

Abstract
While evaluating impact of Au nanoparticles on seed germination and early seedling growth of cowpea, HAuCl4 was used as control. Seedlings of cowpea raised in HAuCl4, even at concentration as high as 1 mM, did not show any suppression in growth. Accordingly, Au3+, despite being a heavy metal, did not alter levels of stress markers (viz. proline and malondialdehyde) in cowpea. Interestingly, cowpea turned clear pale yellow HAuCl4 solutions colloidal purple during the course of seed germination and seedling growth. These purple colloidal suspensions showed Au-nanoparticle specific surface plasmon resonance band in absorption spectra. Transmission electron microscopic and powder X-ray diffraction investigations confirmed presence of crystalline Au-nanoparticles in these purple suspensions. Each germinating seed of cowpea released ∼35 nmoles of GAE of phenolics and since phenolics promote generation of Au-nanoparticles, which are less/non toxic compared to Au3+, it was contemplated that potential of cowpea to withstand Au3+ is linked to phenolics. Of the different components of germinating seed of cowpea tested, seed coat possessed immense power to generate Au-nanoparticles, as it was the key source of phenolics. To establish role of phenolics in generation of Au-nanoparticles (i) seed coat and (ii) the incubation medium in which phenolics were released by germinating seeds, were tested for their efficacy to generate Au-nanoparticles. Interestingly, incubation of either of these components with Au3+ triggered increase in generation of Au-nanoparticles with concomitant decrease in phenolics. Accordingly, with increase in concentration of Au3+, a proportionate increase in generation of Au-nanoparticles and decrease in phenolics was recorded. In summary, our findings clearly established that cowpea possessed potential to withstand Au3+-stress as the phenolics released by seed coat of germinating seeds possess potential to reduce toxic Au3+ to form non/less toxic Au-nanoparticles. Our investigations also pave a novel, simple, green and economically viable protocol for generation of Au-nanoparticles.
Introduction
With the rapid expansion of electronic industry, the demand and cost of gold has increased markedly over past four decades. In general, gold comes into the environment from primary (i.e. ores) and secondary sources such as electronic scrap and waste electroplating solutions [1]–[3]. Au, whose density is 19.32 g cm−3, like other heavy metals, has a negative impact on physiology and biochemistry of microorganisms and animal systems including humans [4], [5]. Researchers working with animal systems could trace Au in various organs including ovaries, hypothalamus, liver, adrenals, kidneys, testes, lymph nodes and pituitary glands [6]. In fact, Au has been shown to be transported even over placental barrier into human embryo. In majority of cases, Au was located in lysosomes and whenever the concentration of Au exceeded a certain level, the lysosomal membrane ruptured releasing its contents into the cytosol [6]. In addition, Au also interferes with functioning of energy transducing system (i.e. mitochondria), nuclei and vacuoles [4]. To the best of our knowledge, no significant studies have been carried to investigate the impact of Au on plant growth and development. However, there are reports on accumulation of Au, synthesis and accumulation of Au-nanoparticles in cells of plants exposed to Au salts [7]–[10].
The degree and mechanism of tolerance to heavy metals vary significantly amongst plant species [11]. The basic mechanisms evolved by plants to counter heavy metal tolerance include (i) formation of exogenous non-toxic metal-chelates with organic acids, polyphosphates and siderphores, which restrict metal uptake [12]–[14]; (ii) interaction of toxic metal species with ligands located on cell surface/wall [15]; (iii) active (i.e. energy demanding) efflux of metal ions [16]; and (iv) endogenous chelation/sequestration involving various biomolecules including citrate, oxalate, malate [15], phytochelatins, metallothioneins [17] and phenolic compounds [18], [19]. Many phenolic compounds have been reported to have superior tendency to form stable complexes with most widespread toxic metals such as Ni, Cu, Co and Mn than many organic acids [19]. Phenolics, characterized by at least one aromatic ring (C6) bearing one or more hydroxyl groups [20], are a group of low molecular weight secondary metabolites that are known to impart heavy metal stress tolerance either by chelating metal ions or by scavenging heavy metal stress induced reactive oxygen species [18], [19], [21].
