

Abstract
In this review, we mainly focus on zoonotic encephalitides caused by arthropod-borne viruses (arboviruses) of the families Flaviviridae (genus Flavivirus) and Togaviridae (genus Alphavirus) that are important in both humans and domestic animals. Specifically, we will focus on alphaviruses (Eastern equine encephalitis virus, Western equine encephalitis virus, Venezuelan equine encephalitis virus) and flaviviruses (Japanese encephalitis virus and West Nile virus). Most of these viruses were originally found in tropical regions such as Africa and South America or in some regions in Asia. However, they have dispersed widely and currently cause diseases around the world. Global warming, increasing urbanization and population size in tropical regions, faster transportation and rapid spread of arthropod vectors contribute in continuous spreading of arboviruses into new geographic areas causing reemerging or resurging diseases. Most of the reemerging arboviruses also have emerged as zoonotic disease agents and created major public health issues and disease epidemics.
Keywords: Arthropod-borne viruses, Arboviruses, Alphaviruses, Flaviviruses, Equine Encephalomyelitis, Encephalitis, Zoonoses
Introduction
"Zoonosis" is defined as a disease or infection that is naturally transmitted from vertebrate animals to humans [1]. Zoonotic diseases can be either transmitted via direct or indirect contact. Transmission of an infectious agent from a vertebrate animal to a human by an arthropod vector is an example of indirect transmission of zoonotic disease. Viruses that maintain transmission cycles between vertebrate animal reservoirs as main amplifying hosts and insects as primary vectors are known as arboviruses (arthropod-borne viruses). Arboviruses must replicate in the arthropod vectors, such as mosquitoes, ticks, midges or sandflies, prior to transmission. Female mosquitoes acquire virus during blood feeding of an infected animal and the virus replicates in the mesenteronal epithelial cells. The virus released from the mesenteronal epithelial cells infects salivary glands after secondary amplification in other cells and tissues. Some arboviruses can infect the salivary glands without secondary amplification in other cells and tissues. Subsequently, the virus released from salivary gland epithelium is transmitted during blood feeding of the vertebrate host. Arboviruses are included in different taxonomic families, including Flaviviridae (genus Flavivirus), Bunyaviridae (genus Nairovirus, Orthobunyavirus, Phlebovirus, and Tospovirus), Togaviridae (genus Alphavirus), Rhabdoviridae (genus Vesiculovirus), Orthomyxoviridae (genus Thogotovirus), and Reoviridae (genus Orbivirus and Coltivirus) (Fig. 1). Many of the important zoonotic arboviruses belong to the families Togaviridae and Flaviviridae [2]. However, there are many other clinically important human and animal arboviruses belonging to the Bunyaviridae family, such as Crimean-Congo hemorrhagic fever virus (tick-borne) in the genus Nairovirus [3] and Toscana virus (sandfly-borne) and Rift Valley fever virus (mosquito-borne) in the genus Phlebovirus [4]. Colorado tick fever virus in the family Reoviridae (genus Coltivirus) is also an important human arbovirus [5-7].
Fig. 1.

Classification of arboviruses. Arboviruses are included in six different taxonomic virus families. a)Arboviruses that cause human encephalitides belong to four genera in four virus families.
Arboviruses are maintained in complex life cycles involving nonhuman primate/vertebrate hosts and primary arthropod vectors (Fig. 2). Mosquitoes are the most important vectors that transmit zoonotic viruses. Different mosquito species (Culex spp., Aedes spp., etc.) may act as vectors for the same virus in different vertebrate hosts depending on different geographical and ecological locations. Ticks, sandflies (Phlebotomus spp.) and gnats (Culicoides spp.) are also important vectors of some arboviruses. Vertical transmission (transovarial and transstadial) occurs in some arthropod vectors as they transmit some arboviruses from parent arthropod to offspring arthropods (Fig. 3). This type of transmission mainly occurs in tick-borne encephalitis viruses (TBEVs) but it has been also reported in some mosquito-borne viruses [8-10]. For example, La Crosse virus, one of the most important viruses among agents causing California encephalitis, is transmitted by its main vector, Aedes triseriatus, not only by transovarial and transstadial routes but also sexually [11].
Fig. 2.

Vertebrate host and vector transmission cycles. (A) Enzootic cycle (sylvatic or jungle cycle). The natural transmission of virus between wild animals (vertebrate hosts) and primary or enzootic insect vectors and that leads to the amplification of the virus in the vector. The vertebrate host is the reservoir host that can harbor a virus indefinitely with no ill effects. Therefore, reservoir host is the primary host of a virus and may be re-infected several times during their life. (B) Epizootic cycle (rural cycle). The virus is transmitted between non-wild or domestic animals and the primary or accessory insect vectors. This can lead to an epidemic outbreak of viral disease in a domestic animal population where the virus is amplified (amplifying host), often with the implication that it may extend to humans by insect vectors (e.g., Japanese encephalitis virus, Venezuelan equine encephalitis virus [VEEV]). (C) Urban cycle. Humans are the source of infection for mosquitoes due to high level of viremia. The virus cycles between humans and insect vectors (urban vector e.g., A. aegypti) repeatedly, as reinfection occurs with every new insect bite (e.g.,dengue virus, yellow fever virus, St. Louis encephalitis virus, VEEV, chikungunya virus, Rift Valley fever virus). (D) Humans are dead-end hosts in the infection chain and do not develop sufficient viremia and do not serve for amplification of the virus to be transmitted again to insect vector (e.g., Eastern equine encephalitis virus, Western equine encephalitis virus, West Nile virus, and Sindbis virus). a)Amplifying host is in which the level of virus can become high enough that an insect vector such as a mosquito that feeds on it will probably become infectious. b)Dead-end host or incidental host is an intermediate host that does generally not allow transmission of the virus to the definitive host. They do not develop sufficient viremia to be picked up by the insect vectors. c)Bridge vector is an arthropod that acquires virus from an infected wild animal and subsequently transmits the agent to human or secondary host.
Fig. 3.

Vertical transmission cycle (transovarial and transstadial). A vertical transmission exists in some arthropod vectors and is epidemiologically important. This type of transmission is found in viruses that belong to tick-borne encephalitis complex. However, it is also found in some of the mosquito-transmitted viruses (e.g., La Crosse encephalitis, Murray Valley encephalitis, St. Louis encephalitis, Japanese encephalitis,West Nile, and Western equine encephalitis).
Most known arboviruses were first isolated in tropical regions such as Africa and South America and in some Asian countries. However, the geographic distribution and frequency of epidemic outbreaks of arboviral diseases have expanded dramatically across the world in the past several decades [12]. Several factors such as changes in viral genetics, host and/or vector population, and climate changes facilitated expansion and transmission of arboviruses resulting in emergence/reemergence of arboviral disease outbreaks in new regions in the world. Extensive tropical urbanization and faster and increased movement of humans and animals with modern transportation helped vectors to be in closer contact with vertebrate reservoir hosts raising transmission potential. Introduction of West Nile virus into the New World and the emergence of Japanese encephalitis virus (JEV) in Australia are a few prominent examples of recent unexpected emerging/reemerging zoonotic diseases [13,14].
Epidemics/epizootics of humans and domestic animals usually occur when the enzootic virus is introduced into rural environments or comes to close contact with humans by a bridge vector (Fig. 2). Usually, humans and domestic animals develop clinical disease but do not develop a sufficient level of viremia to infect arthropods, thus, they are considered dead-end hosts and do not contribute to the transmission cycle [15, 16]. However, some arboviruses such as dengue fever (DF), yellow fever, and chikungunya (CHIKV) viruses cause high levels of viremia in humans and can be transmitted from person to person by mosquitoes (urban cycle) (Fig. 2) [17].
In this review, we will mainly focus on the transmission and epidemiology of mosquito-borne arboviruses, especially alphaviruses and flaviviruses that are pathogenic to humans as well as domestic animals, thus, increasing the public health and economic significance. Although these viruses, with the exception of JEV, are not currently circulating in the Korean peninsula, there is a great chance for other viruses to emerge when a competent vector and vertebrate host populations happen to be temporally and spatially together in a permissive environment.
Alphaviruses
Alphaviruses (formerly, group A arboviruses) are enveloped, positive-sense, single-stranded RNA viruses that belong to the genus Alphavirus in the family Togaviridae. The alphavirus genome varies between 11 and 12 kb in length and is composed of a non-segmented, single-strand RNA with a 7-methylguanosine and a poly A tail at 5'- and 3'-terminus, respectively [18,19]. It encodes four non-structural proteins responsible for genome replication and protein processing, and generates a subgenomic mRNA (26S). The five structural proteins (C, E3, E2, 6K, and E1) are translated from the 26S subgenomic mRNA [18,20].
The alphaviruses are widely distributed throughout the world. They have been classified as belonging to either New and Old World alphaviruses: New World alphaviruses (e.g., Eastern equine encephalitis virus [EEEV], Western equine encephalitis virus [WEEV], and Venezuelan equine encephalitis virus [VEEV]) are distributed across the Americas and cause encephalitis in humans, whereas Old World alphaviruses (e.g., Sindbis virus [SINV], CHIKV, O'nyong-nyong virus [ONNV], Ross River virus [RRV], Barmah Forest virus [BFV], and Semliki Forest virus [SFV]), characterized by fever, rash, and arthritis, are found in Europe, Asia, Australia, and parts of Africa [21,22]. However RRV, SINV, and CHIKV have been occasionally associated with encephalitis. The alphavirus serogroups can be divided into seven antigenically related complexes: Barmah Forest, Eastern equine encephalitis (EEE), Middleburg, Ndumu, Semliki Forest, Venezuelan equine encephalitis (VEE), and Western equine encephalitis (WEE) [21,23-25]. All clinically relevant alphaviruses are transmitted by mosquitoes. More than one mosquito species is usually involved in the alphavirus transmission cycle. The survival of alphaviruses in a certain geographic region depends on the presence of competent vectors (mosquitoes) and of vertebrate hosts that develop viremic infection with low pathogenicity. Important amplification hosts are birds (for SINV, SFV, EEEV, and WEEV), rodents (for RRV, VEEV, BFV), and monkeys (for CHIKV, ONNV, and Mayaro fever virus) [26].
Eastern equine encephalitis virus
EEEV is a zoonotic virus transmitted by mosquitoes and originating in birds [27]. In North America, EEEV is an important cause of disease in domestic animals and humans. The disease is severe in horses, pigs, dogs, and some species of birds. EEEV was first isolated in 1933 from the brains of affected horses during a widespread outbreak in the northeastern US, in New Jersey and Virginia [27,28]. However, horse deaths have recently been reported further north along the eastern coast of the US (New Hampshire and Maine) and Canada. In 1936, South American EEEV was first isolated from a horse in Argentina [29,30]. The EEEV strains present in South and North America are antigenically and genetically different from each other and also differ in human pathogenicity [31,32]. There are four lineages (I, II, III, and IV) of EEEV based on their antigenicity and distribution in various geographic regions (Table 1) [33,34]. The closely related North American EEEV (NA-EEEV) lineage I viruses that occur in the US, Canada and the Caribbean are the most virulent to horses and humans. In contrast, infection of horses or humans with more genetically and antigenically diverse virus strain, enzootic in Central and South America (lineages II, III, and IV [SA-EEEV]), rarely results in significant clinical disease. The lineage II strains are distributed along the coasts of South and Central America, lineage III in the Amazon Basin, and lineage IV in Brazil [34,35].
Table 1.
