassembling the E-cadherin/β-catenin complex in the cytomembrane.
Keywords: Lynch syndrome, Mutation, Epithelial-mesenchymal transition, β-catenin, Mismatch repair gene
Core tip: As a subgroup of colorectal carcinoma (CRC), Lynch syndrome presents better prognosis than sporadic CRC for less invasion and metastasis. Our study found epithelial-mesenchymal transition may play an important role in the development and progression of Lynch syndrome. Lynch syndrome was caused by the mutations of mismatch repair genes, which also beget the mutational inactivation of transforming growth factor-β type II receptor. Therefore, the colorectal cancer of Lynch syndrome can escape the inhibitory effect of transforming growth factor-β type 1 (TGFβ1). However TGFβ1 can up-regulate the expression of matrix metalloproteinase-7 and down-regulate the expression of tissue inhibitor of metalloproteinase-2 in tumors by disassembling the E-cadherin/β-catenin complex in the cytomembrane.
INTRODUCTION
Lynch syndrome is a dominant inherited disease characterized by vertical transmission and familial aggregation of colorectal cancer (CRC)[1-3]. As a subgroup of CRC, Lynch syndrome presents intriguing clinicopathological features[4-7], such as early onset, frequent localization in proximal colon, tendency to develop multiple primary carcinomas, and excessively mucinous and poor differentiation. Nevertheless, the prognosis of Lynch syndrome is known to be better than sporadic CRC. One of the possible explanations depends on the fact that patients with Lynch syndrome are less likely to suffer from metastasis[8].
Many factors have been raised to interpret oncogenesis, tumour invasion, and distant metastasis of CRC. Among these, epithelial-mesenchymal transition (EMT) might play an important role[9-11]. Recent studies have revealed that EMT is not only correlated with tumor growth and differentiation, but also influences tumor invasion and metastasis. Further investigation suggests that EMT might be the downstream of E-cadherin/β-catenin and Transforming growth factor (TGF)β receptor type II (TGFβRII) in the cytomembrane[12]. As a multifunctional cytoplasmic protein β-catenin associates with E-cadherin and cytoskeleton, constituting an E-cadherin/β-catenin complex to maintain normal epithelial polarity and intercellular adhesion; besides, β-catenin also regulates cellular differentiation and proliferation[13]. When the degradation of β-catenin is interfered in the cytoplasm, β-catenin will accumulate in the cytoplasm, resulting in its translocation into the nucleus to promote the transcription of downstream genes involved in the Wnt/β-catenin signaling pathway. Matrix metalloproteinase-7 (MMP-7) is a target gene product of the Wnt/β-catenin signaling pathway[14]. MMP-7 can disassemble the E-cadherin/β-catenin complex to accelerate accumulation of β-catenin in the cytoplasm, resulting in cell proliferation, tumour invasion and metastasis[15,16]. As an endogenous inhibitor of MMP-7, tissue inhibitor of metalloproteinase (TIMP) can induce expression of the developmental EMT transcription factors and lead to down-regulation of epithelial markers and up-regulation of mesenchymal markers[17,18].
We performed this study to investigate whether a different mechanism for regulation of EMT exists in CRC tissues of patients with Lynch syndrome. Moreover, the correlation between EMT and biological behaviour of Lynch syndrome was also our target.
MATERIALS AND METHODS
Ethics
This work has been carried out in accordance with the Declaration of Helsinki (2000) of the World Medical Association. This study was approved ethically by Ethics Committee of the Air Force General Hospital, PLA. All patients provided informed written consent.
Patient tissue
Thirty resected specimens of sporadic CRC, 30 CRC specimens of Lynch syndrome and 8 tumor-adjacent tissues were collected from patients that had undergone operations from May 2007 to June 2012 at the Department of General Surgery, General Hospital of Air Force. Clinical data (including age, gender, and tumor-node-metastasis stage) were collected retrospectively. None of the patients received radiotherapy or chemotherapy before surgery. The clinical diagnosis of Lynch syndrome were established using AmsterdamII criteria. Informed consent was obtained from patients prior to investigation. After surgical resection, all tissue samples were fixed in 4% formalin for 24 h, and then embedded routinely into paraffin for hematoxylin and eosin (H and E) and immunohistochemistry (IHC) staining. Sections were cut from paraffin blocks at 4 μm. H and E-stained samples were individually examined microscopically by two independent pathologists. Clinicopathological characteristics of tumors are provided in Table 1.
Table 1.
Clinical data of patients
| Group | n | Male | Female | Age (χ2 ± SD, yr) |
Lesions |
Dukes stage |
||||
| Colon | Rectum | A | B | C | D | |||||
| Lynch syndrome | 30 | 23 | 7 | 18-68 (46.0 ± 10.6)1 | 21 | 9 | 2 | 19 | 7 | 2 |
| Sporadic CRC | 30 | 20 | 10 | 60-89 (70.3 ± 7.2)2 | 19 | 11 | 1 | 11 | 15 | 3 |
| Adjacent tissues | 8 | 6 | 2 | 45-70 (57.8 ± 8.1)3 | 6 | 2 | ||||
Lynch syndrome vs sporadic CRC (u = -6.637, P = 0.000);
Sporadic CRC vs adjacent tissues (u = -3.447, P = 0.000);
Lynch syndrome vs adjacent tissues (u = -2.238, P = 0.026). CRC: Colorectal carcinoma.