Owing to lack of any detailed studies, present investigations were initiated with an aim to evaluate impact of Au3+ on growth and development on a leguminous crop, cowpea (Vigna unguiculata). Our results showed for the first time that (i) cowpea posesses excellent potential to withstand Au3+-stress due to the presence of phenolics; and (ii) phenolics released during the course of seed germination and early seedling growth of cowpea play a vital role in detoxification of Au3+ by forming Au-nanoparticles.
Materials and Methods
Seeds of cowpea [Vigna unguiculata (L.) Walp., Fabaceae] were obtained from the local farmers of Haldwani (Uttarakhand, India).
Impact of Au3+
Impact of different concentrations (viz. 0, 0.05, 0.1, 0.25, 0.5 and 1 mM) of Au3+ on seed germination and early seedling growth of cowpea was evaluated using HAuCl4. After washing with 0.1% cetrimide and distilled water, seeds were surface sterilized with 0.1% mercuric chloride for 2 min, rinsed with sterile distilled water and inoculated in autoclaved bottles containing 75 g of uniform sized glass beads with 20 ml test solution under sterile conditions. These bottles were incubated at 25±2°C under a 16/8 h light/dark cycle at a light intensity of 60 µmol m−2 s−1. Growth of seedlings was measured in terms of length and fresh weight of root and shoot. Levels of malondialdehyde (MDA) and proline in root and shoot were measured in 4 d old seedlings.
Estimation of Proline and Malondialdehyde
For determining levels of proline and malondialdehyde (MDA), root and shoot of seedlings were homogenized in 5% TCA and centrifuged at 15,000 xg for 15 min. Proline levels were measured according to Bates et al. [22]. MDA levels were determined following the protocol of Heath and Packer [23]. MDA and proline levels were expressed in nmoles or µmoles g−1 fresh weight.
Estimation of Total Phenolic Content
Total phenolics were measured as per Ainsworth and Gillespie [24] using Folin-Ciocalteau reagent and expressed in terms of nmoles of gallic acid equivalents (GAE).
Contribution of Different Components of Cowpea to Generate Au-nanoparticles
Different components namely seed coat and cotyledons were excised carefully from 4 d old cowpea seedlings raised in distilled water under sterile conditions. Seed coat, cotyledons and rest of the seedlings (i.e. seedlings devoid of cotyledons+seed coat) were treated with 10 ml of different concentrations of sterile HAuCl4 for 24 h to test their potential to generate Au-nanoparticles. Seedlings devoid of cotyledons+seed coat were treated by immersing their roots in HAuCl4 solutions. Contol incubation medium (i.e. distilled water), in which seedlings of cowpea were raised for 4 d, was also tested for its efficacy to form Au-nanoparticles by incubating 500 µl of this medium with 10 ml HAuCl4.
Characterization of Au-nanoparticles
UV-Vis spectra of Au3+ solutions (i) in which seedlings of cowpea were raised; and (ii) incubated independently with various components of seedlings of cowpea and distilled water in which seedlings were raised as detailed above, were recorded from 190 to 1100 nm using Specord 200 Analytikjena UV-Vis spectrophotometer. For transmission electron microscopic (TEM) studies, 10 µl of colloidal suspension was drop-coated on 200 mesh copper grid with an ultrathin continuous carbon film and allowed to dry in a desiccator at room temperature. Grids were viewed in the transmission electron microscope (Technai G2 T30) at a voltage of 300 kV. The hardware associated with the machine also allowed (i) energy dispersive X-ray (EDX) analysis to measure the elemental composition; and (ii) selected area electron diffraction (SAED) analysis to determine crystalline/amorphous nature, of nanoparticles.
Powder X-ray Diffraction Studies
For powder X-ray diffraction (PXRD) studies, colloidal suspensions obtained by incubating different components with Au3+ solutions were centrifuged. The pellet obtained was re-suspended in distilled water, drop coated on silica surface, dried in desiccator and used for collecting PXRD pattern using Rigaku Rotaflex RAD-B with copper target CuK(α)1 radiation with tube voltage 40 kV and 60 mA in 2 theta (θ) range of 30–80°.
