

Abstract
Pseudomonas aeruginosa is a major opportunistic pathogen in cystic fibrosis (CF) patients and causes a wide range of infections among other susceptible populations. Its inherent resistance to many antimicrobials also makes it difficult to treat infections with this pathogen. Recent evidence has highlighted the diversity of this species, yet despite this, the majority of studies on virulence and pathogenesis focus on a small number of strains. There is a pressing need for a P. aeruginosa reference panel to harmonize and coordinate the collective efforts of the P. aeruginosa research community. We have collated a panel of 43 P. aeruginosa strains that reflects the organism's diversity. In addition to the commonly studied clones, this panel includes transmissible strains, sequential CF isolates, strains with specific virulence characteristics, and strains that represent serotype, genotype or geographic diversity. This focussed panel of P. aeruginosa isolates will help accelerate and consolidate the discovery of virulence determinants, improve our understanding of the pathogenesis of infections caused by this pathogen, and provide the community with a valuable resource for the testing of novel therapeutic agents.
Keywords: Cystic fibrosis, genotype, pathogen, Pseudomonas aeruginosa
Background
Cystic fibrosis (CF) is a significant health care challenge and an important cause of premature mortality. Chronic lower respiratory tract infections are the major cause of morbidity and mortality in CF. Impaired mucociliary clearance from the lung makes CF patients vulnerable to opportunistic infections. Novel data from nonculture-based techniques suggests that the airway microbiome in CF is polymicrobial with multiple organisms present (Blainey et al. 2012; Fodor et al. 2012; Zhao et al. 2012). Despite these methodologic advances, conventional culture techniques remain the main clinical tool used in managing CF infections. Such conventional culture methods reveal that a relatively limited number of pathogens are isolated during pulmonary infections seen in patients with CF. The pathogens isolated by culture are predominantly Staphylococcus aureus, Haemophilus influenzae, and Pseudomonas aeruginosa. Rarer organisms from the Burkholderia cepacia complex (Bcc) and other pathogens are also encountered (de Soyza et al. 2004; Davies and Rubin 2007). Although Bcc causes less than 10% of CF infections (Lipuma 2010), the established international Bcc reference panel has helped harmonize Burkholderia research by standardizing approaches (Mahenthiralingam et al. 2000; Coenye et al. 2003).
Pseudomonas aeruginosa is the major pathogen in CF, infecting up to 80% of adult patients, and once established, the pathogen is often difficult to treat clinically (Cheng et al. 1996; Fothergill et al. 2012a; Parkins et al. 2012). Surprisingly there are no recognized international reference panels for the more prevalent CF pathogens, such as P. aeruginosa, S. aureus or H. influenzae. Arguably the most pressing need is for an international P. aeruginosa reference panel to reflect the relevance of this pathogen to CF and a range of other infections. Such a panel will encourage researchers to avoid use of isolates with limited availability, and to potentially prevent unnecessary repetition across laboratories. The availability of a standardized reference panel would improve efficiency and reduce experimental animal sacrifice, while also facilitating the search for improved therapeutic approaches. In order to assemble the most appropriate reference panel of P. aeruginosa isolates, we aimed to define consensus on the core characteristics of an international reference panel through an iterative and interactive process involving workshops and a consensus finding exercise. Molecular genotyping was then used to ensure that the panel was broadly representative of the wider population structure of P. aeruginosa.
Methods
Consensus choice of isolates for initial inclusion
A broad range of expertise was assembled including clinicians, clinical microbiologists and basic science microbiology researchers. Requirements for a reference panel were discussed in open forum on two occasions under the auspices of a European Union Co-ordinated Scientific and Technology (COST) action (COST BM1003; http://www.cost-bm1003.info/). Discussion with further researchers active in the field with an international perspective (including coauthors D. D. V., J. P. P., T. K., J. B., and B. T.; see also acknowledgments) was then conducted prior to a final consensus process involving COST action members using prior techniques (RAND consensus tool) (Francis et al. 2007).
The consensus-seeking process used statements identified in the prior workshops, with the assembled experts independently scoring each statement. Statements identified and scored in the RAND process are included in Table 1. Consensus was then sought based on the individual scores as compared to the group average. Indifference was rated as scores 4–6, while 7–9 was rated as a positive consensus and 1–3 as a negative consensus. The group were also asked to rate the ideal number of isolates to be included in the panel with a mean score presented.
Table 1.
Details of the criteria used in the consensus process and the outcome. Please grade the following criteria as regards how necessary they are to necessitate inclusion of a particular Pseudomonas aeruginosa strain into an international reference panel (increasing numbers mean increasing necessity): Consensus grading – NR, consensus not reached; 9, mandatory criteria; 7, 8, necessary criteria; 4, 5, 6, useful but not mandatory; 2, 3, not a necessary criteria; 1, no relevance to international reference panel.
| Consensus grade | |
|---|---|
| The Pseudomonas strain has | |
| Already been genome sequenced | 5 |
| Some prior virulence data in vitro | 6 |
| Extensive prior virulence data in vitro | NR |
| Some prior virulence data in vivo | NR |
| Extensive prior virulence data in vivo | NR |
| A known biofilm formation phenotype | NR |
| Can be easily phenotyped for biofilm phenotype | NR |
| Comes from a clinical source other than CF | 6 |
| Comes only from CF sources | NR |
| Is known to be from early or late stage CF | NR |
| Is of mucoid nature | NR |
| Is of non mucoid nature | NR |
| Is susceptible to bacteriophages | NR |
| The strain panel should include | |
| Range of strains from different sources (geographic) | 8 |
| Range of strains from different sources (CF, non CF clinical and environmental) | 8 |
| ONLY genome sequenced strains | 4 |
| Strains with structural characterization data available | 6 |
| ONLY strains where structure of virulence determinants known | NR |
| Strains where variation in resistance is known or expected | 6 |
| ONLY strains where resistance patterns already known | NR |
| Strains where in vivo virulence patterns where possible | 6 |
| ONLY strains where in vivo virulence patterns already known | NR |
| Strains to allow a variety of Mucoid\ non Mucoid where possible | 6 |
| Strains ONLY where non Mucoid \non Mucoid Status known | NR |
| Strains of epidemic and sporadic clinical infection status | 8 |
| The panel should be provided at no cost | NR |
| The panel should be hosted by a recognized collection (e.g., BCCM) | 8 |
| The ideal number of strains in the panel should be 0–10,11–20, 21–30, 31–40, 41–50 | 21–30 |
Genotyping of isolates using the ArrayTube method
Strains of P. aeruginosa were genotyped using the ArrayTube (AT) system (CLONDIAG, Alere Technologies, Köln, Germany) as described previously (Wiehlmann et al. 2007). The AT microarray chip enables strains to be classified according to 13 core genome single-nucleotide polymorphisms (SNPs), and also screens for 38 variable genetic markers of the P. aeruginosa accessory genome. These include several previously reported genomic islands (Arora et al. 2001; Liang et al. 2001; Larbig et al. 2002; de Chial et al. 2003; Spencer et al. 2003; He et al. 2004; Klockgether et al. 2004; Lee et al. 2006). Data from the 13 SNPs are combined with flagellin type (a/b) and the presence of the genes encoding mutually exclusive type III secretion exotoxins (S or U), to generate a strain-specific “hexadecimal code” represented by four digits (Wiehlmann et al. 2007). This code can be used to search a large database of P. aeruginosa strains (Cramer et al. 2012). Subsequently, eBURST (version 3.0) (Feil et al. 2004; Spratt et al. 2004) analysis of data generated using the AT from the 13 SNPs, flagellin type (a or b) and presence of the mutually exclusive type III secretion exotoxins (S or U), was used to visualize the position of panel strains within the wider P. aeruginosa population structure using a database of 955 genotyped strains (Cramer et al. 2012).
Results
RAND process
A number of strain and reference panel characteristics were identified at the scoping workshops (Table 1). Through the iterative consensus-seeking process we identified a variety of characteristics for individual isolates and also for the overall panel, and then reduced these to criteria that were deemed either mandatory or critical. Consensus was reached where the majority of scores were within the same ranking and was achieved in 12 of 28 statements. The preferred mean number of isolates to be included in the reference panel was 26 ± 4 (10 responders).
