

Abstract
Background
Phages infecting spoilage microorganisms have been considered as alternative biocontrol agents, and the study of their genomes is essential to their safe use in foods. UFV-P2 is a new Pseudomonas fluorescens-specific phage that has been tested for its ability to inhibit milk proteolysis.
Results
The genome of the phage UFV-P2 is composed of bidirectional modules and presented 75 functionally predict ORFs, forming clusters of early and late transcription. Further genomic comparisons of Pseudomonas-specific phages showed that these viruses could be classified according to conserved segments that appear be free from genome rearrangements, called locally collinear blocks (LCBs). In addition, the genome organization of the phage UFV-P2 was shown to be similar to that of phages PaP3 and LUZ24 which have recently been classified as a Luz24likevirus.
Conclusions
We have presented the functional annotation of UFV-P2, a new Pseudomonas fluorescens phage. Based on structural genomic comparison and phylogenetic clustering, we suggest the classification of UFV-P2 in the Luz24likevirus genus, and present a set of shared locally collinear blocks as the genomic signature for this genus.
Background
According to the International Committee of Virus Taxonomy (ICTV) classification scheme based on morphology, biological characteristics and genome organization (http://www.ictvonline.org/virusTaxonomy.asp), the bacteriophage family Podoviridae contains two subfamilies and 11 genera, and the Luz24likevirus genus comprises the Pseudomonas-infecting bacteriophages PaP3 [1] and LUZ24 [2]. Beyond PaP3 and LUZ24, the phages tf [3], MR299-2 [4], PaP4(KC294142), vB_PaeP_p2-10_Or1 (HF543949) and vB_PaeP_C1-14_Or (HE983844) have similar genomic compositions and should be classified to this genus.
Pseudomonas fluorescens bacteriophage UFV-P2 [5], is a virus with a high ability to reduce casein proteolysis in milk. Milk proteolysis is caused by thermo-resistant enzymes produced by psychrotrophs and is responsible for serious losses in the dairy industry due to negative effects on the quality and reduced shelf life of dairy products. In this environment, Pseudomonas spp. are prevalent contaminants [6-8], mainly P. fluorescens[9,10]. The use of phages in biocontrol has been suggested as an alternative to the use of chemicals. For example, P. fluorescens-specific phages had been studied to control Pseudomonas population and as sanitation agents to efficiently remove bacterial biofilms on stainless steel surfaces similar to those used in food industries, where these contaminants are common [11-13]. However, they must be used with caution. In addition to proteolysis reduction and biofilm inhibition studies and, their host range determination, it is necessary to understand phages’ genome and proteome to make possible their use as biocontrol agents.
To expand our understanding about the P. fluorescens-specific phage UFV-P2, we present in detail the analysis of its structural genome and its comparisons to other phage genomes.
Methods
Sampling
The phage UFV-P2 was isolated from wastewater of a dairy industry in Minas Gerais, Brazil, and propagated at 30°C in LB medium in a strain of P. fluorescens 07A, courtesy of Laboratory of Food Microbiol, located at the Federal University of Viçosa, Brazil.
Genome extraction and composition
Phages were propagated in LB medium containing the bacteria in exponential phase. After incubation at 30°C for 8 h, particle assemble was induced with mitomicin and the virions were recovered by centrifugation and filtration. Phage suspensions were incubated with 75 μg/mL of proteinase K in the presence of 0.01% SDS at 56°C for 90 min. Proteins were removed by extraction with phenol, phenol:chloroform (1:1), followed by chloroform. Genetic material was concentrated with an equal volume of isopropanol and resuspended in 30 μL of distilled water. For analysis of viral genome composition, 5 μL of the genomic extracts were submitted to digestion assays with enzymes DNase I (50 μg/mL) or RNaseA (100 μg/mL) for 60 min at 37°C, followed by 1% agarose gel electrophoresis and visualization by staining with GelRed (Biotium, USA).
Genomic DNA sequencing and assembly
UFV-P2 genome was sequenced using an Illumina Genome Analyzer II by CD Genomics (New York, USA) and was assembled and analyzed using CLC Genomics Workbench version 5.1 (CLC bio, Cambridge, MA, USA). The sequence reads were assembled into contigs using stringent parameters, in which 90% of each read had to cover the other read with 90% identity. The data are available in GenBank database under accession number JX863101.
Bioinformatics analysis
The genome of phage UFV-P2 was oriented to be collinear with that of the type species, Pseudomonas phage LUZ24, and manually annotated using Kodon (Applied Maths, Austin, TX, USA.) [14]. The GenBank flat file (gbk) file was exported from Kodon and converted to FASTA-formatted protein sequences using gbk2faa (http://lfz.corefacility.ca/gbk2faa/). The latter were screened for viral homologs using the BLASTP feature of Geneious R6.1 (Biomatters Ltd., Auckland, New Zealand); and, for protein motifs, using TMHMM [15], Phobius [16] and Batch Web CD-Search Tool [17] at http://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi.
Putative promoters were identified using the Kodon sequence similarity search feature employing TTGACA(N15-18)TATAAT and allowing for a 2 bp mismatch. Rho-independent terminators were tentatively identified using ARNold [18,19] at http://rna.igmors.u-psud.fr/toolbox/arnold/index.php.
For comparative purposes at the genomic level EMBOSS Stretcher [20] and progressive Mauve [21] were employed; while at the proteomic level we used CoreGenes [22,23]. Seventeen genomic reference sequences of phages were downloaded from GenBank (Table 1) and compared to UFV-P2 genome.
Table 1.