IHC for hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, MMP-7 and TIMP-2 proteins
Mouse monoclonal antihuman antibodies were used for the detection of the hMSH2 (clone FE11; 1:50 dilution; Zhongshan Golden Bridge Biotechnology Inc.; Beijing, China), hMLH1 (clone 14; 1:50 dilution; Zhongshan Golden Bridge Biotechnology Inc.), E-cadherin (clone 4A2C7; 1:50 dilution; Zhongshan Golden Bridge Biotechnology Inc.), β-catenin (clone CAT-5H10; 1:50 dilution; Zhongshan Golden Bridge Biotechnology Inc.), MMP-7 (clone 1D2; 1:50 dilution; Zhongshan Golden Bridge Biotechnology Inc.) and TIMP-2 (clone 3A4; 1:50 dilution; Zhongshan Golden Bridge Biotechnology Inc.), and rabbit polyclonal antihuman antibody was used for the detection of the TGFβRII(sc-400, 1:100 dilution; Santa Cruz Biotechnology, California, United States). Heat-induced epitope retrieval (microwave prewarmed for 5 min in 1 mmol/L EDTA buffer, pH 8) was employed prior to hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin and TIMP-2 staining. MMP-7 did not need epitope retrieval. Primary antibodies were then added and slides incubated for 2 h at 37 °C. Slides were then processed on an immunostainer (LabVision Autostainer 360, Fujian, China). The primary antibody was replaced by phosphate-buffered saline as a negative control to assess the specificity of the antibodies. Haematoxylin-counterstained sections were mounted in aqueous mounting medium and observed under a light microscope.
The IHC staining scores (percentage of stained cells + staining intensity) for hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, MMP-7 and TIMP-2 are given for each case after semi-quantitative evaluation by two independent pathologists. The following scores have been used. Percentage of stained cells was 0, < 10% (1), < 50% (2), < 80% (3), or ≥ 80% (4). Staining intensity was negative (0), weak (1), moderate (2), or strong (3).
Statistical analysis
The results of IHC are expressed as mean ± SD and examined by the χ2 test. The age between the three groups was examined by the Mann-Whitney rank test. Correlations between the expression levels of hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, MMP-7 and TIMP-2 were evaluated by Spearman bivariate correlation test. In all statistical analyses, P < 0.05 was considered significant.
RESULTS
The expression of hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, MMP-7 and TIMP-2 in the three groups is presented in Table 2. We obtained the following findings.
Table 2.
The expression of hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, matrix metalloproteinase-7 and tissue inhibitor of metalloproteinase-2 in three groups
| Group | n |
hMSH2 |
hMLH1 |
TGFβRII |
E-cadherin |
Cytomembrane β-catenin |
Cytoplasmic β-catenin |
MMP-7 |
TIMP-2 |
|||||||||||||
| - | + | ++ | - | + | ++ | - | + | ++ | - | + | - | + | - | + | - | + | ++ | - | + | ++ | ||
| Lynch syndrome | 30 | 23a | 7 | 0 | 14a | 9 | 7 | 18a | 8 | 4 | 4 | 26a | 5 | 25a | 16a | 14 | 16a | 11 | 3 | 11 | 15a | 4 |
| Sporadic CRC | 30 | 11c | 19 | 0 | 5c | 11 | 14 | 8c | 14 | 8 | 12c | 18 | 15c | 15 | 6 | 24c | 4 | 16 | 10c | 24 | 6 | 0 |
| Adjacent tissues | 8 | 0 | 1 | 7e | 0 | 0 | 8e | 0 | 1 | 7e | 0 | 8 | 0 | 8 | 8e | 0 | 8e | 0 | 0 | 8e | 0 | 0 |
Lynch syndrome vs sporadic CRC;
Sporadic CRC vs adjacent tissues;
Lynch syndrome vs adjacent tissues, P < 0.05. CRC: Colorectal carcinoma.
Expression features of hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, MMP-7 and TIMP-2 in three groups
The proteins of hMLH1, TGFβRII, MMP-7 and TIMP-2 were dispersedly expressed as brown granules in the cytoplasm, hMSH2 protein was expressed in the nucleolus, E-cadherin and β-catenin proteins were expressed in the cytomembrane, and β-catenin and TGFβRII proteins also can be expressed in the cytoplasm (Figure 1). The positive expression of hMSH2, hMLH1, and TGFβRII declined from adjacent tissues, sporadic CRC to Lynch syndrome (P < 0.05). The positive expression of E-cadherin and cytomembrane β-catenin declined from adjacent tissues, Lynch syndrome to sporadic CRC (P < 0.05). The positive expression of cytoplasmic β-catenin and MMP-7 increased significantly from adjacent tissues, Lynch syndrome to sporadic CRC (P < 0.05). The positive expression of TIMP-2 was significantly higher in Lynch syndrome than in adjacent tissues and sporadic CRC (P < 0.05).
Figure 1.