Panel strain selection
Isolates were selected based on the characteristics identified through the Delphi process and literature searches via PubMed. Where possible, isolates were chosen when extensive prior data were available, including those with genome sequencing data (Stover et al. 2000; Mathee et al. 2008; Winstanley et al. 2009; Stewart et al. 2011), in vitro and/or in vivo virulence data (Hajjar et al. 2002; Al-Aloul et al. 2004; Cigana et al. 2009; Carter et al. 2010). Isolates were also selected to include representatives from diverse biologic and geographic origins, including clinical isolates (CF and non-CF clinical infections) and environmental sources (Cigana et al. 2009; Pirnay et al. 2009). Because this is an International panel, a global perspective is essential and isolates from geographically dispersed origins were also chosen (Pirnay et al. 2009). The final selection of 43 strains is listed in Table 2. The strain panel is available from the BCCM/LMG Bacteria Collection (http://bccm.belspo.be/about/lmg.php), Gent, Belgium, and the LMG reference numbers are shown in Table 2.
Table 2.
International reference panel of Pseudomonas aeruginosa isolates.
| Panel no. | Source ID | LMG number | Origin and source | Genome sequence | AT code (Clone) | Details | Reference |
|---|---|---|---|---|---|---|---|
| CF transmissible strains | |||||||
| 1 | LES B58 | 27622 | CF, U.K. (1988) | Yes | 4C12(T) | Earliest isolate of LES; genome sequenced; produces more biofilm than PAO1 and PA14 but less motile; virulent in C. elegans model, reduced virulence in murine model | Kukavica-Ibrulj et al. (2008); Winstanley et al. (2009); Carter et al. (2010) |
| 2 | LES 400 | 27623 | CF, U.K. | Yes | 4C12(T) | Quorum sensing defective lasR mutant of LES; reduced C. elegans killing; upregulation of alginate production | Salunkhe et al. (2005); Carter et al. (2010) |
| 3 | LES 431 | 27624 | Non CF parent of CF patient, U.K. | Yes | 4C12(T) | LES isolate with increased virulence in the murine model; hypervirulent subtype more adapted to acute infections; transmitted to non CF parents; upregulated QS genes | Salunkhe et al. (2005); Carter et al. (2010) |
| 4 | C3719 | 27625 | CF, Manchester, U.K. | Yes | 4012(O) | Manchester epidemic strains associated with increased treatment requirements of CF patients with a highly transmissible strain | Jones et al. (2002); Mathee et al. (2008) |
| 5 | DK2 | 27626 | CF, Denmark; earliest isolates 1973–2007 | Yes | F421(A2) | Initial period of rapid adaption in CF host, followed by long period of some phenotypic changes and genotypic negative selection; low growth rate, loss of motility and reduced regulatory functions | Yang et al. (2011b) |
| 6 | AES-1R | 27627 | Pediatric CF (1992), Melbourne, Australia | Yes | E82A | AUST-01 strain (aka AES-1, Pulsotype 1), ST-649; associated with increased morbidity and treatment; virulence gene expression during chronic CF infection; niche specialization | Armstrong et al. (2002); Manos et al. (2008); Naughton et al. (2011) |
| 7 | AUS23 | 27628 | Adult CF (2007), Brisbane, Australia | Pending | 2D9A | AUST-02 strain (aka AES-2, Pulsotype 2), ST-775; associated with increased treatment requirements and niche specialization | O'Carroll et al. (2004); Kidd et al. (2012) |
| 8 | AUS52 | 27629 | Adult CF (2008), Hobart, Australia | Pending | EC22 | AUST-03 strain (aka AES-3), ST-242; associated with increased exacerbation rate; predominantly detected in CF patients from Tasmania and southern Australia | Bradbury et al. (2008); Kidd et al. (2012) |
| Paired or sequential CF isolates | |||||||
| 9 | AA2 | 27630 | CF (early), Germany | Yes | AF9A | LPS and PGN studied; in vivo virulence | Bragonzi et al. (2006, 2009); Lore et al. (2012) |
| 10 | AA43 | 27631 | CF (late), Germany | Yes | AF9A | As above | Bragonzi et al. (2006, 2009); Lore et al. (2012) |
| 11 | AA44 | 27632 | CF (late), Germany | Yes | AF9A | As above | (Bragonzi et al. 2006, 2009;Lore et al. 2012) |
| 12 | AMT 0023-30 | 27633 | Pediatric CF, Seattle, WA | Yes | 478A | Early isolate taken at 6 months of age; persister cells | Mulcahy et al. (2010) |
| 13 | AMT 0023-34 | 27634 | Pediatric CF, Seattle, WA | Yes | 478A | 96-month isolate from same patient as AMT023-30 showing a 100-fold increase in persister cells; mutS mutation; hypermutator phenotype | Mulcahy et al. (2010) |
| 14 | AMT 0060-1 | 27635 | Pediatric CF, Seattle, WA | No | 6D92(H) | Late isolate identifying hip mutants; late isolates showed high-level persister cells; mutation in mexZ repressor affecting MIC to Ofloxacin, Carbenicillin and Tobramycin. | Mulcahy et al. (2010) |
| 15 | AMT 0060-2 | 27636 | Pediatric CF, Seattle, WA | No | 6D92(H) | Late isolate identifying hip mutants; late isolates from these patients showed high level persister cells, mutation in mexZ repressor affecting MIC to Ofloxacin, Carbenicillin and Tobramycin. | Mulcahy et al. (2010) |
| 16 | AMT 0060-3 | 27637 | Pediatric CF, Seattle, WA | Pending | 6D92(H) | Early isolate from same patient as AMT 0060-1 and -2 were isolated | Mulcahy et al. (2010) |
| Commonly used strains/clones | |||||||
| 17 | PAO11 (ATCC 15692) | 27638 | Genome sequenced isolate | 0002(W) | Widely studied wound isolate from Melbourne, Australia; characterized by Bruce Holloway | Stover et al. (2000) | |
| 18 | UCBPP-PA14 | 27639 | Human Burn isolate | Yes | D421(A) | Severe symptoms in mouse burn model; 58 genes present in PA14 not in PAO1 | Rahme et al. (1995); Lee et al. (2006) |
| 19 | PAK | 27640 | Clinical non CF | Pending | 55AA | Widely studied; expresses pili, flagella and glycosylation islands etc. | Totten and Lory (1990); Semblat and Doerig (2012) |
| 20 | CHA | 27641 | CF | Yes | EC2A | Detailed phenotypic characterization; | Toussaint et al. (1993); Dacheux et al. (2000); Bezuidt et al. (2013) |
| 21 | NN2 | 27642 | CF, Germany | Yes | C40A (C) | Detailed phenotypic characterization; Clone C | Cramer (et al. 2011) |
| Strains with specific phenotypic/virulence characteristics | |||||||
| 22 | IST 27 mucoid | 27643 | Lisbon Portugal, CF patient | Pending | F69A | Pediatric CF patient from Hospital de Santa Maria,Lisbon | Leitao et al. (1996); Tavares et al. (1999) |
| 23 | IST 27N | 27644 | Lisbon Portugal, spontaneous nonmucoid variant of IST27 | Pending | F69A | Spontaneous non-mucoid variant of IST27 with the same macrofragment restriction profile by RFLP-PFGE | Leitao et al. (1996); Tavares et al. (1999) |
| 24 | 968333S | 27645 | UK Non CF bronchiectasis | Pending | 3C52(K) | Non CF bronchiectasis patient from the UK; Mucoid; Detailed phenotypic characterization | De Soyza et al. (Manuscript under review)2 |
| 25 | 679 | 27646 | Non CF Urine sample, male, Wroclaw Poland, 2011 | Pending | 0C4A | Resistant to imipenem and meropenem | Zuzanna Drulis-Kawa, University of Wroclaw |
| 26 | 39016 | 27647 | Keratitis eye isolate, U.K. | Yes | F469 (D) | Clone D by AT; serotype O11; carries distinctive pilA; subpopulation adapted to corneal infections; associated with severe infection; ST-235 | Stewart et al. (2011); Hall et al. (2013) |