Stasistical Analysis
All experiments were carried out independently at least seven times. Duncan's multiple range test was used to to determine the level of significance in physiological and biochemical data [25].
Results and Discussion
Impact of Au3+ on Seedling Growth
While evaluating impact of Au-nanoparticles on seed germination and early seedling growth of cowpea, HAuCl4 was used as control. To our surprise, in spite of being a heavy metal ion, Au3+ was not associated with any significant alteration in seedling growth (measured in terms of length and fresh weight of root and shoot) of cowpea even when present at a concentration of 1 mM (Figure 1). It is well documented that heavy metal ions such as Cd+, Zn2+, Co2+ and Pb2+ inhibit plant growth and development [26]. In general, crop plants exposed to heavy metal stress show enhanced levels of stress markers, proline (an imino acid) and MDA (a cytotoxic byproduct of lipid peroxidation), concomitant with suppression in growth [27], [28]. Heavy metal stress induced enhancement in MDA levels is due to lipid peroxidation by reactive oxygen species (ROS) that are generated due to suppression in electron transport system and/or promotion of Fenton's reaction [29]. Increase in level of proline under heavy metal stress is linked to (i) its synthesis to ensure appropriate recycling of NAD(P)+ for cellular metabolism including its role as terminal acceptor of light mediated photosynthetic electron transport [29], [30]; (ii) its role in scavenging ROS [31]; and (iii) protection of enzymes and other macromolecular structures/complexes [32]. Astonishingly, seedlings of cowpea raised in Au3+, even at concentration as high as 1 mM, did not show any enhancement in levels of proline and MDA, which is in synchronization with unaltered growth (Figure 1). These findings depicted that cowpea possessesremarkable potential to tolerate Au3+ by some unique mechanism.
Figure 1.

Seedlings of cowpea raised in varying concentrations (mM) of Au3+ (a). Length [(b),(c)], fresh weight [(d),(e)], proline [(f),(g)] and MDA levels [(h),(i)] of root (yellow bars) and shoot (green bars) of cowpea seedlings raised in Au3+. Absorption spectra (j) and TEM images [(k),(l)] of Au3+ solutions in which seedlings were raised. Values represent mean of data collected from seven independent experiments and vertical lines on bars represent standard error [n = 35 for (b), (c), (d) & (e); n = 7 for (f), (g), (h) & (i)]. Values designated by different small letters above bars are significantly different at P≤0.05 (Duncan's multiple range test).
Unexpectedly, clear pale yellow Au3+ solutions in which seedlings of cowpea were raised, turned colloidal purple. Such an alteration in color of Au3+ solution (from clear pale yellow to colloidal purple) is due to generation of Au-nanoparticles [33], [34]. Absorption spectra of these colloidal purple suspensions showed distinct peak at ∼550 nm which is well documented to arise due to surface plasmon resonance of Au-nanoparticles [33], [34]. In general, intensity of the purple color and Au-nanoparticle specific absorbance peak of these suspensions increased with increase in concentration of Au3+ (Figure 1). Occasionally, an additional peak in infra red region was recorded in absorption spectra of 1 mM Au3+ solution in which seedlings of cowpea were raised. At present, it is difficult to presume reasons behind the appearance of this IR peak in only 1 mM Au3+ and not other concentrations of Au3+ in which seedlings of cowpea were raised. Transmission electron microscopic investigations confirmed presence of distinct crystalline nanoparticles in range of 20–50 nm in these purple colloidal suspensions (Figure 1). It is known that ionic speciation state of metals is more toxic compared to nanoparticle speciation state [35]. Therefore, we hypothesize that the potential of cowpea to withstand Au3+-stress is linked to its inbuilt potential to generate Au-nanoparticles.
