

Abstract
During thymic development, thymocytes expressing a T cell receptor consisting of an alpha and beta chain (TCRαβ), commit to either the cytotoxic- or T helper-lineage fate. This lineage dichotomy is controlled by key transcription factors, including the T helper (Th) lineage master regulator, the Th-inducing BTB/POZ domain-containing Kruppel-like zinc-finger transcription factor, ThPOK, (formally cKrox or Zfp67; encoded by Zbtb7b), which suppresses the cytolytic program in major histocompatibility complex (MHC) class II–restricted CD4+ thymocytes and the Runt related transcription factor 3 (Runx3), which counteracts ThPOK in MHC class I restricted precursor cells and promotes the lineage commitment of CD8αβ+ cytolytic T lymphocytes (CTL). ThPOK continues to repress the CTL gene program in mature CD4+ T cells, even as they differentiate into effector Th cell subsets. The Th cell fate however is not fixed and two recent studies showed that mature, antigen-stimulated CD4+ T cells have the flexibility to terminate the expression of ThPOK and functionally reprogram to cytotoxic effector cells. This unexpected plasticity of CD4 T cells results in the post-thymic termination of the Th lineage fate and the functional differentiation of distinct MHC class II–restricted CD4+ CTL. The recognition of CD4 CTL as a defined separate subset of effector cells and the identification of the mechanisms and factors that drive their reprogramming finally create new opportunities to explore the physiological relevance of these effector cells in vivo and to determine their pivotal roles in both, protective immunity as well as in immune-related pathology.
Keywords: CD4 T helper cells, CD8αβ Cytotoxic T lymphocyte, ThPOK, Runx3, plasticity, lineage commitment, functional reprogramming, tumor, chronic viral infection, intestine, autoimmunity, protective immunity
1. Introduction
MHC class II restricted CD4+ TCRαβ+ cells have commonly been classified as ‘helper’ T cells based on their function to help promote or dampen cellular or humoral immune responses whereas MHC class I restricted CD8αβ+ T lymphocytes are known to provide direct protection by killing infected or transformed cells. Nevertheless, numerous observations suggest that CD4 T cells may also play direct and indispensable roles in protective immunity. The functional dichotomy of TCRαβ T cells is initially induced during thymic development and coincides with, but does not depend on, the specific MHC restriction and CD4- or CD8αβ co-receptor expression. The molecular mechanism, which controls the lineage-commitment is driven by the action and counteraction of key transcription factors such as the Th cell transcription factor, ThPOK, which drives the CD4+ Th cell fate and prevents thymocytes from differentiating into CTL [1–7]. Runx3, a member of the Runx family of transcription factors, has the opposite effect and terminates CD4 expression while promoting differentiation of MHC class I restricted thymocytes into CD8αβ CTL [8, 9]. ThPOK, continues to be expressed in naïve CD4 T cells and cooperates with additional transcription factors, such as T-bet, Gata3, RORγt or Foxp3, as activated mature CD4 T cells differentiate into Th1, Th2, Th17 or T regulatory cells respectively [4, 7].
Nevertheless, cytotoxicity associated with antigen-experienced CD4+ lymphocytes has been described for decades especially under conditions of chronic viral infections or viral-induced immune pathology and autoimmunity [10, 11]. At first cytolytic CD4+ T cells were considered a potential artifact of in-vitro generated T cell lines and clones but more recent reports provided unambiguous evidence that in-vivo some antigen-specific CD4+ T cells possess direct MHC class II restricted cytotoxic activity [12–18]. These cells were merely viewed however as functional variants of the classical CD4 Th1 cells and consequently they remained unexplored and their physiological relevance was often doubted. Because of this, cytolytic CD4 effector cells have not been fully appreciated nor recognized as possible active contributors in health and disease.
Recently two studies [4, 19], determined that cytotoxic CD4+ T cells form a separate type of CD4 effector cells that is distinct from any known conventional CD4+ Th subset. They showed that, like classical CD8αβ CTL, these mature CD4+ T cells lack expression of the Th master regulator, ThPOK. However, in contrast to the thymic committed CD8αβ CTL, termination of the Thpok gene expression in the cytotoxic CD4+ T cells occurs post-thymically in response to repeated stimulation with their cognate antigen in-vivo [4]. As a consequence of the loss of ThPOK expression, activated CD4+ T cells de-repress the cytolytic-gene expression program leading to the functionally effector differentiation of MHC class II restricted CD4 CTL. The post-thymic reprogramming of mature CD4+ T cells provides a unique mechanism of plasticity not only to generate cytotoxic MHC class II restricted effector T cells but also to redirect Th cells away from becoming either inflammatory- or immunosuppressive cells.
The discovery of the CTL reprogramming of mature CD4+ T cells not only represents a major advance in our understanding of T cell biology but also provides powerful opportunities for the design of new strategies to overcome inflammatory T cell-mediated pathologies or immune suppression as well as to induce pre-existing anti-viral or anti-tumor protective immunity. Together with the observation that defects in the differentiation or regulation of this process may lead to impaired immune protection or aberrant immune functions [19], these significant novel insights have evoked new interest in the cytotoxic CD4+ T cells as potential important beneficial and/or pathogenic contributors of the immune response.
2. Thymic commitment and lineage decision
The thymus is initially seeded by bone marrow-derived uncommitted progenitors, which gradually lose their multipotency and fully commit to the T cell lineage. The initial process involves suppression of gene expression programs characteristic of other lineages as well as the induction of a T cell specific gene expression profile mediated by various transcription factors, including Runx1, Gata3 and E-box proteins, which cooperate with Notch1 to initiate T-lineage differentiation [20]. Immature thymocytes first appear as CD4 and CD8αβ coreceptor double negative (DN) cells that sequentially transition through various stages defined by the expression of CD44 and CD25 as CD44+CD25− DN1, CD44+CD25+ DN2, CD44−CD25+ DN3 and finally CD44−CD25− DN4 cells. During the immature DN stages the T lineage committed progenitors rearrange the loci encoding the TCR and pass through various checkpoints to verify proper rearrangement of the γ and δ or α and β TCR chains. Before expressing a full αβTCR, precursor thymocytes first express a pre-TCR consisting of the CD3 components together with a variable rearranged β-chain and an invariant pre-α chain. Signals received through the pre-TCR, known as β-selection, direct αβTCR progenitors to the next phase and those that pass β-selection undergo a proliferative burst and become CD4 CD8αβ double positive (DP) or triple positive (TP) thymocytes that express CD8αα together with the coreceptors CD8αβ and CD4 [21]. During this phase, the T cell progenitors also rearrange the Tcrα gene, resulting in the surface expression of mature αβTCR complexes and the full commitment of the thymocyte to the TCRαβ lineage. Those thymocytes, which successfully rearrange an αβTCR progress further through a selection process based on the interaction of their TCR with self-MHC and self-antigens. Selected thymocytes segregate into mature DN or ‘single-positive’ (SP) cells that either express the CD8αβ coreceptor together with an MHC class I restricted TCR or the CD4 coreceptor in conjunction with an MHC class II specific TCR. The selection event also coincides with, but does not depend on, the functional commitment of the developing thymocytes, to either the CD8αβ CTL-lineage or the CD4 Th-lineage. The functional fate decision is mediated by key transcription factors, including the master Th transcription factor, ThPOK, which promotes the CD4+ T helper fate and prevents thymocytes from differentiating into CTL and Runx3, which has the opposite effect and terminates CD4 expression while driving differentiation of the maturing thymocyte into the CD8αβ CTL lineage [20] (Fig. 1).
Figure 1. Th and CTL Lineage-commitment in the thymus.

The reciprocal action and counter action of the master transcription factors, ThPOK and Runx3, drive the Th-lineage- and CTL-lineage commitment of MHC class II and MHC class I restricted thymocytes, respectively.