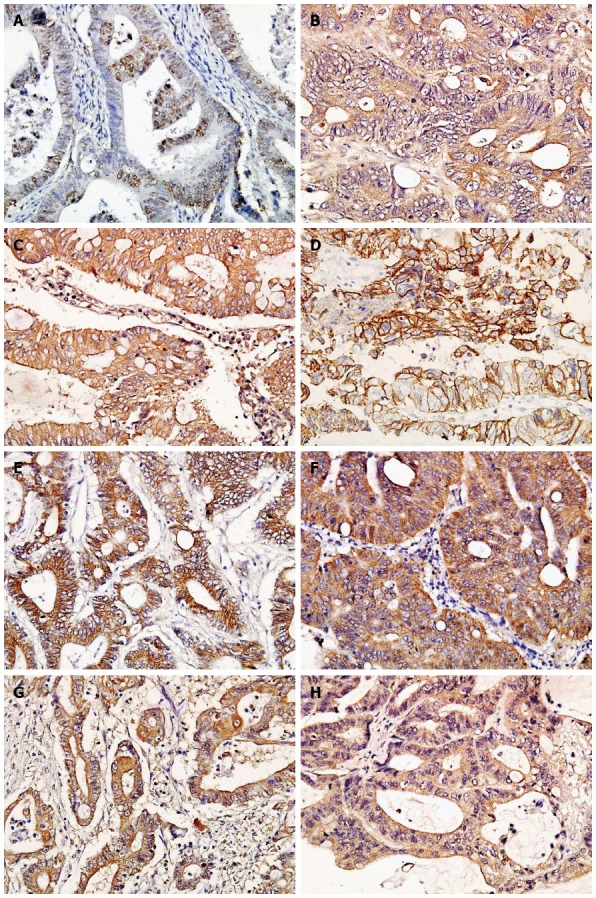
Immunophenotypes of the investigated antigens in Lynch syndrome (× 400 magnification). A: hMSH2 (Positive staining located in the nucleolus); B: hMLH1 (Positive staining located in the cytoplasm); C: TGFβRII (Positive staining located in the cytomembrane and cytoplasm); D: E-cadherin (Positive staining located in the cytomembrane); E: Cytomembrane β-catenin (Positive staining located in the cytomembrane); F: Cytoplasmic β-catenin (Positive staining located in the cytoplasm); G: MMP-7 (Positive staining located in the cytoplasm); H: TIMP-2 (Positive staining located in the cytoplasm).
Correlation between hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, MMP-7, TIMP-2 expression levels and clinicopathological characteristics of tumors
The correlation between protein expression and clinicopathological factors is shown in Table 3. The positive expression rates of hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, MMP-7 and TIMP-2 were significantly related to the depth of invasion and lymph node metastasis, but not to gender or tumour size or location. Positive expression rates of hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, MMP-7 and TIMP-2 were significantly different between samples from sporadic CRC and those from Lynch syndrome.
Table 3.
The relation of expression of hMSH2, hMLH1, TβRII, E-cadherin, β-catenin, matrix metalloproteinase-7 and tissue inhibitor of metalloproteinase-2 with clinicopathological characteristics of Lynch syndrome and sporadic colorectal carcinoma
| Clinicopathological parameter | n | hMSH2 | hMLH1 | TGFβRII | E-cadherin | Cytomembrane β-catenin | Cytoplasmic β-catenin | MMP-7 | TIMP-2 |
| Lynch syndrome | 30 | 7 | 16 | 12 | 26 | 25 | 14 | 14 | 19 |
| Male | 23 | 4 | 10 | 10 | 21 | 20 | 9 | 12 | 14 |
| Female | 7 | 3 | 6 | 2 | 5 | 5 | 5 | 2 | 5 |
| Tumor size | |||||||||
| < 4 cm | 16 | 4 | 8 | 7 | 14 | 13 | 8 | 8 | 10 |
| ≥ 4 cm | 14 | 3 | 8 | 5 | 12 | 12 | 6 | 6 | 9 |
| Position | |||||||||
| Right half of colon | 18 | 4 | 9 | 7 | 16 | 16 | 9 | 10 | 11 |
| Left half of colon | 3 | 1 | 2 | 1 | 3 | 2 | 1 | 1 | 2 |
| Rectum | 9 | 1 | 5 | 4 | 7 | 7 | 4 | 3 | 6 |
| Invasion | |||||||||
| Inner chorion | 21 | 5 | 10 | 11a | 21a | 20a | 7 | 7 | 17ac |
| Outer chorion | 9 | 2 | 6 | 1 | 5 | 5 | 7a | 7a | 2c |
| Metastasis | |||||||||
| No | 21 | 4 | 11 | 11a | 21a | 20a | 7 | 6 | 18ac |
| Yes | 9 | 3 | 5 | 1 | 5 | 5 | 7a | 8a | 1 |
| Sporadic CRC | 30 | 19 | 25 | 22 | 18 | 15 | 24 | 26 | 6 |
| Male | 20 | 12c | 17c | 14 | 11 | 10 | 17 | 18 | 4 |
| Female | 10 | 7 | 8 | 8 | 7 | 5 | 7 | 8 | 2 |
| Tumor size | |||||||||
| < 4 cm | 13 | 9c | 11c | 9 | 8 | 7 | 11 | 10 | 4 |
| ≥ 4 cm | 17 | 10c | 14 | 13 | 10 | 8 | 13 | 16 | 2 |
| Position | |||||||||
| Right half of colon | 14 | 8c | 11 | 12 | 9 | 9 | 12 | 13 | 3 |
| Left half of colon | 5 | 4 | 4 | 3 | 3 | 2 | 3 | 4 | 1 |
| Rectum | 11 | 7c | 10 | 7 | 6 | 4 | 9 | 9 | 2 |
| Invasion | |||||||||
| Inner chorion | 14 | 7 | 11 | 13ac | 12a | 10a | 9 | 10c | 5a |
| Outer chorion | 16 | 12c | 14 | 9b | 6 | 5 | 15a | 16a | 1 |
| Metastasis | |||||||||
| No | 12 | 7c | 10 | 12ac | 10a | 9a | 7 | 8c | 5a |
| Yes | 18 | 12 | 15 | 10c | 8 | 6 | 17a | 18a | 1 |
Compared within the group;
Compared between groups, P < 0.05. CRC: Colorectal carcinoma; TGFβRII: Transforming growth factor-β type II receptor; MMP-7: Matrix metalloproteinase-7.
