

Abstract
Recent research has focused on the different types of compass cues available to ball-rolling beetles for orientation, but little is known about the relative precision of each of these cues and how they interact. In this study, we find that the absolute orientation error of the celestial compass of the day-active dung beetle Scarabaeus lamarcki doubles from 16° at solar elevations below 60° to an error of 29° at solar elevations above 75°. As ball-rolling dung beetles rely solely on celestial compass cues for their orientation, these insects experience a large decrease in orientation precision towards the middle of the day. We also find that in the compass system of dung beetles, the solar cues and the skylight cues are used together and share the control of orientation behaviour. Finally, we demonstrate that the relative influence of the azimuthal position of the sun for straight-line orientation decreases as the sun draws closer to the horizon. In conclusion, ball-rolling dung beetles possess a dynamic celestial compass system in which the orientation precision and the relative influence of the solar compass cues change over the course of the day.
Keywords: orientation, solar compass, polarized light, dung beetle, vision, navigation
1. Introduction
Foraging dung beetles use a keen sense of smell to locate their food of preference [1]. Once a suitable pile of dung has been located, some species quickly sculpt a piece into a ball and start to roll it away along a straight path [2–6]. The end destination for the ball is, at this point, unknown [7]. The straight-line departure helps to minimize the time that the dung beetle spends near the dung pat, where foraging conspecifics congregate and frequently try to steal a ready-made ball. After some distance, the beetle finds a suitable patch of ground and buries itself together with its ball of dung. The ball is then slowly consumed in the safety of the ground [1].
Leaving a defined place might at first glance seem simple but is, as it turns out, impossible without an external directional reference—i.e. some form of compass [8]. This is true even for ensuring progress away from a dung pile. Foraging dung beetles use a number of celestial compass cues to guide them along their straight tracks: the sun, the moon, the pattern of polarized light and even the Milky Way [3,4,6,9–12].
Under thick clouds, however, the position of the sun or the moon can no longer be recognized, the stars are no longer visible and the degree of polarization in the sky decreases dramatically [13,14]. This renders the celestial compass system of the dung beetles effectively useless to the point that, on these rare, overcast days, the beetles roll their food in circles [2]. A similar problem occurs on a more regular basis: as the sun/moon draws closer to the zenith, the accuracy of the celestial compass decreases [15]. To compensate for this loss in accuracy around noon, navigating ants rely instead on the direction of the wind [15], whereas sandhoppers rather rely on their magnetic compass [16,17]. Neither of these seems to be an option for the celestially fixated beetles, which rely on a single compass system for orientation [2]. In the first part of this study, we test how the reliability of a purely celestial compass system changes over the course of the day.
In many of the diurnal dung beetle species tested so far, the primary cue for straight-line orientation appears to be the sun [12]. Consequently, if the apparent position of the sun is changed by 180° using a mirror, the majority of beetles change their bearing [12]. In a recent study on the diurnal dung beetle Scarabaeus lamarcki, however, we found that only half of the beetles changed their course in response to a change in the apparent position of the sun [18]. The beetles that continued along their original paths were most probably guided by one, or several, of the skylight cues that span the entire sky. These cues include the pattern of polarized light, and gradients of colour and intensity [19–22]. Dung beetles have been demonstrated to exploit polarized skylight cues for their orientation [6,10], while the role of the other skylight cues remains to be tested.
Studies on ants navigating towards a nest suggest that the influence of different navigational systems (celestial compass, wind direction and landmarks) varies with the relative precision afforded by each of these cues at different times of the day [15,23]. Dung beetles, however, rely solely on celestial cues. As the sun draws closer to the horizon, the degree of polarization in the zenith of the sky gradually increases and reaches its maximum when the sun is closest to the horizon [15,22]. This raises the question of the relative influence of different celestial cues (the sun, the pattern of polarized light, and gradients of colour and intensity) in a celestial compass system at different elevations of the sun. In the second part of this study, we test how ball-rolling beetles respond to an apparent change in the position of the sun over the course of the day.
2. Material and methods
(a). Animals and field site
Individuals of the diurnal dung beetle S. lamarcki were collected within the game farm ‘Stonehenge’, 70 km northwest of Vryburg, North West province, South Africa (24.3° E, 26.4° S) in February and November 2012, and in February 2013. The experiments were conducted at the same location in February 2012 and 2013, and the animals were tested in the same year as they were caught.