3. Lineage-fate decisions and transcription factors
Although much attention has been focused on ThPOK as the master transcription factor of the CD4 Th lineage fate, another important regulator of early T cell development, β-selection and CD4 Th lineage differentiation is the zinc finger transcription factor GATA-binding protein 3 (Gata3) [3, 5, 22]. Gata3 is expressed throughout T cell development and asymmetrically distributed during thymic selection with increased levels in CD4 Th-lineage differentiating thymocytes [23], but reduced levels in CD8αβ CTL committed progenitors [3, 24]. The increase in Gata3 expression also occurs in Thpok-deficient, MHC-II-restricted precursor cells whereas in contrast, ThPOK is not detected in Gata3-deficient thymocytes, indicating that Gata3 functions upstream of ThPOK as a critical inducer of Thpok in early T cell development. Since, enforced Thpok transgene expression in Gata3-deficient thymocytes does not lead to CD4-Th lineage commitment [25], it also suggests that Gata3 serves additional roles to promote the ThPOK-driven CD4 Th fate. Importantly, Gata3 also represses CD8-lineage gene- expression, including Runx3. Consequently, high-level of Gata3 and Runx3 are mutually exclusive, making Gata3 a critical regulator of the CD4-CD8 lineage commitment. Interestingly, even though Gata3 cooperates with ThPOK to promote the CD4 Th-fate, Gata3 is not required for the ThPOK-mediated suppression of Runx3, and transgenic expression of ThPOK alone is sufficient to block the CD8 differentiation of Gata3-deficient thymocytes [25]. Furthermore, enforced Thpok expression causes precursor cells, including MHC class I–restricted thymocytes, to develop into CD4 Th cells [6, 25]. Similarly, in mice expressing ThPOK constitutively due to Runx deficiency, class I–restricted CD4-expressing lymphocytes develop, which exhibit phenotypic and functional characteristics of CD4+ Th cells [26, 27]. Conversely, in mutant mice, which fail to express functional ThPOK (ThPOKHD/HD, a mutant form of Thpok, found in CD4 Th-deficient mice), MHC class II–restricted CD8 T cells develop, which exhibit a functional CTL phenotype [2], whereas the Cd4 silencer in these cells is derepressed [28]. These findings demonstrate that ThPOK is necessary and sufficient to suppress the CD8αβ lineage fate in developing thymocytes. The re-expression of CD8 in redirected CD4 Th- cells is driven by Runx3; which is an essential transcription factor to drive expression of CD8αβ CTL-lineage associated genes like perforin and granzyme b the cytokine ifn-γ and the transcription factor eomesodermin (eomes) [7, 29]. Runx3 protein is not detected in DP thymocytes, but it is up-regulated during the differentiation of CD8 CTL-lineage cells in the thymus and remains expressed in post-thymic mature CD8αβ+ T cells [9, 30, 31] (Fig. 1); Although Runx3 deletion does not prevent CD8 CTL differentiation, it results in reduced numbers of CD8αβ+ T cells, and most of them appear as CD4CD8αβ DP cells. Furthermore, Runx3-deficient CD8 cells have reduced CD8 expression [30], consistent with the direct binding of Runx3 to the Cd8 enhancer, E8I, located in the Cd8α gene control region [31]. The presence of CD4 Th lineage cells in Thpok-Runx double-deficient mice, but not in ThPOK single-deficient mice, suggests that Runx not only directs the CTL lineage but also inhibits the expression of some Th genes that direct Runx-deficient thymocytes toward the CD4 Th lineage. In line with this, Runx3 is able to bind a silencer located upstream of the distal promoter of the Thpok gene, which results in active repression of ThPOK in immature CD8αβ+ thymocytes [9, 30–32]. This negative feed back loop system, where ThPOK represses Runx3 during the Th lineage commitment and Runx represses ThPOK in CTL-lineage thymocytes, prevents coexpression of both transcription factors in committed thymocytes (Fig. 1 and 2). Runx3 does not require MHC-I-specific signals for its initial induction in thymocytes since it is expressed in MHC-II restricted thymocytes that are Thpok-deficient [33]. On the other hand, Thpok is kept silent in MHC-I-restricted thymocytes before detectable Runx3 up-regulation, whereas enforced expression of Runx3 does not redirect MHC-II-restricted thymocytes into the CD8+ CTL-lineage [34, 35]. These observations indicate that the mutual exclusive regulation of ThPOK and Runx3 is not the only mechanism that drives the lineage choice in developing thymocytes and that additional factors are involved to direct the reciprocal commitment of CD4+ and CD8αβ+ progenitor cells [26]. Such factors may include Mazr, a BTB/POZ domain-containing transcription factor, which also controls thymocyte fate [36] by repressing ThPOK expression in MHC class-I signaled DP thymocytes or NF-κB, and E-box binding proteins, as well as genes involved in the Notch pathway, all of which have been proposed to be involved, positively or negatively, in the CD4/CD8 lineage fate [37–41].
Figure 2. ThPOK is the master regulator of the CD4 Th-lineage fate.
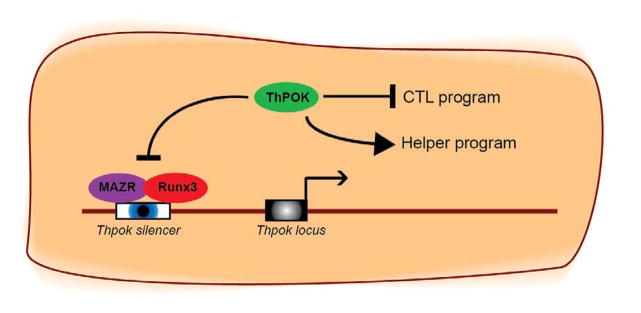
ThPOK drives the Th fate in developing thymocytes by counteracting Runx3. ThPOK continues to suppress the Runx3-controlled cytotoxic program in mature MHC class II restricted CD4 T cells. ThPOK controls its own expression by repressing the Thpok silencer in mature CD4 T cells. Derepression of the Thpok silencer results in the termination of Thpok transcription and loss of ThPOK expression and the activation of the Runx3-controlled CTL gene expression program.
4. Transcriptional regulation of mature CD4+ Th cells in the periphery
Although the mutual exclusive expression of the lineage master transcription factors is strictly regulated in the thymus, the control of these and other factors to maintain the lineage integrity—and prevent derepression of alternative gene programs in mature T cells is more flexible. For example, in activated CD4+ T cells, Runx3, which counteracts ThPOK in the thymus, is induced by T-bet and actively promotes the IFN-γ production by Th1 cells [42, 43]. On the other hand, conditional disruption of ThPOK expression post thymically in CD4+ T cells causes CD8 re-expression and the induction of granzyme B [7], whereas enforced expression of ThPOK in mature T cells results in lineage conversion of MHC class I-restricted T lymphocytes toward the CD4 fate [6]. These observations indicate that ThPOK expression, initially established in the thymus, must be maintained in the periphery in order to prevent the activation-induced Runx3-driven CTL program in CD4+ effector cells [7, 44]. In response to antigens, CD4+ T cells differentiate to various effector subtypes, controlled by master transcription regulators that direct the functional specialization of each specific Th subtype, including T-bet for Th1, Gata3 for Th2 cells and RORγt for Th17 (Fig.3). In general, primary transcription factors will induce a secondary set, such as Gata3-induced ThPOK or T-bet-induced Runx3, after which they cooperate to drive specific gene expression profiles like the Gata-3/ThPOK-driven Th program in thymocytes or the T-bet/Runx3 Th1 program in mature CD4+ T cells. Gata3 is no longer expressed in naïve or resting mature CD4 T cells, however it is auto-induced upon activation [45], suggesting that Gata3 might have additional roles for the functional differentiation of CD4 effector cells. Consistent with this notion, conditional deletion of Gata3 abolishes Th2 differentiation [46] and leads to the instability of Foxp3 Tregs during inflammation [47, 48]. In contrast, enforced Gata3 induced co-expression of Th2 cytokines in Th17 conditioned cells [49] and under Th1 conditions, Gata3 is redistributed to T-bet-binding sites at Th1 genes away from Th2 genes [50]. Hence, in activated CD4+ T cells, Gata3 functions as an inducer or repressor of functional gene expression programs in various CD4 Th subsets. However, even though Gata3 initially induces ThPOK expression in developing thymocytes, it is no longer required for the constitutive expression and maintenance of ThPOK in naïve CD4 T cells [25]. Instead ThPOK regulates its own expression by repressing a silencer in its promoter thus counteracting Runx3-driven transcriptional suppression, which results in continuous expression of ThPOK in mature CD4 Th-lineage T cells (Fig. 2). Activated CD4 T cells however, lacking Thpok, in the presence of normal Runx3 activity still undergo multiple Th effector differentiation, except Th2 polarization, which is inhibited by the re-expression of Runx3. This observation indicates that Runx3 repression by ThPOK is not required for the Th differentiation per se; but rather, it is necessary to suppress Runx3 and preserve specific CD4 Th effector fates. Moreover, a number of studies showed that ThPOK not only counteracts Runx3 but also negatively regulates the expression of other CD8 CTL-lineage genes including Cd8α, perforin, granzyme B [44]. In addition, eomes, which promotes IFN-γ expression and cytotoxic effector differentiation of mature CD8αβ cells, is directly repressed by ThPOK [51–53]. These observations indicate that ThPOK preserves the Th fate by negatively regulating the CD8αβ CTL transcription program in activated mature CD4 T cells. Another transcription factor LRF is also required for the maintenance of the Th program in mature CD4 T cells, however in contrast to the Thpok deletion, disruption of both factors, ThPOK and LRF, resulted in a complete block of Th gene expression and Th cell polarization of MHC class II restricted mature T cells in-vivo. These data identified the ThPOK-LRF pair as master regulators of Th differentiation and function [54].
Figure 3. Functional plasticity of mature CD4 T cells.

Antigen-stimulated CD4 T cells functionally differentiate into various Th subsets, driven by subset-specific transcription factors and cytokines. Repeated antigen stimulation leads to the loss of ThPOK expression and the induction of the Runx3-driven CTL polarization.
5. Diverting from the T helper path: CD4 CTL
Although, it is evident that ThPOK functions to continuously suppress the CTL program in mature CD4 Th lymphocytes, many reports have repeatedly indicated the presence in-vivo of CD4+ T cells with cytolytic features [55–57]. For example, at steady state, effector cells within the epithelium of the intestine show an enrichment for cytotoxic T cells, including CD4+ T cells [4, 19] and small numbers of cytolytic CD4+ T cells can be detected in the blood of healthy individuals [57]. In immune challenges, including viral infections [58], antitumor responses [59, 60] and autoimmune disorders [61, 62], cytotoxic CD4+ T cells with a functional phenotype typical of end-stage differentiated effector cells, significantly expand in the blood and peripheral tissues [57, 63]. The presence of CD4+ cytotoxic T cells in various species, including humans and rodents and their expansion in certain disease conditions, suggest that they might participate in protective and/or pathogenic immune functions. Nonetheless, direct evidence for unique non-redundant roles for these cells in any aspect of the adaptive immune response has not been generated, mainly because cytolytic CD4+ effector cells have usually been viewed and treated as functional variants of the conventional CD4 Th1 subtype. As a result, CD4+ CTL have not been actively targeted as part of clinical strategies to fight or prevent infections or other immune related diseases. New advances, however, indicating that CD4 T cells with cytolytic capacity are separate from any CD4 Th subset [4, 19], together with the fact that the efficiency of MHC-class II effector cells to kill target cells in-vitro matches that of classical CD8αβ CTL, underscores the notion that direct protective immunity might not only be the valuable characteristic of CD8αβ MHC class I, but also of CD4 MHC class II restricted TCRαβ+ T cells.