| 27 | 2192 | 27648 | Chronic CF patient, Boston, MA | Yes | 049A | Large genome, converted to mucoidy; produces LPS which lacks O-side chains; lacks motility | Mathee et al. (2008) |
| 28 | NH57388A | 27649 | Denmark CF | Pending | 6822 | Stable CF sputum mucoid isolate; alginate hyperproducer; has functional QS system; caused chronic lung infection in a mouse model. | Hoffmann et al. (2005) |
| Strains included to represent serotype, genotype, geographic and source diversity | |||||||
| 29 | 1709-12 | 27650 | Leuven Belgium Non CF clinical 2004 | Pending | 7D92 | Serotype 12; multi drug resistant | Pirnay et al. (2009) |
| 30 | Mi 162 | 27651 | Non CF burn, Ann Arbor, MI, 1997 | Pending | D429(Q) | Serotype 11; multi drug resistant | Pirnay et al. (2009) |
| 31 | Jpn 1563 | 27652 | Lake Tamaco, Japan, Lake water, 2003 | Pending | 3C1A | Non serotypeable | Pirnay et al. (2009) |
| 32 | LMG 14084 | 27653 | Bucharest, Romania, Water, 1960-1964 | Pending | E42A (B) | Serotype 17 | Pirnay et al. (2009) |
| 33 | Pr335 | 27654 | Prague, Czech Republic, Hospital environment 1997 | Pending | FA0A | Serotype 1 | Pirnay et al. (2009) |
| 34 | U018a | 27655 | Hobart, Australia, CF patient | Pending | 2A42 | Serotype 1 | Pirnay et al. (2009) |
| 35 | CPHL 9433 | 27656 | Tobacco plant, Philippines | Pending | F462 | Serotype 1 | Pirnay et al. (2009) |
| 36 | RP1 | 27657 | CF, Germany | Yes | OC2E | Abundant in Northern Germany but infrequent in other European countries | Cramer et al. (2012) |
| 37 | 15108/-1 | 27658 | ICU (acute infection), Spain | Yes | E429 | Isolated in Prat de Llobgregat, Spain; Detailed phenotypic characterization | Kohler et al. (2009) |
| 38 | 57P31PA | 27659 | Chronic Obstructive Pulmonary Disease, USA | Yes | 2C22 | Detailed phenotypic characterization | Rakhimova et al. (2009) |
| 39 | 13121/-1 | 27660 | ICU (acute infection), France | Yes | 239A | Isolated inLimoges, France; Detailed phenotypic characterization | Wiehlmann et al. (2007) |
| 40 | 39177 | 27661 | Keratitis, Manchester U.K. | Yes | EA0A | Serotype 1; Detailed phenotypic characterization | Stewart et al. (2011); (Shankar et al. (2012) |
| 41 | KK1 | 27662 | CF, Germany | Yes | 1BAE | Detailed phenotypic characterization | Cramer et al. (2012) |
| 42 | A5803 | 27663 | Community-acquired pneumonia | Yes | F429 | Detailed phenotypic characterization | Wiehlmann et al. (2007) |
| 43 | TBCF10839 | 27664 | CF, Germany | Yes | 3C52 | Detailed phenotypic characterization | Bezuidt et al. (2013); Klockgether et al. (2013) |
MIC, minimum inhibitory concentration; ICU, intensive care unit.
This has alternate strain names of: 1C, ATCC 15692, ATCC 17503, ATCC 25247, ATCC 25375, BCRC 13078, CCRC 13078, CECT 4122, CIP 104116, HER 1146, Holloway 1C, Holloway1, JCM 14847, KCTC 1637, Kemira Oy, LMG 12228, NCCB 4163, NCIMB 10545, NCIMB 10548, PA0 1, PA01, PAO 1, PC 4163, PRS 101, Stanier 131, VTT E-082794, VTT E-84219.
De Soyza, A., A. Perry, A. J. Hall, S. Sunny, K. Walton, N. H. Mustafa, et al. Pseudomonas aeruginosa cross infection in bronchiectasis: a Molecular Epidemiology study. Manuscript under review.
Genotyping to define the distribution of the panel strains among the wider P. aeruginosa population
Each of the panel strains was genotyped using the AT method. Figure 1 shows an eBURST representation of the distribution of the panel strains among the wider population of P. aeruginosa. The panel strains are widely distributed and include representatives of both abundant and less abundant clones. The AT codes are indicated in Table 2.
Figure 1.
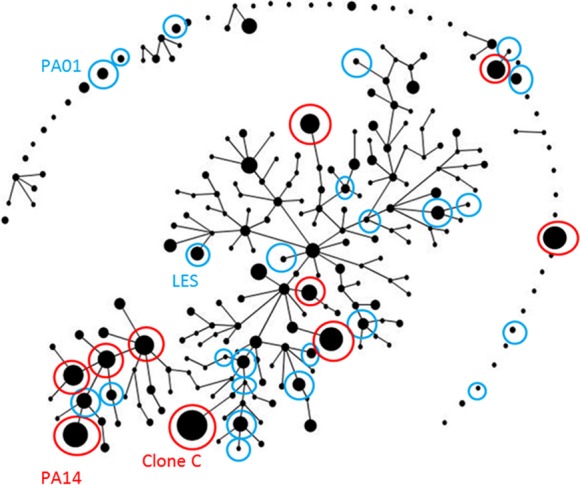
Distribution of panel strains among the wider Pseudomonas aeruginosa population. The figure shows an eBURST analysis based on AT genotyping using a database of 955 P. aeruginosa isolates of independent origin. The locations of the panel isolates are circled. Red circles indicate genotypes corresponding to the 10 most abundant in the database. Blue circles indicate the location of other panel strains. Each dot represents a different genotype; the size of a dot indicates the relative abundance of a genotype. The genotype cluster locations for four commonly studied strains from the panel are indicated.
Brief description of the panel strains
CF transmissible strains
Although it is still widely assumed that most CF patients acquire their infecting strains of P. aeruginosa from environmental sources, there is increasing evidence for the emergence of particularly successful transmissible strains (Fothergill et al. 2012a), some of which have been associated with increased patient morbidity or mortality (Al-Aloul et al. 2004; Aaron et al. 2010) or antimicrobial resistance (Ashish et al. 2012). Hence, we have included representatives of the most widely studied transmissible strains. These include three isolates of the Liverpool Epidemic Strain (LES), first reported in a U.K. children's CF Unit in the 1990s (Cheng et al. 1996), but known to be widespread through the U.K. (Scott and Pitt 2004; Martin et al. 2013), and reported recently in North America (Aaron et al. 2010). This strain is associated with novel transmission events (McCallum et al. 2002), increased patient morbidity (Al-Aloul et al. 2004), and resistance to antimicrobials (Ashish et al. 2012). There are many phenotypic variants of this strain (Mowat et al. 2011), but we have selected three, namely (1) LESB58, the earliest known isolate (from 1988) (Winstanley et al. 2009), (2) LES400, a lasR mutant that is in defective quorum sensing and shows reduced virulence in various infection models (Salunkhe et al. 2005; Carter et al. 2010) and (3) LES431, an isolate associated with the infection of the non-CF parent of a CF patient, an upregulated quorum sensing system and enhanced virulence in infection models (McCallum et al. 2002; Salunkhe et al. 2005; Carter et al. 2010). All three LES isolates are methionine auxotrophs.
In addition, we include a genome-sequenced representative of the other most commonly studied U.K. epidemic strain, the Manchester strain, C3719 (Mathee et al. 2008; Jones et al. 2010). A number of transmissible strains have been reported in Australia, and we include representatives of the three most widely studied, namely AUST-01 (AES-1), AUST-02 (AES-2) and AUST-03 (AES-3). AUST-01, associated with increased patient morbidity, was first reported in Melbourne but has also been reported in Sydney and Brisbane (Armstrong et al. 2003; O'Carroll et al. 2004; Kidd et al. 2013). AUST-02 has been reported as more common in Brisbane (Syrmis et al. 2004; Kidd et al. 2013), whereas AUST-03 is the most common CF strain in Tasmania (Bradbury et al. 2008). Strain DK2, identified as infecting multiple CF patients in Denmark, was the subject of a detailed analysis of genome sequences from multiple isolates (Yang et al. 2011a). Although we recognize that there are other known CF transmissible strains (Fothergill et al. 2012a), and new strains are emerging all the time, we have restricted our choices to those for which there are substantial additional phenotypic and genotypic data.