(b). Precision of the celestial compass
The aim of this experiment was to determine the precision of the celestial compass at different solar elevations: 75–78° (high elevations), 45–60° (medium elevations) and 20–30° (low elevations). A circular wooden board (1 m diameter) with sand glued to its top surface was levelled to create a perfectly flat ball-rolling arena. Individual beetles were placed alongside their balls in the centre of the arena. Each beetle then climbed on top of its ball, performed an ‘orientation dance’ [18] and began to roll its ball towards the edge of the arena along the chosen direction. The beetle was then placed back alongside its ball in the centre of the arena and allowed to roll out a second time. The compass bearings of the two consecutive rolls were recorded as the beetle crossed the edge of the arena, and the absolute angular difference was calculated to determine the orientation error of the celestial compass system (figure 1a). An orientation error of 0° indicates the highest precision possible. The precision of the beetles’ celestial compass was tested under three conditions: (1) with a full view of the sky, (2) with the sun hidden from view by a square board (100 × 75 cm, placed approx. 1.5 m away from the beetle) and (3) with the sun hidden from view only during the second exit. This last condition was designed to test the effect of the removal of the sun once the beetle had set a bearing in its presence.
Figure 1.
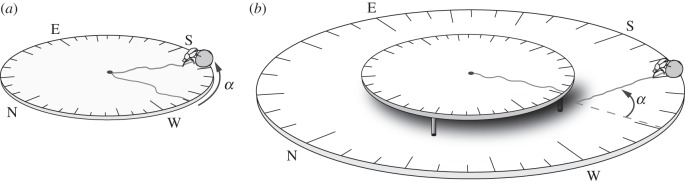
Experimental set-ups. (a) A circular wooden board (1 m diameter) was covered with sand to create a perfectly flat arena. A beetle was placed alongside its ball in the centre of the arena and allowed to roll its ball towards the edge of the arena, were the bearing of its path was recorded. The beetle was then placed back alongside its ball in the centre of the arena and allowed to roll out of the arena a second time. The compass bearing was again recorded at the edge of the arena. The absolute angular differences between the two bearings (α) were analysed to determine the orientation error of the celestial compass system. (b) A 1 m diameter, 5 cm high raised circular arena was positioned at the centre of a second, larger arena (2 m diameter). Both arenas were flat and covered in sand. A beetle was placed alongside its balls at the centre of the raised arena and allowed to roll its ball out, first to the edge of the inner arena (from where it fell off its edge) and then to the edge of the bigger arena. The compass bearing was recorded when the beetle reached the edge of the raised arena, and the bearing taken after the fall was recorded at the edge of the second arena. The absolute angular difference between the two paths (α) was calculated from the two different bearings.
To test for a possible effect of heat stress on orientation performance, the temperature of the surface of the arena was recorded over an 8 cm diameter circle (non-contact infrared thermometer, CIR 350) at the centre of the board just after the beetle commenced rolling the second time with a full view of the sky (condition 1). As can be expected, the temperature of the arena varies with the elevation of the sun (high: 45.5 ± 0.7°C; medium: 41.4 ± 0.4°C; low: 38.7 ± 0.4°C; ANOVA, F2/87 = 49.53, p < 0.001), with a 10°C difference between midday and the late afternoon. At surface temperatures above 50°C, S. lamarcki climb on their dung balls more frequently and use them as a thermal refuge to cool down while rolling across hot soil [24]. In these experiments, the temperature of the arena never passed 50°C, and we did not observe any of the known behaviours related to heat stress. Furthermore, we find no significant positive association between the temperature of the arena and the orientation errors of the beetles (Spearman's rank-order correlation, r = 0.004, d.f. = 87, p = 0.97). Thus, we consider it unlikely that the rise in temperature at higher sun elevations influenced the outcome of the experiments.
In the second set of experiments, the orientation errors at different elevations of the sun were analysed from the bearings taken before and after a 5 cm drop from the edge of an arena. A 1 m diameter, 5 cm high raised circular arena was positioned in the centre of a second, larger arena (2 m diameter). The beetles were individually placed alongside their balls at the centre of the raised arena and allowed to roll out, first to the edge of the inner arena (from where they fell) and then to the edge of the bigger arena. The compass bearing taken before the fall was recorded at the edge of the raised arena and the bearing taken after the fall was calculated from this first bearing and the compass angle at which the beetle exited the edge of the second arena (figure 1b). Again, the absolute angular difference between the two bearings was used to define the orientation errors at the three different ranges of solar elevations.