6. Functional reprogramming of mature CD4 Th cells to CTL
The functional features expressed by cytolytic CD4 T cells suggest that ThPOK does not suppress the Runx3-driven CTL gene expression program in those cells. In agreement with this, Mucida et al. (2013) and Reis et al. (2013), provided the first evidence that activated CD4+ T cells have the capacity to lose ThPOK and gain the Runx3-controlled CTL gene-expression program. Using an adoptive transfer approach these new reports showed that naïve CD4+ T cells isolated from Thpok-GFP reporter mice transferred into lymphopenic (Rag1−/−) recipient mice, lost GFP expression indicating that these CD4 donor cells no longer expressed ThPOK. In parallel with the loss of ThPOK, the transferred cells acquired various CTL features, including the expression of granzymes, perforin, and IFN-γ, together with the cell surface induction of several receptors typical of natural killer cells and conventional CD8αβ CTL. This included the cytotoxicity-related, MHC class I–restricted, T cell–associated molecule CRTAM and the CD2 family member CD244 (2B4), both known to promote the cytolytic function and IFN-γ production of CD8αβ effector cells [64–66]. In contrast, in this transfer model of induced colitis, genes characteristic of Th17 cells, including those encoding the cytokines IL-17A, IL-17F and IL-22, the cytokine receptor IL-23R and the transcription factor RORγt, were expressed by many of the GFP+ThPOK+ pathogenic CD4 donor cells but not by the GFP− subset. Moreover, analysis of genes characteristic of other CD4 Th types, such as Th1 and Th2 cells, demonstrated that the gene-expression profile of those ThPOK− CD4 donor cells was different than that expressed by typical ThPOK+ CD4 Th effector subsets. The reciprocal expression of either ThPOK or the CTL signature genes in the transferred Thpok-GFP CD4 donor cells indicated that the generation of CD4 CTL coincides with post-thymic loss of ThPOK and also confirmed the notion that ThPOK continuously suppressed the CTL program in conventional Th cell. Furthermore, enforced expression via retroviral transduction of either wild type or a mutant form of Thpok (ThPOKHD/HD) showed that this post thymic loss of ThPOK resulted in the derepression of the CTL program in mature CD4 effector cells. Using a genomic deletion approach, Mucida et al. (2013), identified the Thpok silencer as the critical switch that turned off Thpok transcription in mature CD4+ T cells. This was also confirmed in an adoptive transfer experiment using CD4 donor cells, in which a loxP-flanked Thpok silencer (ThpokSfl/Sfl) was deleted at the mature stage. CD4 donor cells with a Thpok silencer-deletion failed to differentiate to CTL upon transfer, which further emphasized the critical role of the Thpok silencer in terminating the transcription of Thpok as part of the CTL-differentiation process. In contrast to ThPOK, which suppresses its own silencer, the transcription factor MAZR activates the Thpok silencer [67], resulting in negative regulation of Thpok transcription (Fig. 2). Accordingly, MAZR-deficient CD4+ donor cells transferred into Rag1−/−recipient mice showed impaired capacity to reprogram to CTL, indicating that transcriptional termination of Thpok expression was critical for the functional differentiation of MHC class II–restricted CD4+ CTL.
Nevertheless, although the loss of ThPOK expression is a key feature in diverting mature CD4 T cells to CTL, it is not sufficient and Reis et al. [19] showed using transfer of CD4 donor cells isolated from Thpok-GFP and Runx3-YFP double reporter mice, that both, the loss of ThPOK and the gain of Runx3 expression, were required for the reprogramming of CD4+ T cells to CTL. Interestingly, whereas in this transfer model of induced colitis, forced loss of ThPOK expression resulted in dampening of the inflammation, although it did not impair the Th17 differentiation, the lack of Runx3 expression due to a Runx3 deletion resulted in severe pathology with enhanced Th17 differentiation. These observations indicated that the Runx3-driven differentiation of mature CD4+ Th cells to CTL diverts them away from becoming pathogenic effector cells. Some CD4+ T cells were Runx3hiThPOKhi, however, none were Runx3loThPOKlo, suggesting that the induction of Runx3 occurred before the full downregulation of ThPOK. Furthermore, conditional deletion of Runx3 in T cells indicated that the activation-induced Runx3 expression was critical for the generation of the ThPOK-negative CD4 CTL in-vivo. The gain of Runx3 expression also coincided with the appearance of the transcription factor T-bet, which is known to promote Runx3 expression in Th1 conditioned CD4 Th cells [19]. Nevertheless, CTL programming is not exclusively driven by T-bet-induced Runx3 expression and other CTL transcription factors, such as Eomes, may also be involved in the development of cytotoxic CD4+ T cells in vivo. Inline with this, since ThPOK directly suppresses eomes in CD4+ Th cells, it is possible that in CTL precursor cells, which lose ThPOK, Eomes becomes expressed and operates either alone or in synergy with Runx3 to drive the functional reprogramming of fully functional CD4 CTL.
7. The CD4 CTL differentiation coincides with chronic activation
Th1-polarized, CD4+ effector cells can acquire perforin-mediated CTL activity in-vitro [68] and cytolytic CD4+ cells have been isolated ex-vivo, in response to acute influenza virus infections [11], however, the typical appearance of CD4 CTL in-vivo associates most often with chronic activation such as in chronic viral infections, including cytomegalovirus (CMV) [55], Epstein–Barr virus (EBV) [56] or Human Immune deficiency Virus (HIV) infections [57] or in autoimmune conditions such as rheumatoid arthritis [62]. This suggests that persistent antigen exposure may drive the progressive differentiation of CD4 CTL during which they lose the expression of Thpok and the typical Th program and derepress the genes encoding the functional cytotoxic phenotype. In line with this, Mucida et al. [4] showed that in mice, transgenic for a TCR (OT-II) specific for ovalbumin amino acids 323–339 [OVA (323–339)] presented by I-Ab or in Rag1−/−recipients that previously received naïve OT-II CD4 donor T cells, that continuous feeding of an OVA-containing diet, resulted in the accumulation of a large number of OT-II CD4 cells in the intestine of these mice that expressed typical features of CTL. Furthermore, many OT-II Thpok-GFP cells isolated from reporter mice and transferred into OVA-fed Rag1−/−recipient mice lost GFP expression indicating that they no longer expressed ThPOK. Instead they showed expression of typical cytolytic markers, such as 2B4 and granzyme B and they displayed an OVA-specific cytolytic response when restimulated in-vitro with the OT-II TCR–specific peptide OVA (323–339) but not when restimulated with the MHC class I–restricted OVA peptide SIINFEKL (amino acids 257–264) [4]. Interestingly, despite their cytolytic potential, OT-II ThPOK CD4+ effector cells remained immunologically quiescent in-vivo even in the continuous presence of OVA in the diet. However, similar to classical CD8αβ CTLs, which upon recall with their cognate antigen in the context of excess amounts of IL-15 as in active celiac disease [69, 70] differentiate into destructive killer cells [71] that produced large amounts of the inflammatory cytokines IFN-γ and tumor-necrosis factor (TNF) [69, 72, 73], chronic OVA diet-induced OT-II CD4 CTL restimulated in-vitro with their cognate antigen (Ag) and IL-15, also considerably upregulated their protective capacity, including a substantial increase of their cytolytic and cytokine response. Similar results were obtained using polyclonal CD4+ T cells isolated from normal lymphocyte-sufficient mice, which showed that natural in-vivo generated CD4 CTL also had the potential to substantially increase their cytolytic and inflammatory immune responses when restimulated with their cognate Ag in the presence of an inflammatory challenge. The ability to generate diet-Ag-induced CD4 CTL that accumulate within the intestinal epithelium and have the potential to display potent protective immune functions when challenged with their cognate diet Ag in the context of pathogen-induced danger signals, provides the mucosal immune system with a powerful arm to prevent invasion of newly encounter pathogens without pre-existing pathogen-specific immunity. Repeated Ag stimulation via the diet is likely not the only condition in-vivo that drives the reprogramming of CD4 Th cells to CTL, since cytotoxic CD4 T cells have frequently been observed in chronic viral infections, anti-tumor responses as well as in various autoimmune conditions. Therefore, it is possible that the repeated stimulation with viral-, tumor- or self-antigens in the absence of excess inflammatory cytokines also leads to the loss of ThPOK and the functional differentiation of CD4 to CTL.
Overall, these new insights demonstrate that thymic committed CD4 Th cells have the flexibility, when chronically stimulated by their cognate Ag in the absence of inflammatory signals, to gradually differentiate into MHC class II restricted CTL that, although at steady state remain quiescent, have the potential to exert potent effector functions when reactivated with their Ag in the presence of inflammatory conditions.