Other CF isolates
In choosing our CF isolates (epidemic and nonepidemic strains) we sought to represent phenotypes typically associated with such isolates, such as mucoidy, hypervirulence, loss of virulence activities (such as quorum sensing), antimicrobial resistance, and auxotrophy.
Adaptation is a hallmark of CF pathogens, enabling them to chronically colonize the challenging host environment and avoid immune detection during chronic colonization (Callaghan and McClean 2012). There have been a number of studies where single-strain sequential isolates for CF patients have been analyzed in order to understand the mechanisms of the adaptation that P. aeruginosa populations undergo during chronic lung infections (Smith et al. 2006; Bragonzi et al. 2009; Cramer et al. 2011; Yang et al. 2011b). Classically, these include examples of isolates from “early” and “late” in an infection, and we have included example isolates from both European and North American studies. We have included isolates AA2 (early), AA43 (mucoid, late) and AA44 (nonmucoid, late), which have been compared using multiple phenotypic tests and in a murine infection model (Bragonzi et al. 2009). We also include five pediatric sequential isolates including the matched “early” and “late” isolates AMT 0023-30 and AMT 0023-34, the latter of which is an example of a hypermutator (mutS mutant) and shows a 100-fold increase in persister levels (enhanced survival upon exposure to antibiotics) (Mulcahy et al. 2010). We further include AMT0060-1, -2 and -3, “early” and “late” isolates from a separate patient, representing sequential isolates obtained when that patient was 15.4 (two distinct phenotypes, AMT0060-1 and -2) and 7.7 (AMT0060-3) years-old.
Widely studied strains
We have included in the panel a number of strains that are frequently studied by the P. aeruginosa research community. We note that there are variants (Klockgether et al. 2010) of the most widely studied strain, PAO1, which was the first to be genome sequenced (Stover et al. 2000). This can lead to difficulties when comparisons are made between laboratories. The variant of PAO1 we have deposited is derived from the culture grown for genome sequence analysis (Stover et al. 2000). The Pseudomonas Genome Database (http://v2.pseudomonas.com) (Winsor et al. 2011) represents a very useful and comprehensive online database for interrogation of the genome of strain PAO1, as well as the genomes of other strains included in this panel (2192, 39016, C3719, DK2, LES B58, and UCBPP-PA14) (Table 2).
It has been shown that PA14-like strains and Clone C are the two most abundant P. aeruginosa clones among CF patients (Romling et al. 2005; Cramer et al. 2011, 2012). Indeed, Clone C is ubiquitous throughout the inanimate environment also, whereas UCBPP-PA14 (PA14) is not so common outside of the CF population (Pirnay et al. 2009). Hence, representatives of both of these clones have been included. Strain PAK has also been widely studied, for example in relation to biofilm formation (Vasseur et al. 2005), flagellar glycosylation (Miller et al. 2008), and gene regulation (Brencic and Lory 2009).
We have also included the highly pathogenic strains CHA (Toussaint et al. 1993) and TBCF10839 (Tummler et al. 1991; Klockgether et al. 2013). Strain CHA is an example of a strain producing the potent exotoxin U, and its type III secretion system has been the subject of much analysis (Dacheux et al. 2000; Ader et al. 2005). The virulence of strain TBCF10839 has also been extensively studied (Bohn et al. 2009).
Strains with specific phenotypic characteristics
Mucoidy is a phenotype associated with chronic colonization, which derives from the production of high concentrations of the exopolysaccharide, alginate. Mucoid strains that chronically colonize the lungs of CF patients may evolve from initial nonmucoid strains, but the mucoid phenotype is unstable in vitro. Consequently, the pair of strains IST27, a mucoid isolate from a CF patient, and its nonmucoid variant IST27N, obtained spontaneously during IST27 cultivation in the laboratory, have been included to directly assess the role of mucoidy in pathogenesis and the regulation of the mucoid phenotype switch. IST27 and IST27N are clonal variants indistinguishable by genomic fingerprinting (Leitao et al. 1996). In contrast to IST27, IST27N has undetectable levels of GDP (guanosine diphosphate)-mannose dehydrogenase (GMD) activity, consistent with the concept that the control of alginate biosynthesis occurs at the level of the encoding gene algD (Tavares et al. 1999). In addition, other mucoid strains such as NH57388A and 968333S have been included (manuscript under review). The latter is a mucoid strain and was selected due to its isolation from a U.K. patient with advanced non-CF bronchiectasis treated with long-term colistin. This will provide a useful comparator to “late phase” CF strains.
Lipopolysaccharide (LPS) composition is another important phenotype that plays a key role in pathogenesis; therefore isolates with defined LPS structures have been included (Cigana et al. 2009). Furthermore, strains 39,106 (Stewart et al. 2011) and 679 were chosen as examples of severe keratitis and urinary tract infection isolates, respectively.
Strains representing genotypic, geographic and source diversity
Although conscious of keeping the reference panel to a reasonable number, we included a number of strains to ensure that we captured the diversity of P. aeruginosa in nature as much as possible. There have been a number of key studies using genotypic and phenotypic approaches to defining the population structure (Pirnay et al. 2002, 2009; Wiehlmann et al. 2007; Cramer et al. 2012; Kidd et al. 2012; Shankar et al. 2012; Martin et al. 2013). We selected seven isolates from a P. aeruginosa study (Pirnay et al. 2009) representing diversity in serotype, drug resistance, and geographic source, including three serotype 1 strains from very different geographic locations, and environmental isolates. Cramer et al. (2012) recently defined the population structure of P. aeruginosa in relation to CF by AT genotyping a collection of 955 isolates from multiple European CF centers. The collection also included isolates from various non-CF clinical and environmental sources for comparison. We have included in our collection representatives of each of the 10 most common clones identified. We have also ensured that we have representatives from diverse types of infection, including various non-CF respiratory infections (pneumonia, including ventilator-associated pneumonia (VAP), chronic obstructive pulmonary disease (COPD)-associated, non-CF bronchiectasis), burn wound infections, eye infections, and urinary tract infections. Although we accept that other genotyping approaches have been used to study large collections of P. aeruginosa, including multilocus sequence typing (MLST) and variable number of typing repeats (VNTR), and that all such methods have limitations, using the AT method we were able to easily place the panel strains in the context of a much wider survey and ensure that common clones are represented. It is likely that all such genotyping methods will be superceded by whole genome sequencing. Hence, we are committed to ensuring that each panel strain is genome sequenced. This is currently being undertaken in collaboration with Roger Levesque (Université Laval) and the data will be made available as soon as possible.
Discussion
There is a need to coordinate our collective efforts in P. aeruginosa research to achieve both rapid and meaningful progress. Arguably the most pressing need for accelerating scientific progress relates to CF due to the clinical burden and prognostic effects of P. aeruginosa infections. Hence, our efforts were focussed on defining a core reference panel of isolates relevant to CF research. However, given the increasing importance of P. aeruginosa in a range of opportunistic infections, we have also chosen a panel with broader relevance.
We realize that no panel will achieve absolute consensus on all of the required parameters that various international research teams may wish to study. However, the consensus-seeking process did define core characteristics necessary for the proposed reference panel of P. aeruginosa. These characteristics included ensuring diversity in the biologic niche by choosing clinical, environmental, and laboratory isolates. We have also ensured that geographic diversity is reflected, and that although the panel includes representatives of the dominant circulating P. aeruginosa clones, we also include more unusual outliers to reflect the diversity of the species. Hence, we believe that within the limitations of keeping the number manageable, we have assembled a representative panel for use by the wider research community.
Infections of the CF lung by P. aeruginosa have been associated with the development of a number of important bacterial characteristics such as induction of mucoid status (Govan and Deretic 1996), evidence of hypermutability (Oliver et al. 2000), changes in cell surface virulence determinants (Cigana et al. 2009), and loss of virulence factors (D'Argenio et al. 2007). However, it is important to note that during chronic infections of the CF lung P. aeruginosa populations are diverse. Hence, there is considerable variability in virulence factor expression or antibiotic resistance among CF isolates of P. aeruginosa, even when they are obtained from the same sputum sample and the patient is infected with a single strain (Foweraker et al. 2005; Mowat et al. 2011). These features of P. aeruginosa associated with CF may be due to the (passive) accumulation or (active) development of mutations that occurs during the chronic lung infections that characterize CF. Furthermore, there are data demonstrating “CF specific” differences in the expression of virulence determinants such as LPS as compared to isolates from other clinical diseases states and environmental isolates (Ernst et al. 2007; Moskowitz and Ernst 2010). We have sought to represent in the panel some of the key phenotypic variations evident in the wider P. aeruginosa community, especially in relation to CF infections.