Expression rates of hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, MMP-7 and TIMP-2 between Lynch syndrome and sporadic CRC
The correlation between expression of hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, MMP-7 and TIMP-2 is shown in Tables 4 and 5. The expression of hMSH2 had a positive correlation with that of hMLH1 in Lynch syndrome and sporadic CRC (rLynch syndrome = 0.835, P = 0.000; rsporadic CRC = 0.549, P = 0.002). The expression of TGFβRII had a positive correlation with that of hMSH2 (rLynch syndrome = 0.592, P = 0.001; rsporadic CRC = 0.444, P = 0.014), hMLH1 (rLynch syndrome = 0.472, P = 0.009. rsporadic CRC = 0.682, P = 0.000) and MMP-7 (rLynch syndrome = 0.735, P = 0.000. rsporadic CRC = 0.792, P = 0.000), and a negative correlation with that of TIMP-2 (rLynch syndrome = - 0.582, P = 0.001; rsporadic CRC = -0.394, P = 0.031). The expression of MMP-7 had a negative correlation with that of TIMP-2 in Lynch syndrome and sporadic CRC (rLynch syndrome = -0.421, P = 0.008; rsporadic CRC = -0.545, P = 0.034). The expression of E-cadherin was positively correlated with that of cytomembrane β-catenin (rLynch syndrome = 0.438, P = 0.015; rsporadic CRC = 0.516, P = 0.004). However, the expression of cytomembrane β-catenin was negatively correlated with that of cytoplasmic β-catenin (rLynch syndrome = -0.449, P = 0.013; rsporadic CRC = -0.389, P = 0.034), and the expression of cytoplasmic β-catenin was positively correlated with that of MMP-7 (rLynch syndrome = 0.393, P=0.031; rsporadic CRC = 0.367, P = 0.046).
Table 4.
The correlation of hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, matrix metalloproteinase-7 and tissue inhibitor of metalloproteinase-2 expression in Lynch syndrome
| Lynch syndrome |
hMSH2 |
hMLH1 |
TGFβRII |
E-cadherin |
Cytomembrane β-catenin |
Cytoplasmic β-catenin |
MMP-7 |
||||||||||||
| (++) | (+) | (-) | (++) | (+) | (-) | (++) | (+) | (-) | (+) | (-) | (+) | (-) | (+) | (-) | (++) | (+) | (-) | ||
| hMLH1 | (++) | 0 | 7 | 0 | |||||||||||||||
| (+) | 0 | 0 | 9 | ||||||||||||||||
| (-) | 0 | 0 | 14 | ||||||||||||||||
| TGFβRII | (++) | 0 | 2 | 2 | 0 | 0 | 4 | ||||||||||||
| (+) | 0 | 4 | 4 | 0 | 0 | 8 | |||||||||||||
| (-) | 0 | 1 | 17 | 7 | 9 | 2 | |||||||||||||
| E-cadherin | (+) | 0 | 7 | 19 | 6 | 7 | 13 | 3 | 7 | 16 | |||||||||
| (-) | 0 | 0 | 4 | 1 | 2 | 1 | 1 | 1 | 2 | ||||||||||
| Cytomembrane β-catenin | (+) | 0 | 5 | 20 | 4 | 8 | 13 | 3 | 6 | 16 | 21 | 4 | |||||||
| (-) | 0 | 2 | 3 | 3 | 1 | 1 | 1 | 2 | 2 | 5 | 0 | ||||||||
| Cytoplasmic β-catenin | (+) | 0 | 3 | 11 | 2 | 6 | 6 | 1 | 2 | 11 | 14 | 0 | 10 | 4 | |||||
| (-) | 0 | 4 | 12 | 5 | 3 | 8 | 3 | 6 | 7 | 12 | 4 | 15 | 1 | ||||||
| MMP-7 | (++) | 0 | 2 | 1 | 2 | 0 | 1 | 1 | 2 | 0 | 1 | 2 | 1 | 2 | 2 | 1 | |||
| (+) | 0 | 4 | 7 | 4 | 1 | 6 | 3 | 2 | 6 | 10 | 1 | 9 | 2 | 9 | 2 | ||||
| (-) | 0 | 1 | 15 | 1 | 8 | 7 | 0 | 4 | 12 | 15 | 1 | 15 | 1 | 3 | 13 | ||||
| TIMP-2 | (++) | 0 | 2 | 2 | 2 | 1 | 1 | 0 | 0 | 4 | 3 | 1 | 3 | 1 | 2 | 2 | 1 | 1 | 2 |
| (+) | 0 | 2 | 13 | 2 | 8 | 5 | 0 | 4 | 11 | 13 | 2 | 13 | 2 | 8 | 7 | 1 | 6 | 8 | |
| (-) | 0 | 3 | 8 | 3 | 0 | 8 | 4 | 4 | 3 | 10 | 1 | 9 | 2 | 4 | 7 | 1 | 4 | 6 | |
Table 5.