A list of important alphaviruses
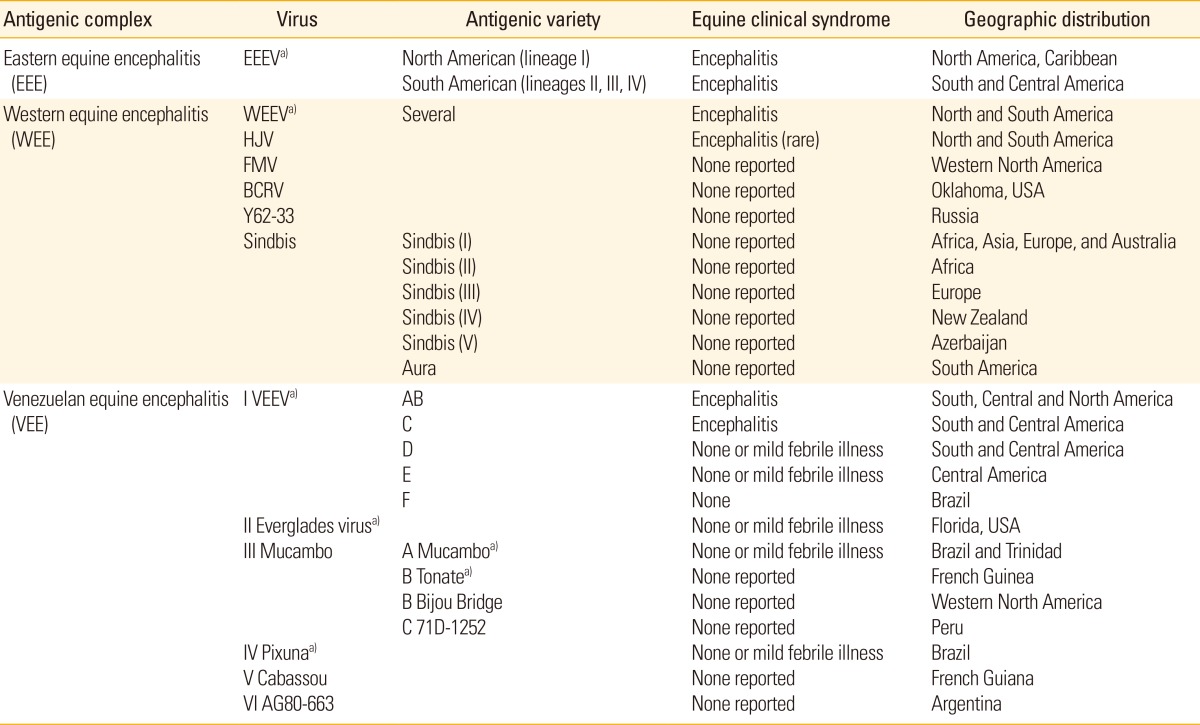
ArgentinaAdapted from Weaver et al. [33], with permission from Elsevier Ltd.
EEEV, Eastern equine encephalitis virus; WEEV, Western equine encephalitis virus; HJV, Highland J virus; FMV, Fort Morgan virus; BCRV, Buggy Creek virus; VEEV, Venezuelan equine encephalitis virus.
a)Encephalitis and febrile illness in humans.
In North America, EEEV is enzootic from the eastern and gulf coasts as far as to inland sites (Texas) [36,37]. NA-EEEV strains are genetically highly conserved, with only one major lineage (lineage I) from the first isolation in 1933 [38]. Most EEE outbreaks in North America occur in the late summer and early fall, often associated with heavy rainfall. Outbreaks in horses are common and often accompanied by high case-fatality rates. Eighty to 90% of the infected horses develop acute and lethal disease, and about 66% of the survivors develop severe neurologic sequelae [27]. During outbreaks or epidemics of EEE, horses do not serve as amplifying hosts but they tend to be the first to develop clinical signs and often serve as an indicator of the start of an outbreak or epidemic. Thus, the rapid detection of EEEV in equine specimens is critical for control of disease outbreaks in humans, horses, and other animal species. The NA-EEEV strains are responsible for most human cases [31]. Human infections are usually asymptomatic, but some progress to severe encephalitis accompanied by high fatality rate or incapacitating sequelae. The disease is generally more severe in the elderly and infants [5,39]. Although only few cases of human EEEV infection have been reported annually since the 1960s, the high mortality rate and severe neurologic sequelae in infected patients make EEEV an important human pathogen.
In South America, enzootic EEEV is widely distributed in most areas of tropical forests, in the Amazon Basin in Brazil, and in Northern Argentina [40]. In these regions, EEEV is principally an equine pathogen and equine cases can occur year-round. However, human EEEV infections were rarely detected even during major equine epizootics [41-43]. In temperate regions of South America (e.g., Argentina), EEEV infections often occur during the summer [44].
EEEV transmission cycle
The EEEV transmission cycle in North America is maintained between the passerine birds as reservoir/amplification hosts and ornithophilic mosquito, Culiseta melanura, as the main enzootic vector in swamp habitats (Fig. 4) [27]. In addition, studies have shown that C. melanura, regarded as a bridge vector in human and equine infections so far, may also serve as a main epizootic vector as well [45,46]. Mosquito species such as C. peccator, C. erraticus, and Uranotaenia sapphirina may also serve as enzootic vectors in some regions of the southeastern US [47]. These mosquitoes are known to feed on reptiles and amphibians. Recently, snakes have been suggested to play a role in the enzootic EEEV transmission cycle as over-wintering hosts [48-50]. EEEV infections in birds are usually asymptomatic; however, disease with high-titered viremia and high mortality rate has been reported in chukar partridges, pheasants, egrets, glossy ibises (Plegadis falcinellus), rock doves, house sparrows, psittacine birds, ratites (emus, ostriches), African penguins, chicken (<14 days old), pigeons, Pekin ducks, and whooping cranes [51-55]. Passerine birds develop extremely high levels of viremia, enough to infect both enzootic vectors as well as a variety of bridge vectors (e.g., Aedes and Coquillettidia) that transmit the virus from enzootic cycle to humans and horses [27]. In pheasants, EEEV is transmitted through feather picking and cannibalism. Humans and equids are dead-end hosts since they do not develop sufficient viremia to transmit the virus [10].
Fig. 4.

Enzootic and epizootic/epidemic transmission cycles of Eastern equine encephalitis virus (EEEV). The enzootic EEEV transmission cycle is maintained between passerine birds as reservoir/amplification hosts and Culiseta melanura, as the main enzootic vector in swamp habitats. Rodents/marsupials may serve as principal enzootic vectors and reservoirs in South America. Passerine birds develop extremely high levels of viremia, enough to infect both enzootic vectors as well as a variety of bridge vectors. Humans and equids are dead-end hosts since they do not develop sufficient viremia to transmit the virus.
In South America, reservoirs and amplification hosts involved in the enzootic EEEV transmission cycle are not known yet. However, seroprevalence and experimental studies suggest thatthe Culex (Melanoconion) subgenus and rodents/marsupials may serve as principal enzootic vectors and reservoirs, respectively, and they may play a more important role in enzootic EEEV transmission in South America [56]. The virus mainly causes disease in horses and occasional cases of encephalitis have also been reported in sheep, cattle, deer, South American camelids (llamas and alpacas), and pigs [57,58]. In addition, infections have been seen in dogs, goats, bats, and small mammals including rodents [59-62].
EEEV vaccine
There is a formalin-inactivated vaccine based on an NA-EEEV strain (PE-6) used in horses and emus, however it does not induce significant neutralizing anti-E2 antibody to SA-EEEV [63]. The vaccine is used in laboratory workers to protect from accidental exposure. A similar formalin-inactivated vaccine is available for horses. There is no specific therapy for EEE at the moment.
Western equine encephalitis virus
WEEV is genetically diverse and both epizootic and enzootic strains have been identified. Epizootic North American strains are more virulent than strains that are enzootic in South America (sporadic cases of WEE). WEE was the first equine encephalitic arbovirus identified in North America. It is closely related to Sindbis and SFV since it emerged from a recombination of viruses in the EEE and the Sindbis lineages [64,65]. WEEV was first isolated from brains of affected horses during an equine epizootic outbreak in the San Joaquin Valley of California in 1930 [66]. In 1938, the first lethal human infection of WEEV was confirmed [67] and since then it spread to the west of North America and the American Midwest with periodic equine epizootics and epidemics [68,69]. Epidemiological studies have shown that WEEV occurs throughout most of the Americas from the western half of North to South America, including Guyana, Ecuador, Brazil, Uruguay, and Argentina [65]. In South America, with the exception of Argentina, only small equine epizootics, but no human WEE cases have been reported [29].
WEEV continues to cause equine encephalitis in northern South America and Central America with occasional outbreaks in Florida and the southwestern US, but only a few human cases of WEE have been reported, with low fatality rate, in the past several decades in North America [70]. Most WEEV infections in humans and equines occur in summer, June and July, and slightly later in temperate regions like Canada. Although most human cases of WEE are asymptomatic, infants and children are highly susceptible to WEEV infection and are most likely to develop severe encephalitis. Clinical manifestations develop after 2 to 10 days of incubation and are characterized by nonspecific febrile viremia, malaise, and headache often in association with meningismus. The case fatality rate in humans is about 3% to 4% [71]. The case fatality rate in horses is 20% to 30%, but can be up to 50% in some epidemics [72]. For horses, WEEV is less virulent than EEEV.
In addition, Highland J virus (HJV), Fort Morgan virus (FMV), and related Buggy Creek virus (BCRV), distinct but closely related to WEEV, were also isolated in North America [33]. HJV has been identified in the eastern US (Florida) and is transmitted from C. melanura mosquitoes to songbirds in freshwater swamps [73,74]. It has a low pathogenicity in mammals and is rarely seen in humans or horses. Exposure to HJV has not been directly associated with human illness. However, HJV can cause sporadic encephalitis in horses, and is also pathogenic to turkeys and partridges [53,75-78]. Similar to WEEV, BCRV is a natural recombinant virus derived from Old World SINV and New World EEEV [65]. BCRV (and the closely related FMV) is apparently widely distributed in North America, having been found in Texas, Oklahoma, Nebraska, Colorado, South Dakota, and Washington State. It was first isolated in 1980 at Buggy Creek in Grady County, Oklahoma. However, the ecologically very similar FMV was discovered in the 1970s in Colorado [79]. BCRV is commonly associated with the cimicid swallow bug (Oeciacus vicarius) [79]. The bug is an ectoparasite of the colonially nesting cliff swallow (Petrochelidon pyrrhonota) and, to a lesser extent, the house sparrow (Passer domesticus), with both birds serving as hosts to BCRV. FMV is also associated with swallow bugs, cliff swallows, and house sparrows. These two viruses are pathogenic to swallows but not to humans or horses. These four viruses in North America (WEEV, BCRV, FMV, and HJV), the Aura virus in South America, and SINV with its four subtypes found in Africa, Asia, Australia, and Europe, are regarded as members of the WEE complex [80].
WEEV transmission cycle
WEEV is maintained in an enzootic cycle between passerine birds as reservoirs and its specific mosquito vector, Culex tarsalis, abundant in agricultural settings in the western US. Domestic and wild birds are considered important reservoir and epizootic amplifying hosts (Fig. 5). It has been also suggested that lagomorphs and rodents can serve as amplification hosts when they are infected with WEEV by Aedes mosquitoes [10, 44,79].
Fig. 5.

Enzootic and epizootic/epidemic transmission cycles of Western equine encephalitis virus (WEEV). WEEV is maintained in an enzootic cycle between passerine birds as reservoirs and its specific mosquito vector, C. tarsalis. Domestic and wild birds are considered important reservoir and epizootic amplifying hosts. It has also been suggested that lagomorphs and rodents can serve as amplification hosts when they are infected with WEEV by Aedes mosquitos.
Epizootic transmission to horses and humans is mediated by bridge vectors, such as Ochlerotatus melanimon in California, Aedes dorsalis in Utah and New Mexico and A. campestris in New Mexico. The seasonal continuation of the natural WEEV transmission cycle in temperate regions is not clear. However, the annual reintroduction of migratory birds and vertical transmission among A. dorsalis mosquitoes are suspected for the maintenance mechanism in temperate regions [81].
WEEV vaccine
Formalin-inactivated vaccines have been developed experimentally for the protection of laboratory workers and other persons at high risk [82]. There is a formalin-inactivated vaccine that is available as a double vaccine in combination with EEEV only for veterinary use (horses) [83]. Horses are vaccinated twice a year due to low immunogenicity of the inactivated vaccine. Presence of neutralizing antibodies is used as a correlate of protection and to monitor the success of immunization.
Venezuelan equine encephalitis virus
Of the three equine encephalitis alphaviruses, VEEV is the most important zoonotic pathogen, with several reported outbreaks in South and Central America [33,84-89] and some of the Central American and Mexican outbreaks have spread into southern Texas in 1971 [90,91]. The VEEV was first isolated from brains of affected horses during an equine encephalitis outbreak that occurred in central Colombia and Venezuela in 1936 [92]. The VEEV antigenic complex is divided into six distinct antigenic subtypes (I-VI) (Table 1) [88,93]. The "epizootic subtypes" of VEEV (types IAB and IC), responsible for the large outbreaks of encephalitis in horses in the Western hemisphere in the last 25 years, are believed to originate from subtype ID strains with an adaptive mutation in the E2 envelope glycoprotein gene [94]. This mutation increases the ability to infect the epidemic mosquito vector, Aedes taeniorhynchus and replication in equids [95,96]. The "enzootic subtypes" of VEEV are considered to be of low pathogenicity for equids under most circumstances. The enzootic subtypes of VEEV include ID, IE, and IF strains from South and Central America, Central America and Brazil, respectively, and type II (Everglades) virus circulating in Florida. Phylogenetic studies indicate that the epidemic subtypes IAB and IC strains emerged from one of six subtypes of ID VEEV lineages, but there was no epidemic strains emerged from IE lineages [86,97-99]. This epidemic-progenitor subtype ID lineage occurs in western Venezuela, Colombia, and the Amazon areas of Ecuador and Peru [84, 85,100]. Enzootic cycles of VEEV subtypes ID and IE are dispersed from southern Florida to northern Argentina [88,101].