8. CD4 CTL in health and disease
The functional differentiation of CD4 Th cells to CTL suggests that they have the capacity to participate in direct protective immunity to control infections or eliminate transformed cells. Especially the ability of cytolytic CD4 effector cells to function in an MHC class II restricted fashion, renders them potentially capable of detaining viral infections trophic for class II+ target cells. Furthermore, the gradual appearance of cytolytic CD4 effector cells in close association with persistent viral infections, when the CD8 response is weakened, inadequate or totally absent, suggests that CD4 CTL might be key immune cells for the control of chronic infections or acute secondary infections. Many viruses establish persistent infections, which are mostly innocuous [74]. However, in immune compromised individuals, they often reactivate and cause lethal diseases, such as the reactivation of CMV, a leading cause of mortality in AIDS patients and transplant recipients [75]. Although it is known that the cause of viral reactivation is due to suppression and/or deletion of protective immune cells, the specific type(s) of cell(s), which keep these persistent viruses under control in immune competent conditions, has not been identified. Nevertheless, even though the precise function of CD4 CTL in the maintenance of immunity against pathogens still remains to be established, the knowledge we have gained so far from in-vitro and in-vivo studies suggests strongly that they play non-redundant roles in controlling chronic viral infections. Consistent with this, studies with lymphocytic choriomeningitis virus (LCMV) in mice, which have led to profound insights that directly apply to viral infections in humans [76], have shown that in a chronic infection with LCMV clone 13, the time preceding viral clearance is associated with exhaustion of the CD8 CTL and an expansion of CD4+ T cells that are indispensible for controlling the chronic infection [77]. Similarly, CD4+ T cells also play a principal role in controlling persistent MCMV infections in the salivary gland in mice [78], and depletion of CD4, but not CD8 T cells, in immune competent animals significantly increased the accumulation of persistent MCMV [79]. Early in infection, MCMV-specific CD4 T cells display a typical Th1 phenotype (IFNγ+/IL2+), whereas during latency, MCMV-specific cytolytic CD4 T cells (IFNγ+/IL2−) emerge suggesting that CD4 CTL might be key to control persistent CMV infections [80, 81]. In agreement with this for example, the predominant specificity of human cytolytic CD4 T cells is for the CMV structural proteins, gB and gH [82, 83], which can be processed by endosomal proteases and loaded onto MHC class II molecules [84]. Furthermore, CD4+CD28−T cells isolated from the blood of individuals infected with CMV display direct antigen-specific cytotoxicity [63] and when analyzed ex-vivo, such cells were able to kill CMV-peptide pulsed autologous target cells in an MHC class II-restricted and pathogen-dependent manner without the need for prior in-vitro stimulation [55, 81, 84]. Similarly, CD4+ T cell clones derived from patients infected with EBV, were also shown to directly lyse EBV infected B cells, or transformed B cells expressing EBV proteins [56]. Furthermore, human cytolytic CD4+ T cell clones with specificity for CMV, EBV, Polyomavirus (BKV), vaccinia virus, HIV and mycobacteria and cryptococcus neoformans [56, 85–90] have been generated in-vitro and in some instances, cytolytic CD4+ T cells generated in culture, have been adoptively transferred to patients as part of an anti-tumor therapy [91, 92] or to control infections in stem cell recipients infected with EBV or CMV [93]. Also in mice, transfer of influenza-specific, in-vitro generated CD4 CTL resulted in a reduced viral titer [94], even in the absence of CD8 or B cells, or before the primary response in normal mice could develop. This suggests that CD4 CTL might have active non-redundant roles in direct protective immunity. Perhaps more importantly, their MHC class II restriction could also be of particular importance in the control of pathogens such as the herpes viruses, EBV, CMV and especially HIV-1, that have developed sophisticated evasion strategies to avoid recognition by MHC class I restricted CD8αβ + T cells or NK cells. On the other hand, the fact that CMV has developed mechanisms to avoid the MHC class II pathway [95] could equally reflect the importance and active involvement of cytotoxic CD4+ T cells in anti-viral protective immunity.
One problem in understanding or appreciating the significance of CD4 CTL-mediated protection is the restricted MHC class II expression on target cells. Some viruses, such as EBV and LCMV can infect immune cells that constitutively express MHC class II on their cell surface [11]. However, many other viruses, such as CMV or influenza, target cells that normally do not express MHC class II molecules. Nevertheless, several published studies have shown that under inflammatory conditions, MHC class II can be drastically upregulated on various cell types including, β-cells [96], or on infected or IFNγ treated lung epithelial cells [97–99]. Also in humans, local upregulation of HLA MHC class II in various tissues seems to be a common consequence of inflammation [100, 101]. CD4 CTL might also function as bystanders and destroy infected MHC class II negative cells or other target cells indirectly through excessive production of IFNγ and TNF [102].
Recently, much attention has been paid on cytotoxic CD4+ T cells for tumor vaccination therapies as well. Indeed, not only infected but also malignant cells often induce MHC class II expression thus becoming CD4+ CTL target cells and strategies using anti-tumor specific CD4+ CTL are now beginning to be exploited. Studies of mouse γ -herpes virus 68 (MHV-68) have shown that CD4+ T cells can control herpes virus-induced malignancies in-vivo, independently of CD8+ T cells and B cells [103, 104]. Also in human, the growth inhibition of EBV-transformed B cells in healthy EBV carriers, was shown to be controlled in part by EBV-specific CD4+ T cells [105] and cytolitic CD4+ T cells, with specificity for various EBV epitopes expressed in the latent or the lytic phase of infection, have been isolated from PBMC of healthy donors [106]. Interestingly, EBV-specific CD4+ CTL have also been shown to directly target Burkitt’s lymphoma, an EBV-associated B cell lymphoma, which resists recognition by CD8αβ+ CTL due to down-regulation of the MHC class I antigen processing pathway [107]. In addition to MHC class II-based epitope vaccinations, also cell therapy approaches have been met with some success, including the treatment of EBV-associated Hodgkin’s lymphoma and nasopharyngeal carcinoma using in-vitro-generated EBV-specific CD4 CTL. In some cases, complete remission was reached for a sustained period [56, 108], supporting a significant potent and long-lasting role for anti-tumor CD4 CTL in-vivo.
In addition to the potential direct participation of CD4 CTL, as direct protective immune cells to control infections and eliminate malignant cells, CD4 CTL themselves or the mechanisms that drive their differentiation, might also have regulatory capacity. Like conventional CD8 CTL, CD4 T cells with cytolytic functions, have the potential to modulate or control immune responses by eliminating MHC class II expressing professional antigen presenting cells, thus curtailing antigen presentation during priming and preventing further activation of naive T cells. Furthermore, the accumulation of CD4 CTL in the small intestine of recipient mice in the transfer model of induced colitis, correlated with less tissue destruction, suggesting that the CTL reprogramming resulted in less pathogenic effector cells compared to the inflammatory Th17 and Th1 effector cells that differentiated in parallel in this model [19]. Consistent with this, transfer of naive CD4+ T with forced deletion of the Thpok gene in all activated CD4 T cells, resulted in increased differentiation of CD8α-expressing CD4 donor T cells and less inflammation in the intestine of the RAG−/− recipients [19]. In contrast, mice that received CD4 T cells with forced deletion of Runx3, showed accelerated wasting disease, which correlated with an increased IL-17 production [19]. Similarly, mice with a forced deletion of Runx3 in T cells, showed more inflammation, with transmural infiltrates and epithelial cell damage, in response to a Citrobacter rodentium infection, whereas they cleared the pathogen more efficiently than wild-type mice. This was likely due to the greater frequency of IL-17-producing CD4+ effector cells in the absence of Runx3 induction [19]. These observations suggested that, in addition to the promotion of the CTL program, Runx3 might also counteract the Th17 pathway in activated CD4 T cells. Together, these results indicated that the reprogramming of CD4 T cells controlled by the gain of Runx3 and the loss of ThPOK, not only leads to effector cells with direct protective capacity but the mechanism also deviate those cells away from becoming inflammatory Th cells, which have the capacity to cause tissue damage and other pathology.
Nevertheless, the increased accumulation of cytotoxic CD4+ T cells under chronic inflammatory conditions, such as auto-immune [109] and inflammatory bowel disease (IBD) [110], suggests that next to their potential beneficial protective functions, these cells might also participate in or contribute to inflammatory diseases. Indeed, whereas functional studies raised the possibility that disruption of this process may lead to uncontrolled CD4 Th responses, they also highlighted the potential of CD4 CTL to become potent pathogenic killer cells in the context of inflammatory cytokines, such as IL-15, which is excessively produced in IBD and Celiac disease or in virus-induced immune pathology. Although in general, most viral- or vaccine-induced CTL responses result in protective, sterilizing immunity for their hosts, they can also induce disease due to excessive or aberrant cytotoxicity. Virus control, as well as dampening of the inflammatory response, is important for minimizing pathology, however, whether an infection will result in severe or minimal tissue damage depends on numerous factors, not in the least on the inflammatory or (self)-regulatory potential of the responder T cells. For example, whereas normally, intra cranial (i.c.) LCMV infections in immune competent mice causes lethal meningitis and wasting disease mediated by excessive CD8 CTL responses, vaccinated immune competent mice elicit a CD8 CTL response that can either protect from immune pathology, or enhance immune pathology, depending on the potency and kinetics of the acute response [76]. Male mice show a low response and are not susceptible to immune-mediated pathology unless their responses have been boosted by vaccination. Female mice display much stronger responses and pre-existing immunity will protect them from immune pathology due to an effective and rapid CD8 CTL memory response that clears the virus before the immune response becomes excessive. Interestingly, in β2m-deficient mice, which do not have CD8+ T cells, i.c. LCMV infections still lead to lethality in female mice as well as in vaccinated male mice, strongly suggesting that CD4 CTL could develop into pathogenic effector cells. In agreement with this, in autoimmune diseases such as rheumatoid arthritis and ankylosing spondylitis, the amount of CD4+ T cells that display cytolytic features is related to the severity of disease, whereas treatment with anti-TNF, reduces their numbers[111–113]. Nevertheless, as of yet, there is no evidence that links the expanded CD4 CTL with self-antigen specificity. It is therefore also possible that the increase in CD4 CTL numbers associated with inflammation and autoimmunity is cytokine-driven or the consequence of an indirect bystander effect.