The correlation of hMSH2, hMLH1, TGFβRII, E-cadherin, β-catenin, matrix metalloproteinase-7 and tissue inhibitor of metalloproteinase-2 expression in sporadic colorectal carcinoma
| Sporadic CRC |
hMSH2 |
hMLH1 |
TGFβRII |
E-cadherin |
Cytomembrane β-catenin |
Cytoplasmic β-catenin |
MMP-7 |
||||||||||||
| (++) | (+) | (-) | (++) | (+) | (-) | (++) | (+) | (-) | (+) | (-) | (+) | (-) | (+) | (-) | (++) | (+) | (-) | ||
| hMLH1 | (++) | 0 | 14 | 0 | |||||||||||||||
| (+) | 0 | 5 | 6 | ||||||||||||||||
| (-) | 0 | 0 | 5 | ||||||||||||||||
| TGFβRII | (++) | 0 | 4 | 4 | 7 | 0 | 1 | ||||||||||||
| (+) | 0 | 10 | 4 | 7 | 5 | 2 | |||||||||||||
| (-) | 0 | 5 | 3 | 0 | 6 | 2 | |||||||||||||
| E-cadherin | (+) | 0 | 12 | 6 | 12 | 4 | 2 | 6 | 9 | 3 | |||||||||
| (-) | 0 | 7 | 5 | 2 | 7 | 3 | 2 | 5 | 5 | ||||||||||
| Cytomembrane β-catenin | (+) | 0 | 13 | 2 | 9 | 5 | 1 | 6 | 8 | 1 | 12 | 3 | |||||||
| (-) | 0 | 6 | 9 | 5 | 6 | 4 | 2 | 6 | 7 | 6 | 9 | ||||||||
| Cytoplasmic β-catenin | (+) | 0 | 14 | 10 | 13 | 8 | 3 | 7 | 11 | 6 | 15 | 9 | 10 | 14 | |||||
| (-) | 0 | 5 | 1 | 1 | 3 | 2 | 1 | 3 | 2 | 3 | 3 | 5 | 1 | ||||||
| MMP-7 | (++) | 0 | 6 | 4 | 6 | 2 | 2 | 7 | 3 | 0 | 8 | 2 | 5 | 5 | 7 | 3 | |||
| (+) | 0 | 12 | 4 | 6 | 9 | 1 | 1 | 11 | 4 | 10 | 6 | 10 | 6 | 14 | 2 | ||||
| (-) | 0 | 1 | 3 | 2 | 0 | 2 | 0 | 0 | 4 | 0 | 4 | 0 | 4 | 3 | 1 | ||||
| TIMP-2 | (++) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| (+) | 0 | 4 | 2 | 4 | 0 | 2 | 2 | 2 | 2 | 5 | 1 | 4 | 2 | 1 | 5 | 4 | 2 | 0 | |
| (-) | 0 | 15 | 9 | 10 | 11 | 3 | 6 | 12 | 6 | 13 | 11 | 11 | 13 | 23 | 1 | 6 | 14 | 4 | |
DISCUSSION
Lynch syndrome is caused by germline mutations in mismatch repair genes, mainly hMSH2 and hMLH1[19-21]. Patients with Lynch syndrome have better prognosis due to less metastases compared with sporadic CRC. However, specimens from Lynch syndrome patients are reported to be less differentiated than sporadic CRC, and the survival benefits of Lynch syndrome remain unexplained. Immunological response of intraepithelial cytotoxic T lymphocytes against tumor cells in Lynch syndrome has been proposed as a possible explanation. Studies also demonstrated that EMT might also be an important regulator in tumorigenesis[22]. The EMT status in Lynch syndrome was investigated in the present study, and we demonstrated significantly lower expression levels of hMSH2, hMLH1, cytoplasmic β-catenin, TGFβRII and MMP-7, and higher expression levels of E-cadherin, cytomembrane β-catenin and TIMP-2 in Lynch syndrome. The mutations of hMSH2 and hMLH1 in Lynch syndrome are in accordance with AmsterdamII criteria. So ICH might be an easy and cheap way to detect hMLH1 and hMSH2 expression for the diagnosis of Lynch syndrome.
Many alterations of structural or functional genes may exist in tumorigenesis. TGFβRII is a downstream gene of MMR[23]. When the mutations of hMSH2 and hMLH1 results in Lynch syndrome tumorigenesis, mutations of TGFβRII often occur. This may make the tumors escape the inhibition by TGF-β and promote the development of tumors[24]. The anti-mitogenic and pro-metastatic effects of TGF-β can co-exist simultaneously with TGF-β to accelerate metastatic tumour progression[25]. According to this theory, the onset age of Lynch syndrome will be earlier than sporadic CRC, and the developing speed of Lynch syndrome tumours will be faster than sporadic CRC. This may be a reason why Lynch syndrome has an early onset, tendency to develop multiple primary carcinomas, and excessively mucinous and poor differentiation. Our study showed that the abnormal expression of TGFβRII is significantly correlated to the depth of invasion and lymph node metastasis. This indicates that mutation of TGFβRII not only associates with carcinogenesis of colorectal cancer, but also affects the invasion and metastasis of the tumors.