(c). Cue hierarchy of the celestial compass
In the third set of experiments, the impact of the sun on straight-line orientation was evaluated by artificially changing its position during a beetle's second exit. Similar to the experiments above, beetles were first allowed to exit the 1 m diameter wooden arena with a full view of the sun. When a beetle reached the edge of the arena, its bearing was recorded and the beetle was placed back into the centre of the arena. During the second exit of the beetle, the sun was reflected by 180° from its natural position in the sky by the use of a mirror (30 × 30 cm), while keeping its elevation constant. Simultaneously, the beetle's view of the real sun was occluded using a wooden board (100 × 75 cm). The exit bearing of beetles rolling under the reflected sun was recorded, and the absolute angular difference between the first and the second bearing was calculated to determine the sun's importance relative to other orientation cues in the compass system of the beetles. An angular difference around 180° between the two paths indicates that the beetles use the sun as their main orientation cue, while a difference around 0° indicates that they instead preferentially orient to skylight cues, such as the pattern of polarized light, spectral gradients and intensity gradients. For each category of sun elevations (high, medium and low), we would expect 95% of the population to fall within two and a half orientation errors (as given by experiment (a)) around this expected bearing (grey sectors in figure 3).
Figure 3.
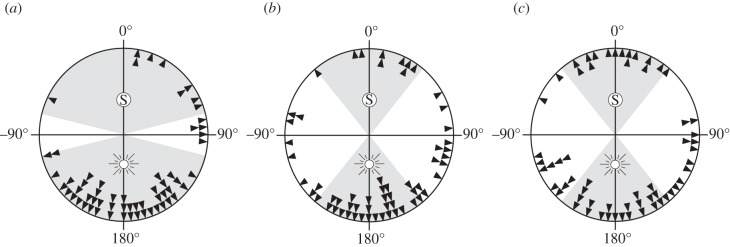
The effect of the mirrored sun on straight-line orientation at different elevations of the sun. To investigate the influence that the solar compass has on straight-line orientation in S. lamarcki, the difference between the bearings taken with a view of the real sun and with a view of a mirrored sun (black arrows) was determined for 60 beetles at different elevations of the sun: (a) 75–78° (high elevations), (b) 45–60° (medium elevations) and (c) 15–30° (low elevations). We expect beetles that use the sun as their primary orientation cue to change their rolling direction by 180° ± the error of the compass (grey sectors with sun symbol), while beetles orienting preferentially to the skylight cues should not change their rolling direction by more than the error of the compass (grey sectors marked with S). At high solar elevations, 77% of the dung beetles chose a direction consistent with a 180° turn (a), and at medium and low solar elevations, 52 and 37% of the population showed a similar response (b,c).
The experimental conditions were presented to the beetles in randomized orders and the beetles were put in from the south side of the arena. The dataset obtained from the experiments above can be downloaded from datadryad.org.
3. Precision of the celestial compass at different sun elevations
The precision of the dung beetle celestial compass was tested at different elevations of the sun (figure 1a). We found that the orientation errors at low (16.0 ± 2.2°, mean ± s.e.m., n = 30) and medium solar elevations (16.0 ± 3.4°) were remarkably similar, and nearly half the size of the errors recorded at high solar elevations (29.3 ± 5.5°; figure 2). The orientation precision of the beetles’ celestial compass at solar elevations above 75° was significantly worse than that at solar elevations below 60° (ANOVA, F2/87 = 3.83, p = 0.03).
Figure 2.
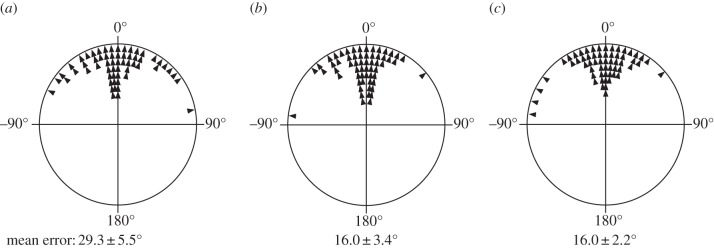
The precision of the dung beetle celestial compass at different elevations of the sun. The orientation error of the dung beetle celestial compass was determined by letting 60 beetles roll out twice from the centre to the edge of the arena at different elevations of the sun: (a) 75–78° (high elevations), (b) 45–60° (medium elevations) and (c) 15–30° (low elevations). The angular difference between the two bearings was calculated (black arrows) and used to define the absolute orientation errors at the three different ranges of solar elevations (a–c). Numbers in text and figure show mean ± s.e.m.