9. Conclusion and perspectives
Despite the irrefutable observations of CD4+ T cells with cytolytic capacity in conditions of viral infections, autoimmunity and cancers as well as their significant presence among the mucosal T cells that reside within the gut epithelium, we still lack convincing evidence that CD4 CTL display important direct cytolytic functions in-vivo. Furthermore, continuous skepticism labeling these cells as an in-vitro artifact and doubting their physiological relevance has greatly overshadowed the need to investigate, track or target these cells as part of medical intervention strategies. Notwithstanding this, increasing evidence linking cytolytic CD4 T cells to the control of persistent viral infections and their noticeable inhibitory effects on malignant cell growth, have finally began to turn the spotlight on them. More importantly, the new insights gained from the recent studies [4, 19], identifying these cytolytic CD4 effector cells as a true, separate T cell subset, marked by a unique network of transcription factors and the characterization of the mechanisms that drive their differentiation in-vivo, have opened up new avenues of research, to understand the biology and potentially pivotal roles of CD4 CTL in health and disease. The new appreciation of the functional potential of these cells has also created the necessity to rethink vaccine design and to aim for alternative strategies that also include CD4 CTL. Without a doubt, much progress will be made in the near future in improving medical intervention based on strategies that will exploit CD4+ CTL as an important target cell to prevent or treat infections, cancers and devastating autoimmune and other immune-related diseases.
Highlights.
The thymic CD4 Th-lineage commitment is not fixed and mature MHC class-II restricted CD4 T cells have the flexibility to reprogram to CTL.
The transcription factor, ThPOK, directs the Th-lineage fate of MHC class II restricted thymocytes and continues to suppress the Runx3-driven CTL program in mature CD4 Th cells.
Chronic antigen stimulation of mature CD4 T cells leads to the loss of the ThPOK controlled Th fate and the gain of the Runx3-driven CTL reprogramming.
CD4 CTL activated by their cognate antigen in the context of IL-15 become potent killer cells.
CD4 CTL are associated with chronic viral infections, malignancies and autoimmune disorders.
Chronic antigen-induced CD4 CTL have regulatory capacity.
The reprogram mechanism of CD4 T cells diverts them away from becoming pathogenic inflammatory effector cells.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Aliahmad P, KA, de la Torre B, Kappes D, Kaye J. TOX is required for development of the CD4 T cell lineage gene program. J Immunol. 2011;187:5931–40. doi: 10.4049/jimmunol.1101474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.He X, HX, Dave VP, Zhang Y, Hua X, Nicolas E, Xu W, RoeKD BA. The zinc finger transcription factor Th-POK regulates CD4 versus CD8 T-cell lineage commitment. nature. 2005;433:826–833. doi: 10.1038/nature03338. [DOI] [PubMed] [Google Scholar]
- 3.Hernández-Hoyos G, AM, Wang C, Rothenberg EV, Alberola-Ila J. GATA-3 expression is controlled by TCR signals and regulates CD4/CD8 differentiation. immunity. 2003;19:83–94. doi: 10.1016/s1074-7613(03)00176-6. [DOI] [PubMed] [Google Scholar]
- 4.Mucida D, Husain MM, Muroi S, van Wijk F, Shinnakasu R, Naoe Y, Reis BS, Huang, et al. Transcriptional reprogramming of mature CD4+ helper T cells generates distinct MHC class II–restricted cytotoxic T lymphocytes. Nat Immunol. 2013;14:281–89. doi: 10.1038/ni.2523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Pai SY, TM, Ting CN, Leiden JM, Glimcher LH, Ho IC. Critical roles for transcription factor GATA-3 in thymocyte development. immunity. 2003;19:863–75. doi: 10.1016/s1074-7613(03)00328-5. [DOI] [PubMed] [Google Scholar]
- 6.Sun G, LX, Mercado P, Jenkinson SR, Kypriotou M, Feigenbaum L, Galéra P, Bosselut R. The zinc finger protein cKrox directs CD4 lineage differentiation during intrathymic T cell-positive selection. Nat Immunol. 2005;6:373–81. doi: 10.1038/ni1183. [DOI] [PubMed] [Google Scholar]
- 7.Wang L, WK, Castro E, Xiong Y, Feigenbaum L, TessarolloBR L. The Zinc Finger Transcription Factor Zbtb7b Represses CD8-Lineage Gene Expression in Peripheral CD4+ T Cells. immunity. 2008;29:876–887. doi: 10.1016/j.immuni.2008.09.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Taniuchi I, OM, Egawa T, Sunshine MJ, Bae SC, Komori T, Ito Y, Littman DR. Differential requirements for Runx proteins in CD4 repression and epigenetic silencing during T lymphocyte development. Cell. 2002;111:621–633. doi: 10.1016/s0092-8674(02)01111-x. [DOI] [PubMed] [Google Scholar]
- 9.Woolf E, XC, Fainaru O, Lotem J, Rosen D, Negreanu V, Bernstein Y, Goldenberg D, Brenner O, Berke G, Levanon D, Groner Y. Runx3 and Runx1 are required for CD8 T cell development during thymopoiesis. Proc Natl Acad Sci U S A. 2003;100:7731–6. doi: 10.1073/pnas.1232420100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.van de Berg PJ, vLE, ten Berge IJ, van Lier R. Cytotoxic human CD4+ T cells. Curr Opin Immunol. 2008;20:339–343. doi: 10.1016/j.coi.2008.03.007. [DOI] [PubMed] [Google Scholar]
- 11.Brown DM. Cytolytic CD4 Cells: Direct Mediators in Infectious Disease and Malignancy. Cell Immunol. 2010;262:89–95. doi: 10.1016/j.cellimm.2010.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Krensky AM, CSR, Mier JW, Strominger JL, Burakoff SJ. Long-term human cytolytic T-cell lines allospecific for HLA-DR6 antigen are OKT4. Proc Natl Acad Sci U S A. 1982;79:2365–2369. doi: 10.1073/pnas.79.7.2365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lukacher AE, ML, Braciale VL, Malissen B, Braciale TJ. Expression of specific cytolytic activity by H-2I region-restricted, influenza virus-specific T lymphocyte clones. J Exp Med. 1985;162:171–87. doi: 10.1084/jem.162.1.171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.McCarthy SA. Recognition of MHC class I allodeterminants regulates the generation of MHC class II-specific CTL. J Immunol. 1986;137:3087–92. [PubMed] [Google Scholar]
- 15.Man S, Lechler RI, Batchelor JR, Sharrock CE. Individual variation in the frequency of HLA class II-specific cytotoxic T lymphocyte precursors. Eur J Immunol. 1990;20:847. doi: 10.1002/eji.1830200420. [DOI] [PubMed] [Google Scholar]
- 16.Suni MA, Ghanekar SA, Houck DW, Maecker HT, Wormsley SB, Picker LJ, Moss RB, Maino VC. CD4_CD8dim T lymphocytes exhibit enhanced cytokine expression, proliferation and cytotoxic activity in response to HCMV and HIV-1 antigens. Eur J Immunol. 2001;31:2512. doi: 10.1002/1521-4141(200108)31:8<2512::aid-immu2512>3.0.co;2-m. [DOI] [PubMed] [Google Scholar]
- 17.Fleischer B. Acquisition of specific cytotoxic activity by human T4+ T lymphocytes in culture. Nature. 1984;308:365–367. doi: 10.1038/308365a0. [DOI] [PubMed] [Google Scholar]
- 18.Williams NS, V, Engelhard H. Identification of a population of CD4+ CTL that utilizes a perforin, rather than a Fas ligand-dependent cytotoxic mechanism. J Immunol. 1996;156:153–9. [PubMed] [Google Scholar]
- 19.Reis BS, RA, Costa-Pinto FA, Taniuchi I, Mucida D. Mutual expression of the transcription factors Runx3 and ThPOK regulates intestinal CD4+ T cell immunity. Nat Immunol. 2013;14:271– 280. doi: 10.1038/ni.2518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Naito T, TH, Naoe Y, Taniuchi I. Transcriptional control of T-cell development. Int Immunol. 2011;23:661–68. doi: 10.1093/intimm/dxr078. [DOI] [PubMed] [Google Scholar]
- 21.Gangadharan D, LF, Attinger A, Wang-Zhu Y, Sullivan BA, Cheroutre H. Identification of pre- and postselection TCRalphabeta+ intraepithelial lymphocyte precursors in the thymus. immunity. 2006;25:631–41. doi: 10.1016/j.immuni.2006.08.018. [DOI] [PubMed] [Google Scholar]
- 22.Zhu J, MB, Hu-Li J, Watson CJ, Grinberg A, Wang Q, Killeen N, Urban JF, Jr, Guo L, Paul WE. Conditional deletion of Gata3 shows its essential function in T(H)1–T(H)2 responses. nat Immunol. 2004;5:1157–65. doi: 10.1038/ni1128. [DOI] [PubMed] [Google Scholar]