Pairwise comparisons of phage UFV-P2 and others phage genomes
| |
|
|
|
Phage UFV-P2 |
|
|---|---|---|---|---|---|
| Phage | GenBank accession | GC content (%) | Genome density (genes/kbp) | Identities | % |
|
UFV-P2 |
JX863101 |
51.5 |
1.65 |
- |
- |
|
vB_PaeP_p2-10_Or1 |
HF543949 |
52.0 |
1.32 |
27,253 |
57.46 |
|
vB_PaeP_C1-14_Or |
HE983844 |
52.0 |
1.41 |
27,672 |
57.31 |
|
LUZ24 |
NC_010325 |
52.2 |
1.49 |
27,510 |
56.80 |
|
PaP4 |
KC294142 |
52.5 |
1.59 |
27,015 |
56.73 |
|
PaP3 |
NC_004466 |
52.2 |
1.56 |
27,358 |
56.20 |
|
MR299-2 |
JN254801 |
52.0 |
1.52 |
27,192 |
56.19 |
|
Tf |
NC_017971 |
53.2 |
1.51 |
23,750 |
49.55 |
|
Phi-2 |
NC_013638 |
58.9 |
1.00 |
22,672 |
46.73 |
|
phiKMV |
NC_005045 |
62.3 |
1.13 |
22,392 |
46.43 |
|
phiIBB-PF7A |
NC_015264 |
56.3 |
1.27 |
22,150 |
46.41 |
|
Bf7 |
NC_016764 |
58.4 |
1.15 |
21,825 |
46.16 |
|
PaP2 |
NC_005884 |
45.4 |
1.32 |
22,523 |
46.11 |
|
119X |
NC_007807 |
44.9 |
1.29 |
22,434 |
46.03 |
|
T7 (
Enterobacteria
) |
NC_001604 |
48.4 |
1.50 |
21,602 |
45.66 |
|
gh-1 |
NC_004665 |
57.4 |
1.12 |
21,218 |
45.35 |
|
F116 |
NC_006552 |
63.2 |
1.07 |
27,102 |
41.20 |
| LUZ7 | NC_013691 | 53.2 | 1.54 | 29,160 | 38.74 |
Phylogenetic clustering
For clustering UFV-P2 phage in an evolutionary way, a phylogenetic hypothesis was inferred by Bayesian inference (BI) using MrBayes v3.2.2 [24]. The genomic sequences of phages were aligned using ClustalW [25], and a pairwise distance matrix was calculated MEGA version 5 [26] (Table 1). The alignment was manually inspected, and the sites with gaps were excluded. To expedite the construction of phylogenetic trees, a model of nucleotide substitution was estimated using the jModelTest 2 program [27]. The GTR + G substitution model was selected as the best DNA evolution model for genomic sequences, according to the Akaike Information Criterion (AIC) and Bayesian Information Criterion (BIC).
The BI phylogenetic tree was calculated using the Bayesian Markov Chain Monte Carlo (MCMC) method, in two runs with 5,000,000 generations. The convergence of the parameters was analyzed in TRACER v1.5.0 (http://beast.bio.ed.ac.uk/tracer), and the chains reached a stationary distribution after 50,000 generations. Then, a total of 1% of the generated trees was burned to produce the consensus tree. To root the phylogenetic tree, the Enterobacteria phage T7 (NC_001604) was selected as outgroup taxa.
Results and Discussion
Transmission electron microscopy of the UFV-P2 virions (data not shown) showed that this virus has isometric capsids and very short tails, with morphological similarity to the P. aeruginosa phages Pap3 and MR299-2. Thus, UFV-P2 can be inserted in the Podoviridae family, order Caudovirales.
Functional genomic organization
The viral genome was extracted and sequenced
The phage UFV-P2 has a linear 45,517 bp DNA genome with a GC content of 51.5%, and was sequenced with coverage of 30,655 fold. One of the interesting characteristics of members of the Luz24likevirus genus is the presence of localized single-stranded breaks associated with the consensus sequence TACTRTGMC [28]. Fourteen of these sequences were found in the top strand of the tf DNA, while the genome of UFV-P2 contains 15.
At first, bioinformatics analyses had showed that the UFV-P2 genome has a bidirectional organization with 92 predicted open reading frames (ORFs) larger than 100 bp, but only 41 ORFs (44.75%) could be identified as coding sequences (CDS) by similarity searches against known proteins in the GenBank and UniProt databases [5]. However, we propose a new annotation of the genome of this virus based on different tools, which were able to functionally predict 75 ORFs also bidirectionally oriented and forming clusters of early and late transcription (Figure 1 and Table 2).
Figure 1.
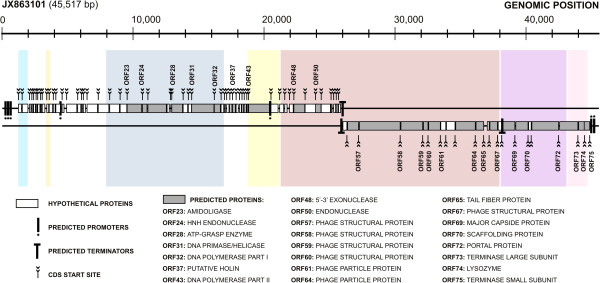
The genomic organization of the phage UFV-P2. The colored squares in the background correspond to the conserved locally collinear blocks (LCB) found for the UFV-P2 in relation to the other members of the Luz24likevirus genus (Figure 2). The detailed annotation is shown in Table 2.
Table 2.