In this first set of experiments, the beetles were picked up, separated from their balls and placed back at the centre of the arena by the experimenter. Although this method allows us to precisely measure and compare the orientation performance of the beetles under a range of conditions (see §4), the extensive handling of the beetles could potentially induce an artificially large orientation error. To test for this, we also measured the orientation errors of beetles that rolled out from the centre of a 5 cm high, raised circular platform until they fell from its edge (figure 1b). This experiment simulates a more natural situation because, when rolling in natural terrain, dung beetles frequently fall into depressions in the uneven ground. The angular difference between the directions taken before and after the drop from the platform thus defines the orientation errors made by the beetles after experiencing a more natural, less intrusive disturbance to the roll path. Again, we found that the orientation error at high solar elevations (31.0 ± 4.1°) was significantly larger than that at medium (16.9 ± 2.7°) or low solar elevations (20.7 ± 2.9°) (n = 60, t-test, high–medium: T118 = 3.12, p < 0.01; high–low: T118 = 2.21, p = 0.01; ANOVA, F2/177 = 5.06, p < 0.01). Overall, the orientation performance of beetles that fell from the raised platform was somewhat worse (possibly owing to sideways movements during the fall or a slight change in the shape of the ball after the fall) than the performance of beetles that were picked up, separated from their balls and placed back at the centre of the arena. This demonstrates that the additional handling of beetles in the first set of experiments did not induce an artificially large orientation error.
Considering the geometry of the sky surrounding an orienting dung beetle, it does not come as a surprise that the precision of the sun compass decreases at higher solar elevations. In an eye, the optical axes of two adjacent receptors differ from each other horizontally by the inter-receptor angle Δφ. This angle covers a certain azimuthal range, which defines the optimal resolution of the sun compass. At higher elevations, the azimuthal range covered by Δφ becomes larger—about twice as large at 75° than that at 30° elevation—making it more difficult to pinpoint the exact azimuth of the sun. In other words, the higher the elevation of the sun, the lower the accuracy with which its azimuthal position can be determined. High sun elevations also result in low degrees of polarization in the zenith of the sky [22]. This reduces the input to the skylight compass, while at the same time the accuracy of the solar compass is low.
A decrease in the performance of a celestial compass at high solar elevations has also been observed in the homing desert ant Cataglyphis fortis [15] and in beach-dwelling sandhoppers [16,19]. In nature, however, these animals continue to navigate with high precision in the middle of the day by extracting additional directional information from terrestrial landmarks, wind direction or the magnetic field [15,16,23]. These options do not seem to be available to dung beetles, which demonstrate a drastic decrease in orientation performance even when given full access to all of these cues. This finding supports the results from a recent study, which suggests that dung beetles rely exclusively on their celestial compass to orient [2]. However, the orientation precision that this single system offers, with a drastic decrease in precision around noon, will only temporarily impair the foraging efficiency of a ball-rolling beetle that can bury itself in the ground at any time. For a homing insect, the same error could instead be fatal. In the field, we rarely see S. lamarcki active in the middle of the day, and a similar bimodal activity pattern has been described for many other dung beetle species [25]. A break around midday not only serves to minimize the effective orientation errors of a purely sky-based compass system, but also spares the beetles from being active during some of the hottest hours of the day. In the field, high elevations of the sun always correspond with this midday drop in activity, and while we cannot exclude that the orientation precision of the ball-rolling beetles varies with the time of day we consider it very unlikely.
In conclusion, during the greater part of the day, the compass system of S. lamarcki offers an orientation precision of 16°. Orientation errors of less than 7° have been recorded for other diurnal dung beetle species in similar experiments [12], but whether these differences in precision can be accounted for by optical differences (i.e. divergence angle), variations in the size of the dorsal rim area used for polarized light detection, additional compass cues, small differences in the ball-rolling techniques or experimental set-ups remains to be investigated.