- 23.Ho IC, TT, Pai SY. GATA3 and the T-cell lineage: essential functions before and after T-helper-2-cell differentiation. nat Rev Immunol. 2009;9:125–35. doi: 10.1038/nri2476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hendriks RW, NM, Engel JD, van Doorninck H, Grosveld F, Karis A. Expression of the transcription factor GATA-3 is required for the development of the earliest T cell progenitors and correlates with stages of cellular proliferation in the thymus. European J Immunol. 1999;29:1912–8. doi: 10.1002/(SICI)1521-4141(199906)29:06<1912::AID-IMMU1912>3.0.CO;2-D. [DOI] [PubMed] [Google Scholar]
- 25.Wang L, WK, Zhu J, Zhang X, Feigenbaum L, Tessarollo L, Paul WE, Fowlkes BJ, Bosselut R. Distinct functions for the transcription factors GATA-3 and ThPOK during intrathymic differentiation of CD4+ T cells. nat immunol. 2008;9:1122–30. doi: 10.1038/ni.1647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Egawa T, LD ThPOK acts late in specification of the helper T cell lineage and suppresses Runx-mediated commitment to the cytotoxic T cell lineage. nat Immunol. 2008;9:1131–39. doi: 10.1038/ni.1652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Setoguchi R, Tachibana M, Naoe Y, Muroi S, Akiyama K, Tezuka C, Okuda T, Taniuchi I. Repression of the transcription factor Th-POK by Runx complexes in cytotoxic T cell development. Science. 2008;319:822–825. doi: 10.1126/science.1151844. [DOI] [PubMed] [Google Scholar]
- 28.Kappes DJ, HX, He X. Role of the transcription factor Th-POK in CD4:CD8 lineage commitment. immunol rev. 2006;209:237–52. doi: 10.1111/j.0105-2896.2006.00344.x. [DOI] [PubMed] [Google Scholar]
- 29.Cruz-Guilloty F, PM, Djuretic IM, Levanon D, Lotem J, Lichtenheld MG, Groner Y, Rao A. Runx3 and T-box proteins cooperate to establish the transcriptional program of effector CTLs. J Exp Med. 2009;206:51–59. doi: 10.1084/jem.20081242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Egawa T, Tillman RE, Naoe Y, Taniuchi I, Littman DR. The role of the Runx transcription factors in thymocyte differentiation and in homeostasis of naive T cells. J Expl Med. 2007;204:1945–1957. doi: 10.1084/jem.20070133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sato T, OS, Hayashi T, Sato C, Kohu K, Satake M, Habu S. Dual functions of Runx proteins for reactivating CD8 and silencing CD4 at the commitment process into CD8 thymocytes. immunity. 2005;22:317–328. doi: 10.1016/j.immuni.2005.01.012. [DOI] [PubMed] [Google Scholar]
- 32.Collins A, Littman DR, Taniuchi I. RUNX proteins in transcription factor networks that regulate T-cell lineage choice. nat Rev Immunol. 2009;9:106–115. doi: 10.1038/nri2489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Liu X, TB, Sun G, Bosselut R. Analyzing expression of perforin, Runx3, and Thpok genes during positive selection reveals activation of CD8-differentiation programs by MHC II-signaled thymocytes. J Immunol. 2005;175:4465–74. doi: 10.4049/jimmunol.175.7.4465. [DOI] [PubMed] [Google Scholar]
- 34.Grueter B, PM, Egawa T, Laule-Kilian K, Aldrian CJ, Wuerch A, Ludwig Y, Fukuyama H, Wardemann H, Waldschuetz R, Möröy T, Taniuchi I, Steimle V, Littman DR, Ehlers M. Runx3 regulates integrin alpha E/CD103 and CD4 expression during development of CD4-/CD8+ T cells. J Immunol. 2005;175:1694–1705. doi: 10.4049/jimmunol.175.3.1694. [DOI] [PubMed] [Google Scholar]
- 35.Kohu K, ST, Ohno S, Hayashi K, Uchino R, Abe N, Nakazato M, Yoshida N, Kikuchi T, Iwakura Y, Inoue Y, Watanabe T, Habu S, Satake M. Overexpression of the Runx3 transcription factor increases the proportion of mature thymocytes of the CD8 singlepositive lineage. J Immunol. 2005;174:2627–2636. doi: 10.4049/jimmunol.174.5.2627. [DOI] [PubMed] [Google Scholar]
- 36.Bilic I, EW The role of BTB domain-containing zinc finger proteins in T cell development and function. Immunol Lett. 2007;108:1–9. doi: 10.1016/j.imlet.2006.09.007. [DOI] [PubMed] [Google Scholar]
- 37.Bilic I, Koesters C, Unger B, Sekimata M, Hertweck A, Maschek R, Wilson CB, Ellmeier aW. Negative regulation of CD8 expression via Cd8 enhancermediated recruitment of the zinc finger protein MAZR. nat Immunol. 2006;7:392–400. doi: 10.1038/ni1311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Laky K, Fowlkes BJ. Notch signaling in CD4 and CD8 T cell development. Curr Opin Immunol. 2008;20:197–202. doi: 10.1016/j.coi.2008.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Laky K, Fowlkes BJ. Presenilins regulate alphabeta T cell development by modulating TCR signaling. J Exp Med. 2007;204:2115–29. doi: 10.1084/jem.20070550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Jones ME, Zhuang Y. Acquisition of a functional T cell receptor during T lymphocyte development is enforced by HEB and E2A transcription factors. immunity. 2007;27:860–870. doi: 10.1016/j.immuni.2007.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Jimi E, Strickland I, Voll RE, Long M, Ghosh S. Differential Role of the Transcription Factor NF-κB in Selection and Survival of CD4+ and CD8+ Thymocytes. immunity. 2008;29:523–537. doi: 10.1016/j.immuni.2008.08.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Djuretic IM, Levanon D, Negreanu V, Groner Y, Rao A, Ansel KM. Transcription factors T-bet and Runx3 cooperate to activate Ifng and silence Il4 in T helper type 1 cells. nat Immunol. 2007;8:145–153. doi: 10.1038/ni1424. [DOI] [PubMed] [Google Scholar]
- 43.Naoe Y, Setoguchi R, Akiyama K, Muroi S, Kuroda M, Hatam F, Littman DR, Taniuchi I. Repression of interleukin-4 in T helper type 1 cells by Runx/Cbfβ binding to the Il4 silencer. J Exp Med. 2007;204:1749–1755. doi: 10.1084/jem.20062456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Jenkinson SR, Intlekofer AM, Sun G, Feigenbaum L, Reiner SL, Bosselut R. Expression of the transcription factor cKrox in peripheral CD8 T cells reveals substantial postthymic plasticity in CD4-CD8 lineage differentiation. J Exp Med. 2007;204:267–272. doi: 10.1084/jem.20061982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Ouyang W, LM, Gao Z, Assenmacher M, Ranganath S, Radbruch A, Murphy KM. Stat6-independent GATA-3 autoactivation directs IL-4-independent Th2 development and commitment. immunity. 2000;12:27–37. doi: 10.1016/s1074-7613(00)80156-9. [DOI] [PubMed] [Google Scholar]
- 46.Pai SY, TM, Ho IC. GATA-3 deficiency abrogates the development and maintenance of T helper type 2 cells. Proc Natl Acad Sci U S A. 2004;101:1993–8. doi: 10.1073/pnas.0308697100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wang Y, SM, Wan YY. An essential role of the transcription factor GATA-3 for the function of regulatory T cells. immunity. 2011;35:337–48. doi: 10.1016/j.immuni.2011.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wohlfert EA, GJ, Bouladoux N, Konkel JE, Oldenhove G, Ribeiro CH, Hall JA, Yagi R, Naik S, Bhairavabhotla R, Paul WE, Bosselut R, Wei G, Zhao K, Oukka M, Zhu J, Belkaid Y. GATA3 controls Foxp3+ regulatory T cell fate during inflammation in mice. J Clin Invest. 2011;121:4503–15. doi: 10.1172/JCI57456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.van Hamburg JP, dBM, Ribeiro de Almeida C, van Zwam M, van Meurs M, de Haas E, Boon L, Samsom JN, Hendriks RW. Enforced expression of GATA3 allows differentiation of IL-17-producing cells, but constrains Th17-mediated pathology. Eur J Immunol. 2008;38:2573–86. doi: 10.1002/eji.200737840. [DOI] [PubMed] [Google Scholar]
- 50.Kanhere A, HA, Bhatia U, Gökmen MR, Perucha E, Jackson I, Lord GM, Jenner RG. T-bet and GATA3 orchestrate Th1 and Th2 differentiation through lineage-specific targeting of distal regulatory elements. nature communications. 2012;3:1268. doi: 10.1038/ncomms2260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Pearce EL, MA, Martins GA, Krawczyk CM, Hutchins AS, Zediak VP, Banica M, DiCioccio CB, Gross DA, Mao CA, Shen H, Cereb N, Yang SY, Lindsten T, Rossant J, Hunter CA, Reiner SL. Control of effector CD8+ T cell function by the transcription factor Eomesodermin. Science. 2003;302:1041–3. doi: 10.1126/science.1090148. [DOI] [PubMed] [Google Scholar]