E-cadherin, β-catenin, MMP-7 and TIMP-2 are proteins correlating with EMT in CRC[26-29]. Our study showed that as tumour invasion became deeper and lymph node metastasis occurred, the expression of E-cadherin and plasmamembrane β-catenin markedly decreased, and the expression of MMP-7 and cytoplasmic β-catenin markedly increased. Moreover, there have been significant differences in the expression of E-cadherin, β-catenin, MMP-7 and TIMP-2 between Lynch syndrome and sporadic CRC. The proportional unbalance of MMP-7(+)/TIMP-2(-) and cytoplasmic β-catenin/cytomembrane β-catenin was obviously heightened. This may explain why Lynch syndrome has less invasive/metastasis and better prognosis than sporadic CRC. First, sporadic CRC is mainly caused by the mutation of the APC gene[30,31], and the inactive APC protein cannot disassemble the cytoplasmic β-catenin complex. As a consequence, β-catenin will accumulate in the cytoplasm and promote the expression of MMP-7, and this will accelerate the invasion and metastasis of tumours. Second, there is crosstalk between TGF-β/Smad and Wnt/β-catenin signalling pathways[32-34], and TGF-β can make use of Smad2 to disassemble the E-cadherin/β-catenin complex. But in Lynch syndrome, the mutations of MMR lead to mutational inactivation of TGFβRII. This will reduce the function of TGF-β to disassemble the E-cadherin/β-catenin complex, so the expression of E-cadherin and plasmamembrane β-catenin will increase in Lynch syndrome than in sporadic CRC. Recently there is a study reporting that exon 3 of the β-catenin gene in Lynch syndrome is more likely to be mutated than in sporadic CRC without functional inhibition[35].
The chemosynthetic inhibitors of MMP and β-cat
enin[36-40], such as batimastat and marimastat, have been used in clinical treatment for the invasion and metastasis of tumours. But this kind of drug is very expensive now. Our study shows that MMP-7 and cytoplasmic β-catenin were not expressed in all CRC specimens, and their expression levels are not similar in different stages of tumogenesis. So it is worthy to optimize the timing to use the inhibitors of MMP and β-catenin to cure patients with CRC. We think it is feasible to examine the levels of MMP-7 and β-catenin in tumour tissue and (or) serum before the prescription of inhibitors of MMP-7 and β-catenin to patients. In a word, our study suggests that MMP-7 and β-catenin inhibitors might be tomorrow agents for the treatment of tumour invasion and metastasis.
COMMENTS
Background
Lynch syndrome significantly differs from sporadic colorectal cancer (CRC) in biological behavior, but the reasons remain largely unexplained. This study explored multiple molecules regulating the epithelial-mesenchymal transition in CRC by using Immunohistochemical staining, which can help us interpret the biological behaviour of Lynch syndrome tumors.
Research frontiers
In this study, the authors investigated the expression of seven molecules by IHC staining, which is very easy and cheap to be used in clinic for examining molecular targets of Lynch syndrome at the protein level, and the results are helpful in understanding the biological behaviour of Lynch syndrome.
Innovations and breakthroughs
In this study, the authors explored the expression of seven molecules during Lynch syndrome carcinogenesis, development, invasion and metastasis, and the expression of these molecules was previously seldom simultaneously detected in Lynch syndrome. The results can help explain the biological behavior of Lynch syndrome.
Applications
IHC staining of hMSH2 and hMLH1 for diagnosing Lynch syndrome can facilitate rapid translation of molecular discoveries to clinical applications. IHC staining of hMSH2 and hMLH1 for diagnosing Lynch syndrome is fast, convenient and economical.
Terminology
Lynch syndrome is a dominant inherited disease with vertical transmission and aggregation of cancers. Lynch syndrome presents several particular clinicopathological features, such as early onset, frequent localization in proximal colon, proclivity of synchronous and metachronous tumours, excessive mucinous and poorly differentiated tumours, and high incidence of extra-colonic malignancies. But the prognosis of Lynch syndrome is better than that of sporadic CRC.
Peer review
The manuscript entitled “Epithelial-mesenchymal transition in cancerous tissue of hereditary nonpolyposis colorectal carcinoma” was carefully reviewed. The authors conducted an IHC analysis of seven proteins between HNPCC and sporadic CRC patients. The study is very interesting.
Footnotes
Supported by Capital Citizen Health Cultivation Project, No. Z131100004013021; and General Projects of the Chinese PLA “Twelfth Five-Year” Logistics Research Subject, No. CWS11J193
P- Reviewers: Gruber SB, Lehmann U S- Editor: Qi Y L- Editor: Wang TQ E- Editor: Ma S
References
- 1.Petersen HV, Ladelund S, Carlsson C, Nilbert M. Sense of coherence and self-concept in Lynch syndrome. Hered Cancer Clin Pract. 2013;11:7. doi: 10.1186/1897-4287-11-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bansidhar BJ, Silinsky J. History and pathogenesis of lynch syndrome. Clin Colon Rectal Surg. 2012;25:63–66. doi: 10.1055/s-0032-1313776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Vasen HF, de Vos Tot Nederveen Cappel WH. A hundred years of Lynch syndrome research (1913-2013) Fam Cancer. 2013;12:141–142. doi: 10.1007/s10689-013-9654-7. [DOI] [PubMed] [Google Scholar]