4. Relative importance of the sun for orientation at different elevations
(a). The effect of a mirrored sun at different sun elevations
To investigate the influence of the solar compass on straight-line orientation, we recorded the bearings of beetles rolling first with a view of the real sun and then with a view of a mirrored sun. While the sun's apparent azimuthal position is changed by 180° in this treatment, the directional information from the skylight will remain unaffected. We expect beetles that use the sun as their primary orientation cue to change their rolling direction by 180° (figure 3, grey sectors with sun should include 95% of the population). Beetles orienting preferentially to the skylight cues, however, should not change their rolling direction (figure 3, grey sectors with S). It is also possible that beetles are using a combination of the sun and skylight cues to orientate. If they weigh these cues equally, we expect them to change their bearing by around 90°, but the greater the relative impact of the sun, the larger the angular deviation between the two bearings. We found that the angular deviation between the two bearings was 128.0 ± 5.7° at high solar elevations (more than 75°), 120.6 ± 6.7° at medium solar elevations (45–60°) and 104.2 ± 7.4° at low solar elevations (less than 30°) (n = 60; figure 3). This indicates that the sun is one of the main orientation cues for straight-line orientation in S. lamarcki, but the relative influence of this cue varies for different sun elevations (ANOVA, F2/177 = 3.37, p = 0.04) and is stronger at higher elevations.
In an orientation system where the sun compass dominates over other compass systems, we expect the animals to change their direction by 180° when the position of the sun is set in conflict with their other compass systems by the use of a mirror. This type of response can be observed in shore-living sandhoppers [26] and in some ant species [27]. At high solar elevations, 77% of the dung beetles chose a direction consistent with a 180° turn. At medium and low solar elevations, 52 and 37% of the population followed the direction of the sun (figure 3a–c, grey sectors with sun). If, instead, the skylight compass has a larger influence than the sun compass, as in the desert ant Cataglyphis [28,29], the animals should not change from their original selected bearing. For S. lamarcki, only 15% (at high and medium elevations) to 25% (at low elevations) of the population followed their original bearing when the sun was set in conflict with their skylight compass (figure 3a–c, grey sectors with S). This indicates that the skylight compass rarely has a larger influence than the sun compass.
When the directions given by skylight and sun information were set in conflict, many beetles also took an intermediate course between the two (figure 3b–d, white sectors). In similar conflict experiments, dancing bees also choose an intermediate course between skylight information and sun information [30] and navigating ants steer an intermediate course between the direction of the sun and the direction of the wind [15]. In both of these insects, the relative change in the orientation direction can be controlled by the relative strengths of the two stimuli. As the sun draws closer to the horizon, the degree of polarization in the zenith of the sky increases [22]. It is thus possible that the observed decrease in the relative influence of the sun at low solar elevations is a consequence of the increased input to the skylight compass.
Finally, there is a large variability in how ball-rolling dung beetles respond to the two conflicting stimuli, skylight and sun. This could be a consequence of the direction in which a beetle initially chooses to roll [7]. For example, beetles rolling directly towards the sun will not necessarily be able to see the disc of the sun behind their ball (as they move backwards with their head down when rolling) and may accord less weight to this cue. The circular–circular correlation coefficient [31] between the initial bearing and the orientation response to the mirrored sun, however, is only 0.03. This indicates that a beetle's initial bearing does not influence its response to the mirrored sun. A similar spread in the orientation response has been described for sandhoppers, when the sun and the magnetic field are set in conflict [16]. Those authors conclude that there are ‘solar’ individuals and ‘magnetic’ individuals, and we cannot exclude that there are similar individual differences in the orientation system of dung beetles.
(b). The effect of a shaded sun on straight-line orientation
If exposed to the full sky during training, the desert ant Cataglyphis has difficulties finding its way home if its main orientation cue, the celestial polarization pattern, is removed on the homebound journey [28]. To test the orientation performance of beetles in the absence of one of their main orientation cues, the sun, the orientation errors of beetles rolling in the shade were determined using the same method as above: beetles were allowed to roll out from the centre of the circular arena twice and the difference between the two tracks was analysed (figure 1a). Again, we found that orientation errors were significantly larger at higher elevations of the shaded sun: high 25.5 ± 4.1°, medium 17.5 ± 3.5° and low 11.7 ± 2.2° (n = 30; ANOVA, F2/87 = 4.35, p = 0.02; figure 4, grey bars). Further, they do not differ significantly from those recorded when the beetles had a full view of the sky (two-way ANOVA, F1/134 = 0.56, p = 0.46). This indicates that, even if the sun is used as the primary cue for straight-line orientation, the precision of the compass system is not significantly affected by its absence. This finding supports the results from previous studies of re-orientation in ball-rolling dung beetles in the sun and in the shade after induced falls or passive rotations [12].