Hypothesizing the Involvemenmt of Phenolics in Imparting Au3+-Stress Tolerance
Control incubation medium (i.e. distilled water) turned brown during the course of seed germination and early seedling growth. It is known that germinating legume seeds release phenolics [36], which impart brown coloration to the incubation medium. Interestingly, each seedling of cowpea released ∼35 nmoles GAE of phenolics during the course of seed germination and early seedling growth. It is well documented that phenolics such as gallic acid, catechin promote generation of Au-nanoparticles [37], [38]. This prompted us to believe that phenolics released by germinating seeds of cowpea could be responsible for generation of Au-nanoparticles. Owing to electron donating capacity of phenolics [21], we believe that phenolics released in large quantities during seed germination and early seedling growth must have reduced Au3+ and promoted formation of Au-nanoparticles. This seems to be an ideal Au3+-tolerance mechanism exhibited by cowpea, wherein toxic Au3+ is converted to less/non-toxic Au-nanoparticles by phenolics released during seed germination and early seedling growth.
Identifying the Key Component(s) Involved in Generation of Au-nanoparticles
To identify the key component(s) responsible for formation of Au-nanoparticles and imparting Au3+-stress tolerance to cowpea seedlings, four different components namely (i) control incubation medium (i.e. distilled water) in which seedlings were raised; (ii) seed coat; (iii) cotyledons; and (iv) seedlings devoid of cotyledons+seed coat, were tested for their efficacy to generate Au-nanoparticles by independently incubating them in different levels of Au3+. As anticipated, brown colored incubation medium possessed abundant potential to turn pale yellow Au3+ solutions purple indicating the generation of Au-nanoparticles (Figure 2). Of the three components of 4 d old seedlings (viz. seed coat, cotyledons and seedlings devoid of cotyledons+seed coat), seed coat possessed maximum potential to turn pale yellow Au3+ solutions purple (Figure 2). Accordingly, Au-nanoparticle specific plasmon resonance band in absorprion spectra of purple suspensions formed by (i) incubation medium in which seedlings were raised and (ii) seed coat, was more intense compared to those formed by cotyledons and seedlings devoid of cotyledons+seed coat.
Figure 2.

Color (a–d) and absorption spectra (e–f) of Au3+ solutions (mM) incubated for 24 h with incubation medium in which cowpea seedlings were raised (a,e); seed coat (b,f); cotyledons (c,g); and roots of intact cowpea seedlings (devoid of cotyledons+seed coat) immersed (d,h).
Transmission electron microscopic investigations confirmed presence of nanoparticles in purple colloidal suspensions formed independently by all four components when incubated with Au3+ solutions. However, the size and morphology of nanoparticles varied depending on the component. Seed coat and incubation medium generated nanoparticles in range of 10–30 nm, while cotyledons and seedlings devoid of cotyledons+seed coat generated nanoparticles in the range of 10–25 and 5–10 nm, repectively (Figure 3). These results indicated that the mechanism of generation of nanoparticles in the former two cases is similar, but, vary distinctly from the latter two cases. However, irrespective of the component responsible for generation of Au-nanoparticles, EDX spectra showed two prominent peaks of Au confirming that these nanoparticles were composed of Au (Figure 3). The prominent peaks of Cu and C seen in these EDX spectra arose from carbon coated copper grids, on which samples were loaded. Irrespective of the component responsible for generation of Au-nanoparticles, the nanoparticles were crystalline as revealed by SAED pattern (Figure 3). Similarly, PXRD analysis showed that Au-nanoparticles formed in all cases were crystalline and had face centered cubic structure corresponding to (111), (200), (220), and (311) gold crystalline facets which matched with the JCPDS (Joint Committee on Powder Diffraction Studies) File No. 04–0784 (Figure 4).
Figure 3.

TEM images (a–d,i–l), SAED pattern (e,g,m,o) and EDX spectra (f,h,n,p) of Au-nanoparticles in Au3+ solutions incubated with incubation medium in which cowpea seedlings were raised (a,b,e,f); seed coat (c,d,g,h); cotyledons (i,j,m,n); and the roots of intact cowpea seedlings (devoid of cotyledons+seed coat) immersed (k,l,o,p).
Figure 4.