Approximately 150 animal species may become infected under natural conditions and may remain subclinical. Horses are most severely affected by VEE infection and half of infected cases (50%) develop encephalitis. However, a subclinical to mild course of the disease is possible. Dogs and pigs can become clinically ill and infection in dogs with epizootic VEEV can be fatal [101].
Infected patients manifest "flu-like" clinical signs (e.g., fever, severe headache, myalgia) after 1- to 4-days of incubation. Most clinical cases are self-limited illnesses with recovery after about a week [25,101]. A subset of symptomatic cases can progress to neurologic disease with convulsions, drowsiness, and disorientation and sometimes followed by incapacitating sequelae (e.g., paralysis, epilepsy, and tremor) [102,103]. Signs of encephalitis are rare and are more frequently seen in children (4%) than adults (0.4%) [102]. Thus the disease is less severe in adults than children. The case fatality rate in patients is approximately 20% but it can reach up to 35% in children under 5 years of age. In horses, enzootic and epizootic VEEV strains present a major difference in terms of pathogenesis, but they show similar virulence in humans. Generally, VEE clinical manifestations are indistinguishable from other tropical infectious diseases such as DF; therefore, the number of cases may be underestimated [104].
VEEV transmission cycle
The enzootic transmission cycle of VEEV is maintained among rodents and other vertebrates (e.g., cotton rats, spiny rats, bats, and opossums) as reservoirs and mosquitoes in the subgenus Culex (Melanoconion) as primary vectors (Fig. 6)[88,98]. These enzootic (sylvatic) cycles are limited in distribution, and typically found in humid, tropical forests or swamps throughout the Americas [104-107]. In contrast, epizootic VEEV strains (IAB and IC) are transmitted by several mosquito vectors (e.g., Aedes and Psorophora spp.) abundant in agricultural habitats, to susceptible amplification hosts such as equids [33]. During epizootic or epidemic cycles, equids are efficient amplification hosts with high-titered viremia for mosquito transmission. Thus, infections from horses will be transmitted to humans during epizootics of VEE in horses. In addition, humans, dogs, pigs, cats, cattle, goats, bats, and birds can be infected during an epizootic [26,33]. They develop viremia that can be a source of mosquito infections.
Fig. 6.

Enzootic and epizootic/epidemic transmission cycles of Venezuelan equine encephalitis virus (VEEV). The enzootic transmission cycle of VEEV is maintained among rodents and other vertebrates (e.g., cotton rats, spiny rats, bats, and opossums) as reservoirs and mosquitoes in the subgenus Culex (Melanoconion) as primary vectors. In contrast, epizootic VEEV strains are transmitted by several mosquito vectors (e.g., Aedes and Psorophora spp.) to susceptible amplification hosts, horses. During epizootic or epidemic cycles, horses are efficient amplification hosts with high titered viremia for mosquito transmission.
Epidemiological studies indicate that major human VEEV outbreaks emerge from humans being infected from spillover of enzootic rodent cycles [87,104,108,109]. Moreover, VEEV infected horses can shed virus in body fluids and the virus can spread by direct contact or via aerosols. Although direct transmission between horses, person-to person, or from horses to humans was not confirmed, there are however, reported cases of people infected in laboratories via aerosols [87]. VEEV is also important as a biological weapon due to the ease of growth in large quantities and the high infectivity when aerosolized [110,111]. Transplacental transmission of VEE infections has been observed in horses.
VEEV vaccine
The VEEV TC-83 strain is the only licensed live-attenuated vaccine available to protect horses in endemic regions. This vaccine strain has been developed by passaging the Trinidad donkey (TrD) IA/B strains 83 times in guinea pig heart cells [112]. Genetic characterization of the TC-83 strain indicates that the attenuation phenotype relies on only two point mutations (nucleotide position 3 in 5'-noncoding region and amino acid position 120 in E2 glycoprotein) [113,114]. Currently, there is no licensed human VEEV vaccine, but TC-83 live-attenuated vaccine is used for laboratory workers and military personnel [115].
In efforts to develop a live-attenuated VEEV vaccine for human use [116], the US Army Medical Research Institute of Infectious Diseases in collaboration with the University of North Carolina has introduced mutations into an infectious clone of the TrD IA/B strain [117-120]. The new vaccine candidate, V3526, was shown to be effective in protecting rodent, horses and nonhuman primates against virulent challenge with VEEV IA/B and other related subtypes (e.g., IE, IIA, and IC) [116,120-122].
Flaviviruses
Flaviviruses (formerly group B arboviruses) include agents pathogenic for humans and animals that are transmitted by mosquitoes or ticks [123,124]. Flaviviruses are enveloped viruses containing a single-stranded positive-sense RNA genome of approximately 11 kb in length. The viral genome encodes three structural proteins (capsid [C], premembrane [prM], and envelope [E] proteins) and seven non-structural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) [125]. The genus Flavivirus is comprised of more than sixty-six virus species, many of which are arthropod-borne human pathogens (mainly mosquitoes and ticks) and are highly pathogenic for both humans and animals (Table 2) [126,127]. Flaviviruses are further classified into antigenic complexes and subcomplexes [128] that are related serologically, genetically, and etiologically. The diseases caused by flaviviruses range from fevers and encephalitides to hemorrhagic fever. There are three complexes of flaviviruses that include important agents pathogenic for humans [126].
Table 2.
List of mosquito- and tick-borne flaviviruses (genus Flavivirus, [126])a)
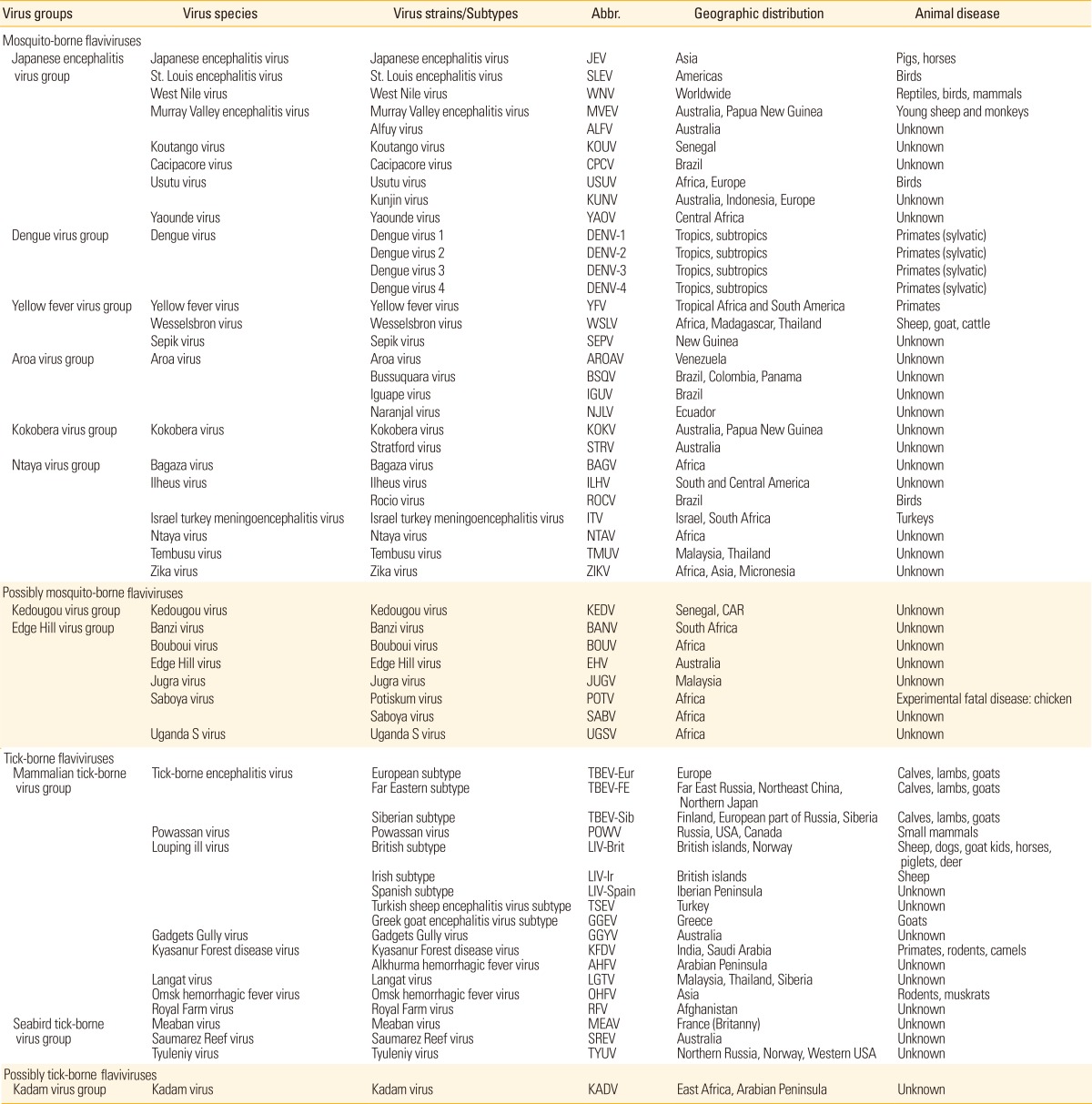
a)Adapted from Weissenbock et al. [127], with permission from Elsevier Ltd.
'Virus complexes transmitted by mosquitoes' (JEV and related encephalitis virus complex)-The mosquito-borne flaviviruses infect a variety of animal species and humans, with birds being most important to the enzootic transmission cycle of many of these viruses. Culex mosquitoes are the main vectors of these viruses. The following flaviviruses are members of this group [126]: 1) JEV (Japanese encephalitis [JE]) in Southeast Asia, 2) West Nile virus (WNV; West Nile fever/encephalitis) in Africa, North America, Europe, and Asia, 3) St. Louis encephalitis virus (SLEV; St. Louis encephalitis) in North and South America, 4) Kunjin virus (KUN; Kunjin virus encephalitis) in Australia, 5) Murray Valley encephalitis virus (MVEV; Murray Valley encephalitis) in Australia and New Guinea, 6) Rocio virus (Rocio encephalitis) in South America, and 7) Usutu virus in Africa and Europe.
'Virus complexes transmitted by ticks' (tick-borne encephalitis [TBE] complex)-Rodents are the most important vertebrate hosts. Hedgehogs, deer, and livestock may also be inapparently infected. Ixodes, Dermacentor, and Haemaphysalis ticks are the principal vectors of these viruses. The following flavivirus zoonoses are transmitted by ticks [126,128]: 1) TBE, European subtype (Central European encephalitis), 2) TBE, Eastern subtype (Russian spring-summer encephalitis), 3) Powassan encephalitis and Modoc virus encephalitis in North America, 4) louping ill in Scotland and Negishi virus encephalitis in Japan, 5) Omsk hemorrhagic fever in Siberia, 6) Kyasanur Forest disease in India, and 7) Alkhurma hemorrhagic fever in Saudi Arabia.
'Agents causing yellow fever and dengue which form two closely related virus complexes'-Simians and humans are viremic hosts and Aedes mosquitoes are the vectors. Following are the zoonotic diseases caused by viruses in this complex [126]: 1) yellow fever virus (YFV; yellow fever) in Central Africa and South America, 2) dengue virus (DENV) serotypes 1-4 in Asia, Africa, and Central and South America (dengue hemorrhagic fever, dengue shock syndrome), and 3) Wesselsbron fever in Africa.
In this review article, we focus only on two of the most important mosquito-borne flaviviruses (JEV and WNV) and briefly discuss TBE and other zoonotic encephalitides caused by other arboviruses.