Figure 4.

The precision of the dung beetle celestial compass in the presence and absence of the sun. The orientation error of the dung beetle celestial compass was determined by letting beetles roll out twice from the centre to the edge of the arena under different experimental conditions: with a full view of the sky (sun–sun, white bars), with the sun hidden from view (shade–shade, grey bars) or with the sun hidden from view only during the second roll (sun–shade, white/grey bars). The absolute angular difference between the two paths determines the orientation error. The orientation error in each condition was determined for different elevations of the sun: 75–78° (high elevations), 45–60° (medium elevations) and 15–30° (low elevations). In all experimental conditions, the orientation error at high elevations of the sun is significantly larger than that at medium or low solar elevations. Numbers in text and figure show mean ± s.e.m.
While we find that the dung beetle's compass is equally precise when orienting in the sun or in the shade, we also wanted to test how robust the dung-beetle celestial compass system is to the removal of the sun after the initial bearing has been set in its presence. In this part of the experiment, the sun was shaded only during the second exit of the beetle. In line with our previous results, we find that the mean orientation error at high elevations (38 ± 7.2°, n = 15) is almost twice as large as that at medium (19.3 ± 3.3°, n = 15) or low elevations (17.3 ± 2.4°, n = 15; ANOVA, F2/42 = 5.67, p < 0.01; figure 4, white/grey bars). Even though the orientation errors in this experimental condition are consistently larger compared with repeated rolls in the sun, they do not differ significantly (two-way ANOVA, F1/179 = 3.24, p = 0.07). Thus, even if the sun serves as one of the main orientation cues, the magnitude of the orientation error of the beetle is not significantly affected by a sudden removal of this cue or by its absence. Or, in more functional terms, the orientation performance of the dung beetle's celestial compass does not vary between the shady and the sun-lit parts of its path (figure 4).
5. Conclusion
The orientation system of ball-rolling dung beetles relies exclusively on celestial information. Consequently, they experience an unavoidable loss in orientation precision in the middle of the day, when celestial cues give less precise directional information. From morning to midday (with solar elevations above 75°) the error given by the celestial compass of the beetles nearly doubles from 16° to 29°. During these hours of the day, the beetles will either have to stay underground or forage with decreased orientation precision.
In the ball-rolling dung beetles S. lamarcki, the skylight compass and the solar compass are active together and share the control of behaviour. The solar compass dominates the skylight compass at all elevations of the sun, but the relative influence of the sun in the celestial compass system is flexible and changes with sun elevation. A recent model of navigational control suggests that a guidance system that produces a sharp activation pattern will have a stronger influence on the behavioural output than a system that produces a weaker activation pattern [23]. Our observations of the different compass cues within the dung beetle compass support this idea. In the beetles, the influence of the sun decreases at lower elevations. We suggest that this is because the polarized light in the zenith of the sky creates a stronger activation pattern the closer the sun moves to the horizon.
Within the observed population of dung beetles, we find large individual differences in the relative influence of the solar compass. At all elevations of the sun, we find instances in which the solar compass dominates over the skylight compass and vice versa. Whether there are separate ‘solar individuals’, who will repeatedly follow the direction of the sun, and ‘skylight’ individuals, who follow the direction of polarized light or other celestial cues, remains to be investigated. In nature, of course, celestial orientation cues are never set in conflict with each other. Thus, the pooling of information from multiple cues would help to reduce errors in a directional decision, irrespective of the nature of the main orientation cue. Individual differences between the relative influence of different cues in the celestial compass system of beetles will thus not necessarily be reflected in orientation precision at the individual level.
While the sun is a local point-source that occupies only one ommatidium and can easily be obscured by clouds, the skylight cues span the entire sky. Despite these differences in reliability, S. lamarcki and many other ball-rolling dung beetles [12] preferentially follow the direction given by the sun. The homing dung beetle Pachysoma, which needs to navigate back to a tiny nest entrance among the sand dunes, seems to follow the direction given by its skylight compass instead [32]. This raises interesting questions regarding the evolution and precision of different types of compass systems. Is the celestial compass of Pachysoma more precise than that of ball-rolling beetles, and do they complement their celestial compass with landmark cues to increase their orientation precision? These questions will be the focus of our future studies.