We achieved consensus on the requirement that the panel must include CF epidemic strains (Fothergill et al. 2012a) and sporadic isolates responsible for clinical infections. We also achieved consensus on the need for multiple isolates of certain important strains (e.g., LES), including subtypes with different virulence characteristics.
Antimicrobial resistance and biofilm information are both characteristics central to the clinical challenges in managing CF and other infections. The panel therefore includes multidrug-resistant strains and an isogenic parent and mutant strain which are, respectively, biofilm-forming and biofilm-deficient. It is the intention that such a panel should be used for further in-depth analysis and comparisons of phenotypes such as biofilm formation and antimicrobial resistance, and there was agreement that this requires further study. It is also envisaged that the panel will represent an excellent strain reference set for the testing of novel therapeutic approaches to the treatment of P. aeruginosa infections, which are desperately needed (Fothergill et al. 2012b).
Making use of prior data is scientifically and economically mandatory and improves the effective use of research funding. The proposed panel has aimed to include isolates that had already been well-characterized, such as the commonly used strains PAO1, PA14, PAK, and LESB58, and isolates where there has been comprehensive pathogenicity work undertaken.
Importantly, there was consensus that all of the identified factors did not need to be defined for each strain in the panel at inclusion. Full genome sequencing was not felt mandatory for inclusion in the panel. However, the majority of strains included have been sequenced and the remainder will be sequenced in the near future for completion. Furthermore, the number of strains included is beyond the consensus achieved for an “ideal” number. It proved difficult to reduce the number of strains included without losing the richness of the existing data already available on many of these strains.
Significant challenges lie ahead in understanding the biology of P. aeruginosa. The coordination of scientific efforts across research groups and avoiding the use of widely differing random isolates which result in unhelpful repetition is imperative. Better coordination and definitive replicate data fulfil an unwritten demand of the scientific community, the patient population and life science research funders. The aim of this proposed panel is to harmonize and coordinate the ongoing efforts of the research community to fulfil these goals. The international reference panel of B. cepacia complex isolates undoubtedly led to more streamlined approaches with this less prevalent group of pathogens. The Bcc reference panel grew with time to reflect the needs of the bioscience community, as well as to mediate the discovery of newer species within the complex being defined. This proposed panel of P. aeruginosa isolates has a particular focus on CF and human disease. As occurred with the original Bcc reference panel, which comprised 30 strains originally (Mahenthiralingam et al. 2000), but was updated with time and new discoveries (Coenye et al. 2003), this panel may similarly need to be extended in time. However, it is clear that a focussed panel of P. aeruginosa isolates as assembled is needed to help accelerate discovery and assessment of virulence determinants, and to develop better strategies to counter this successful pathogen.
Acknowledgments
The following authors (A. D. S., E. M., P. D., W. K., Z. D. K., S. R. S., V. T., T. C., I. S. C., S.Mc.C., and C.W.) are all members of the EU COST Action BM1003: Microbial cell surface determinants of virulence as targets for new therapeutics in cystic fibrosis (http://www.cost-bm1003.info/) and acknowledge this support in the collation of this panel. A. D. S. was supported by a Higher Education Funding Council for England Senior Lectureship, J. L. B. was supported by a grant from the National Institutes of Health (NIDDK P30 DK 89507). We also acknowledge the helpful advice and encouragement from Peter Vandamme, Ghent University, David Speert, University of British Columbia, Scott Bell, The Prince Charles Hospital, Brisbane, and John R. W. Govan. University of Edinburgh. We also gratefully acknowledge Marvin Whiteley, University of Texas at Austin, and Niels Hoiby, University of Copenhagen, who kindly supplied strains.
Conflict of Interest
None declared.
References
- Aaron SD, Vandemheen KL, Ramotar K, Giesbrecht-Lewis T, Tullis E, Freitag A, et al. Infection with transmissible strains of Pseudomonas aeruginosa and clinical outcomes in adults with cystic fibrosis. JAMA. 2010;304:2145–2153. doi: 10.1001/jama.2010.1665. [DOI] [PubMed] [Google Scholar]
- Ader F, Faure R, le Berre K, Gosset P, Epaulard O, Toussaint B, et al. Alveolar response to Pseudomonas aeruginosa: role of the type III secretion system. Infect. Immun. 2005;73:4263–4271. doi: 10.1128/IAI.73.7.4263-4271.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al-Aloul M, Crawley J, Winstanley C, Hart CA, Ledson MJ, Walshaw MJ. Increased morbidity associated with chronic infection by an epidemic Pseudomonas aeruginosa strain in CF patients. Thorax. 2004;59:334–336. doi: 10.1136/thx.2003.014258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armstrong DS, Nixon GM, Carzino R, Bigham A, Carlin JB, Robins-Browne RM, et al. Detection of a widespread clone of Pseudomonas aeruginosa in a pediatric cystic fibrosis clinic. Am. J. Respir. Crit. Care Med. 2002;166:983–987. doi: 10.1164/rccm.200204-269OC. [DOI] [PubMed] [Google Scholar]
- Armstrong D, Bell S, Robinson M, Bye P, Rose B, Harbour C, et al. Evidence for spread of a clonal strain of Pseudomonas aeruginosa among cystic fibrosis clinics. J. Clin. Microbiol. 2003;41:2266–2267. doi: 10.1128/JCM.41.5.2266-2267.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arora SK, Bangera M, Lory S, Ramphal R. A genomic island in Pseudomonas aeruginosa carries the determinants of flagellin glycosylation. Proc. Natl Acad. Sci. USA. 2001;98:9342–9347. doi: 10.1073/pnas.161249198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashish A, Shaw M, Winstanley C, Ledson MJ, Walshaw MJ. Increasing resistance of the Liverpool Epidemic Strain (LES) of Pseudomonas aeruginosa (Psa) to antibiotics in cystic fibrosis (CF)–a cause for concern? J. Cyst. Fibros. 2012;11:173–179. doi: 10.1016/j.jcf.2011.11.004. [DOI] [PubMed] [Google Scholar]
- Bezuidt OK, Klockgether J, Elsen S, Attree I, Davenport CF, Tummler B. Intraclonal genome diversity of Pseudomonas aeruginosa clones CHA and TB. BMC Genomics. 2013;14:416. doi: 10.1186/1471-2164-14-416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blainey PC, Milla CE, Cornfield DN, Quake SR. Quantitative analysis of the human airway microbial ecology reveals a pervasive signature for cystic fibrosis. Sci. Transl. Med. 2012;4:153ra130. doi: 10.1126/scitranslmed.3004458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bohn Y-ST, Brandes G, Rakhimova E, Horatzek S, Salunkhe P, Munder A, et al. Multiple roles of Pseudomonas aeruginosa TBCF10839 PilY1 in motility, transport and infection. Mol. Microbiol. 2009;71:730–747. doi: 10.1111/j.1365-2958.2008.06559.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradbury R, Champion A, Reid DW. Poor clinical outcomes associated with a multi-drug resistant clonal strain of Pseudomonas aeruginosa in the Tasmanian cystic fibrosis population. Respirology. 2008;13:886–892. doi: 10.1111/j.1440-1843.2008.01383.x. [DOI] [PubMed] [Google Scholar]