Zoonosis caused by mosquito-borne flaviviruses
Japanese encephalitis virus (JEV)
JE (formerly known as Japanese B encephalitis) is the most common mosquito-transmitted encephalitic disease in Asian countries where it is regarded as a major health threat [129,130]. JEV was first documented as viral encephalitis in the 1870s and isolated in 1935 from the brain of a fatal human encephalitis case in Tokyo, Japan [131]. Recently, cases of JEV have been reported in Pakistan, Papua New Guinea, and Australia indicating expansion of the virus into new geographic regions [13,14]. JEV is classified into one single serotype with at least four distinct genotypes (I-IV) [132-134]. Genotype I strains have been isolated in Australia, Cambodia, China, Thailand, Japan, Korea, Taiwan, and Vietnam since 1967. JEV strains isolated in Australia, Indonesia, Malaysia, Papua New Guinea, and Thailand between 1951 and 1999 were included in Genotype II. Genotype III isolates have been found in temperate areas of Asia including China, Japan, Taiwan, and the Philippines since 1935, and genotype IV was found only in Indonesia in 1980 and 1981. Recently, Genotype V has been suggested for an isolate from the Muar region of Malaysia [132-135]. Genotypes I and III frequently occur in epidemic regions whereas genotypes II and IV are mostly associated with endemic transmission [133].
Up to 70% of adults in tropical regions of Asia have JEV antibodies. An estimated 50,000 cases occur every year where the most acutely infected patients are children or non-immune individuals [136]. Clinical infections are severe with a case fatality rate of 30% or severe neurologic sequels in up to 50% of the patients [137]. JEV infections occur all year long in South Asian countries while they occur during summer in temperate regions [138]. The infection in animals is subclinical, however, it causes abortion in pregnant sows and death in newborn piglets [139]. JEV causes inapparent infection in horses, but neurologic signs are observed occasionally with a high fatality rate. Seroepidemiological studies show that JEV can incidentally infect a variety of other vertebrates, including dogs, ducks, chickens, cattle, bats, snakes, and frogs [130].
JEV transmission cycle
JEV is maintained in a zoonotic cycle and transmitted by mosquitoes, primarily Culex tritaeniorhynchus [139]. This mosquito vector mainly hatches in rice fields in India and other Asian countries, with multiplication increasing up to 50% in fertilized fields. JEV also can be transmitted by C. annulus, C. annulirostris, and Aedes mosquitoes [139]. Pigs and aquatic birds are the efficient amplification and reservoir hosts of JEV that develop high-titered viremia providing a source of infection for mosquitoes (Fig. 7)[139]. In enzootic regions, pigs, frogs, and waterbirds (e.g., egrets and herons) increase the risk of transmission to humans and equines, especially in agricultural settings such as rice cultivation areas [139]. The combination of increased rice crops and pig farming gives JEV an epidemiological advantage. Humans and horses are dead-end hosts since the level of viremia is insufficient for mosquito transmission and they succumb to encephalitis after infection.
Fig. 7.

Enzootic and epizootic/epidemic transmission cycles of Japanese encephalitis virus (JEV) and West Nile virus (WNV). Left: JEV is transmitted by primarily Culex tritaeniorhynchus. Pigs and aquatic birds are the efficient amplification and reservoir hosts of JEV that develop high-titered viremia. Humans and horses are dead-end hosts since the level of viremia is insufficient for mosquito transmission. Right: WNV maintains an enzootic transmission cycle between Culex mosquitoes and birds (reservoir host). Horses, humans, and other mammals infected in a spillover transmission are "dead-end" hosts.
JEV vaccines
Despite the use of effective vaccines including both inactivated whole virus and live attenuated vaccines, JEV remains as an important cause of arthropod-transmitted viral encephalitis. The first JE vaccines available were inactivated vaccines prepared in mouse brains or primary hamster kidney cells with protection efficacy of 76% to 95% [140]. Due to safety concerns, the inactivated vaccines were replaced by the live-attenuated vaccine SA14-14-2. Recently, a new purified inactivated JE vaccine derived from Vero cell-adapted SA14-14-2 strain (IXIARO, Intercell AG, Vienna, Austria) has been licensed in the US, Europe, Canada, and Australia [141]. In addition, a live chimeric vaccine containing prM and E proteins of JEV in a backbone of attenuated YFV 17D strain was developed by Sanofi Pasteur (Chimerivax/IMOJEV, Lyon, France) [142]. The Chimerivax/IMOJEV showed outstanding immunogenicity without concerning adverse effects, thus, it was recently licensed in Australia and is currently under review in Thailand [143].
West Nile virus (WNV)
WNV was first isolated from blood of a woman in West Nile district, Uganda in 1937 [144]. Before its appearance in the New World, WNV caused sporadic outbreaks in Africa, the Middle East, Asia, and Australia. Since it first emergence in New York in 1999, West Nile virus has spread rapidly across the US, Canada, Central and South America and is currently one of the most common causes of epidemic encephalitis in the US [17]. Now, WNV has spread to Europe, the Mediterranean, Asia, and the Americas and has a global distribution like DENV, which occurs in tropical and subtropical areas of southeast Asia, the Pacific, and the Americas [145]. WNV can be divided into five lineages based on the nucleotide sequence and phylogenetic analysis. Lineages 1 and 2 are considered the main genetic lineages. Lineage 1 is highly invasive and in the 1990's the virulent strain of lineage 1 WNV emerged in the Middle East and invaded Eastern Europe and subsequently the US, resulting in a worldwide distribution currently. KUN is a variant of lineage 1 WNV that is enzootic in Australia. Lineage 2 has remained enzootic in Africa. The nucleotide sequence identity between the lineages 1 and 2 is approximately 75% [146]. Taxonomic status and virulence of lineages 3 and 4 are not clear but lineage 5 appears to be confined to India [147].
Most of the WNV infections in humans are subclinical (80%), but some develop clinical signs (20%) without neurologic signs characterized by biphasic fever, malaise, headache, nausea, anorexia, vomiting, myalgia, and arthralgia (West Nile fever). Less than 1% of the people will develop neurologic disease (1 neurologic case in ~150 total infections) and of these 10% are fatal (<0.1% of total infections) [148-150]. Neurological disease is more common in elderly people and immunosuppressed individuals and is characterized by meningitis, encephalitis, or meningoencephalitis. Some people develop a poliomyelitis-like syndrome of acute flaccid paralysis following WNV infection. Horses are the most important domestic animal affected by WNV, but severity of the disease is dependent on the neurovirulence of the infecting virus strain [151]. The strain of lineage 1 WNV that emerged in North America and Europe is highly pathogenic to horses and affected horses show a variety of neurological signs. About 1 in 12 horses develops clinical signs following infection characterized by fever, anorexia, depression, and encephalomyelitis (muscle fasciculation of face and neck, hyperexcitabilty, apprehension, lameness, ataxia, paresis and recumbency) with low-grade viremia and 90% of the horses will recover from infection. However, lineage 2 WNV only causes subclinical disease in horses [26,151]. Similarly, KUN causes mild disease in horses. WNV infection has been reported in many other wild and domestic species without clinical signs. However, there are some reports of West Nile disease in dogs, cats, sheep, squirrels, and alligators [151].
WNV transmission cycle
WNV maintains an enzootic transmission cycle between Culex mosquitoes and birds (reservoir host). Horses, humans, and other mammals infected in a spillover transmission are "dead-end" hosts (Fig. 7) [152]. In temperate regions, human infections usually coincide with the seasonal activity of Culex mosquito vectors that peaks in August and September. The virus amplification in birds occurs mostly in the late spring and early summer. In tropical regions, transmission occurs all year long. Due to high number of asymptomatic cases of WNV infection, there is a high risk of virus transmission by blood transfusion or organ transplantation [153,154].
WNV vaccine
West Nile-Innovator is a formalin-inactivated veterinary vaccine produced by Fort Dodge Animal Health (now Zoetis, Kalamazoo, MI, USA) conditionally licensed by the United States Department of Agriculture (USDA) in 2001 and received full-licensed status in 2003. Acambis Inc. (Cambridge, MA, USA) has developed a live-attenuated vaccine based on its ChimericVax technology, which has also been used in the development of vaccines against JE and DENVs. The first Chimerivax West Nile, WN01, was developed by Intervet as a single dose vaccine for horses (PreveNile, Schering-Plough Animal Health/Merck) and has been commercially available since 2006 in the US [142]. In 2010, there was an urgent recall of PreveNile due to increased incidences of adverse effects and it has been off the market since then. As a part of the development of a WNV vaccine for human use (WN02), three mutations were introduced into the WN01 E gene (L107F, A316V, and K440R). These mutations are known to attenuate the closely related JEV. In the human WN02 vaccine, these mutations were shown to independently enhance neuroattenuation of the chimera, so that reversion at one or two residues would maintain the attenuated phenotype [155]. At this time, there are no vaccines against WNV available for human use.
Zoonotic encephalitides caused by other mosquito-borne viruses
There are several other flaviviruses that can cause milder cases of human febrile illness with rash and occasional mild encephalitis. These include MVEV, the causative agent of Murray Valley encephalitis in Australia, and New Guinea and KUN virus, which is a close relative of WNV that circulates in Australia. The antibodies to MVEV cross neutralize JEV and this may explain, in part, the absence of JE in Australia. The KUN virus may cause neurological disease in horses, where as MVEV is not pathogenic to horses.
SLEV also belongs to the family Flaviviridae, genus Flavivirus and cross-reacts serologically with other mosquito-borne flaviviruses, especially with JEV and WNV, which makes serological diagnosis of these viral infections difficult. SLEV occurs in North, Central and South America and it is one of the most important arbovirus infections in North America [125]. SLEV is maintained in a mosquito-bird-mosquito cycle and wild birds (sparrow, pigeon, blue jay, and robin) are the primary vertebrate hosts [156]. Birds sustain inapparent infections but develop sufficient viremia to infect mosquito vectors. The principal vectors are C. pipiens and C. quinquefasciatus in the east, C. nigripalpus in Florida, and C. tarsalis and members of the C. pipiens complex in western states in the US [125]. Humans and domestic mammals acquire SLEV infection, but are dead-end hosts. Most SLEV infections are silent but clinical severity can vary from mild nonspecific febrile illnesses to meningitis or encephalitis. The severity of the disease and fatality rate increases with age. There is no vaccine available but SLEV infection is thought to confer life-long immunity against subsequent infection.
California encephalitis serogroup in the genus Orthobunyavirus include at least fourteen different viruses (e.g., La Crosse [LAC], Jamestown Canyon virus, snowshoe hare virus and Tahyna virus [TAHV]), each of which is transmitted by mosquitoes and has a narrow range of vertebrate hosts and a limited geographic distribution [157-159]. The LAC virus is the most important zoonotic pathogen in the California encephalitis serogroup, which is maintained by transovarial transmission in A. triseriatus, a tree-hole-breeding woodland mosquito, and is amplified by a mosquito-vertebrate-mosquito cycle involving inapparent infection of woodland rodents (e.g., squirrels and chipmunks) [160,161]. Humans are dead-end hosts, and there is no human-to-human transmission. The LAC and Jamestown Canyon virus occur in the US, snowshoe hare virus occurs in Canada. The LAC virus cause encephalitis in humans and disease is severe in children (childhood encephalitis). TAHV has been isolated in the former Soviet Union (USSR), where it caused influenza-like disease in humans [162]. There is no evidence that there is any clinical disease associated with these viruses other than humans.
Zoonosis caused by tick-borne flaviviruses
Tick-borne flaviviruses form a major group within viruses transmitted by ticks. Mosquito-borne and tick-borne flaviviruses, although distinct, appear to have evolved via a common ancestral line that diverged from viruses with no known arthropod vector [163]. Tick-borne flaviviruses are medically important arboviruses causing 10,000 to 15,000 human cases every year in Europe and Asia [164]. These viruses circulate mostly in forests, moorlands, or in steppe regions and are transmitted between ticks feeding on rodents and other mammals. Tick-borne flaviviruses can cause a wide range of disease, from mild febrile illness (with biphasic fever) to encephalitis or hemorrhagic fever, in humans and animals [165]. Two inactivated vaccines against TBEV, FSME-IMMUN (Baxter Biosciences), and Encepur (Novartis) [166,167], are used in Europe, but are not licensed in the US [168,169].