Functional genomic annotation of phage UFV-P2
| Gene | Predicted protein | Genomic coordinates | Strand | Protein mass (Da) | Protein pI | AA residues | Homologs (a) & Motifs | E-value |
|---|---|---|---|---|---|---|---|---|
| ORF01 |
Conserved hypothetical protein |
1205..1483 |
+ |
10486 |
10.5 |
92 |
NP_955002 hypothetical protein PaP3p71 [Pseudomonas phage PaP3] |
7.34E-09 |
| ORF02 |
Conserved hypothetical protein |
1501..1983 |
+ |
17752 |
9.4 |
160 |
YP_006382463 hypothetical protein tf_02 [Pseudomonas phage tf]; protein motifs: cl10333 PHA01782 |
3.22E-37 |
| ORF03 |
Conserved hypothetical protein |
2046..2216 |
+ |
6515 |
7.8 |
56 |
YP_007183264 hypothetical protein BN425_ORF_56 [Pseudomonas phage vB_PaeP_p2-10_Or1] |
7.83E-02 |
| ORF04 |
Hypothetical protein |
2231..2368 |
+ |
5276 |
9.8 |
45 |
- |
- |
| ORF05 |
Hypothetical protein |
2365..2556 |
+ |
7171 |
9.2 |
63 |
- |
- |
| ORF06 |
Hypothetical membrane protein |
2553..2687 |
+ |
4708 |
9.5 |
44 |
protein motifs: one transmembrane domain discovered using TMHMM and Phobius |
|
| ORF07 |
Conserved hypothetical protein |
2677..2898 |
+ |
8374 |
9.8 |
73 |
YP_006382470 hypothetical protein tf_10 [Pseudomonas phage tf] |
2.09E-04 |
| ORF08 |
Conserved hypothetical protein |
2898..3020 |
+ |
5178 |
9.3 |
40 |
AGC35239 hypothetical protein PaP4_008 [Pseudomonas phage PaP4] |
1.19E-05 |
| ORF09 |
Hypothetical protein |
3021..3275 |
+ |
9120 |
4.1 |
84 |
- |
- |
| ORF10 |
Hypothetical protein |
3368..3568 |
+ |
7493 |
9.6 |
66 |
- |
- |
| ORF11 |
Hypothetical protein |
3555..3869 |
+ |
11403 |
5.0 |
104 |
- |
- |
| ORF12 |
Hypothetical protein |
3869..4063 |
+ |
7139 |
9.0 |
64 |
- |
- |
| ORF13 |
Conserved hypothetical protein |
4060..4458 |
+ |
14906 |
9.4 |
132 |
YP_006382473 hypothetical protein tf_13 [Pseudomonas phage tf] |
9.55e-15 |
| ORF14 |
Hypothetical membrane protein |
4539..4736 |
+ |
7618 |
9.1 |
65 |
protein motifs: one to two transmembrane domains discovered using TMHMM and Phobius |
|
| ORF15 |
Conserved hypothetical protein |
4884..5714 |
+ |
29824 |
8.5 |
276 |
AGC35249 transposase fusion protein [Pseudomonas phage PaP4]; protein motifs: pfam01145 Band_7, & cl02525 Band_7 |
1.15E-113 |
| ORF16 |
Conserved hypothetical protein |
5727..6017 |
+ |
10708 |
9.1 |
96 |
YP_006659979 hypothetical protein tf_14 [Pseudomonas phage tf] |
2.29E-11 |
| ORF17 |
Conserved hypothetical protein |
6021..6149 |
+ |
4783 |
11.0 |
42 |
NP_775206 hypothetical protein PaP3p51 [Pseudomonas phage PaP3] |
1.32E-03 |
| ORF18 |
Hypothetical protein |
6159..6374 |
+ |
7803 |
5.0 |
71 |
- |
- |
| ORF19 |
Conserved hypothetical protein |
6441..7289 |
+ |
31291 |
9.2 |
282 |
YP_006382477 hypothetical protein tf_17 [Pseudomonas phage tf] |
1.75E-03 |
| ORF20 |
Conserved hypothetical protein |
7296..8159 |
+ |
32906 |
4.9 |
287 |
YP_006382478 hypothetical protein tf_18 [Pseudomonas phage tf] |
2.06E-07 |
| ORF21 |
Conserved hypothetical protein |
8171..8968 |
+ |
29423 |
5.0 |
265 |
YP_006382479 hypothetical protein tf_19 [Pseudomonas phage tf]; protein motifs: PF14395.1 COOH-NH2_lig |
8.75E-112 |
| ORF22 |
Conserved hypothetical protein |
8965..9525 |
+ |
21203 |
7.5 |
186 |
NP_775210 hypothetical protein PaP3p47 [Pseudomonas phage PaP3] |
2.53E-40 |
| ORF23 |
Amidoligase |
9501..10658 |
+ |
43407 |
5.5 |
385 |
YP_001671891 hypothetical protein [Pseudomonas phage LUZ24]; protein motifs: PF12224.3 Amidoligase_2 |
4.46E-96 |
| ORF24 |
HNH endonuclease |
10667..11071 |
+ |
15340 |
9.6 |
134 |
YP_002003475 gp2.8 [Enterobacteria phage BA14]; protein motifs: PF13392.1 HNH_3 |
2.71E-20 |
| ORF25 |
Glutamine amidotransferase |
11084..12544 |
+ |
54036 |
5.9 |
486 |
YP_006382482 glutamine amidotransferase [Pseudomonas phage tf]; protein motifs: PF13522.1 GATase_6, & cd00352 Gn_AT_II |
2.72E-151 |
| ORF26 |
Conserved hypothetical protein |
12547..12765 |
+ |
8166 |
4.8 |
72 |
NP_775213 hypothetical protein PaP3p44 [Pseudomonas phage PaP3] |
2.25E-20 |
| ORF27 |
Hypothetical protein |
12805..12909 |
+ |
3505 |
5.3 |
34 |
- |
- |
| ORF28 |
ATP-grasp enzyme |
12906..13781 |
+ |
31797 |
7.7 |
291 |
YP_006382486 hypothetical protein tf_24 [Pseudomonas phage tf] |
2.60E-96 |
| ORF29 |
Conserved hypothetical protein |
13774..14169 |
+ |
15076 |
5.5 |
131 |
AGC35259 hypothetical protein PaP4_028 [Pseudomonas phage PaP4]; proteins motifs: PF06094.7 AIG2, & cd06661 GGCT_like |
5.55E-27 |
| ORF30 |
Conserved hypothetical protein |
14169..14465 |
+ |
11295 |
5.4 |
98 |
YP_006382488 hypothetical protein tf_026 [Pseudomonas phage tf] |
6.03E-23 |
| ORF31 |
DNA primase/helicase |
14425..16176 |
+ |
65673 |
5.9 |
583 |
YP_001671897 primase/helicase [Pseudomonas phage LUZ24]; protein motifs: PF13155.1 Toprim_2, & PF03796.10 DnaB_C |
0 |
| ORF32 |
DNA polymerase part I |
16151..16666 |
+ |
20133 |
4.5 |
171 |
YP_006382490 3′-5′ exonuclease [Pseudomonas phage tf] & YP_001671898 DNA polymerase part I [Pseudomonas phage LUZ24] |
2.82E-75; 3.72e-68 |