- Bragonzi A, Wiehlmann L, Klockgether J, Cramer N, Worlitzsch D, Doring G, et al. Sequence diversity of the mucABD locus in Pseudomonas aeruginosa isolates from patients with cystic fibrosis. Microbiology. 2006;152:3261–3269. doi: 10.1099/mic.0.29175-0. [DOI] [PubMed] [Google Scholar]
- Bragonzi A, Paroni M, Nonis A, Cramer N, Montanari S, Rejman J, et al. Pseudomonas aeruginosa microevolution during cystic fibrosis lung infection establishes clones with adapted virulence. Am. J. Respir. Crit. Care Med. 2009;180:138–145. doi: 10.1164/rccm.200812-1943OC. [DOI] [PubMed] [Google Scholar]
- Brencic A, Lory S. Determination of the regulon and identification of novel mRNA targets of Pseudomonas aeruginosa RsmA. Mol. Microbiol. 2009;72:612–632. doi: 10.1111/j.1365-2958.2009.06670.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Callaghan M, McClean S. Bacterial host interactions in cystic fibrosis. Curr. Opin. Microbiol. 2012;15:71–77. doi: 10.1016/j.mib.2011.11.001. [DOI] [PubMed] [Google Scholar]
- Carter ME, Fothergill JL, Walshaw MJ, Rajakumar K, Kadioglu A, Winstanley C. A subtype of a Pseudomonas aeruginosa cystic fibrosis epidemic strain exhibits enhanced virulence in a murine model of acute respiratory infection. J. Infect. Dis. 2010;202:935–942. doi: 10.1086/655781. [DOI] [PubMed] [Google Scholar]
- Cheng K, Smyth RL, Govan JR, Doherty C, Winstanley C, Denning N, et al. Spread of beta-lactam-resistant Pseudomonas aeruginosa in a cystic fibrosis clinic. Lancet. 1996;348:639–642. doi: 10.1016/S0140-6736(96)05169-0. [DOI] [PubMed] [Google Scholar]
- de Chial M, Ghysels B, Beatson SA, Geoffroy V, Meyer JM, Pattery T, et al. Identification of type II and type III pyoverdine receptors from Pseudomonas aeruginosa. Microbiology. 2003;149:821–831. doi: 10.1099/mic.0.26136-0. [DOI] [PubMed] [Google Scholar]
- Cigana C, Curcuru L, Leone MR, Ierano T, Lore NI, Bianconi I, et al. Pseudomonas aeruginosa exploits lipid A and muropeptides modification as a strategy to lower innate immunity during cystic fibrosis lung infection. PLoS ONE. 2009;4:e8439. doi: 10.1371/journal.pone.0008439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coenye T, Vandamme P, Lipuma JJ, Govan JR, Mahenthiralingam E. Updated version of the Burkholderia cepacia complex experimental strain panel. J. Clin. Microbiol. 2003;41:2797–2798. doi: 10.1128/JCM.41.6.2797-2798.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cramer N, Klockgether J, Wrasman K, Schmidt M, Davenport CF, Tummler B. Microevolution of the major common Pseudomonas aeruginosa clones C and PA14 in cystic fibrosis lungs. Environ. Microbiol. 2011;13:1690–1704. doi: 10.1111/j.1462-2920.2011.02483.x. [DOI] [PubMed] [Google Scholar]
- Cramer N, Wiehlmann L, Ciofu O, Tamm S, Hoiby N, Tummler B. Molecular epidemiology of chronic Pseudomonas aeruginosa airway infections in cystic fibrosis. PLoS ONE. 2012;7:e50731. doi: 10.1371/journal.pone.0050731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dacheux D, Toussaint B, Richard M, Brochier G, Croize J, Attree I. Pseudomonas aeruginosa cystic fibrosis isolates induce rapid, type III secretion-dependent, but ExoU-independent, oncosis of macrophages and polymorphonuclear neutrophils. Infect. Immun. 2000;68:2916–2924. doi: 10.1128/iai.68.5.2916-2924.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Argenio DA, Wu M, Hoffman LR, Kulasekara HD, Deziel E, Smith EE, et al. Growth phenotypes of Pseudomonas aeruginosa lasR mutants adapted to the airways of cystic fibrosis patients. Mol. Microbiol. 2007;64:512–533. doi: 10.1111/j.1365-2958.2007.05678.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies JC, Rubin BK. Emerging and unusual gram-negative infections in cystic fibrosis. Semin. Respir. Crit. Care Med. 2007;28:312–321. doi: 10.1055/s-2007-981652. [DOI] [PubMed] [Google Scholar]
- Ernst RK, Moskowitz SM, Emerson JC, Kraig GM, Adams KN, Harvey MD, et al. Unique lipid a modifications in Pseudomonas aeruginosa isolated from the airways of patients with cystic fibrosis. J. Infect. Dis. 2007;196:1088–1092. doi: 10.1086/521367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feil EJ, Li BC, Aanensen DM, Hanage WP, Spratt BG. eBURST: inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data. J. Bacteriol. 2004;186:1518–1530. doi: 10.1128/JB.186.5.1518-1530.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fodor AA, Klem ER, Gilpin DF, Elborn JS, Boucher RC, Tunney MM, et al. The adult cystic fibrosis airway microbiota is stable over time and infection type, and highly resilient to antibiotic treatment of exacerbations. PLoS ONE. 2012;7:e45001. doi: 10.1371/journal.pone.0045001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fothergill JL, Walshaw MJ, Winstanley C. Transmissible strains of Pseudomonas aeruginosa in Cystic Fibrosis lung infections. Eur. Respir. J. 2012a;40:227–238. doi: 10.1183/09031936.00204411. [DOI] [PubMed] [Google Scholar]
- Fothergill JL, Winstanley C, James CE. Novel therapeutic strategies to counter Pseudomonas aeruginosa infections. Expert Rev. Anti Infect Ther. 2012b;10:219–235. doi: 10.1586/eri.11.168. [DOI] [PubMed] [Google Scholar]
- Foweraker JE, Laughton CR, Brown DF, Bilton D. Phenotypic variability of Pseudomonas aeruginosa in sputa from patients with acute infective exacerbation of cystic fibrosis and its impact on the validity of antimicrobial susceptibility testing. J. Antimicrob. Chemother. 2005;55:921–927. doi: 10.1093/jac/dki146. [DOI] [PubMed] [Google Scholar]
- Francis HC, Prys-Picard CO, Fishwick D, Stenton C, Burge PS, Bradshaw LM, et al. Defining and investigating occupational asthma: a consensus approach. Occup. Environ. Med. 2007;64:361–365. doi: 10.1136/oem.2006.028902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Govan JR, Deretic V. Microbial pathogenesis in cystic fibrosis: mucoid Pseudomonas aeruginosa and Burkholderia cepacia. Microbiol. Rev. 1996;60:539–574. doi: 10.1128/mr.60.3.539-574.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hajjar AM, Ernst RK, Tsai JH, Wilson CB, Miller SI. Human Toll-like receptor 4 recognizes host-specific LPS modifications. Nat. Immunol. 2002;3:354–359. doi: 10.1038/ni777. [DOI] [PubMed] [Google Scholar]
- Hall AJ, Fothergill JL, Kaye SB, Neal TJ, McNamara PS, Southern KW, et al. Intraclonal genetic diversity amongst cystic fibrosis and keratitis isolates of Pseudomonas aeruginosa. J. Med. Microbiol. 2013;62:208–216. doi: 10.1099/jmm.0.048272-0. [DOI] [PubMed] [Google Scholar]
- He J, Baldini RL, Deziel E, Saucier M, Zhang Q, Liberati NT, et al. The broad host range pathogen Pseudomonas aeruginosa strain PA14 carries two pathogenicity islands harboring plant and animal virulence genes. Proc. Natl Acad. Sci. USA. 2004;101:2530–2535. doi: 10.1073/pnas.0304622101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffmann N, Rasmussen TB, Jensen PO, Stub C, Hentzer M, Molin S, et al. Novel mouse model of chronic Pseudomonas aeruginosa lung infection mimicking cystic fibrosis. Infect. Immun. 2005;73:2504–2514. doi: 10.1128/IAI.73.4.2504-2514.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones AM, Dodd ME, Doherty CJ, Govan JR, Webb AK. Increased treatment requirements of patients with cystic fibrosis who harbour a highly transmissible strain of Pseudomonas aeruginosa. Thorax. 2002;57:924–925. doi: 10.1136/thorax.57.11.924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones AM, Dodd ME, Morris J, Doherty C, Govan JR, Webb AK. Clinical outcome for cystic fibrosis patients infected with transmissible pseudomonas aeruginosa: an 8-year prospective study. Chest. 2010;137:1405–1409. doi: 10.1378/chest.09-2406. [DOI] [PubMed] [Google Scholar]