Conclusion
Many arboviruses are present in different environments across the world and virulent forms of these viruses are constantly emerging and invading new habitats. Thus, understanding the ecology and epidemiology of these arboviruses is critical to predict future emergence mechanisms and to implement preventive and control measures. In order to control and prevent the urban spread, it is extremely important to understand the mechanisms how the enzootic (sylvatic) strains reemerge and reinitiate urban transmission cycles that infect humans and domestic animals. Besides the above-mentioned viruses, there are many arboviruses with very little information available. Therefore, it is only a matter of time before we see the emergence of one of these viruses with a virulent phenotype and spread into an immunologically naïve population. Introduction of severe fever with thrombocytopenia syndrome virus (SFTSV) just in this summer of South Korea is an excellent example of a constant geographical expansion of these viruses. This virus is a novel member of the genus Phlebovirus in the family Bunyaviridae and causes fever with thrombocytopenia syndrome in humans [169]. The transmission cycle of SFTSV is not clear yet, but Haemaphysalis longicornis ticks are considered as vectors [169,170]. There have been 13 confirmed cases including 8 fatalities from SFTSV in South Korea [171].
Currently, prevention of infection with most arboviruses relies primarily on efforts to control vector populations by spraying repellents, wearing protective clothing and reducing breeding places. Despite the great efforts to make vaccines against arboviral infections, especially those that cause severe encephalitis, only a few were successful. Currently, several new generation live-attenuated vaccines based on modern molecular biology techniques are under development. ChimeriVax candidates developed against DENV, WNV, and JEV showed satisfactory safety and immunogenicity results both in vitro and in pre-clinical tests [172]. Among these, the JE-CV (chimeric vaccine) candidate is closest to registration in some endemic countries. Pre-clinical trials have proven that JE-CV provides protection against all four major genotypes currently in circulation [173]. Moreover, this vaccine candidate has advantages of being convenient and affordable since it is given as a single-dose vaccine.
Antiviral drug development is another area in need of a breakthrough. There are no specific effective antiviral treatments for any of the arboviruses, and thus only supportive cares like fluid and electrolyte management are available. However, compound screening has identified potential antiviral therapies for evaluation in animals.
Footnotes
No potential conflict of interest relevant to this article was reported.
Preparation of this review by Dr. Udeni B.R. Balasuriya was supported by the Department of Homeland Security (DHS) through Center of Excellence for Emerging and Zoonotic Animal Diseases (CEEZAD) Cooperative Agreement 2010-ST-061-AG0001, Kansas State University, Manhattan, KS, USA. The authors would like to thank Ms. Kathleen M. Shuck and Dr. Mariano Carossino for critical reading of the manuscript.
References
- 1.JOINT WHO/FAO expert committee on zoonoses. World Health Organ Tech Rep Ser. 1959;58:1–84. [PubMed] [Google Scholar]
- 2.Hollidge BS, Gonzalez-Scarano F, Soldan SS. Arboviral encephalitides: transmission, emergence, and pathogenesis. J Neuroimmune Pharmacol. 2010;5:428–442. doi: 10.1007/s11481-010-9234-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Soldan SS, Gonzalez-Scarano F. Emerging infectious diseases: the Bunyaviridae. J Neurovirol. 2005;11:412–423. doi: 10.1080/13550280591002496. [DOI] [PubMed] [Google Scholar]
- 4.Elliott R. Bunyaviruses: general features. In: Mahy BW, van Regenmortel MH, editors. Encyclopedia of virology. London: Elsevier-Academic Press; 2008. pp. 390–399. [Google Scholar]
- 5.Calisher CH. Medically important arboviruses of the United States and Canada. Clin Microbiol Rev. 1994;7:89–116. doi: 10.1128/cmr.7.1.89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gunther G, Haglund M. Tick-borne encephalopathies: epidemiology, diagnosis, treatment and prevention. CNS Drugs. 2005;19:1009–1032. doi: 10.2165/00023210-200519120-00004. [DOI] [PubMed] [Google Scholar]
- 7.Romero JR, Simonsen KA. Powassan encephalitis and Colorado tick fever. Infect Dis Clin North Am. 2008;22:545–559. doi: 10.1016/j.idc.2008.03.001. [DOI] [PubMed] [Google Scholar]
- 8.Miller BR, Nasci RS, Godsey MS, et al. First field evidence for natural vertical transmission of West Nile virus in Culex univittatus complex mosquitoes from Rift Valley province, Kenya. Am J Trop Med Hyg. 2000;62:240–246. doi: 10.4269/ajtmh.2000.62.240. [DOI] [PubMed] [Google Scholar]
- 9.Rosen L. Further observations on the mechanism of vertical transmission of flaviviruses by Aedes mosquitoes. Am J Trop Med Hyg. 1988;39:123–126. doi: 10.4269/ajtmh.1988.39.123. [DOI] [PubMed] [Google Scholar]
- 10.Pfeffer M, Dobler G. Emergence of zoonotic arboviruses by animal trade and migration. Parasit Vectors. 2010;3:35. doi: 10.1186/1756-3305-3-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Watts DM, Thompson WH, Yuill TM, DeFoliart GR, Hanson RP. Overwintering of La Crosse virus in Aedes triseriatus. Am J Trop Med Hyg. 1974;23:694–700. doi: 10.4269/ajtmh.1974.23.694. [DOI] [PubMed] [Google Scholar]
- 12.Gubler DJ. The global resurgence of arboviral diseases. Trans R Soc Trop Med Hyg. 1996;90:449–451. doi: 10.1016/s0035-9203(96)90286-2. [DOI] [PubMed] [Google Scholar]
- 13.Hanna JN, Ritchie SA, Phillips DA, et al. Japanese encephalitis in north Queensland, Australia, 1998. Med J Aust. 1999;170:533–536. doi: 10.5694/j.1326-5377.1999.tb127878.x. [DOI] [PubMed] [Google Scholar]
- 14.Hanna JN, Ritchie SA, Phillips DA, et al. An outbreak of Japanese encephalitis in the Torres Strait, Australia, 1995. Med J Aust. 1996;165:256–260. doi: 10.5694/j.1326-5377.1996.tb124960.x. [DOI] [PubMed] [Google Scholar]
- 15.Dobler G. Arboviruses causing neurological disorders in the central nervous system. Arch Virol Suppl. 1996;11:33–40. doi: 10.1007/978-3-7091-7482-1_4. [DOI] [PubMed] [Google Scholar]
- 16.Gubler DJ. Human arbovirus infections worldwide. Ann N Y Acad Sci. 2001;951:13–24. doi: 10.1111/j.1749-6632.2001.tb02681.x. [DOI] [PubMed] [Google Scholar]
- 17.Weaver SC, Barrett AD. Transmission cycles, host range, evolution and emergence of arboviral disease. Nat Rev Microbiol. 2004;2:789–801. doi: 10.1038/nrmicro1006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kuhn RJ. Togaviridae: the viruses and their replication. In: Knipe DM, Howley PM, editors. Fields virology. Philadelphia: Lippincott Williams & Wilkins; 2012. pp. 629–650. [Google Scholar]
- 19.Strauss EG, Rice CM, Strauss JH. Complete nucleotide sequence of the genomic RNA of Sindbis virus. Virology. 1984;133:92–110. doi: 10.1016/0042-6822(84)90428-8. [DOI] [PubMed] [Google Scholar]
- 20.Simmons DT, Strauss JH. Replication of Sindbis virus. I. Relative size and genetic content of 26S and 49S RNA. J Mol Biol. 1972;71:599–613. [PubMed] [Google Scholar]
- 21.Powers AM, Brault AC, Shirako Y, et al. Evolutionary relationships and systematics of the alphaviruses. J Virol. 2001;75:10118–10131. doi: 10.1128/JVI.75.21.10118-10131.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Strauss JH, Strauss EG. The alphaviruses: gene expression, replication, and evolution. Microbiol Rev. 1994;58:491–562. doi: 10.1128/mr.58.3.491-562.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.La Linn M, Gardner J, Warrilow D, et al. Arbovirus of marine mammals: a new alphavirus isolated from the elephant seal louse, Lepidophthirus macrorhini. J Virol. 2001;75:4103–4109. doi: 10.1128/JVI.75.9.4103-4109.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Travassos da Rosa AP, Turell MJ, Watts DM, et al. Trocara virus: a newly recognized Alphavirus (Togaviridae) isolated from mosquitoes in the Amazon Basin. Am J Trop Med Hyg. 2001;64:93–97. doi: 10.4269/ajtmh.2001.64.93. [DOI] [PubMed] [Google Scholar]
- 25.Zacks MA, Paessler S. Encephalitic alphaviruses. Vet Microbiol. 2010;140:281–286. doi: 10.1016/j.vetmic.2009.08.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Krauss H, Weber A, Appel M, et al. Zoonoses: infectious diseases transmissible from animals to humans. Washington: ASM Press; 2003. [Google Scholar]
- 27.Scott TW, Weaver SC. Eastern equine encephalomyelitis virus: epidemiology and evolution of mosquito transmission. Adv Virus Res. 1989;37:277–328. doi: 10.1016/s0065-3527(08)60838-6. [DOI] [PubMed] [Google Scholar]
- 28.Hanson RP. An epizootic of equine encephalomyelitis that occurred in Massachusetts in 1831. Am J Trop Med Hyg. 1957;6:858–862. doi: 10.4269/ajtmh.1957.6.858. [DOI] [PubMed] [Google Scholar]
- 29.Sabattini MS, Monath TP, Mitchell CJ, et al. Arbovirus investigations in Argentina, 1977-1980. I. Historical aspects and description of study sites. Am J Trop Med Hyg. 1985;34:937–944. [PubMed] [Google Scholar]
- 30.Weaver SC, Winegar R, Manger ID, Forrester NL. Alphaviruses: population genetics and determinants of emergence. Antiviral Res. 2012;94:242–257. doi: 10.1016/j.antiviral.2012.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Aguilar PV, Paessler S, Carrara AS, et al. Variation in interferon sensitivity and induction among strains of eastern equine encephalitis virus. J Virol. 2005;79:11300–11310. doi: 10.1128/JVI.79.17.11300-11310.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Aguilar PV, Robich RM, Turell MJ, et al. Endemic eastern equine encephalitis in the Amazon region of Peru. Am J Trop Med Hyg. 2007;76:293–298. [PubMed] [Google Scholar]
- 33.Weaver SC, Powers AM, Brault AC, Barrett AD. Molecular epidemiological studies of veterinary arboviral encephalitides. Vet J. 1999;157:123–138. doi: 10.1053/tvjl.1998.0289. [DOI] [PubMed] [Google Scholar]
- 34.Brault AC, Powers AM, Chavez CL, et al. Genetic and antigenic diversity among eastern equine encephalitis viruses from North, Central, and South America. Am J Trop Med Hyg. 1999;61:579–586. doi: 10.4269/ajtmh.1999.61.579. [DOI] [PubMed] [Google Scholar]
- 35.Weaver SC, Vasilakis N. Molecular evolution of dengue viruses: contributions of phylogenetics to understanding the history and epidemiology of the preeminent arboviral disease. Infect Genet Evol. 2009;9:523–540. doi: 10.1016/j.meegid.2009.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chang GJ, Trent DW. Nucleotide sequence of the genome region encoding the 26S mRNA of eastern equine encephalomyelitis virus and the deduced amino acid sequence of the viral structural proteins. J Gen Virol. 1987;68(Pt 8):2129–2142. doi: 10.1099/0022-1317-68-8-2129. [DOI] [PubMed] [Google Scholar]