Acknowledgements
We thank Ted and Winnie Harvey for their invaluable help at the field site, the Swedish Research Council, the Carl Tryggers Foundations, the Royal Physiographic Society of Lund and the South African National Research Foundation.
References
- 1.Tribe GD, Burger BV. 2011. Olfactory ecology. In Ecology and evolution of dung beetles (eds Simmons LW, Ridsdill-Smith TJ.), pp. 87–105. Oxford, UK: Wiley-Blackwell. [Google Scholar]
- 2.Dacke M, Byrne M, Smolka J, Warrant E, Baird E. 2012. Dung beetles ignore landmarks for straight-line orientation. J. Comp. Physiol. A 199, 17–23. ( 10.1007/s00359-012-0764-8) [DOI] [PubMed] [Google Scholar]
- 3.Byrne MJ, Dacke M. 2011. The visual ecology of dung beetles. In Ecology and evolution of dung beetles (eds Simmons LW, Ridsdill-Smith TJ.), pp. 177–198. Oxford, UK: Wiley-Blackwell. [Google Scholar]
- 4.Dacke M, Baird E, Byrne M, Scholtz CH, Warrant EJ. 2013. Dung beetles use the Milky Way for orientation. Curr. Biol. 23, 1–3. ( 10.1016/j.cub.2012.12.034) [DOI] [PubMed] [Google Scholar]
- 5.Dacke M, Byrne MJ, Baird E, Scholtz CH, Warrant EJ. 2011. How dim is dim? Precision of the celestial compass in moonlight and sunlight. Phil. Trans. R. Soc. B 366, 697–702. ( 10.1098/rstb.2010.0191) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dacke M, Nilsson D-E, Scholtz CH, Byrne M, Warrant EJ. 2003. Animal behaviour: insect orientation to polarized moonlight. Nature 424, 33 ( 10.1038/424033a) [DOI] [PubMed] [Google Scholar]
- 7.Baird E, Byrne MJ, Scholtz CH, Warrant EJ, Dacke M. 2010. Bearing selection in ball-rolling dung beetles: is it constant? J. Comp. Physiol. A 196, 801–806. ( 10.1007/s00359-010-0559-8) [DOI] [PubMed] [Google Scholar]
- 8.Cheung A, Zhang S, Stricker C, Srinivasan MV. 2007. Animal navigation: the difficulty of moving in a straight line. Biol. Cybern. 97, 47–61. ( 10.1007/s00422-007-0158-0) [DOI] [PubMed] [Google Scholar]
- 9.Dacke M, Nordström P, Scholtz CH, Warrant EJ. 2002. A specialized dorsal rim area for polarized light detection in the compound eye of the scarab beetle Pachysoma striatum. J. Comp. Physiol. A 188, 211–216. ( 10.1007/s00359-002-0295-9) [DOI] [PubMed] [Google Scholar]
- 10.Dacke M, Nordström P, Scholtz CH. 2003. Twilight orientation to polarised light in the crepuscular dung beetle Scarabaeus zambesianus. J. Exp. Biol. 206, 1535–1543. ( 10.1242/jeb.00289) [DOI] [PubMed] [Google Scholar]
- 11.Dacke M, Byrne MJ, Scholtz CH, Warrant EJ. 2003. Lunar orientation in a beetle. Proc. R. Soc. Lond. B 271, 361–365. ( 10.1098/rspb.2003.2594) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Byrne M, Dacke M, Nordstr mP, Scholtz C, Warrant E. 2003. Visual cues used by ball-rolling dung beetles for orientation. J. Comp. Physiol. A 189, 411–418. ( 10.1007/s00359-003-0415-1) [DOI] [PubMed] [Google Scholar]
- 13.Hegedüs RR, Akesson SS, Horváth GG. 2007. Polarization patterns of thick clouds: overcast skies have distribution of the angle of polarization similar to that of clear skies. J. Opt. Soc. Am. A 24, 2347–2356. ( 10.1364/JOSAA.24.002347) [DOI] [PubMed] [Google Scholar]
- 14.Pomozi II, Horváth GG, Wehner RR. 2001. How the clear-sky angle of polarization pattern continues underneath clouds: full-sky measurements and implications for animal orientation. J. Exp. Biol. 204, 2933–2942. [DOI] [PubMed] [Google Scholar]