- Kidd TJ, Ritchie SR, Ramsay KA, Grimwood K, Bell SC, Rainey PB. Pseudomonas aeruginosa Exhibits Frequent Recombination, but Only a Limited Association between Genotype and Ecological Setting. PLoS ONE. 2012;7:e44199. doi: 10.1371/journal.pone.0044199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kidd TJ, Ramsay KA, Hu H, Marks GB, Wainwright CE, Bye PT, et al. Shared Pseudomonas aeruginosa genotypes are common in Australian cystic fibrosis centres. Eur. Respir. J. 2013;41:1091–1100. doi: 10.1183/09031936.00060512. [DOI] [PubMed] [Google Scholar]
- Klockgether J, Reva O, Larbig K, Tummler B. Sequence analysis of the mobile genome island pKLC102 of Pseudomonas aeruginosa C. J. Bacteriol. 2004;186:518–534. doi: 10.1128/JB.186.2.518-534.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klockgether J, Munder A, Neugebauer J, Davenport CF, Stanke F, Larbig KD, et al. Genome diversity of Pseudomonas aeruginosa PAO1 laboratory strains. J. Bacteriol. 2010;192:1113–1121. doi: 10.1128/JB.01515-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klockgether J, Miethke N, Kubesch P, Bohn YS, Brockhausen I, Cramer N, et al. Intraclonal diversity of the Pseudomonas aeruginosa cystic fibrosis airway isolates TBCF10839 and TBCF121838: distinct signatures of transcriptome, proteome, metabolome, adherence and pathogenicity despite an almost identical genome sequence. Environ. Microbiol. 2013;15:191–210. doi: 10.1111/j.1462-2920.2012.02842.x. [DOI] [PubMed] [Google Scholar]
- Kohler T, Buckling A, van Delden C. Cooperation and virulence of clinical Pseudomonas aeruginosa populations. Proc. Natl Acad. Sci. USA. 2009;106:6339–6344. doi: 10.1073/pnas.0811741106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kukavica-Ibrulj I, Bragonzi A, Paroni M, Winstanley C, Sanschagrin F, O'toole GA, et al. In vivo growth of Pseudomonas aeruginosa strains PAO1 and PA14 and the hypervirulent strain LESB58 in a rat model of chronic lung infection. J. Bacteriol. 2008;190:2804–2813. doi: 10.1128/JB.01572-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larbig KD, Christmann A, Johann A, Klockgether J, Hartsch T, Merkl R, et al. Gene islands integrated into tRNA(Gly) genes confer genome diversity on a Pseudomonas aeruginosa clone. J. Bacteriol. 2002;184:6665–6680. doi: 10.1128/JB.184.23.6665-6680.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee DG, Urbach JM, Wu G, Liberati NT, Feinbaum RL, Miyata S, et al. Genomic analysis reveals that Pseudomonas aeruginosa virulence is combinatorial. Genome Biol. 2006;7:R90. doi: 10.1186/gb-2006-7-10-r90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leitao JH, Alvim T, Sa-Correia I. Ribotyping of Pseudomonas aeruginosa isolates from patients and water springs and genome fingerprinting of variants concerning mucoidy. FEMS Immunol. Med. Microbiol. 1996;13:287–292. doi: 10.1111/j.1574-695X.1996.tb00253.x. [DOI] [PubMed] [Google Scholar]
- Liang X, Pham XQ, Olson MV, Lory S. Identification of a genomic island present in the majority of pathogenic isolates of Pseudomonas aeruginosa. J. Bacteriol. 2001;183:843–853. doi: 10.1128/JB.183.3.843-853.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipuma JJ. The changing microbial epidemiology in cystic fibrosis. Clin. Microbiol. Rev. 2010;23:299–323. doi: 10.1128/CMR.00068-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lore NI, Cigana C, Riva I, de Fino C, Juhas M, Schwager S, et al. Cystic fibrosis-niche adaptation of Pseudomonas aeruginosa reduces virulence in multiple infection hosts. PLoS ONE. 2012;7:e35648. doi: 10.1371/journal.pone.0035648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mahenthiralingam E, Coenye T, Chung JW, Speert DP, Govan JR, Taylor P, et al. Diagnostically and experimentally useful panel of strains from the Burkholderia cepacia complex. J. Clin. Microbiol. 2000;38:910–913. doi: 10.1128/jcm.38.2.910-913.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manos J, Arthur J, Rose B, Tingpej P, Fung C, Curtis M, et al. Transcriptome analyses and biofilm-forming characteristics of a clonal Pseudomonas aeruginosa from the cystic fibrosis lung. J. Med. Microbiol. 2008;57:1454–1465. doi: 10.1099/jmm.0.2008/005009-0. [DOI] [PubMed] [Google Scholar]
- Martin K, Baddal B, Mustafa N, Perry C, Underwood A, Constantidou C, et al. Clusters of genetically similar isolates of Pseudomonas aeruginosa from multiple hospitals in the United Kingdom. J. Med. Microbiol. 2013;62(Pt.7):988–1000. doi: 10.1099/jmm.0.054841-0. [DOI] [PubMed] [Google Scholar]
- Mathee K, Narasimhan G, Valdes C, Qiu X, Matewish JM, Koehrsen M, et al. Dynamics of Pseudomonas aeruginosa genome evolution. Proc. Natl Acad. Sci. USA. 2008;105:3100–3105. doi: 10.1073/pnas.0711982105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCallum SJ, Gallagher MJ, Corkill JE, Hart CA, Ledson MJ, Walshaw MJ. Spread of an epidemic Pseudomonas aeruginosa strain from a patient with cystic fibrosis (CF) to non-CF relatives. Thorax. 2002;57:559–560. doi: 10.1136/thorax.57.6.559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller WL, Matewish MJ, McNally DJ, Ishiyama N, Anderson EM, Brewer D, et al. Flagellin glycosylation in Pseudomonas aeruginosa PAK requires the O-antigen biosynthesis enzyme WbpO. J. Biol. Chem. 2008;283:3507–3518. doi: 10.1074/jbc.M708894200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moskowitz SM, Ernst RK. The role of Pseudomonas lipopolysaccharide in cystic fibrosis airway infection. Subcell. Biochem. 2010;53:241–253. doi: 10.1007/978-90-481-9078-2_11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mowat E, Paterson S, Fothergill JL, Wright EA, Ledson MJ, Walshaw MJ, et al. Pseudomonas aeruginosa population diversity and turnover in cystic fibrosis chronic infections. Am. J. Respir. Crit. Care Med. 2011;183:1674–1679. doi: 10.1164/rccm.201009-1430OC. [DOI] [PubMed] [Google Scholar]
- Mulcahy LR, Burns JL, Lory S, Lewis K. Emergence of Pseudomonas aeruginosa strains producing high levels of persister cells in patients with cystic fibrosis. J. Bacteriol. 2010;192:6191–6199. doi: 10.1128/JB.01651-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naughton S, Parker D, Seemann T, Thomas T, Turnbull L, Rose B, et al. Pseudomonas aeruginosa AES-1 exhibits increased virulence gene expression during chronic infection of cystic fibrosis lung. PLoS ONE. 2011;6:e24526. doi: 10.1371/journal.pone.0024526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Carroll MR, Syrmis MW, Wainwright CE, Greer RM, Mitchell P, Coulter C, et al. Clonal strains of Pseudomonas aeruginosa in paediatric and adult cystic fibrosis units. Eur. Respir. J. 2004;24:101–106. doi: 10.1183/09031936.04.00122903. [DOI] [PubMed] [Google Scholar]
- Oliver A, Canton R, Campo P, Baquero F, Blazquez J. High frequency of hypermutable Pseudomonas aeruginosa in cystic fibrosis lung infection. Science. 2000;288:1251–1254. doi: 10.1126/science.288.5469.1251. [DOI] [PubMed] [Google Scholar]
- Parkins MD, Rendall JC, Elborn JS. Incidence and risk factors for pulmonary exacerbation treatment failures in patients with cystic fibrosis chronically infected with Pseudomonas aeruginosa. Chest. 2012;141:485–493. doi: 10.1378/chest.11-0917. [DOI] [PubMed] [Google Scholar]