- 37.Gibney KB, Robinson S, Mutebi JP, et al. Eastern equine encephalitis: an emerging arboviral disease threat, Maine, 2009. Vector Borne Zoonotic Dis. 2011;11:637–639. doi: 10.1089/vbz.2010.0189. [DOI] [PubMed] [Google Scholar]
- 38.Arrigo NC, Adams AP, Weaver SC. Evolutionary patterns of eastern equine encephalitis virus in North versus South America suggest ecological differences and taxonomic revision. J Virol. 2010;84:1014–1025. doi: 10.1128/JVI.01586-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Deresiewicz RL, Thaler SJ, Hsu L, Zamani AA. Clinical and neuroradiographic manifestations of eastern equine encephalitis. N Engl J Med. 1997;336:1867–1874. doi: 10.1056/NEJM199706263362604. [DOI] [PubMed] [Google Scholar]
- 40.Causey OR, Causey CE, Maroja OM, Macedo DG. The isolation of arthropod-borne viruses, including members of two hitherto undescribed serological groups, in the Amazon region of Brazil. Am J Trop Med Hyg. 1961;10:227–249. doi: 10.4269/ajtmh.1961.10.227. [DOI] [PubMed] [Google Scholar]
- 41.Corniou B, Ardoin P, Bartholomew C, Ince W, Massiah V. First isolation of a South American strain of Eastern Equine virus from a case of encephalitis in Trinidad. Trop Geogr Med. 1972;24:162–167. [PubMed] [Google Scholar]
- 42.Sabattini MS, Daffner JF, Monath TP, et al. Localized eastern equine encephalitis in Santiago del Estero Province, Argentina, without human infection. . Medicina (B Aires) 1991;51:3–8. [PubMed] [Google Scholar]
- 43.Aguilar PV, Greene IP, Coffey LL, et al. Endemic Venezuelan equine encephalitis in northern Peru. Emerg Infect Dis. 2004;10:880–888. doi: 10.3201/eid1005.030634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Monath TP, Sabattini MS, Pauli R, et al. Arbovirus investigations in Argentina, 1977-1980. IV. Serologic surveys and sentinel equine program. Am J Trop Med Hyg. 1985;34:966–975. [PubMed] [Google Scholar]
- 45.Armstrong PM, Andreadis TG. Eastern equine encephalitis virus in mosquitoes and their role as bridge vectors. Emerg Infect Dis. 2010;16:1869–1874. doi: 10.3201/eid1612.100640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Molaei G, Oliver J, Andreadis TG, Armstrong PM, Howard JJ. Molecular identification of blood-meal sources in Culiseta melanura and Culiseta morsitans from an endemic focus of eastern equine encephalitis virus in New York. Am J Trop Med Hyg. 2006;75:1140–1147. [PubMed] [Google Scholar]
- 47.Estep LK, McClure CJ, Burkett-Cadena ND, et al. A multi-year study of mosquito feeding patterns on avian hosts in a southeastern focus of eastern equine encephalitis virus. Am J Trop Med Hyg. 2011;84:718–726. doi: 10.4269/ajtmh.2011.10-0586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Bingham AM, Graham SP, Burkett-Cadena ND, White GS, Hassan HK, Unnasch TR. Detection of eastern equine encephalomyelitis virus RNA in North American snakes. Am J Trop Med Hyg. 2012;87:1140–1144. doi: 10.4269/ajtmh.2012.12-0257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Graham SP, Hassan HK, Chapman T, White G, Guyer C, Unnasch TR. Serosurveillance of eastern equine encephalitis virus in amphibians and reptiles from Alabama, USA. Am J Trop Med Hyg. 2012;86:540–544. doi: 10.4269/ajtmh.2012.11-0283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.White G, Ottendorfer C, Graham S, Unnasch TR. Competency of reptiles and amphibians for eastern equine encephalitis virus. Am J Trop Med Hyg. 2011;85:421–425. doi: 10.4269/ajtmh.2011.11-0006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Day JF, Stark LM. Eastern equine encephalitis transmission to emus (Dromaius novaehollandiae) in Volusia County, Florida: 1992 through 1994. J Am Mosq Control Assoc. 1996;12(3 Pt 1):429–436. [PubMed] [Google Scholar]
- 52.Fothergill LD, Dingle JH, Fellow JJ. A fatal disease of pigeons caused by the virus of the eastern variety of equine encephalomyelitis. Science. 1938;88:549–550. doi: 10.1126/science.88.2293.549-a. [DOI] [PubMed] [Google Scholar]
- 53.Guy JS, Ficken MD, Barnes HJ, Wages DP, Smith LG. Experimental infection of young turkeys with eastern equine encephalitis virus and highlands J virus. Avian Dis. 1993;37:389–395. [PubMed] [Google Scholar]
- 54.Luginbuhl RE, Satriano SF, Helmboldt CF, Lamson AL, Jungherr EL. Investigation of eastern equine encephalomyelitis. II. Outbreaks in Connecticut pheasants. Am J Hyg. 1958;67:4–9. doi: 10.1093/oxfordjournals.aje.a119917. [DOI] [PubMed] [Google Scholar]
- 55.Satriano SF, Luginbuhl RE, Wallis RC, Jungherr EL, Williamson LA. Investigation of eastern equine encephalomyelitis. IV. Susceptibility and transmission studies with virus of pheasant origin. Am J Hyg. 1958;67:21–34. [PubMed] [Google Scholar]
- 56.Arrigo NC, Adams AP, Watts DM, Newman PC, Weaver SC. Cotton rats and house sparrows as hosts for North and South American strains of eastern equine encephalitis virus. Emerg Infect Dis. 2010;16:1373–1380. doi: 10.3201/eid1609.100459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Bauer RW, Gill MS, Poston RP, Kim DY. Naturally occurring eastern equine encephalitis in a Hampshire wether. J Vet Diagn Invest. 2005;17:281–285. doi: 10.1177/104063870501700314. [DOI] [PubMed] [Google Scholar]
- 58.Tate CM, Howerth EW, Stallknecht DE, Allison AB, Fischer JR, Mead DG. Eastern equine encephalitis in a free-ranging white-tailed deer (Odocoileus virginianus) J Wildl Dis. 2005;41:241–245. doi: 10.7589/0090-3558-41.1.241. [DOI] [PubMed] [Google Scholar]
- 59.Farrar MD, Miller DL, Baldwin CA, Stiver SL, Hall CL. Eastern equine encephalitis in dogs. J Vet Diagn Invest. 2005;17:614–617. doi: 10.1177/104063870501700619. [DOI] [PubMed] [Google Scholar]
- 60.Freier JE. Eastern equine encephalomyelitis. Lancet. 1993;342:1281–1282. doi: 10.1016/0140-6736(93)92366-2. [DOI] [PubMed] [Google Scholar]
- 61.McGee ED, Littleton CH, Mapp JB, Brown RJ. Eastern equine encephalomyelitis in an adult cow. Vet Pathol. 1992;29:361–363. doi: 10.1177/030098589202900414. [DOI] [PubMed] [Google Scholar]
- 62.Walder R, Suarez OM, Calisher CH. Arbovirus studies in southwestern Venezuela during 1973-1981. II. Isolations and further studies of Venezuelan and eastern equine encephalitis, Una, Itaqui, and Moju viruses. Am J Trop Med Hyg. 1984;33:483–491. [PubMed] [Google Scholar]
- 63.Strizki JM, Repik PM. Differential reactivity of immune sera from human vaccinees with field strains of eastern equine encephalitis virus. Am J Trop Med Hyg. 1995;53:564–570. doi: 10.4269/ajtmh.1995.53.564. [DOI] [PubMed] [Google Scholar]
- 64.Hahn CS, Lustig S, Strauss EG, Strauss JH. Western equine encephalitis virus is a recombinant virus. Proc Natl Acad Sci U S A. 1988;85:5997–6001. doi: 10.1073/pnas.85.16.5997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Weaver SC, Kang W, Shirako Y, Rumenapf T, Strauss EG, Strauss JH. Recombinational history and molecular evolution of western equine encephalomyelitis complex alphaviruses. J Virol. 1997;71:613–623. doi: 10.1128/jvi.71.1.613-623.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Meyer KF, Haring CM, Howitt B. The etiology of epizootic encephalomyelitis of horses in the San Joaquin Valley, 1930. Science. 1931;74:227–228. doi: 10.1126/science.74.1913.227. [DOI] [PubMed] [Google Scholar]
- 67.Howitt B. Recovery of the virus of equine encephalomyelitis from the brain of a child. Science. 1938;88:455–456. doi: 10.1126/science.88.2289.455. [DOI] [PubMed] [Google Scholar]
- 68.Reisen WK, Chiles RE. Prevalence of antibodies to western equine encephalomyelitis and St. Louis encephalitis viruses in residents of California exposed to sporadic and consistent enzootic transmission. Am J Trop Med Hyg. 1997;57:526–529. doi: 10.4269/ajtmh.1997.57.526. [DOI] [PubMed] [Google Scholar]
- 69.Reisen WK, Hardy JL, Lothrop HD. Landscape ecology of arboviruses in southern California: patterns in the epizootic dissemination of western equine encephalomyelitis and St. Louis encephalitis viruses in Coachella Valley, 1991-1992. J Med Entomol. 1995;32:267–275. doi: 10.1093/jmedent/32.3.267. [DOI] [PubMed] [Google Scholar]
- 70.Reimann CA, Hayes EB, DiGuiseppi C, et al. Epidemiology of neuroinvasive arboviral disease in the United States, 1999-2007. Am J Trop Med Hyg. 2008;79:974–979. [PubMed] [Google Scholar]
- 71.Longshore WA, Jr, Stevens IM, Hollister AC, Jr, Gittelsohn A, Lennette EH. Epidemiologic observations on acute infectious encephalitis in California, with special reference to the 1952 outbreak. Am J Hyg. 1956;63:69–86. doi: 10.1093/oxfordjournals.aje.a119793. [DOI] [PubMed] [Google Scholar]
- 72.Doby PB, Schnurrenberger PR, Martin RJ, Hanson LE, Sherrick GW, Schoenholz WK. Western encephalitis in Illinois horses and ponies. J Am Vet Med Assoc. 1966;148:422–427. [PubMed] [Google Scholar]
- 73.Jennings WL, Allen RH, Lewis AL. Western equine encephalomyelitis in a Florida horse. Am J Trop Med Hyg. 1966;15:96–97. doi: 10.4269/ajtmh.1966.15.96. [DOI] [PubMed] [Google Scholar]
- 74.Karabatsos N, Lewis AL, Calisher CH, Hunt AR, Roehrig JT. Identification of Highlands J virus from a Florida horse. Am J Trop Med Hyg. 1988;39:603–606. doi: 10.4269/ajtmh.1988.39.603. [DOI] [PubMed] [Google Scholar]
- 75.Ficken MD, Wages DP, Guy JS, Quinn JA, Emory WH. High mortality of domestic turkeys associated with Highlands J virus and eastern equine encephalitis virus infections. Avian Dis. 1993;37:585–590. [PubMed] [Google Scholar]
- 76.Guy JS, Barnes HJ, Ficken MD, Smith LG, Emory WH, Wages DP. Decreased egg production in turkeys experimentally infected with eastern equine encephalitis virus or Highlands J virus. Avian Dis. 1994;38:563–571. [PubMed] [Google Scholar]
- 77.Guy JS, Barnes HJ, Smith LG. Experimental infection of young broiler chickens with eastern equine encephalitis virus and Highlands J virus. Avian Dis. 1994;38:572–582. [PubMed] [Google Scholar]
- 78.Wages DP, Ficken MD, Guy JS, Cummings TS, Jennings SR. Egg-production drop in turkeys associated with alphaviruses: eastern equine encephalitis virus and Highlands J virus. Avian Dis. 1993;37:1163–1166. [PubMed] [Google Scholar]
- 79.Reisen WK, Monath TP. Western equine encephalomyelitis. In: Monath TP, editor. The arboviruses: epidemiology and ecology. Boca Raton: CRC Press; 1988. pp. 89–138. [Google Scholar]
- 80.Calisher CH, Karabatsos N, Lazuick JS, Monath TP, Wolff KL. Reevaluation of the western equine encephalitis antigenic complex of alphaviruses (family Togaviridae) as determined by neutralization tests. Am J Trop Med Hyg. 1988;38:447–452. doi: 10.4269/ajtmh.1988.38.447. [DOI] [PubMed] [Google Scholar]
- 81.Fulhorst CF, Hardy JL, Eldridge BF, Presser SB, Reeves WC. Natural vertical transmission of western equine encephalomyelitis virus in mosquitoes. Science. 1994;263:676–678. doi: 10.1126/science.8303276. [DOI] [PubMed] [Google Scholar]