| ORF33 |
Conserved hypothetical membrane protein |
16669..16920 |
+ |
9085 |
10.0 |
83 |
YP_006382491 hypothetical protein tf_29 [Pseudomonas phage tf]; protein motifs: one transmembrane domain discovered using TMHMM and Phobius |
1.99E-06 |
| ORF34 |
Hypothetical protein |
16931..17068 |
+ |
5096 |
3.4 |
45 |
- |
- |
| ORF35 |
Hypothetical protein |
17095..17325 |
+ |
8505 |
9.0 |
76 |
- |
- |
| ORF36 |
Conserved hypothetical protein |
17336..17539 |
+ |
7596 |
5.8 |
67 |
YP_001671900 hypothetical protein [Pseudomonas phage LUZ24] |
2.23E-02 |
| ORF37 |
Putative holin |
17536..17817 |
+ |
10475 |
9.8 |
93 |
YP_006382493 hypothetical protein tf_32 [Pseudomonas phage tf] & YP_001671904 putative holin [Pseudomonas phage LUZ24]; protein motifs: three transmembrane domains discovered using TMHMM and Phobius |
5.03e-35; 1.92e-18 |
| ORF38 |
Hypothetical membrane protein |
17814..18032 |
+ |
8079 |
9.9 |
72 |
protein motifs: one or two transmembrane domains discovered using TMHMM and Phobius |
|
| ORF39 |
Conserved hypothetical protein |
18069..18263 |
+ |
6728 |
3.5 |
64 |
YP_006382494 hypothetical protein tf_34 [Pseudomonas phage tf] |
6.48E-04 |
| ORF40 |
Conserved hypothetical protein |
18263..18433 |
+ |
5628 |
8.2 |
56 |
YP_007112538 hypothetical protein MAR_61 [Vibrio phage vB_VpaM_MAR] |
9.20E-05 |
| ORF41 |
Conserved hypothetical protein |
18433..18627 |
+ |
7272 |
10.9 |
64 |
YP_006382495 hypothetical protein tf_35 [Pseudomonas phage tf] |
1.52E-03 |
| ORF42 |
Hypothetical protein |
18630..18764 |
+ |
4887 |
6.0 |
44 |
- |
- |
| ORF43 |
DNA polymerase part II |
18765..20405 |
+ |
60865 |
9.6 |
546 |
YP_007183240 DNA polymerase1 [Pseudomonas phage vB_PaeP_p2-10_Or1] & YP_001671907 DNA polymerase part II [Pseudomonas phage LUZ24]; protein motifs: PF00476.15 DNA_pol_A |
0; 0 |
| ORF44 |
DNA binding protein |
20472..21068 |
+ |
21853 |
4.8 |
198 |
YP_006382500 DNA binding protein [Pseudomonas phage tf] |
6.61E-70 |
| ORF45 |
Conserved hypothetical protein |
21139..21495 |
+ |
13200 |
6.3 |
118 |
AGC35272 hypothetical protein PaP4_041 [Pseudomonas phage PaP4] |
4.02E-47 |
| ORF46 |
Hypothetical protein |
21525..21755 |
+ |
8335 |
4.7 |
76 |
- |
- |
| ORF47 |
Hypothetical protein |
21887..22222 |
+ |
12649 |
9.4 |
111 |
- |
- |
| ORF48 |
5′-3′ exonuclease |
22219..23103 |
+ |
33498 |
5.3 |
294 |
NP_775229 exonuclease [Pseudomonas phage PaP3]; protein motifs: PF01367.15 5_3_exonuc, & cd09898 H3TH_53EXO |
1.47E-149 |
| ORF49 |
Conserved hypothetical protein |
23078..24073 |
+ |
37154 |
4.7 |
331 |
YP_006659984 conserved hypothetical protein [Pseudomonas phage tf] |
1.15E-39 |
| ORF50 |
Endonuclease |
23874..24341 |
+ |
17655 |
5.4 |
155 |
YP_006382505 endonuclease [Pseudomonas phage tf] |
1.11E-41 |
| ORF51 |
Conserved hypothetical protein |
24307..25062 |
+ |
28981 |
6.2 |
251 |
YP_001671917 hypothetical protein [Pseudomonas phage LUZ24] |
2.15E-129 |
| ORF52 |
Hypothetical protein |
25059..25184 |
+ |
4844 |
10.5 |
41 |
- |
- |
| ORF53 |
Conserved hypothetical protein |
25255..25458 |
+ |
7717 |
6.5 |
67 |
YP_006659986 hypothetical protein tf_48 [Pseudomonas phage tf] |
7.14E-14 |
| ORF54 |
Hypothetical protein |
25455..25631 |
+ |
6814 |
4.6 |
58 |
- |
- |
| ORF55 |
Conserved hypothetical protein |
25628..25834 |
+ |
8109 |
4.4 |
68 |
YP_001671920 hypothetical protein [Pseudomonas phage LUZ24] |
1.57E-12 |
| ORF56 |
Conserved hypothetical protein |
25963..26319 |
- |
13124 |
5.9 |
118 |
AGC35282 hypothetical protein PaP4_051 [Pseudomonas phage PaP4] |
2.11E-45 |
| ORF57 |
Phage structural protein |
26321..27208 |
- |
32029 |
5.5 |
295 |
AFD10698 hypothetical protein I7C_020 [Pseudomonas phage MR299-2] |
3.43E-152 |
| ORF58 |
Phage structural protein |
27219..30383 |
- |
111420 |
5.3 |
1054 |
YP_001671923 phage particle protein [Pseudomonas phage LUZ24] |
0 |
| ORF59 |
Phage structural protein |
30389..32101 |
- |
60235 |
5.5 |
570 |
YP_001671924 phage particle protein [Pseudomonas phage LUZ24] |
1.35E-86 |
| ORF60 |
Phage structural protein |
32103..32507 |
- |
13485 |
6.5 |
134 |
YP_001671925 phage particle protein [Pseudomonas phage LUZ24] |
9.26E-11 |
| ORF61 |
Phage particle protein |
32504..33448 |
- |
32240 |
4.8 |
314 |
AFD10694 hypothetical protein I7C_016 [Pseudomonas phage MR299-2] |
3.23E-112 |
| ORF62 |
Conserved hypothetical protein |
33429..33863 |
- |
16522 |
4.5 |
144 |
AGC35288 hypothetical protein PaP4_057 [Pseudomonas phage PaP4] |
2.95E-57 |
| ORF63 |
Conserved hypothetical protein |
33860..34570 |
- |
25585 |
4.8 |
236 |
YP_001671928 hypothetical protein [Pseudomonas phage LUZ24] |
3.73E-56 |
| ORF64 |
Phage particle protein |
34570..36111 |
- |
57542 |
5.1 |
513 |
YP_006382515 phage particle protein [Pseudomonas phage tf] & YP_001671929 hypothetical protein [Pseudomonas phage LUZ24] |
0; 0 |
| ORF65 |
Tail fiber protein |
36119..36754 |
- |
21879 |
6.7 |
211 |
NP_775246 hypothetical protein PaP3p12 [Pseudomonas phage PaP3] |