- 52.Intlekofer AM, BA, Takemoto N, Gordon SM, Dejong CS, Shin H, HunterWE CA, Lindsten T, Reiner SL. Anomalous type 17 response to viral infection by CD8+ T cells lacking T-bet and eomesodermin. Science. 2008;321:408–11. doi: 10.1126/science.1159806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Li Y, TA, Gao Z, Han Z, Gao Y, Li Z, Lin F, Wang Y, Wei G, Yao Z, Li B. 60-kDa Tat-interactive protein (TIP60) positively regulates Th-inducing POK (ThPOK)-mediated repression of eomesodermin in human CD4+ T cells. J Biol Chem. 2013;288(22):15537–46. doi: 10.1074/jbc.M112.430207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Carpenter AC, GJ, Xiong Y, Kanno Y, Chu HH, Wang L, Naik S, dos Santos L, Wei L, Jenkins MK, O’Shea JJ, Belkaid Y, Bosselut R. The Transcription Factors Thpok and LRF Are Necessary and Partly Redundant for T Helper Cell Differentiation. immunity. 2012;37:622–33. doi: 10.1016/j.immuni.2012.06.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Casazza JP, BM, Price DA, Precopio ML, Ruff LE, Brenchley JM, Hill BJ, Roederer M, Douek DC, Koup RA. Acquisition of direct antiviral effector functions by CMV-specific CD4+ T lymphocytes with cellular maturation. J Exp Med. 2006;203:2865–77. doi: 10.1084/jem.20052246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Haigh TA, LX, Jia H, Hui EP, Chan AT, Rickinson AB, Taylor GS. EBV latent membrane proteins (LMPs) 1 and 2 as immunotherapeutic targets: LMP-specific CD4+ cytotoxic T cell recognition of EBV-transformed B cell lines. J Immunol. 2008;180:1643–54. doi: 10.4049/jimmunol.180.3.1643. [DOI] [PubMed] [Google Scholar]
- 57.Appay V, ZJ, Papagno L, Sutton J, Jaramillo A, Waters A, Easterbrook P, Grey P, Smith D, McMichael AJ, Cooper DA, Rowland-Jones SL, Kelleher AD. Characterization of CD4+ CTLs Ex Vivo. J Immunol. 2002;168:5954–58. doi: 10.4049/jimmunol.168.11.5954. [DOI] [PubMed] [Google Scholar]
- 58.Swain NBMaSL. Cytotoxic CD4 T Cells in Antiviral Immunity. Journal of Biomedicine and Biotechnology. 2011 doi: 10.1155/2011/954602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Fu J, ZZ, Zhou L, Qi Z, Xing S, Lv J, Shi J, Fu B, Liu Z, Zhang JY, Jin L, Zhao Y, Lau GK, Zhao J, Wang FS. Impairment of CD4+ cytotoxic T cells predicts poor survival and high recurrence rates in patients with hepatocellular carcinoma. Hepatology. 2013;58:139–49. doi: 10.1002/hep.26054. [DOI] [PubMed] [Google Scholar]
- 60.Martorelli D, ME, Merlo A, Turrini R, Rosato A, Dolcetti R. Role of CD4+ Cytotoxic T Lymphocytes in the Control of Viral Diseases and Cancer. Int Rev Immunol. 2010;29:371–402. doi: 10.3109/08830185.2010.489658. [DOI] [PubMed] [Google Scholar]
- 61.Broux B, PK, Zhang X, Markovic-Plese S, Broekmans T, Eijnde BO, Van Wijmeersch B, Somers V, Geusens P, van der Pol S, van Horssen J, Stinissen P, Hellings N. CX(3)CR1 drives cytotoxic CD4(+)CD28(-) T cells into the brain of multiple sclerosis patients. j Autoimmun. 2012;38(1):10–9. doi: 10.1016/j.jaut.2011.11.006. [DOI] [PubMed] [Google Scholar]
- 62.Pawlik A, OL, Brzosko I, Brzosko M, Masiuk M, Machalinski B, Gawronska-Szklarz B. The expansion of CD4+CD28- T cells in patients with rheumatoid arthritis. Arthritis Res Ther. 2003;5:R210–3. doi: 10.1186/ar766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.van Leeuwen EM, RE, Vossen MT, Rowshani AT, Wertheim-van Dillen PM, van Lier RA, ten Berge IJ. Emergence of a CD4+CD28- granzyme B+, cytomegalovirus-specific T cell subset after recovery of primary cytomegalovirus infection. journal of Immunology. 2004;173:1834–41. doi: 10.4049/jimmunol.173.3.1834. [DOI] [PubMed] [Google Scholar]
- 64.Kennedy J, VA, Saylor V, Zurawski SM, Copeland NG, Gilbert DJ, Jenkins NA, Zlotnik A. A molecular analysis of NKT cells: identification of a class-I restricted T cell-associated molecule (CRTAM) J Leuk Biol. 2000;67:725–34. doi: 10.1002/jlb.67.5.725. [DOI] [PubMed] [Google Scholar]
- 65.Boles KS, BW, Diacovo T, Cella M, Colonna M. The tumor suppressor TSLC1/NECL-2 triggers NK-cell and CD8+ T-cell responses through the cell-surface receptor CRTAM. Blood. 2005;106:779–86. doi: 10.1182/blood-2005-02-0817. [DOI] [PubMed] [Google Scholar]
- 66.Boles KS, SS, Bennett M, Kumar V, Mathew PA. 2B4 (CD244) and CS1: novel members of the CD2 subset of the immunoglobulin superfamily molecules expressed on natural killer cells and other leukocytes. Immunol rev. 2001;181:234–49. doi: 10.1034/j.1600-065x.2001.1810120.x. [DOI] [PubMed] [Google Scholar]
- 67.Sakaguchi S, HM, Bilic I, Naoe Y, Schebesta A, TaniuchiEW I. The zinc-finger protein MAZR is part of the transcription factor network that controls the CD4 versus CD8 lineage fate of double-positive thymocytes. nat Immunol. 2010;11(5):442–448. doi: 10.1038/ni.1860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Jacobson S, RJ, Biddison WE, Satinsky A, Hartzman RJ, McFarland HF. Measles virus-specificT4+ human cytotoxic T cell clones are restricted by class II HLA antigens. J Immunol. 1984;133:754–7. [PubMed] [Google Scholar]
- 69.DePaolo RW, AV, Tang F, Fehlner-Peach H, Hall JA, Wang W, Marietta EV, Kasarda DD, Waldmann TA, Murray JA, Semrad C, Kupfer SS, Belkaid Y, Guandalini S, Jabri B. Co-adjuvant effects of retinoic acid and IL-15 induce inflammatory immunity to dietary antigens. Nature. 2011;471:220–4. doi: 10.1038/nature09849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Mention JJ, BAM, Bègue B, Barbe U, Verkarre V, Asnafi V, Colombel JF, Cugnenc PH, Ruemmele FM, McIntyre E, Brousse N, Cellier C, Cerf-Bensussan N. Interleukin 15: a key to disrupted intraepithelial lymphocyte homeostasis and lymphomagenesis in celiac disease. Gastroenterol. 2003;125:730–45. doi: 10.1016/s0016-5085(03)01047-3. [DOI] [PubMed] [Google Scholar]
- 71.ECE. Interleukin 15 is a potent stimulant of intraepithelial lymphocytes. Gastroenterol. 1998;115:1439–45. doi: 10.1016/s0016-5085(98)70022-8. [DOI] [PubMed] [Google Scholar]
- 72.Ye W, YJ, Liu CC. Interleukin-15 induces the expression of mRNAs of cytolytic mediators and augments cytotoxic activities in primary murine lymphocytes. cell immunol. 1996;174:54–62. doi: 10.1006/cimm.1996.0293. [DOI] [PubMed] [Google Scholar]
- 73.Abadie V, DV, Jabri B. Intraepithelial lymphocytes in celiac disease immunopathology. Semin Immunopathol. 2012;34:551–66. doi: 10.1007/s00281-012-0316-x. [DOI] [PubMed] [Google Scholar]
- 74.MHA. Patterns of persistent viral infections. N Engl J Med. 1986;315:939–48. doi: 10.1056/NEJM198610093151506. [DOI] [PubMed] [Google Scholar]
- 75.MSH. Cytomegalovirus and its role in the pathogenesis of acquired immunodeficiency syndrome. Transplant Proc. 1991;23:118–21. [PubMed] [Google Scholar]
- 76.Kang SS, MD Lymphocytic choriomeningitis infection of the central nervous system. Front Biosci. 2008;13:4529–43. doi: 10.2741/3021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Jellison ER, KS, Welsh RM. Cutting edge: MHC class II-restricted killing in vivo during viral infection. J Immunol. 2005;174:614–8. doi: 10.4049/jimmunol.174.2.614. [DOI] [PubMed] [Google Scholar]
- 78.Arens R, WP, Sidney J, Loewendorf A, Sette A, Schoenberger SP, Peters B, Benedict CA. Cutting edge: murine cytomegalovirus induces a polyfunctional CD4 T cell response. J Immunol. 2008;180:6472–6. doi: 10.4049/jimmunol.180.10.6472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Jonji S, PI, Lucin P, Rukavina D, Koszinowski UH. Efficacious control of cytomegalovirus infection after long-term depletion of CD8+ T lymphocytes. J virol. 1990;64:5457–64. doi: 10.1128/jvi.64.11.5457-5464.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Rentenaar RJ, GL, van DerHoek N, van Diepen FN, Boom R, Weel JF, Wertheim-van Dillen PM, van Lier RA, ten Berge IJ. Development of virus-specific CD4(+) T cells during primary cytomegalovirus infection. J Clin Invest. 2000;105:541–8. doi: 10.1172/JCI8229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.van Leeuwen EM, RE, Heemskerk MH, ten Berge IJ, van Lier RA. Strong selection of virus-specific cytotoxic CD4+ T-cell clones during primary human cytomegalovirus infection. Blood. 2006;108:3121–7. doi: 10.1182/blood-2006-03-006809. [DOI] [PubMed] [Google Scholar]
- 82.Crompton L, KN, Khanna R, Nayak L, Moss PA. CD4+ T cells specific for glycoprotein B from cytomegalovirus exhibit extreme conservation of T-cell receptor usage between different individuals. Blood. 2008;111:2053–61. doi: 10.1182/blood-2007-04-079863. [DOI] [PubMed] [Google Scholar]
- 83.Elkington R, SN, Walker S, Crough T, Fazou C, Kaur A, Walker CM, Khanna R. Cross-reactive recognition of human and primate cytomegalovirus sequences by human CD4 cytotoxic T lymphocytes specific for glycoprotein B and H. European J Immunol. 2004;34:3216–26. doi: 10.1002/eji.200425203. [DOI] [PubMed] [Google Scholar]