- 82.Pittman P, Plotkin SA. Biodefense and special pathogen vaccines. In: Plotkin SA, Orenstein WA, Offit PA, editors. Vaccines. Philadelphia: Elsevier; 2008. pp. 1123–1133. [Google Scholar]
- 83.Barber TL, Walton TE, Lewis KJ. Efficacy of trivalent inactivated encephalomyelitis virus vaccine in horses. Am J Vet Res. 1978;39:621–625. [PubMed] [Google Scholar]
- 84.Dickerman RW, Cupp EW, Groot H, et al. Venezuelan equine encephalitis virus activity in northern Colombia during April and May 1983. Bull Pan Am Health Organ. 1986;20:276–283. [PubMed] [Google Scholar]
- 85.Estrada-Franco JG, Navarro-Lopez R, Freier JE, et al. Venezuelan equine encephalitis virus, southern Mexico. Emerg Infect Dis. 2004;10:2113–2121. doi: 10.3201/eid1012.040393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Oberste MS, Schmura SM, Weaver SC, Smith JF. Geographic distribution of Venezuelan equine encephalitis virus subtype IE genotypes in Central America and Mexico. Am J Trop Med Hyg. 1999;60:630–634. doi: 10.4269/ajtmh.1999.60.630. [DOI] [PubMed] [Google Scholar]
- 87.Quiroz E, Aguilar PV, Cisneros J, Tesh RB, Weaver SC. Venezuelan equine encephalitis in Panama: fatal endemic disease and genetic diversity of etiologic viral strains. PLoS Negl Trop Dis. 2009;3:e472. doi: 10.1371/journal.pntd.0000472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Weaver SC, Ferro C, Barrera R, Boshell J, Navarro JC. Venezuelan equine encephalitis. Annu Rev Entomol. 2004;49:141–174. doi: 10.1146/annurev.ento.49.061802.123422. [DOI] [PubMed] [Google Scholar]
- 89.Weaver SC, Pfeffer M, Marriott K, Kang W, Kinney RM. Genetic evidence for the origins of Venezuelan equine encephalitis virus subtype IAB outbreaks. Am J Trop Med Hyg. 1999;60:441–448. doi: 10.4269/ajtmh.1999.60.441. [DOI] [PubMed] [Google Scholar]
- 90.Sneider JM, Kinney RM, Tsuchiya KR, Trent DW. Molecular evidence that epizootic Venezuelan equine encephalitis (VEE) I-AB viruses are not evolutionary derivatives of enzootic VEE subtype I-E or II viruses. J Gen Virol. 1993;74(Pt 3):519–523. doi: 10.1099/0022-1317-74-3-519. [DOI] [PubMed] [Google Scholar]
- 91.Zehmer RB, Dean PB, Sudia WD, Calisher CH, Sather GE, Parker RL. Venezuelan equine encephalitis epidemic in Texas, 1971. Health Serv Rep. 1974;89:278–282. [PMC free article] [PubMed] [Google Scholar]
- 92.Beck CE, Wyckoff RW. Venezuelan equine encephalomyelitis. Science. 1938;88:530. doi: 10.1126/science.88.2292.530. [DOI] [PubMed] [Google Scholar]
- 93.Kinney RM, Trent DW, France JK. Comparative immunological and biochemical analyses of viruses in the Venezuelan equine encephalitis complex. J Gen Virol. 1983;64(Pt 1):135–147. doi: 10.1099/0022-1317-64-1-135. [DOI] [PubMed] [Google Scholar]
- 94.Young NA, Johnson KM. Antigenic variants of Venezuelan equine encephalitis virus: their geographic distribution and epidemiologic significance. Am J Epidemiol. 1969;89:286–307. doi: 10.1093/oxfordjournals.aje.a120942. [DOI] [PubMed] [Google Scholar]
- 95.Brault AC, Powers AM, Holmes EC, Woelk CH, Weaver SC. Positively charged amino acid substitutions in the e2 envelope glycoprotein are associated with the emergence of Venezuelan equine encephalitis virus. J Virol. 2002;76:1718–1730. doi: 10.1128/JVI.76.4.1718-1730.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Brault AC, Powers AM, Weaver SC. Vector infection determinants of Venezuelan equine encephalitis virus reside within the E2 envelope glycoprotein. J Virol. 2002;76:6387–6392. doi: 10.1128/JVI.76.12.6387-6392.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Adams AP, Navarro-Lopez R, Ramirez-Aguilar FJ, et al. Venezuelan equine encephalitis virus activity in the Gulf Coast region of Mexico, 2003-2010. PLoS Negl Trop Dis. 2012;6:e1875. doi: 10.1371/journal.pntd.0001875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Deardorff ER, Weaver SC. Vector competence of Culex (Melanoconion) taeniopus for equine-virulent subtype IE strains of Venezuelan equine encephalitis virus. Am J Trop Med Hyg. 2010;82:1047–1052. doi: 10.4269/ajtmh.2010.09-0556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Powers AM, Oberste MS, Brault AC, et al. Repeated emergence of epidemic/epizootic Venezuelan equine encephalitis from a single genotype of enzootic subtype ID virus. J Virol. 1997;71:6697–6705. doi: 10.1128/jvi.71.9.6697-6705.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Aguilar PV, Adams AP, Suarez V, et al. Genetic characterization of Venezuelan equine encephalitis virus from Bolivia, Ecuador and Peru: identification of a new subtype ID lineage. PLoS Negl Trop Dis. 2009;3:e514. doi: 10.1371/journal.pntd.0000514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Johnson KM, Martin DH. Venezuelan equine encephalitis. Adv Vet Sci Comp Med. 1974;18:79–116. [PubMed] [Google Scholar]
- 102.Rivas F, Diaz LA, Cardenas VM, et al. Epidemic Venezuelan equine encephalitis in La Guajira, Colombia, 1995. J Infect Dis. 1997;175:828–832. doi: 10.1086/513978. [DOI] [PubMed] [Google Scholar]
- 103.Weaver SC, Salas R, Rico-Hesse R, et al. VEE Study Group. Re-emergence of epidemic Venezuelan equine encephalomyelitis in South America. Lancet. 1996;348:436–440. doi: 10.1016/s0140-6736(96)02275-1. [DOI] [PubMed] [Google Scholar]
- 104.Aguilar PV, Estrada-Franco JG, Navarro-Lopez R, Ferro C, Haddow AD, Weaver SC. Endemic Venezuelan equine encephalitis in the Americas: hidden under the dengue umbrella. Future Virol. 2011;6:721–740. doi: 10.2217/FVL.11.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Ferro C, Boshell J, Moncayo AC, et al. Natural enzootic vectors of Venezuelan equine encephalitis virus, Magdalena Valley, Colombia. Emerg Infect Dis. 2003;9:49–54. doi: 10.3201/eid0901.020136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Galindo P, Grayson MA. Culex (Melanoconion) aikenii: natural vector in Panama of endemic Venezuelan encephalitis. Science. 1971;172:594–595. doi: 10.1126/science.172.3983.594. [DOI] [PubMed] [Google Scholar]
- 107.Grayson MA, Galindo P. Epidemiologic studies of Venezuelan equine encephalitis virus in Almirante, Panama. Am J Epidemiol. 1968;88:80–96. doi: 10.1093/oxfordjournals.aje.a120870. [DOI] [PubMed] [Google Scholar]
- 108.Forshey BM, Guevara C, Laguna-Torres VA, et al. Arboviral etiologies of acute febrile illnesses in Western South America, 2000-2007. PLoS Negl Trop Dis. 2010;4:e787. doi: 10.1371/journal.pntd.0000787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Franck PT, Johnson KM. An outbreak of Venezuelan encephalitis in man in the Panama Canal Zone. Am J Trop Med Hyg. 1970;19:860–865. doi: 10.4269/ajtmh.1970.19.860. [DOI] [PubMed] [Google Scholar]
- 110.Hawley RJ, Eitzen EM., Jr Biological weapons: a primer for microbiologists. Annu Rev Microbiol. 2001;55:235–253. doi: 10.1146/annurev.micro.55.1.235. [DOI] [PubMed] [Google Scholar]
- 111.Reed DS, Lind CM, Sullivan LJ, Pratt WD, Parker MD. Aerosol infection of cynomolgus macaques with enzootic strains of venezuelan equine encephalitis viruses. J Infect Dis. 2004;189:1013–1017. doi: 10.1086/382281. [DOI] [PubMed] [Google Scholar]
- 112.Berge TO, Banks IS, Tigertt WD. Attenuation of Venezuelan equine encephalomyelitis virus by in vitro cultivation in guinea-pig heart cells. Am J Hyg. 1961;73:209–218. [Google Scholar]
- 113.Kinney RM, Chang GJ, Tsuchiya KR, et al. Attenuation of Venezuelan equine encephalitis virus strain TC-83 is encoded by the 5'-noncoding region and the E2 envelope glycoprotein. J Virol. 1993;67:1269–1277. doi: 10.1128/jvi.67.3.1269-1277.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Kinney RM, Johnson BJ, Welch JB, Tsuchiya KR, Trent DW. The full-length nucleotide sequences of the virulent Trinidad donkey strain of Venezuelan equine encephalitis virus and its attenuated vaccine derivative, strain TC-83. Virology. 1989;170:19–30. doi: 10.1016/0042-6822(89)90347-4. [DOI] [PubMed] [Google Scholar]
- 115.Paessler S, Weaver SC. Vaccines for Venezuelan equine encephalitis. Vaccine. 2009;27(Suppl 4):D80–D85. doi: 10.1016/j.vaccine.2009.07.095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Fine DL, Roberts BA, Teehee ML, et al. Venezuelan equine encephalitis virus vaccine candidate (V3526) safety, immunogenicity and efficacy in horses. Vaccine. 2007;25:1868–1876. doi: 10.1016/j.vaccine.2006.10.030. [DOI] [PubMed] [Google Scholar]
- 117.Davis NL, Brown KW, Greenwald GF, et al. Attenuated mutants of Venezuelan equine encephalitis virus containing lethal mutations in the PE2 cleavage signal combined with a second-site suppressor mutation in E1. Virology. 1995;212:102–110. doi: 10.1006/viro.1995.1458. [DOI] [PubMed] [Google Scholar]
- 118.Davis NL, Grieder FB, Smith JF, et al. A molecular genetic approach to the study of Venezuelan equine encephalitis virus pathogenesis. Arch Virol Suppl. 1994;9:99–109. doi: 10.1007/978-3-7091-9326-6_11. [DOI] [PubMed] [Google Scholar]
- 119.Davis NL, Powell N, Greenwald GF, et al. Attenuating mutations in the E2 glycoprotein gene of Venezuelan equine encephalitis virus: construction of single and multiple mutants in a full-length cDNA clone. Virology. 1991;183:20–31. doi: 10.1016/0042-6822(91)90114-q. [DOI] [PubMed] [Google Scholar]
- 120.Hart MK, Lind C, Bakken R, Robertson M, Tammariello R, Ludwig GV. Onset and duration of protective immunity to IA/IB and IE strains of Venezuelan equine encephalitis virus in vaccinated mice. Vaccine. 2001;20:616–622. doi: 10.1016/s0264-410x(01)00337-1. [DOI] [PubMed] [Google Scholar]
- 121.Pratt WD, Davis NL, Johnston RE, Smith JF. Genetically engineered, live attenuated vaccines for Venezuelan equine encephalitis: testing in animal models. Vaccine. 2003;21:3854–3862. doi: 10.1016/s0264-410x(03)00328-1. [DOI] [PubMed] [Google Scholar]
- 122.Reed DS, Lind CM, Lackemeyer MG, Sullivan LJ, Pratt WD, Parker MD. Genetically engineered, live, attenuated vaccines protect nonhuman primates against aerosol challenge with a virulent IE strain of Venezuelan equine encephalitis virus. Vaccine. 2005;23:3139–3147. doi: 10.1016/j.vaccine.2004.12.023. [DOI] [PubMed] [Google Scholar]
- 123.Gould EA, Solomon T. Pathogenic flaviviruses. Lancet. 2008;371:500–509. doi: 10.1016/S0140-6736(08)60238-X. [DOI] [PubMed] [Google Scholar]
- 124.Shope RE. Epidemiology of other arthropod-borne flaviviruses infecting humans. Adv Virus Res. 2003;61:373–391. doi: 10.1016/s0065-3527(03)61009-2. [DOI] [PubMed] [Google Scholar]
- 125.Gubler DJ, Kuno G, Markoff L. Flaviviruses. In: Knipe DM, Howley PM, Griffin DE, et al., editors. Fields virology. Philadelphia: Lippincott William & Wilkins; 2007. pp. 1153–1253. [Google Scholar]