1.68E-62 |
| ORF66 |
Conserved hypothetical protein |
36976..37170 |
- |
6838 |
6.6 |
64 |
YP_001671932 hypothetical protein LUZ24 [Pseudomonas phage LUZ24] |
3.02E-28 |
| ORF67 |
phage structural protein |
37175..37804 |
- |
23887 |
5.0 |
209 |
AFD10687 putative constituent protein [Pseudomonas phage MR299-2] |
2.54E-82 |
| ORF68 |
Conserved hypothetical protein |
37808..38128 |
- |
11847 |
5.8 |
106 |
YP_007183215 hypothetical protein BN425_ORF_07 [Pseudomonas phage vB_PaeP_p2-1 0_Or1] |
1.26E-50 |
| ORF69 |
Major capsid protein |
38180..39133 |
- |
34740 |
6.0 |
317 |
NP_775251 major head protein [Pseudomonas phage PaP3] & YP_001671935 major head protein [Pseudomonas phage LUZ24] |
0; 0 |
| ORF70 |
Scaffolding protein |
39152..40138 |
- |
36190 |
4.3 |
328 |
YP_001671936 scaffolding protein [Pseudomonas phage LUZ24] |
4.90E-101 |
| ORF71 |
Conserved hypothetical protein |
40128..40379 |
- |
9559 |
5.2 |
83 |
AGC35298 hypothetical protein PaP4_067 [Pseudomonas phage PaP4] & YP_001671937 hypothetical protein [Pseudomonas phage LUZ24] |
8.9E-30; 2.00e-29 |
| ORF72 |
Portal protein |
40379..42469 |
- |
79405 |
5.0 |
696 |
YP_007183212 putative portal protein [Pseudomonas phage vB_PaeP_p2-10_Or1] YP_001671938 portal protein [Pseudomonas phage LUZ24] |
0; 0 |
| ORF73 |
Terminase, large subunit |
42496..43941 |
- |
54330 |
6.0 |
481 |
YP_001671939 terminase large subunit [Pseudomonas phage LUZ24]; protein motifs: PF03237.10 Terminase_6 |
0 |
| ORF74 |
Lysozyme |
43945..44451 |
- |
18961 |
8.6 |
168 |
YP_006382529 lysozyme [Pseudomonas phage tf]; protein motifs: PF00959.14 Phage_lysozyme |
3.27E-56 |
| ORF75 | Terminase, small subunit | 44372..44845 | - | 17291 | 5.9 | 157 | YP_007183209 hypothetical protein BN425_ORF_01 [Pseudomonas phage vB_PaeP_p2-1 0_Or1] & YP_006382530 terminase small subunit [Pseudomonas phage tf] | 5.76E-66; 2.59e-64 |
The searches for consensus sequences of transcriptional promoters revealed the presence of seven promoters, five in the positive strand initiating the transcription of ORFs that encode early proteins, which is a common feature of viral genomes that need bacterial transcription factors to start their infection cycle. The two other promoters are located in late genes modules. These genes are usually transcribed by viral transcription factors.
Three rho-independent transcription terminators were predicted using ARNold, one in the positive and two in the negative strand (Figure 1). A bidirectional termination region was found in the region from 25,922 to 25,964. Interestingly, this pattern of termination is also found in the genomes of the phages PaP3 [1] and LUZ24 [2]. The last terminator sequence is located at the terminal end of the gene encoding the major head protein. The low number of sequences of rho-independent terminators compared to the number of predicted ORFs may be due to the existence of other types of terminators or the presence of transcriptional modules and the generation of polycistronic mRNAs, a very common feature of viral genomes.
The predicted UFV-P2 genes were functionally classified as its promoters, predicted order of transcription, and its annotated functions.
Nucleotide biosynthesis and DNA replication (positive-stranded ORFs)
Fifty-five genes (ORFs 01–55) involved in nucleotide biosynthesis and viral replication process were found in the UFV-P2 genome positive strand, named early genes (Figure 1). Among viral replication genes, ORF31 encodes a primase/helicase; ORF44, a DNA-binding protein; ORF48, a 5′-3′ exonuclease; ORF50, a putative endonuclease; and ORFs 32 and 43 encode the two exons of the viral DNA polymerase, between which there is an ORF encoding a putative holin with three transmembrane domains similar to those from the phages tf and LUZ24. Holins are small membrane proteins that accumulate in the membrane until, at a specific time that is “programmed” into the holin, the membrane suddenly becomes permeabilized to the fully folded endolysin [29]. In addition, the UFV-P2 genome contains two endonucleases encoded by ORF24 and ORF50. The first is a HNH endonuclease, a group I homing endonuclease, which may be related to the presence of introns in the UFV-P2 genome [30], like those between the two parts of DNA polymerase. Other enzymes predicted in the positive strand include ORFs 23, 25 and 28, which encode, respectively, an amidoligase, a glutamine amidotransferase and an ATP-grasp enzyme. The other 45 proteins of the early genes module are hypothetical proteins.
Virion assembly and host lysis (negative-stranded ORFs)
Twenty genes (ORFs 56–75) related to the composition and assembly of the viral particle, DNA packaging, and host lysis were found in the UFV-P2 genome negative strand, named late genes (Figure 1). Two transcriptional modules were found based on predicted terminators. The first is located in the regions comprising the ORFs 75–69, and the second module corresponding to the ORFs 75–56.