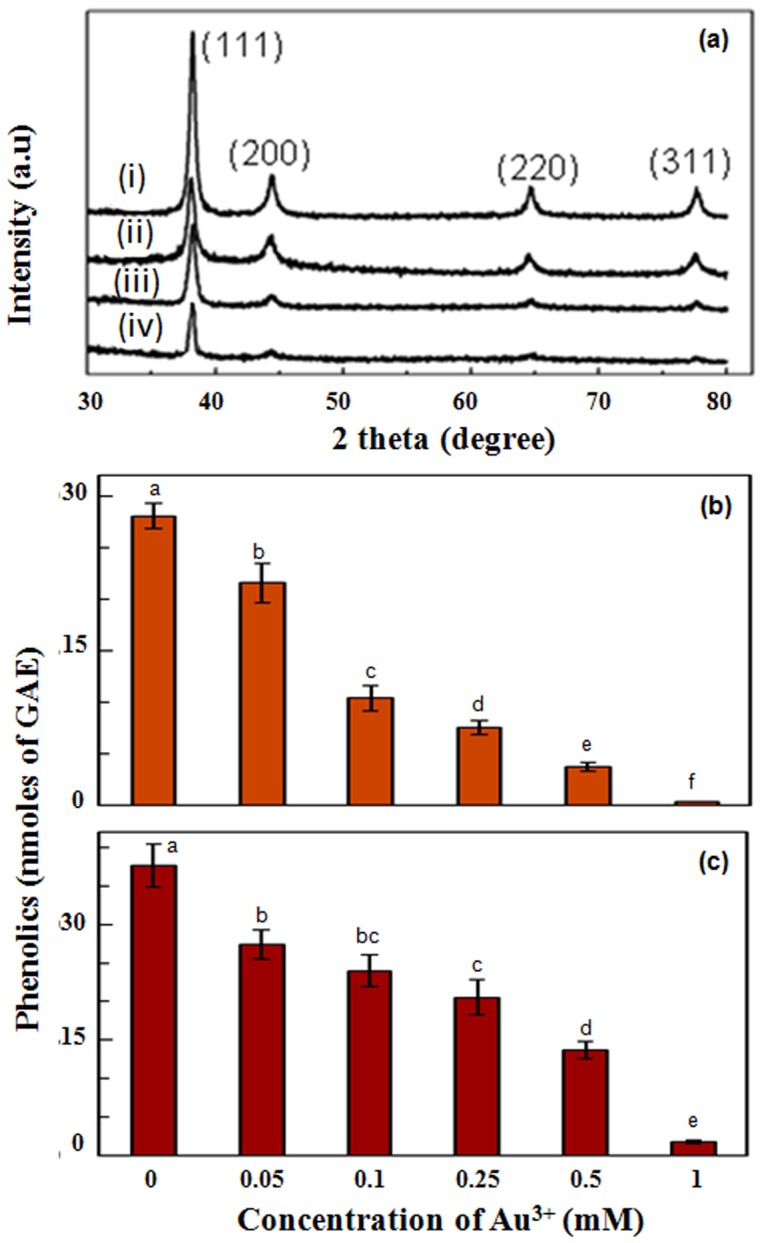
PXRD pattern of Au-nanoparticles (a) in Au3+ solutions incubated with incubation medium in which cowpea seedlings were raised (i); seed coat (ii); cotyledons (iii); and roots of intact cowpea seedlings (devoid of cotyledons+seed coat) immersed (iv); Phenolic content [(b),(c)] in Au3+ solutions (mM) incubated with incubation medium (b); and seed coat (c). Values represent mean of data collected from seven independent experiments and vertical lines on bars represent standard error (n = 7). Values designated by different small letters above bars are significantly different at P≤0.05 (Duncan's multiple range test).
Establishing Role of Phenolics in Imparting Au3+-stress Tolerance
Interestingly, as evident from figure 2d, seedlings devoid of cotyledons+seed coat showed significant suppression in growth upon exposure to Au3+ solutions at concentration above 0.1 mM. This is in contrast to the unperturbed growth response shown by seedlings raised in presence of different levels of Au3+ (Figure 1) and seedlings with cotyledons+seed coat incubated in different levels of Au3+ (data not shown). Potential of cowpea seedlings with seed coat to generate 5–6 fold higher level of Au-nanoparticles compared to the seedlings devoid of seed coat is evident from the intensity of Au-nanoparticle specific absorption peak of purple colloidal suspensions formed by them (Figures 1 and 2). This clearly established that seed coat, which is the key source of phenolics play a pivotal role in imparting Au3+ tolerance to cowpea.