- 15.Müller M, Wehner R. 2007. Wind and sky as compass cues in desert ant navigation. Naturwissenschaften 94, 589–594. ( 10.1007/s00114-007-0232-4) [DOI] [PubMed] [Google Scholar]
- 16.Ugolini A. 2001. Relationship between compass systems of orientation in equatorial sandhoppers. Anim. Behav. 62, 193–199. ( 10.1006/anbe.2001.1766) [DOI] [Google Scholar]
- 17.Ugolini A. 2002. The orientation of equatorial sandhoppers during the zenithal culmination of the sun. Ethol. Ecol. Evol. 14, 269–273. ( 10.1080/08927014.2002.9522745) [DOI] [Google Scholar]
- 18.Baird E, Byrne MJ, Smolka J, Warrant EJ, Dacke M. 2012. The dung beetle dance: an orientation behaviour? PLoS ONE 7, e30211 ( 10.1371/journal.pone.0030211) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ugolini A, Galanti G, Mercatelli L. 2009. Difference in skylight intensity is a new celestial cue for sandhopper orientation (Amphipoda, Talitridae). Anim. Behav. 77, 171–175. ( 10.1016/j.anbehav.2008.09.035) [DOI] [Google Scholar]
- 20.Rossel S, Wehner R. 1984. Celestial orientation in bees: the use of spectral cues. J. Comp. Physiol. A 155, 605–613. ( 10.1007/BF00610846) [DOI] [Google Scholar]
- 21.Edrich W, Neumeyer C, von Heiversen O. 1979. ‘Anti-sun orientation’ of bees with regard to a field of ultraviolet light. J. Comp. Physiol. A 134, 151–157. ( 10.1007/BF00610473) [DOI] [Google Scholar]
- 22.Brines ML, Gould JL. 1982. Skylight polarization patterns and animal orientation. J. Exp. Biol. 96, 69–91. [Google Scholar]
- 23.Collett M. 2012. How navigational guidance systems are combined in a desert ant. Curr. Biol. 22, 927–932. ( 10.1016/j.cub.2012.03.049) [DOI] [PubMed] [Google Scholar]
- 24.Smolka J, Baird E, Byrne MJ, el Jundi B, Warrant EJ, Dacke M. 2012. Dung beetles use their dung ball as a mobile thermal refuge. Curr. Biol. 22, R863–R864. ( 10.1016/j.cub.2012.08.057) [DOI] [PubMed] [Google Scholar]
- 25.Hanski I, Cambefort Y. 1991. Dung beetle ecology. Princeton, NJ: Princeton University Press. [Google Scholar]
- 26.Pardi L, Ercolini A. 1986. Zonal recovery mechanisms in talitrid crustaceans. B. Zool. 53, 139–160. ( 10.1080/11250008609355499) [DOI] [Google Scholar]
- 27.Santschi F. 1911. Observations et remarques critiques sur le mécanisme de l'orientation chez les fourmis. Rev. Suisse. Zool. 19, 305–338. [Google Scholar]
- 28.Wehner R, Müller M. 2006. The significance of direct sunlight and polarized skylight in the ant's celestial system of navigation. Proc. Natl Acad. Sci. USA 103, 12 575–12 579. ( 10.1073/pnas.0604430103) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Duelli P, Wehner R. 1973. The spectral sensitivity of polarized light orientation in Cataglyphis bicolor (Formicidae, Hymenoptera). J. Comp. Physiol. A 86, 37–53. ( 10.1007/BF00694476) [DOI] [Google Scholar]
- 30.von Frisch K. 1967. The dance language and orientation of bees. London, UK: Oxford University Press. [Google Scholar]
- 31.Batschelet E. 1981. Circular statistics in biology. London, UK: Academic Press. [Google Scholar]
- 32.Dacke M, Nordström P, Scholtz C. 1999. Two sets of compound eyes in the dung beetles Pachysoma: a specialistaion for skylight navigation. In Proc. 1st Göttingen Conf. of the German Neuroscience Society Stuttgart, Germany: Georg Thiem Verlag. [Google Scholar]