- 4.Bansidhar BJ. Extracolonic manifestations of lynch syndrome. Clin Colon Rectal Surg. 2012;25:103–110. doi: 10.1055/s-0032-1313781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Mishra N, Hall J. Identification of patients at risk for hereditary colorectal cancer. Clin Colon Rectal Surg. 2012;25:67–82. doi: 10.1055/s-0032-1313777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bleiker EM, Esplen MJ, Meiser B, Petersen HV, Patenaude AF. 100 years Lynch syndrome: what have we learned about psychosocial issues? Fam Cancer. 2013;12:325–339. doi: 10.1007/s10689-013-9653-8. [DOI] [PubMed] [Google Scholar]
- 7.Lynch HT, Lynch PM. Colorectal cancer: Update on the clinical management of Lynch syndrome. Nat Rev Gastroenterol Hepatol. 2013;10:323–324. doi: 10.1038/nrgastro.2013.70. [DOI] [PubMed] [Google Scholar]
- 8.Barrow E, Hill J, Evans DG. Cancer risk in Lynch Syndrome. Fam Cancer. 2013;12:229–240. doi: 10.1007/s10689-013-9615-1. [DOI] [PubMed] [Google Scholar]
- 9.Todosi AM, Gavrilescu MM, Aniţei GM, Filip B, Scripcariu V. Colon cancer at the molecular level--usefulness of epithelial-mesenchymal transition analysis. Rev Med Chir Soc Med Nat Iasi. 2012;116:1106–1111. [PubMed] [Google Scholar]
- 10.Peláez-García A, Barderas R, Torres S, Hernández-Varas P, Teixidó J, Bonilla F, de Herreros AG, Casal JI. FGFR4 role in epithelial-mesenchymal transition and its therapeutic value in colorectal cancer. PLoS One. 2013;8:e63695. doi: 10.1371/journal.pone.0063695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Takahashi Y, Sawada G, Kurashige J, Matsumura T, Uchi R, Ueo H, Ishibashi M, Takano Y, Akiyoshi S, Iwaya T, et al. Tumor-derived tenascin-C promotes the epithelial-mesenchymal transition in colorectal cancer cells. Anticancer Res. 2013;33:1927–1934. [PubMed] [Google Scholar]
- 12.Zheng D, Song T, Zhongliu X, Wu M, Liang J, Liu Y. Downregulation of transforming growth factor-β type II receptor prohibit epithelial-to-mesenchymal transition in lens epithelium. Mol Vis. 2012;18:1238–1246. [PMC free article] [PubMed] [Google Scholar]
- 13.Okumura N, Akutsu H, Sugawara T, Miura T, Takezawa Y, Hosoda A, Yoshida K, Ichida JK, Yamada M, Hamatani T, et al. β-catenin functions pleiotropically in differentiation and tumorigenesis in mouse embryo-derived stem cells. PLoS One. 2013;8:e63265. doi: 10.1371/journal.pone.0063265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.He W, Tan RJ, Li Y, Wang D, Nie J, Hou FF, Liu Y. Matrix metalloproteinase-7 as a surrogate marker predicts renal Wnt/β-catenin activity in CKD. J Am Soc Nephrol. 2012;23:294–304. doi: 10.1681/ASN.2011050490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kang YJ, Park HJ, Chung HJ, Min HY, Park EJ, Lee MA, Shin Y, Lee SK. Wnt/β-catenin signaling mediates the antitumor activity of magnolol in colorectal cancer cells. Mol Pharmacol. 2012;82:168–177. doi: 10.1124/mol.112.078535. [DOI] [PubMed] [Google Scholar]
- 16.Matono H, Oda Y, Nakamori M, Tamiya S, Yamamoto H, Yokoyama R, Saito T, Iwamoto Y, Tsuneyoshi M. Correlation between beta-catenin widespread nuclear expression and matrix metalloproteinase-7 overexpression in sporadic desmoid tumors. Hum Pathol. 2008;39:1802–1808. doi: 10.1016/j.humpath.2008.05.005. [DOI] [PubMed] [Google Scholar]
- 17.Aresu L, Benali S, Garbisa S, Gallo E, Castagnaro M. Matrix metalloproteinases and their role in the renal epithelial mesenchymal transition. Histol Histopathol. 2011;26:307–313. doi: 10.14670/HH-26.307. [DOI] [PubMed] [Google Scholar]
- 18.Jung YS, Liu XW, Chirco R, Warner RB, Fridman R, Kim HR. TIMP-1 induces an EMT-like phenotypic conversion in MDCK cells independent of its MMP-inhibitory domain. PLoS One. 2012;7:e38773. doi: 10.1371/journal.pone.0038773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tanyi M, Olasz J, Tanyi JL, Tóth L, Antal-Szalmás P, Bubán T, András C, Urbancsek H, Garami Z, Csuka O, et al. Q48P mutation in the hMLH1 gene associated with Lynch syndrome in three Hungarian families. Fam Cancer. 2012;11:519–524. doi: 10.1007/s10689-012-9515-9. [DOI] [PubMed] [Google Scholar]
- 20.Wei W, Liu F, Liu L, Li Z, Zhang X, Jiang F, Shi Q, Zhou X, Sheng W, Cai S, et al. Distinct mutations in MLH1 and MSH2 genes in hereditary non-polyposis colorectal cancer (HNPCC) families from China. BMB Rep. 2011;44:317–322. doi: 10.5483/BMBRep.2011.44.5.317. [DOI] [PubMed] [Google Scholar]
- 21.Liu T. Mutational screening of hMLH1 and hMSH2 that confer inherited colorectal cancer susceptibility using denature gradient gel electrophoresis (DGGE) Methods Mol Biol. 2010;653:193–205. doi: 10.1007/978-1-60761-759-4_11. [DOI] [PubMed] [Google Scholar]
- 22.Sánchez-Tilló E, de Barrios O, Siles L, Cuatrecasas M, Castells A, Postigo A. β-catenin/TCF4 complex induces the epithelial-to-mesenchymal transition (EMT)-activator ZEB1 to regulate tumor invasiveness. Proc Natl Acad Sci USA. 2011;108:19204–19209. doi: 10.1073/pnas.1108977108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Best A, Matheson E, Minto L, Hall AG, Irving JA. Mismatch repair and the downstream target genes, PAX5 and Ikaros, in childhood acute lymphoblastic leukemia. Leuk Res. 2010;34:1098–1102. doi: 10.1016/j.leukres.2010.02.017. [DOI] [PubMed] [Google Scholar]