Close correlation between potential of seed coat to release large quantity of phenolics with its potential to generate large proportion of Au-nanoparticles, as evident from figure 2, strengthens our hypothesis that phenolics released during seed germination play a vital role in imparting Au3+-stress tolerance through formation of Au-nanopartoicles. This hypothesis was further strengthened by the potential of control incubation medium (distilled water in which seeds were germinated and seedlings raised) that contained large quantity of phenolics to generate large proportion of Au-nanoparticles (Figure 2).
To establish direct role of phenolics in generating Au-nanoparticles, the level of phenolics in Au3+ solutions incubated with seed coat and incubation medium (in which seeds were germinated and seedlings raised) were determined. As evident from figure 4, level of phenolics decreased progressively with increase in concentration of Au3+. This clearly established that phenolics played a critical role in reduction of Au3+ and generation of Au-nanoparticles. As stated earlier, phenolics possess potential to donate electrons to metal ions such as Au3+ and promote synthesis of Au-nanoparticles.
In nutshell, our results convincingly demonstrated that seed coat, being key source of phenolics, is the most powerful component of developing seedling responsible for generation of Au-nanoparticles. In light of above elaborated experimental evidences, we believe that seed coat plays most vital role in imparting Au3+-stress tolerance to cowpea. Earlier researchers had reported that seed coat plays important role in chemical protection from oxidative damage as it possesses phenolics which act as antioxidants [39], [40]. Although, we do not rule out the possible role of phenolics in acting as antioxidants as has been established by these researchers, our results convincingly demonstrated for the first time that phenolics released by seed coat impart Au3+-stress tolerance by rapidly converting toxic ionic speciation state into less/non-toxic nanoparticle speciation state.
Conclusions
Our above findings demonstrated for the first time that cowpea has substantial potential to withstand Au3+-stress during seed germination and early seedling growth. Our results established that (i) the potential of cowpea to withstand Au3+-stress is linked to phenolics; (ii) seed coat is the key source of phenolics during seed germination and early seedling growth and hence is important for imparting Au3+-stress tolerance; and (iii) phenolics impart Au3+-stress tolerance by converting toxic ionic speciation state of Au to less/non-toxic nanoparticle speciation state. Owing to the proven role of phenolics in imparting metal ion stresss tolerance, we believe that it is important to understand molecular and genetic basis of modulating the synthesis of phenolics for improving tolerance of agricultural/forestry plant species against metal ion stress. Our findings also furnish a novel, simple, green and economically viable protocol for using seed/seed coat of legume seeds for synthesis of metal nanoparticles.
Acknowledgments
Support rendered by University Science Instrumentation Facility, University of Delhi is duly acknowledged. We are thankful to Mr. Rahul Bhardwaj for providing assistance during TEM analysis and Mr. Harsh Kumar for PXRD studies.