In the first module, ORF75 and ORF73 encode the small and large terminase subunits, respectively. The terminase is the motor component that assists the translocation of viral genomic DNA to the inner of the capsid during packaging via ATP hydrolysis. There is an ongoing discussion about the role of terminase structure in determining the points for cleavage of the viral DNA, which would influence the entire viral genome organization [31]. Recently, Shen and coworkers [32] functionally identified the two genes encoding PaP3 terminase subunits, located in ORFs 1 and 3, respectively, which have high sequence similarity with ORFs 75 and 73 of the UFV-P2 genome. The PaP3 genome have been annotated as opposing transcriptional gene clusters in relation to the UFV-P2 genome, what explains the difference observed for the numbering of similar ORFs. The same occurred for the earlier annotation of phage UFV-P2 [5], which is revised in this work to correspond to the annotation of phage LUZ24, which represents the genus.
ORF72 encodes the portal protein; ORF69 encodes the major head protein; and ORF70 encodes a scaffolding protein, which is a chaperone possibly related to viral particle assembly. In the second module, beyond the ORFs from the first, the ORFs 57–61, 64 and 67 encode particle/structural proteins; ORF65 encodes the tail fiber protein; and the other six ORFs encode hypothetical proteins.
ORF74 encodes a lysozyme that is used in the process of host cell breakage through the lysis of the peptidoglycan layer. The occurrence of a lysin, not associated with its cognate holin, is unusual but also found in other members of the Luz24likevirus genus.
Structural genomic comparisons and evolutionary clustering
Pairwise genomic comparisons has been a useful approach for genotyping and classification of viruses like Circoviridae[33] and Geminiviridae[34]. The alignment of phages genomic sequences and pairwise comparisons revealed that vb_PaeP_p2-10_Or1, vb_PaeP_C1-14_Or, LUZ24, PaP4, PaP3, MR299-2 and tf are the phages most closely related to UFV-P2. Genomic sequences of these phages presented an identity to the UFV-P2 genome ranging from 49.5% to 57.5% (see Table 1).
The structural genomic comparisons in Mauve showed that these phages shared a set of conserved locally collinear blocks (LCB) (Figure 2 and Additional file 1: Figure S2). LCBs are conserved segments that appear be free from genome rearrangements, since the orthologous regions of genomes can be reordered or inverted by recombination processes [21]. In addition, a specific comparison between UFV-P2 and LUZ24 showed colinearity across their genomes (Figures 2 and 3).
Figure 2.
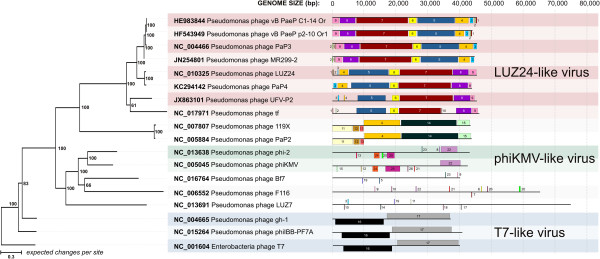
Phylogenetic clustering and structural genomic comparisons among the UFV-P2 and other phages. Phylogenetic tree of phage genomes (left) was calculated by Bayesian MCMC coalescent analysis. The posterior probability values (PP) (expressed as percentages) calculated using the best trees found by MrBayes are shown beside each node. The outgroup taxon is the Enterobacteria phage T7 (NC_001604). The colored squares in the schematic view of genomes (right) correspond to the conserved locally collinear blocks (LCBs) predicted by Mauve. The numbers and colors indicate the LCBs that are shared between the phages genomes.
Figure 3.
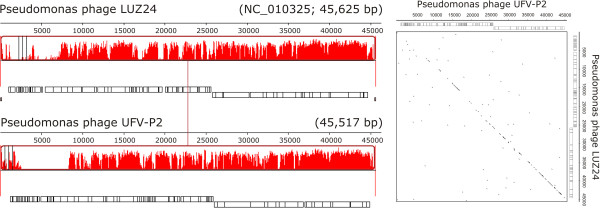
Comparison of the genomes of the phages UFV-P2 and LUZ24. The collinearity between genomes is represented by the conserved locally collinear block (left) and Dot plot alignment (right). Dot plot alignment was calculated using Nucleic Acid Dot Plots (http://www.vivo.colostate.edu/molkit/dnadot/index.html), considering a window size of 13 and a mismatch limit of 0.
Phages LUZ24, PaP4, and UFV-P2 present a conserved bidirectional genomic organization, which is showed by the shared LCBs (blocks 3–9) (Figure 2). Phage tf also presents this organization, but with some differences in the shared LCBs. On the other hand, phages MR299-2, PaP3, vb_PaeP_p2-10_Or1, and vb_PaeP_C1-14_Or present an inverted set of LCBs (blocks 9–3), representing an opposing arrangement of the gene modules. Proteins of these seven phages were the top hits with the UFV-P2 sequences (Table 2) and can collaborate with each other’s functional annotations. In addition to genomic comparisons, a search for direct terminal repeats (DTRs) indicated the presence of patterns at the ends of the UFV-P2 genome, as described for the phages LUZ24, tf, and vB_PaeP_C1-14_Or1. These repeats are responsible for the recognition and cleavage of the phage genome at the end of the repeat region during packaging. Interestingly, one of the unique features of this group of phages is that PaP3 possesses 20 bp 5′-protuding cohesive ends [1], while LUZ24 has 184 bp DTRs, yet there does not appear to be a significant difference in the amino acid sequence of their terminases.
As suggested by the structural genomic comparisons, phylogenetic tree of genomic sequences grouped the phages according the shared LCBs (Figure 2). Phages PaeP_p2-10_Or1, vb_PaeP_C1-14_Or, LUZ24, PaP4, PaP3, MR299-2, tf, and UFV-P2 were included in a distinct monophyletic clade in BI phylogenetic tree, which possibly represents the Luz24likevirus genus. The shared LCBs, blocks 3–9 (Figure 2), may be considered as a genomic signature for this genus. In UFV-P2 genome (Figure 1), as for the other phages, the genes for biosynthesis and DNA replication are included in blocks 5 and 6; genes for virion structure and assembly are in blocks 7 and 8; and genes for host lysis are block 9. In blocks 3 and 4 are included only hypothetical genes. Then, we propose the classification of the phage UFV-P2 in the Luz24likevirus genus. In fact, these analyzes showed that other viruses were also grouped in distinct monophyletic clades or according to specific shared locally collinear blocks (LCB), as those from the T7likevirus (blocks 16 and 17) and Phikmvlikevirus (blocks 22, 24, and 25) genera, beyond a possibly genus including the phages PaP2 and 199X (blocks 4 and 11–15).