- 24.Siegel PM, Shu W, Cardiff RD, Muller WJ, Massagué J. Transforming growth factor beta signaling impairs Neu-induced mammary tumorigenesis while promoting pulmonary metastasis. Proc Natl Acad Sci USA. 2003;100:8430–8435. doi: 10.1073/pnas.0932636100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Drabsch Y, ten Dijke P. TGF-β signaling in breast cancer cell invasion and bone metastasis. J Mammary Gland Biol Neoplasia. 2011;16:97–108. doi: 10.1007/s10911-011-9217-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ludwig K, Tse ES, Wang JY. Colon cancer cells adopt an invasive phenotype without mesenchymal transition in 3-D but not 2-D culture upon combined stimulation with EGF and crypt growth factors. BMC Cancer. 2013;13:221. doi: 10.1186/1471-2407-13-221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kouso H, Yano T, Maruyama R, Shikada Y, Okamoto T, Haro A, Kakeji Y, Maehara Y. Differences in the expression of epithelial-mesenchymal transition related molecules between primary tumors and pulmonary metastatic tumors in colorectal cancer. Surg Today. 2013;43:73–80. doi: 10.1007/s00595-012-0344-0. [DOI] [PubMed] [Google Scholar]
- 28.Cho SH, Park YS, Kim HJ, Kim CH, Lim SW, Huh JW, Lee JH, Kim HR. CD44 enhances the epithelial-mesenchymal transition in association with colon cancer invasion. Int J Oncol. 2012;41:211–218. doi: 10.3892/ijo.2012.1453. [DOI] [PubMed] [Google Scholar]
- 29.Forsyth CB, Tang Y, Shaikh M, Zhang L, Keshavarzian A. Alcohol stimulates activation of Snail, epidermal growth factor receptor signaling, and biomarkers of epithelial-mesenchymal transition in colon and breast cancer cells. Alcohol Clin Exp Res. 2010;34:19–31. doi: 10.1111/j.1530-0277.2009.01061.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Curia MC, De Iure S, De Lellis L, Veschi S, Mammarella S, White MJ, Bartlett J, Di Iorio A, Amatetti C, Lombardo M, et al. Increased variance in germline allele-specific expression of APC associates with colorectal cancer. Gastroenterology. 2012;142:71–77.e1. doi: 10.1053/j.gastro.2011.09.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Roy S, Majumdar AP. Cancer Stem Cells in Colorectal Cancer: Genetic and Epigenetic Changes. J Stem Cell Res Ther. 2012;(6):pii: 10342. doi: 10.4172/2157-7633.S7-006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mauviel A, Nallet-Staub F, Varelas X. Integrating developmental signals: a Hippo in the (path)way. Oncogene. 2012;31:1743–1756. doi: 10.1038/onc.2011.363. [DOI] [PubMed] [Google Scholar]
- 33.Hirota M, Watanabe K, Hamada S, Sun Y, Strizzi L, Mancino M, Nagaoka T, Gonzales M, Seno M, Bianco C, et al. Smad2 functions as a co-activator of canonical Wnt/beta-catenin signaling pathway independent of Smad4 through histone acetyltransferase activity of p300. Cell Signal. 2008;20:1632–1641. doi: 10.1016/j.cellsig.2008.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Osaki LH, Gama P. MAPKs and Signal Transduction in the Control of Gastrointestinal Epithelial Cell Proliferation and Differentiation. Int J Mol Sci. 2013;14:10143–10161. doi: 10.3390/ijms140510143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Johnson V, Volikos E, Halford SE, Eftekhar Sadat ET, Popat S, Talbot I, Truninger K, Martin J, Jass J, Houlston R, et al. Exon 3 beta-catenin mutations are specifically associated with colorectal carcinomas in hereditary non-polyposis colorectal cancer syndrome. Gut. 2005;54:264–267. doi: 10.1136/gut.2004.048132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Park CH, Shin TK, Lee HY, Kim SJ, Lee WS. Matrix metalloproteinase inhibitors attenuate neuroinflammation following focal cerebral ischemia in mice. Korean J Physiol Pharmacol. 2011;15:115–122. doi: 10.4196/kjpp.2011.15.2.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Chaudhary AK, Pandya S, Ghosh K, Nadkarni A. Matrix metalloproteinase and its drug targets therapy in solid and hematological malignancies: an overview. Mutat Res. 2013;753:7–23. doi: 10.1016/j.mrrev.2013.01.002. [DOI] [PubMed] [Google Scholar]
- 38.Sinno M, Biagioni S, Ajmone-Cat MA, Pafumi I, Caramanica P, Medda V, Tonti G, Minghetti L, Mannello F, Cacci E. The matrix metalloproteinase inhibitor marimastat promotes neural progenitor cell differentiation into neurons by gelatinase-independent TIMP-2-dependent mechanisms. Stem Cells Dev. 2013;22:345–358. doi: 10.1089/scd.2012.0299. [DOI] [PubMed] [Google Scholar]
- 39.Benjamin MM, Khalil RA. Matrix metalloproteinase inhibitors as investigative tools in the pathogenesis and management of vascular disease. EXS. 2012;103:209–279. doi: 10.1007/978-3-0348-0364-9_7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Halbedl S, Kratzer MC, Rahm K, Crosta N, Masters KS, Zippert J, Bräse S, Gradl D. Synthesis of novel inhibitors blocking Wnt signaling downstream of β-catenin. FEBS Lett. 2013;587:522–527. doi: 10.1016/j.febslet.2013.01.034. [DOI] [PubMed] [Google Scholar]