Funding Statement
Financial assistance provided to (i) Nisha Shabnam by University Grants Commission (Govt. of India); and (ii) P. Sharmila by Department of Biotechnology (Govt. of India) under Bio-CARe Women Scientist Scheme is duly acknowledged. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Ishikawa S, Suyama K, Arihara K, Itoh M (2002) Uptake and recovery of gold ions from electroplating wastes by using egg shell membrane. Biores Technol 81: 201–206. [DOI] [PubMed] [Google Scholar]
- 2. Baba AA, Adekola FA, Ojutemieden DO, Dada FK (2011) Solvent extraction of gold from hydrochloric acid-leached Nigerian gold ore by tributylphosphate. Chem Bull 1: 1–9. [Google Scholar]
- 3. Syed S (2012) Recovery of gold from secondary sources-A review. Hydrometallurgy 115–116: 30–51. [Google Scholar]
- 4. Ainsworth SK, Swain RP, Watabe N, Brackett NC Jr, Pilia P, et al. (1981) Gold nephropathy, ultrastructural fluorescent and energy-dispersive x-ray microanalysis study. Arch Pathol Lab Med 105: 73–78. [PubMed] [Google Scholar]
- 5. Lengke MF, Southam G (2005) The effect of thiosulfate-oxidizing bacteria on the stability of the gold-thiosulfate complex. Geochim Cosmochim Acta 69: 3759–3772. [Google Scholar]
- 6. Danscher G, Stoltenberg M (2006) Silver enhancement of quantum dots resulting from (1) metabolism of toxic metals in animals and humans, (2) in vivo, in vitro and immersion created zinc–sulphur/zinc–selenium nanocrystals, (3) metal ions liberated from metal implants and particles. Prog Histochem Cytochem 41: 57–139. [DOI] [PubMed] [Google Scholar]
- 7. Anderson CWN, Brooks RR, Stewart RB, Simcock R (1998) Harvesting a crop of gold in plants. Nature 395: 553–554. [Google Scholar]
- 8. Beattie IR, Haverkamp RG (2011) Silver and gold nanoparticles in plants: sites for the reduction to metal. Metallomics 3: 628–632. [DOI] [PubMed] [Google Scholar]
- 9. Sharma NC, Sahi SV, Nath S, Parsons JG, Gardea-Torresday JL, et al. (2007) Synthesis of plant-mediated gold nanoparticles and catalytic role of biomatrix-embedded nanomaterials. Environ Sci Technol 41: 5137–5142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Bali R, Harris AT (2010) Biogenic synthesis of Au nanoparticles using vascular plants. Ind Eng Chem Res 49: 12762–12772. [Google Scholar]
- 11. Maestri E, Marmiroli M, Visioli G, Marmiroli N (2010) Metal tolerance and hyperaccumulation: Costs and trade-offs between traits and environment. Environ Expt Bot 68: 1–13. [Google Scholar]
- 12. Neilands JB (1995) Siderophores: structure and function of microbial iron transport compounds. J Biol Chem 270: 26723–26726. [DOI] [PubMed] [Google Scholar]
- 13. Ma JF, Ryan PR, Delhaize E (2001) Aluminium tolerance in plants and the complexing role of organic acids. Trends Plant Sci 6: 273–271. [DOI] [PubMed] [Google Scholar]
- 14. Wenzl P, Patino GM, Chaves AL, Mayer JE, Rao IM (2001) The high level of aluminum resistance in signalgrass is not associated with known mechanisms of external aluminum detoxification in root apices. Plant Physiol 125: 1473–1484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Bringezu K, Litchenberger O, Leopold I, Neumann D (1999) Heavy metal tolerance of Silene vulgaris . J Plant Physiol 154: 536–546. [Google Scholar]
- 16. Migocka M, Papiernia A, Kosatka E, Klobus G (2011) Comparative study of the active cadmium efflux systems operating at the plasma membrane and tonoplast of cucumber root cells. J Exp Bot 62: 4903–4916. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 17. Cobbett C, Goldsbrough PB (2002) Phytochelatins and metallothioneins: roles in heavy metal detoxification and homeostasis. Annu Rev Plant Biol 53: 159–182. [DOI] [PubMed] [Google Scholar]
- 18. Lavid N, Schwartz A, Yarden O, Tel-Or E (2001) The involvement of polyphenols and peroxidase activities in epidermal glands of water lily (Nymphaeceae). Planta 212: 323–333. [DOI] [PubMed] [Google Scholar]
- 19. Jung C, Maeder V, Funk F, Frey B, Sticher H, et al. (2003) Release of phenols from Lupinus albus L. roots exposed to Cu and their possible role in Cu detoxification. Plant Soil 252: 301–312. [Google Scholar]