Conclusions
We have presented the functional annotation of UFV-P2, a new Pseudomonas fluorescens phage. Based on structural genomic comparison and phylogenetic clustering, we suggest the classification of UFV-P2 in the Luz24likevirus genus, and present a set of shared locally collinear blocks as the genomic signature for this genus.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
MRE carried out the phage isolation, propagation, DNA extration, participated in the sequence alignment and drafted the manuscript. PMPV and RLS carried out the bioinformatic analysis and drafted the manuscript. MPA and RSD participated in the design of the study and initial processes of phage isolation, propagation, DNA extration and analysis. CCS and AFC were essential on the concept of the study, and participated in its design and coordination. AK participated in the bioinformatic analysis and concepts of phage classification. SOP is the responsible for the design and coordination of the project. All authors read and approved the final manuscript.
Supplementary Material
Transmission Electron Microscopy of the phage UFV-P2. Virions have isometric capsids of 40-50 nm and very short tails (arrows). Scale bars = 100 nm. Figure S2. Comparison of the genomes of phages classified in LUZ24likevirus genus. The collinearity among genomes is represented by the conserved locally collinear blocks (LCBs). In the main block (blue), the regions of similarity plot with high identity corresponds to the set of shared LCBs (see Figure 2). The connection line between blocks correspond to the central point of LCB of the reference genome (phage LUZ24 genome).
Contributor Information
Monique R Eller, Email: monique.eller@gmail.com.
Pedro M P Vidigal, Email: pmvidigal@gmail.com.
Rafael L Salgado, Email: rafalocsalg@gmail.com.
Maura P Alves, Email: maurapalves@gmail.com.
Roberto S Dias, Email: rosousa318@yahoo.com.br.
Cynthia C da Silva, Email: ccanedosilva@yahoo.com.br.
Antônio F de Carvalho, Email: afc1800@yahoo.com.
Andrew Kropinski, Email: kropinsk@queensu.ca.
Sérgio O De Paula, Email: depaula@ufv.br.
Acknowledgments
This study was supported by grants from the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). The funders had no role in the study design, data collection, analysis, decision to publish, or preparation of this manuscript.
References
- Tan Y, Zhang K, Rao X, Jin X, Huang J, Zhu J, Chen Z, Hu X, Shen X, Wang L, Hu F. Whole genome sequencing of a novel temperate bacteriophage of P. aeruginosa: evidence of tRNA gene mediating integration of the phage genome into the host bacterial chromosome. Cell Microbiol. 2007;15:479–491. doi: 10.1111/j.1462-5822.2006.00804.x. [DOI] [PubMed] [Google Scholar]
- Ceyssens P-J, Hertveldt K, Ackermann H-W, Noben J-P, Demeke M, Volckaert G, Lavigne R. The intron-containing genome of the lytic Pseudomonas phage LUZ24 resembles the temperate phage PaP3. Virology. 2008;15:233–238. doi: 10.1016/j.virol.2008.04.038. [DOI] [PubMed] [Google Scholar]
- Kulakov LA, Kochetkov VV, Ksenzenko VN, Krylov VN, Boronin AM. [Physical map of the DNA of bacteriophage tf of Pseudomonas putida] Mol Gen Mikrobiol Virusol. 1988. pp. 12–16. [PubMed]
- Alemayehu D, Casey PG, McAuliffe O, Guinane CM, Martin JG, Shanahan F, Coffey A, Ross RP, Hill C. Bacteriophages φMR299-2 and φNH-4 can eliminate Pseudomonas aeruginosa in the murine lung and on cystic fibrosis lung airway cells. mBio. 2012;15:e00029–12. doi: 10.1128/mBio.00029-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eller MR, Salgado RL, Vidigal PMP, Alves MP, Dias RS, De Oliveira LL, Da Silva CC, De Carvalho AF, De Paula SO. Complete genome sequence of the Pseudomonas fluorescens bacteriophage UFV-P2. Genome Announcement. 2013;15:1 e00006–12. doi: 10.1128/genomeA.00006-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rasolofo EA, St-Gelais D, LaPointe G, Roy D. Molecular analysis of bacterial population structure and dynamics during cold storage of untreated and treated milk. Int J Food Microbiol. 2010;15:108–118. doi: 10.1016/j.ijfoodmicro.2010.01.008. [DOI] [PubMed] [Google Scholar]
- Munsch-Alatossava P, Alatossava T. Phenotypic characterization of raw milk-associated psychrotrophicbacteria. Microbiol Res. 2006;15:334–346. doi: 10.1016/j.micres.2005.12.004. [DOI] [PubMed] [Google Scholar]
- Baruzzi F, Lagonigro R, Quintieri L, Morea M, Caputo L. Occurrence of non-lactic acid bacteria populations involved in protein hydrolysis of cold-stored high moisture Mozzarella cheese. Food Microbiol. 2012;15:37–44. doi: 10.1016/j.fm.2011.10.009. [DOI] [PubMed] [Google Scholar]
- Arcuri EF, Aparecida M, Paiva V, Lange CC. Contagem, isolamento e caracterização de bactériaspsicrotróficascontaminantes de leite cru refrigerado. Ciência Rural. 2008;15:2250–2255. doi: 10.1590/S0103-84782008000800025. [DOI] [Google Scholar]
- Dogan B, Boor KJ. Genetic diversity and spoilage potentials among Pseudomonas spp. isolated from fluid milk products and dairy processing plants. Appl Environ Microbiol. 2003;15:130–138. doi: 10.1128/AEM.69.1.130-138.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baum MM, Kainović A, O’Keeffe T, Pandita R, McDonald K, Wu S, Webster P. Characterization of structures in biofilms formed by a Pseudomonas fluorescensisolated from soil. BMC Microbiol. 2009;15:103. doi: 10.1186/1471-2180-9-103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sillankorva S, Neubauer P, Azeredo J. Isolation and characterization of a T7-like lytic phage for Pseudomonas fluorescens. BMC Biotechnol. 2008;15:80. doi: 10.1186/1472-6750-8-80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sillankorva S, Neubauer P, Azeredo J. Pseudomonas fluorescens biofilms subjected to phage phiIBB-PF7A. BMC Biotechnol. 2008;15:79. doi: 10.1186/1472-6750-8-79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lingohr EJ, Villegas A, She Y-M, Ceyssens P-J, Kropinski AM. The genome and proteome of the Kluyverabacteriophage Kvp1–another member of the T7-like Autographivirinae. Virol J. 2008;15:122. doi: 10.1186/1743-422X-5-122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sonnhammer EL, Von Heijne G, Krogh A. A hidden Markov model for predicting transmembrane helices in protein sequences.Proceedings/. International Conference on Intelligent Systems for Molecular Biology; ISMB International Conference on Intelligent Systems for. Mol Biol. 1998;15:175–182. [PubMed] [Google Scholar]