- 126.Simmonds P, Becher P, Collet MS, et al. Family Flaviviridae. In: King AM, Adams MJ, Carstens EB, Lefkowitz EJ, editors. Virus taxonomy classification and nomenclature of viruses. Ninth report of the International Commitee on Taxonomy of Viruses. San Diego: Elsevier Academic Press; 2011. pp. 1003–1020. [Google Scholar]
- 127.Weissenbock H, Hubalek Z, Bakonyi T, Nowotny N. Zoonotic mosquito-borne flaviviruses: worldwide presence of agents with proven pathogenicity and potential candidates of future emerging diseases. Vet Microbiol. 2010;140:271–280. doi: 10.1016/j.vetmic.2009.08.025. [DOI] [PubMed] [Google Scholar]
- 128.Calisher CH. Antigenic classification and taxonomy of flaviviruses (family Flaviviridae) emphasizing a universal system for the taxonomy of viruses causing tick-borne encephalitis. Acta Virol. 1988;32:469–478. [PubMed] [Google Scholar]
- 129.van-den-Hurk AF, Ritchie SA, Johansen CA, Mackenzie JS, Smith GA. Domestic pigs and Japanese encephalitis virus infection, Australia. Emerg Infect Dis. 2008;14:1736–1738. doi: 10.3201/eid1411.071368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.van den Hurk AF, Ritchie SA, Mackenzie JS. Ecology and geographical expansion of Japanese encephalitis virus. Annu Rev Entomol. 2009;54:17–35. doi: 10.1146/annurev.ento.54.110807.090510. [DOI] [PubMed] [Google Scholar]
- 131.Lewis L, Taylor HG, Sorem MB, Norcross JW, Kindsvatter VH. Japanese B encephalitis: clinical observations in an outbreak on Okinawa Shima. Arch Neurol Psychiatry. 1947;57:430–463. [PubMed] [Google Scholar]
- 132.Chen WR, Rico-Hesse R, Tesh RB. A new genotype of Japanese encephalitis virus from Indonesia. Am J Trop Med Hyg. 1992;47:61–69. doi: 10.4269/ajtmh.1992.47.61. [DOI] [PubMed] [Google Scholar]
- 133.Chen WR, Tesh RB, Rico-Hesse R. Genetic variation of Japanese encephalitis virus in nature. J Gen Virol. 1990;71(Pt 12):2915–2922. doi: 10.1099/0022-1317-71-12-2915. [DOI] [PubMed] [Google Scholar]
- 134.Tsarev SA, Sanders ML, Vaughn DW, Innis BL. Phylogenetic analysis suggests only one serotype of Japanese encephalitis virus. Vaccine. 2000;18(Suppl 2):36–43. doi: 10.1016/s0264-410x(00)00039-6. [DOI] [PubMed] [Google Scholar]
- 135.Mohammed MA, Galbraith SE, Radford AD, et al. Molecular phylogenetic and evolutionary analyses of Muar strain of Japanese encephalitis virus reveal it is the missing fifth genotype. Infect Genet Evol. 2011;11:855–862. doi: 10.1016/j.meegid.2011.01.020. [DOI] [PubMed] [Google Scholar]
- 136.Tamada K, Kasai H, Hara M, Aoyama T, Takeuchi Y, Yamashita R. Japanese encephalitis in a 7-month-old Japanese boy. Acta Paediatr Jpn. 1990;32:98–103. doi: 10.1111/j.1442-200x.1990.tb00791.x. [DOI] [PubMed] [Google Scholar]
- 137.Kumar R, Mathur A, Singh KB, et al. Clinical sequelae of Japanese encephalitis in children. Indian J Med Res. 1993;97:9–13. [PubMed] [Google Scholar]
- 138.Mackenzie JS. Emerging zoonotic encephalitis viruses: lessons from Southeast Asia and Oceania. J Neurovirol. 2005;11:434–440. doi: 10.1080/13550280591002487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Rosen L. The natural history of Japanese encephalitis virus. Annu Rev Microbiol. 1986;40:395–414. doi: 10.1146/annurev.mi.40.100186.002143. [DOI] [PubMed] [Google Scholar]
- 140.Halstead SB, Thomas SJ. Japanese encephalitis: new options for active immunization. Clin Infect Dis. 2010;50:1155–1164. doi: 10.1086/651271. [DOI] [PubMed] [Google Scholar]
- 141.Tauber E, Kollaritsch H, Korinek M, et al. Safety and immunogenicity of a Vero-cell-derived, inactivated Japanese encephalitis vaccine: a non-inferiority, phase III, randomised controlled trial. Lancet. 2007;370:1847–1853. doi: 10.1016/S0140-6736(07)61780-2. [DOI] [PubMed] [Google Scholar]
- 142.Guy B, Guirakhoo F, Barban V, Higgs S, Monath TP, Lang J. Preclinical and clinical development of YFV 17D-based chimeric vaccines against dengue, West Nile and Japanese encephalitis viruses. Vaccine. 2010;28:632–649. doi: 10.1016/j.vaccine.2009.09.098. [DOI] [PubMed] [Google Scholar]
- 143.Halstead SB, Thomas SJ. New Japanese encephalitis vaccines: alternatives to production in mouse brain. Expert Rev Vaccines. 2011;10:355–364. doi: 10.1586/erv.11.7. [DOI] [PubMed] [Google Scholar]
- 144.Smithburn KC, Hughes TP, Burke AW, Paul JH. A neurotropic virus isolated from the blood of a native of Uganda. Am J Trop Med. 1940;20:471–492. [Google Scholar]
- 145.Dengue and dengue hemorrhagic fever. Factsheet no. 117 [Internet] Geneva: World Health Organization; 2008. [cite 2013 Sep 1]. Available from: http://www.who.int/mediacentre/factsheets/fs117/en/ [Google Scholar]
- 146.Lanciotti RS, Ebel GD, Deubel V, et al. Complete genome sequences and phylogenetic analysis of West Nile virus strains isolated from the United States, Europe, and the Middle East. Virology. 2002;298:96–105. doi: 10.1006/viro.2002.1449. [DOI] [PubMed] [Google Scholar]
- 147.Kramer LD, Styer LM, Ebel GD. A global perspective on the epidemiology of West Nile virus. Annu Rev Entomol. 2008;53:61–81. doi: 10.1146/annurev.ento.53.103106.093258. [DOI] [PubMed] [Google Scholar]
- 148.Hayes EB, Gubler DJ. West Nile virus: epidemiology and clinical features of an emerging epidemic in the United States. Annu Rev Med. 2006;57:181–194. doi: 10.1146/annurev.med.57.121304.131418. [DOI] [PubMed] [Google Scholar]
- 149.Mostashari F, Bunning ML, Kitsutani PT, et al. Epidemic West Nile encephalitis, New York, 1999: results of a household-based seroepidemiological survey. Lancet. 2001;358:261–264. doi: 10.1016/S0140-6736(01)05480-0. [DOI] [PubMed] [Google Scholar]
- 150.Hayes EB, Komar N, Nasci RS, Montgomery SP, O'Leary DR, Campbell GL. Epidemiology and transmission dynamics of West Nile virus disease. Emerg Infect Dis. 2005;11:1167–1173. doi: 10.3201/eid1108.050289a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Bowen RA. Flaviviridae. In: MacLachlan NJ, Dubovi EJ, editors. Fenner's veterinary virology. Oxford: Elsevier; 2011. pp. 467–481. [Google Scholar]
- 152.Blitvich BJ. Transmission dynamics and changing epidemiology of West Nile virus. Anim Health Res Rev. 2008;9:71–86. doi: 10.1017/S1466252307001430. [DOI] [PubMed] [Google Scholar]
- 153.Iwamoto M, Jernigan DB, Guasch A, et al. Transmission of West Nile virus from an organ donor to four transplant recipients. N Engl J Med. 2003;348:2196–2203. doi: 10.1056/NEJMoa022987. [DOI] [PubMed] [Google Scholar]
- 154.Stramer SL, Fang CT, Foster GA, Wagner AG, Brodsky JP, Dodd RY. West Nile virus among blood donors in the United States, 2003 and 2004. N Engl J Med. 2005;353:451–459. doi: 10.1056/NEJMoa044333. [DOI] [PubMed] [Google Scholar]
- 155.Arroyo J, Miller C, Catalan J, et al. ChimeriVax-West Nile virus live-attenuated vaccine: preclinical evaluation of safety, immunogenicity, and efficacy. J Virol. 2004;78:12497–12507. doi: 10.1128/JVI.78.22.12497-12507.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Reisen WK. Epidemiology of St. Louis encephalitis virus. Adv Virus Res. 2003;61:139–183. doi: 10.1016/s0065-3527(03)61004-3. [DOI] [PubMed] [Google Scholar]
- 157.Campbell GL, Eldridge BF, Reeves WC, Hardy JL. Isolation of Jamestown Canyon virus from boreal Aedes mosquitoes from the Sierra Nevada of California. Am J Trop Med Hyg. 1991;44:244–249. doi: 10.4269/ajtmh.1991.44.244. [DOI] [PubMed] [Google Scholar]
- 158.Fulhorst CF, Hardy JL, Eldridge BF, Chiles RE, Reeves WC. Ecology of Jamestown Canyon virus (Bunyaviridae: California serogroup) in coastal California. Am J Trop Med Hyg. 1996;55:185–189. doi: 10.4269/ajtmh.1996.55.185. [DOI] [PubMed] [Google Scholar]
- 159.Hardy JL, Eldridge BF, Reeves WC, Schutz SJ, Presser SB. Isolations of Jamestown canyon virus (Bunyaviridae: California serogroup) from mosquitoes (Diptera: Culicidae) in the western United States, 1990-1992. J Med Entomol. 1993;30:1053–1059. doi: 10.1093/jmedent/30.6.1053. [DOI] [PubMed] [Google Scholar]
- 160.Beaty BJ, Calisher CH. Bunyaviridae: natural history. Curr Top Microbiol Immunol. 1991;169:27–78. [PubMed] [Google Scholar]
- 161.Grimstad P. California group viruses. In: Monath TP, editor. The arboviruses: epidemiology and ecology. Boca Raton: CRC Press; 1988. pp. 99–136. [Google Scholar]
- 162.Bardos V, Danielova V. The Tahyna virus: a virus isolated from mosquitoes in Czechoslovakia. J Hyg Epidemiol Microbiol Immunol. 1959;3:264–276. [PubMed] [Google Scholar]
- 163.Kuno G, Chang GJ, Tsuchiya KR, Karabatsos N, Cropp CB. Phylogeny of the genus Flavivirus. J Virol. 1998;72:73–83. doi: 10.1128/jvi.72.1.73-83.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Dobler G. Zoonotic tick-borne flaviviruses. Vet Microbiol. 2010;140:221–228. doi: 10.1016/j.vetmic.2009.08.024. [DOI] [PubMed] [Google Scholar]
- 165.Gritsun TS, Nuttall PA, Gould EA. Tick-borne flaviviruses. Adv Virus Res. 2003;61:317–371. doi: 10.1016/s0065-3527(03)61008-0. [DOI] [PubMed] [Google Scholar]
- 166.Bock HL, Klockmann U, Jungst C, Schindel-Kunzel F, Theobald K, Zerban R. A new vaccine against tick-borne encephalitis: initial trial in man including a dose-response study. Vaccine. 1990;8:22–24. doi: 10.1016/0264-410x(90)90172-i. [DOI] [PubMed] [Google Scholar]
- 167.Klockmann U, Bock HL, Franke V, Hein B, Reiner G, Hilfenhaus J. Preclinical investigations of the safety, immunogenicity and efficacy of a purified, inactivated tick-borne encephalitis vaccine. J Biol Stand. 1989;17:331–342. doi: 10.1016/s0092-1157(89)80004-6. [DOI] [PubMed] [Google Scholar]
- 168.Hubalek Z, Rudolf I. Tick-borne viruses in Europe. Parasitol Res. 2012;111:9–36. doi: 10.1007/s00436-012-2910-1. [DOI] [PubMed] [Google Scholar]
- 169.Yu XL, Jiang XL, Wang T, et al. Establishment of minireplicon system for severe fever with thrombocytopenia syndrome bunyavirus. Bing Du Xue Bao. 2012;28:246–251. [PubMed] [Google Scholar]
- 170.Jiang XL, Wang XJ, Li JD, et al. Isolation, identification and characterization of SFTS bunyavirus from ticks collected on the surface of domestic animals. Bing Du Xue Bao. 2012;28:252–257. [PubMed] [Google Scholar]
- 171.Kim KH, Yi J, Kim G, et al. Severe fever with thrombocytopenia syndrome, South Korea, 2012. Emerg Infect Dis. 2013;19:1892–1894. doi: 10.3201/eid1911.130792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Monath TP, McCarthy K, Bedford P, et al. Clinical proof of principle for ChimeriVax: recombinant live, attenuated vaccines against flavivirus infections. Vaccine. 2002;20:1004–1018. doi: 10.1016/s0264-410x(01)00457-1. [DOI] [PubMed] [Google Scholar]
- 173.Monath TP, Guirakhoo F, Nichols R, et al. Chimeric live, attenuated vaccine against Japanese encephalitis (ChimeriVax-JE): phase 2 clinical trials for safety and immunogenicity, effect of vaccine dose and schedule, and memory response to challenge with inactivated Japanese encephalitis antigen. J Infect Dis. 2003;188:1213–1230. doi: 10.1086/378356. [DOI] [PubMed] [Google Scholar]