- 20. Michalak A (2006) Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Polish J Environ Stud 15: 523–530. [Google Scholar]
- 21. Rice-Evans CA, Miller NJ, Paganga G (1996) Structures-antioxidant activity relationships of flavonoids and phenolic acids. Free Rad Biol Med 20: 933–956. [DOI] [PubMed] [Google Scholar]
- 22. Bates LS, Walderen RD, Taere ID (1973) Rapid determination of free proline for water stress studies. Plant Soil 39: 205–207. [Google Scholar]
- 23. Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplast. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125: 189–198. [DOI] [PubMed] [Google Scholar]
- 24. Ainsworth EA, Gillespie KM (2007) Estimation of total phenolic content and other oxidation substances in plant tissues using Folin- Ciocalteu reagent. Nat Protoc 2: 875–877. [DOI] [PubMed] [Google Scholar]
- 25. Duncan DB (1955) Multiple range and multiple F tests. Biometrics 39: 205–207. [Google Scholar]
- 26. Alia, Pardha Saradhi P (1991) Proline accumulation under heavy metal stress. J Plant Physiol 13: 554–558. [Google Scholar]
- 27. Alia, Prasad KVSK, Pardha Saradhi P (1995) Effect of zinc on free radicals and proline in Brassica and Cajanus . Phytochemistry 39: 45–47. [Google Scholar]
- 28. Pardha Saradhi P, Alia, Arora S, Prasad KVSK (1995) Proline accumulates in plants exposed to UV radiation and protect them against UV induced peroxidation. Biochem Biophys Res Com 209: 1–5. [DOI] [PubMed] [Google Scholar]
- 29. Alia, Pardha Saradhi P (1993) Suppression in mitochondrial electron transport is the prime cause behind stress induced proline accumulation. Biochem Biophys Res Com 193: 54–58. [DOI] [PubMed] [Google Scholar]
- 30.Sharmila P, Anwar F, Sharma KR, Pardha Saradhi P (2008) Management of abiotic stresses in grain legumes through manipulation of genes for compatible solutes. In: Kirti PB (ed) Handbook of New Technologies for Genetic Improvement of Legumes, CRC Press, USA, pp 577–603.
- 31. Alia, Pardha Saradhi P, Mohanty P (1997) Involvement of proline in protecting thylakoid membranes against free radical-induced photodamage. J Photochem Photobiol B: Biology 38: 253–257. [Google Scholar]
- 32. Sivakumar P, Sharmila P, Pardha Saradhi P (1998) Proline suppresses Rubisco activity in higher plants. Biochem Biophys Res Commun. 252: 428–432. [DOI] [PubMed] [Google Scholar]
- 33. Shabnam N, Pardha-Saradhi P (2013) Photosynthetic Electron Transport System Promotes Synthesis of Au-Nanoparticles. PLoS ONE 8(8): e71123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Yamal G, Sharmila P, Rao KS, Pardha-Saradhi P (2013) Yeast Extract Mannitol medium and its constituents promote synthesis of Au nanoparticles. Process Biochem 48: 532–538. [Google Scholar]
- 35. Klaus-Joerger T, Joerger R, Olsson E, Granqvist C-G (2001) Bacteria as workers in the living factory: metal-accumulating bacteria and their potential for materials science. Trends Biotechnol 9: 15–20. [DOI] [PubMed] [Google Scholar]
- 36. Bekkara F, Jay M, Viricel MR, Rome S (1998) Distribution of phenolic compounds within seed and seedlings of two Vicia faba cvs differing in their seed tannin content, and study of their seed and root phenolic exudations. Plant Soil 203: 27–36. [Google Scholar]
- 37. Wang W, Chen Q, Jiang C, Yang D, Liu X, et al. (2007) One-step synthesis of biocompatible gold nanoparticles using gallic acid in the presence of poly-(N-vinyl-2-pyrrolidone). Coll. Surf. Physicochem Eng Asp 301: 73–79. [Google Scholar]
- 38. Nune SK, Chanda N, Shukla R, Katti K, Kulkarni RR, et al. (2009) Green nanotechnology from tea: phytochemicals in tea as building blocks for production of biocompatible gold nanoparticles. J Mat Chem 19: 2912–2920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Duenas M, Hernandez T, Estrella I (2006) Assessment of in vitro antioxidant capacity and seed coat and the cotyledon of legumes in relation to their phenolic contents. Food Chem 98: 95–103. [Google Scholar]
- 40. Siddhuraju P, Becker K (2007) The antioxidant and free radical scavenging activities of processed cowpea (Vigna unguiculata (L.) Walp) seed extracts. Food Chem 101: 10–19. [Google Scholar]