- Pirnay JP, Cochez D, De Vos C, Bilocq F, Vanderkelen A, Zizi M, et al. Pseudomonas aeruginosa displays an epidemic population structure. Environ. Microbiol. 2002;4:898–911. doi: 10.1046/j.1462-2920.2002.00321.x. [DOI] [PubMed] [Google Scholar]
- Pirnay JP, Bilocq F, Pot B, Cornelis P, Zizi M, van Eldere J, et al. Pseudomonas aeruginosa population structure revisited. PLoS ONE. 2009;4:e7740. doi: 10.1371/journal.pone.0007740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahme LG, Stevens EJ, Wolfort SF, Shao J, Tompkins RG, Ausubel FM. Common virulence factors for bacterial pathogenicity in plants and animals. Science. 1995;268:1899–1902. doi: 10.1126/science.7604262. [DOI] [PubMed] [Google Scholar]
- Rakhimova E, Wiehlmann L, Brauer AL, Sethi S, Murphy TF, Tummler B. Pseudomonas aeruginosa population biology in chronic obstructive pulmonary disease. J. Infect. Dis. 2009;200:1928–1935. doi: 10.1086/648404. [DOI] [PubMed] [Google Scholar]
- Romling U, Kader A, Sriramulu DD, Simm R, Kronvall G. Worldwide distribution of Pseudomonas aeruginosa clone C strains in the aquatic environment and cystic fibrosis patients. Environ. Microbiol. 2005;7:1029–1038. doi: 10.1111/j.1462-2920.2005.00780.x. [DOI] [PubMed] [Google Scholar]
- Salunkhe P, Smart CH, Morgan JA, Panagea S, Walshaw MJ, Hart CA, et al. A cystic fibrosis epidemic strain of Pseudomonas aeruginosa displays enhanced virulence and antimicrobial resistance. J. Bacteriol. 2005;187:4908–4920. doi: 10.1128/JB.187.14.4908-4920.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott FW, Pitt TL. Identification and characterization of transmissible Pseudomonas aeruginosa strains in cystic fibrosis patients in England and Wales. J. Med. Microbiol. 2004;53:609–615. doi: 10.1099/jmm.0.45620-0. [DOI] [PubMed] [Google Scholar]
- Semblat JP, Doerig C. PAK in pathogen-host interactions. Cell Logist. 2012;2:126–131. doi: 10.4161/cl.20222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shankar J, Sueke H, Wiehlmann L, Horsburgh MJ, Tuft S, Neal TJ, et al. Genotypic analysis of UK keratitis-associated Pseudomonas aeruginosa suggests adaptation to environmental water as a key component in the development of eye infections. FEMS Microbiol. Lett. 2012;334:79–86. doi: 10.1111/j.1574-6968.2012.02621.x. [DOI] [PubMed] [Google Scholar]
- Smith EE, Buckley DG, Wu Z, Saenphimmachak C, Hoffman LR, D'argenio DA, et al. From the Cover: genetic adaptation by Pseudomonas aeruginosa to the airways of cystic fibrosis patients. Proc. Natl Acad. Sci. USA. 2006;103:8487–8492. doi: 10.1073/pnas.0602138103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Soyza A, Morris K, McDowell A, Doherty C, Archer L, Perry J, et al. Prevalence and clonality of Burkholderia cepacia complex genomovars in UK patients with cystic fibrosis referred for lung transplantation. Thorax. 2004;59:526–528. doi: 10.1136/thx.2003.010801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spencer DH, Kas A, Smith EE, Raymond CK, Sims EH, Hastings M, et al. Whole-genome sequence variation among multiple isolates of Pseudomonas aeruginosa. J. Bacteriol. 2003;185:1316–1325. doi: 10.1128/JB.185.4.1316-1325.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spratt BG, Hanage WP, Li B, Aanensen DM, Feil EJ. Displaying the relatedness among isolates of bacterial species – the eBURST approach. FEMS Microbiol. Lett. 2004;241:129–134. doi: 10.1016/j.femsle.2004.11.015. [DOI] [PubMed] [Google Scholar]
- Stewart RM, Wiehlmann L, Ashelford KE, Preston SJ, Frimmersdorf E, Campbell BJ, et al. Genetic characterization indicates that a specific subpopulation of Pseudomonas aeruginosa is associated with keratitis infections. J. Clin. Microbiol. 2011;49:993–1003. doi: 10.1128/JCM.02036-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stover CK, Pham XQ, Erwin AL, Mizoguchi SD, Warrener P, Hickey MJ, et al. Complete genome sequence of Pseudomonas aeruginosa PA01, an opportunistic pathogen. Nature. 2000;406:959–964. doi: 10.1038/35023079. [DOI] [PubMed] [Google Scholar]
- Syrmis MW, O'carroll MR, Sloots TP, Coulter C, Wainwright CE, Bell SC, et al. Rapid genotyping of Pseudomonas aeruginosa isolates harboured by adult and paediatric patients with cystic fibrosis using repetitive-element-based PCR assays. J. Med. Microbiol. 2004;53:1089–1096. doi: 10.1099/jmm.0.45611-0. [DOI] [PubMed] [Google Scholar]
- Tavares IM, Leitao JH, Fialho AM, Sa-Correia I. Pattern of changes in the activity of enzymes of GDP-D-mannuronic acid synthesis and in the level of transcription of algA, algC and algD genes accompanying the loss and emergence of mucoidy in Pseudomonas aeruginosa. Res. Microbiol. 1999;150:105–116. doi: 10.1016/s0923-2508(99)80028-x. [DOI] [PubMed] [Google Scholar]
- Totten PA, Lory S. Characterization of the type a flagellin gene from Pseudomonas aeruginosa PAK. J. Bacteriol. 1990;172:7188–7199. doi: 10.1128/jb.172.12.7188-7199.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toussaint B, Delic-Attree I, Vignais PM. Pseudomonas aeruginosa contains an IHF-like protein that binds to the algD promoter. Biochem. Biophys. Res. Commun. 1993;196:416–421. doi: 10.1006/bbrc.1993.2265. [DOI] [PubMed] [Google Scholar]
- Tummler B, Koopmann U, Grothues D, Weissbrodt H, Steinkamp G, von der Hardt H. Nosocomial acquisition of Pseudomonas aeruginosa by cystic fibrosis patients. J. Clin. Microbiol. 1991;29:1265–1267. doi: 10.1128/jcm.29.6.1265-1267.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vasseur P, Vallet-Gely I, Soscia C, Genin S, Filloux A. The pel genes of the Pseudomonas aeruginosa PAK strain are involved at early and late stages of biofilm formation. Microbiology. 2005;151:985–997. doi: 10.1099/mic.0.27410-0. [DOI] [PubMed] [Google Scholar]
- Wiehlmann L, Wagner G, Cramer N, Siebert B, Gudowius P, Morales G, et al. Population structure of Pseudomonas aeruginosa. Proc. Natl Acad. Sci. USA. 2007;104:8101–8106. doi: 10.1073/pnas.0609213104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winsor GL, Lam DK, Fleming L, Lo R, Whiteside MD, Yu NY, et al. Pseudomonas Genome Database: improved comparative analysis and population genomics capability for Pseudomonas genomes. Nucleic Acids Res. 2011;39:D596–D600. doi: 10.1093/nar/gkq869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winstanley C, Langille MG, Fothergill JL, Kukavica-Ibrulj I, Paradis-Bleau C, Sanschagrin F, et al. Newly introduced genomic prophage islands are critical determinants of in vivo competitiveness in the Liverpool Epidemic Strain of Pseudomonas aeruginosa. Genome Res. 2009;19:12–23. doi: 10.1101/gr.086082.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang L, Jelsbak L, Marvig RL, Damkiaer S, Workman CT, Rau MH, et al. Evolutionary dynamics of bacteria in a human host environment. Proc. Natl Acad. Sci. USA. 2011a;108:7481–7486. doi: 10.1073/pnas.1018249108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang L, Jelsbak L, Marvig RL, Damkiaer S, Workman CT, Rau MH, et al. Evolutionary dynamics of bacteria in a human host environment. Proc. Natl Acad. Sci. USA. 2011b;108:7481–7486. doi: 10.1073/pnas.1018249108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao J, Schloss PD, Kalikin LM, Carmody LA, Foster BK, Petrosino JF, et al. Decade-long bacterial community dynamics in cystic fibrosis airways. Proc. Natl Acad. Sci. USA. 2012;109:5809–5814. doi: 10.1073/pnas.1120577109. [DOI] [PMC free article] [PubMed] [Google Scholar]