- 84.Hegde NR, DC, Lewinsohn DM, Jarvis MA, Nelson JA, Johnson DC. Endogenous human cytomegalovirus gB is presented efficiently by MHC class II molecules to CD4+ CTL. J exp med. 2005;202:1109–19. doi: 10.1084/jem.20050162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Zheng CF, ML, Jones GJ, Gill MJ, Krensky AM, Kubes P, Mody CH. Cytotoxic CD4+ T cells use granulysin to kill Cryptococcus neoformans, and activation of this pathway is defective in HIV patients. Blood. 2007;109:2049–57. doi: 10.1182/blood-2006-03-009720. [DOI] [PubMed] [Google Scholar]
- 86.Sáez-Borderías A, GM, Angulo A, Bellosillo B, Pende D, López-Botet M. Expression and function of NKG2D in CD4+ T cells specific for human cytomegalovirus. Eur J Immunol. 2006;36:3198–206. doi: 10.1002/eji.200636682. [DOI] [PubMed] [Google Scholar]
- 87.Zhou W, SM, Martinez J, Srivastava T, Diamond DJ, Knowles W, Lacey SF. Functional characterization of BK virus-specific CD4+ T cells with cytotoxic potential in seropositiveadults. viral immunol. 2007;20:379–88. doi: 10.1089/vim.2007.0030. [DOI] [PubMed] [Google Scholar]
- 88.Milikan JC, KP, Baarsma GS, Kuijpers RW, Osterhaus AD, Verjans GM. Identification of viral antigens recognized by ocular infiltrating T cells from patients with varicella zoster virus-induced uveitis. Invest Ophthalmol Vis Sci. 2007;48:3689–97. doi: 10.1167/iovs.07-0020. [DOI] [PubMed] [Google Scholar]
- 89.Aslan N, YC, Wiegand J, Greten T, Ciner A, Meyer MF, Heiken H, Kuhlmann B, Kaiser T, Bozkaya H, Tillmann HL, Bozdayi AM, Manns MP, Wedemeyer H. Cytotoxic CD4 T cells in viral hepatitis. J Viral Hepat. 2006;13:505–14. doi: 10.1111/j.1365-2893.2006.00723.x. [DOI] [PubMed] [Google Scholar]
- 90.Silva CL, LD Identification and characterization of murine cytotoxic T cells that kill Mycobacterium tuberculosis. Infect Immun. 2000;68:3269–74. doi: 10.1128/iai.68.6.3269-3274.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Landmeier S, AB, Pscherer S, Eing BR, Kuehn J, Rooney CM, Juergens H, Rossig C. Gene-engineered varicella-zoster virus reactive CD4+ cytotoxic T cells exert tumor-specific effector function. Cancer Res. 2007;67:8335–43. doi: 10.1158/0008-5472.CAN-06-4426. [DOI] [PubMed] [Google Scholar]
- 92.Ohminami H, YM, Kaneko S, Yakushijin Y, Abe Y, Kasahara Y, Ishida Y, FS Fas-independent and nonapoptotic cytotoxicity mediated by a human CD4(+) T-cell clone directed against an acute myelogenous leukemia-associated DEK-CAN fusion peptide. Blood. 1999;93:925–35. [PubMed] [Google Scholar]
- 93.Hanley PJ, CC, Savoldo B, Leen AM, Stanojevic M, Khalil M, Decker W, LH, Molldrem JJ, Gee AP, Rooney CM, Heslop HE, Dotti G, Brenner MK, Shpall EJ, BCM Functionally active virus-specific T cells that target CMV, adenovirus, and EBV can be expanded from naive T-cell populations in cord bloodand will target a range of viral epitopes. Blood. 2009;114:1958–67. doi: 10.1182/blood-2009-03-213256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.McKinstry KK, ST, Kuang Y, Brown DM, Sell S, Dutton RW, Swain SL. Memory CD4+ T cells protect against influenza through multiple synergizing mechanisms. J Clin Invest. 2012;122:2847–56. doi: 10.1172/JCI63689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Hegde NR, JD Human cytomegalovirus US2 causes similar effects on both major histocompatibility complex class I and II proteins in epithelial and glial cells. j virol. 2003;77:9287–94. doi: 10.1128/JVI.77.17.9287-9294.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.von Herrath M, HA Pathological changes in the islet milieu precede infiltration of islets and destruction of beta-cells by autoreactive lymphocytes in a transgenic model of virus-induced IDDM. J Autoimmun. 1997;10:231–8. doi: 10.1006/jaut.1997.0131. [DOI] [PubMed] [Google Scholar]
- 97.Debbabi H, GS, Kamath AB, Alt J, Demello DE, Dunsmore S, Behar SM. Primary type II alveolar epithelial cells present microbial antigens to antigen-specific CD4+ T cells. Am J Physiol Lung Cell Mol Physiol. 2005;289:L274–9. doi: 10.1152/ajplung.00004.2005. [DOI] [PubMed] [Google Scholar]
- 98.Gao J, DB, Banerjee AK. Human parainfluenza virus type 3 up-regulates major histocompatibility complex class I and II expression on respiratory epithelial cells: involvement of a STAT1- and CIITA-independent pathway. J virol. 1999;72:1411–8. doi: 10.1128/jvi.73.2.1411-1418.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Ibrahim L, DM, Yacoub M. Primary human adult lung epithelial cells in vitro: response to interferon-gamma and cytomegalovirus. immunol. 1993;79:119–24. [PMC free article] [PubMed] [Google Scholar]
- 100.Bottazzo GF, DB, McNally JM, MacKay EH, Swift PG, Gamble DR. In situ characterization of autoimmune phenomena and expression of HLA molecules in the pancreas in diabetic insulitis. N Engl J Med. 1985;313:353–60. doi: 10.1056/NEJM198508083130604. [DOI] [PubMed] [Google Scholar]
- 101.Foulis AK, FM Aberrant expression of HLA-DR antigens by insulin-containing beta-cells in recent-onset type I diabetes mellitus. Diabetes. 1986;35:1215–24. doi: 10.2337/diab.35.11.1215. [DOI] [PubMed] [Google Scholar]
- 102.Seewaldt S, TH, Ejrnaes M, Christen U, Wolfe T, Rodrigo E, Coon B, KT, Michelsen B, von Herrath MG. Virus-induced autoimmune diabetes: most beta-cells die through inflammatory cytokines and not perforin from autoreactive (antiviral) cytotoxic T-lymphocytes. Diabetes. 2000;49:1801–9. doi: 10.2337/diabetes.49.11.1801. [DOI] [PubMed] [Google Scholar]
- 103.Robertson KA, UE, Nash AA. Regression of a murine gammaherpesvirus 68-positive b-cell lymphoma mediated by CD4 T lymphocytes. J virol. 2001;75:3480–2. doi: 10.1128/JVI.75.7.3480-3482.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Sparks-Thissen RL, BD, Kreher S, Speck SH, Virgin HW., 4th An optimized CD4 T-cell response can control productive and latent gammaherpesvirus infection. j virol. 2004;78:6827–35. doi: 10.1128/JVI.78.13.6827-6835.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Nikiforow S, BK, Miller G. CD4+ T-cell effectors inhibit Epstein-Barr virus-induced B-cell proliferation. j virol. 2001;75:3740–52. doi: 10.1128/JVI.75.8.3740-3752.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Heller KN, GC, Münz C. Virus-specific CD4+ T cells: ready for direct attack. j exp med. 2006;203:805–8. doi: 10.1084/jem.20060215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Paludan C, BK, Nikiforow S, Tsang ML, Goodman K, Hanekom WA, FonteneauSS JF, Münz C. Epstein-Barr nuclear antigen 1-specific CD4(+) Th1 cells kill Burkitt’s lymphoma cells. J Immuno. 2002;169:1593–603. doi: 10.4049/jimmunol.169.3.1593. [DOI] [PubMed] [Google Scholar]
- 108.Bollard CM, GS, Leen AM, Weiss H, Straathof KC, Carrum G, Khalil M, HM, Wu MF, Chang CC, Gresik MV, Gee AP, Brenner MK, Rooney CM, Heslop HE. Complete responses of relapsed lymphoma following genetic modification of tumor-antigen presenting cells and T-lymphocyte transfer. Blood. 2007;110:2838–45. doi: 10.1182/blood-2007-05-091280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Thewissen M, SV, Hellings N, Fraussen J, Damoiseaux J, Stinissen P. CD4+CD28null T cells in autoimmune disease: pathogenic features and decreased susceptibility to immunoregulation. J Immunol. 2007;179:6514–23. doi: 10.4049/jimmunol.179.10.6514. [DOI] [PubMed] [Google Scholar]
- 110.Kobayashi T, OS, Iwakami Y, Nakazawa A, Hisamatsu T, Chinen H, KamadaIT N, Goto H, Hibi T. Exclusive increase of CX3CR1+CD28-CD4+ T cells in inflammatory bowel disease and their recruitment as intraepithelial lymphocytes. Inflamm Bowel Dis. 2007;13:837–46. doi: 10.1002/ibd.20113. [DOI] [PubMed] [Google Scholar]
- 111.Rizzello V, LG, Brugaletta S, Rebuzzi A, Biasucci LM, Crea F. Modulation of CD4(+)CD28null T lymphocytes by tumor necrosis factor-alpha blockade in patients with unstable angina. Circulation. 2006;113:2272–7. doi: 10.1161/CIRCULATIONAHA.105.588533. [DOI] [PubMed] [Google Scholar]
- 112.Raffeiner B, DC, Duftner C, Kullich W, Goldberger C, Vega SC, Keller M, SM, Grubeck-Loebenstein B. Between adaptive and innate immunity: TLR4-mediated perforin production by CD28null T-helper cells in ankylosing spondylitis. Arthritis Res Ther. 2005;7:R1412–20. doi: 10.1186/ar1840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Gerli R, SG, Giordano A, Bocci EB, Bistoni O, Vaudo G, Marchesi S, RF, Pirro M, Shoenfeld Y, Mannarino E. CD4+CD28- T lymphocytes contribute to early atherosclerotic damage in rheumatoid arthritis patients. Circulation. 2004;109:2744–8. doi: 10.1161/01.CIR.0000131450.66017.B3. [DOI] [PubMed] [Google Scholar]