- Käll L, Krogh A, Sonnhammer ELL. A combined transmembrane topology and signal peptide prediction method. J Mol Biol. 2004;15:1027–1036. doi: 10.1016/j.jmb.2004.03.016. [DOI] [PubMed] [Google Scholar]
- Derbyshire MK, Lanczycki CJ, Bryant SH, Marchler-Bauer A. Annotation of functional sites with the Conserved Domain Database. Database (Oxford) 2012;15:bar058. doi: 10.1093/database/bar058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macke TJ, Ecker DJ, Gutell RR, Gautheret D, Case DA, Sampath R. RNAMotif, an RNA secondary structure definition and search algorithm. Nucleic Acids Res. 2001;15:4724–4735. doi: 10.1093/nar/29.22.4724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gautheret D, Lambert A. Direct RNA motif definition and identification from multiple sequence alignments using secondary structure profiles. J Mol Biol. 2001;15:1003–1011. doi: 10.1006/jmbi.2001.5102. [DOI] [PubMed] [Google Scholar]
- Rice P, Longden I, Bleasby A. EMBOSS: the European Molecular Biology Open Software Suite. TIG. 2000;15:276–277. doi: 10.1016/S0168-9525(00)02024-2. [DOI] [PubMed] [Google Scholar]
- Darling AE, Mau B, Perna NT. progressiveMauve: multiple genome alignment with gene gain, loss and rearrangement. PloS one. 2010;15:e11147. doi: 10.1371/journal.pone.0011147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mahadevan P, King JF, Seto D. Data mining pathogen genomes using GeneOrder and CoreGenes and CGUG: gene order, synteny and in silicoproteomes. Int J Comput Biol Drug Des. 2009;15:100–114. doi: 10.1504/IJCBDD.2009.027586. [DOI] [PubMed] [Google Scholar]
- Kropinski AM, Borodovsky M, Carver TJ, Cerdeño-Tárraga AM, Darling A, Lomsadze A, Mahadevan P, Stothard P, Seto D, Van Domselaar G, Wishart DS. In silico identification of genes in bacteriophage DNA. Methods Mol Biol. 2009;15:57–89. doi: 10.1007/978-1-60327-565-1_6. [DOI] [PubMed] [Google Scholar]
- Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Hohna S, Larget B, Liu L, Suchard MA, Huelsenbeck JP. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol. 2012;15(3):539–542. doi: 10.1093/sysbio/sys029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;15:4673–4680. doi: 10.1093/nar/22.22.4673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;15:2731–2739. doi: 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darriba D, Taboada GL, Doallo R, Posada D. jModelTest 2: more models, new heuristics and parallel computing. Nat Methods. 2012;15(8):772. doi: 10.1038/nmeth.2109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glukhov AS, Krutilina AI, Shlyapnikov MG, Severinov K, Lavysh D, Kochetkov VV, McGrath JW, De Leeuwe C, Shaburova OV, Krylov VN, Akulenko NV, L a K. Genomic analysis of Pseudomonas putida phage tf with localized single-strand DNA interruptions. PloS one. 2012;15:e51163. doi: 10.1371/journal.pone.0051163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang I, Smith DL, Young R. HOLINS: The Protein Clocks of Bacteriophage Infections. Annu Rev Microbiol. 2000;15:799–825. doi: 10.1146/annurev.micro.54.1.799. [DOI] [PubMed] [Google Scholar]
- Hertveldt K, Lavigne R, Pleteneva E, Sernova N, Kurochkina L, Korchevskii R, Robben J, Mesyanzhinov V, Krylov VN, Volckaert G. Genome comparison of Pseudomonas aeruginosa large phages. J Mol Biol. 2005;15:536–545. doi: 10.1016/j.jmb.2005.08.075. [DOI] [PubMed] [Google Scholar]
- Feiss M, Rao VB. The bacteriophage DNA packaging machine. Advances in experimental medicine and biology. 2012;15:489–509. doi: 10.1007/978-1-4614-0980-9_22. [DOI] [PubMed] [Google Scholar]
- Shen X, Li M, Zeng Y, Hu X. Functional identification of the DNA packaging terminase from Pseudomonas aeruginosa phage PaP3. Arch Virol. 2012;15(11):2133–2141. doi: 10.1007/s00705-012-1409-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Segalés J, Olvera A, Grau-Roma L, Charreyre C, Nauwynck H, Larsen L, Dupont K, McCullough K, Ellis J, Krakowka S, Mankertz A, Fredholm M, Fossum C, Timmusk S, Stockhofe-Zurwieden N, Beattie V, Armstrong D, Grassland B, Baekbo P, Allan G. PCV-2 genotype definition and nomenclature. Vet Rec. 2008;15:867–868. doi: 10.1136/vr.162.26.867. [DOI] [PubMed] [Google Scholar]
- Fauquet CM, Stanley J. Revising the way we conceive and name viruses below the species level: a review of geminivirus taxonomy calls for new standardized isolate descriptors. Arch Virol. 2005;15:2151–2179. doi: 10.1007/s00705-005-0583-0. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Transmission Electron Microscopy of the phage UFV-P2. Virions have isometric capsids of 40-50 nm and very short tails (arrows). Scale bars = 100 nm. Figure S2. Comparison of the genomes of phages classified in LUZ24likevirus genus. The collinearity among genomes is represented by the conserved locally collinear blocks (LCBs). In the main block (blue), the regions of similarity plot with high identity corresponds to the set of shared LCBs (see Figure 2). The connection line between blocks correspond to the central point of LCB of the reference genome (phage LUZ24 genome).