

Abstract
Hydrogen sulfide (H2S) has proved to be a multifunctional signaling molecule in plants and animals. Here, we investigated the role of H2S in the decay of fresh-cut pears (Pyrus pyrifolia). H2S gas released by sodium hydrosulfide (NaHS) prolonged the shelf life of fresh-cut pear slices in a dose-dependent manner. Moreover, H2S maintained higher levels of reducing sugar and soluble protein in pear slices. H2S significantly reduced the accumulation of hydrogen peroxide (H2O2), superoxide radicals (•O2 −) and malondialdehyde (MDA). Further investigation showed that H2S fumigation up-regulated the activities of antioxidant enzymes ascorbate peroxidase (APX), catalase (CAT), and guaiacol peroxidase (POD), while it down-regulated those of lipoxygenase (LOX), phenylalanine ammonia lyase (PAL) and polyphenol oxidase (PPO). Furthermore, H2S fumigation effectively inhibited the growth of two fungal pathogens of pear, Aspergillus niger and Penicillium expansum, suggesting that H2S can be developed as an effective fungicide for postharvest storage. The present study implies that H2S is involved in prolonging postharvest storage of pears by acting as an antioxidant and fungicide.
Introduction
Hydrogen sulfide (H2S), traditionally been thought of as poisonous gas, has proved to be a gaseous signaling molecule in animals [1], [2]. Accumulating evidence unveiled its role as a gaseous regulator involved in various processes in plants, including seed germination, root organogenesis, abiotic stress tolerance, guard cell movement and autophagy [3], [4], [5], [6], [7], [8]. H2S can be generated from cysteine or sulfite by the enzymatic actions of O-acetylserine (thiol) lyase or sulfite reductase respectively, further suggesting that it might act as an endogenous regulator in plants [9]. In our previous studies, we found the role of H2S in delaying senescence of cut flowers in a wide spectrum of botanical species including herbaceous and woody plants [10]. More recently, H2S was found to prolong the postharvest shelf life of nonclimacteric fruit strawberry by acting as an antioxidant [11]. The concentration of the applied H2S required to delay senescence in strawberry is quite low and the endogenous content of H2S in H2S-fumigated strawberry is comparable to that of control fruit, suggesting that H2S fumigation to fruits could be safe [11]. However, whether H2S is implicated in ripening and senescence of climacteric fruits remains unknown.
Pear (Pyrus pyrifolia) is one of the most economically important fruit crops in temperate zones, but has a limited postharvest life. Pear is a typical climacteric fruit, where the respiratory climacteric precedes rapid fruit ripening. Many physiological disorders of pear fruit and microbial caused rot, appear during fruit ripening in storage and are considered to be associated with fruit senescence [12], [13]. Postharvest fruits undergo programmed senescence accompanied by various physiological and biochemical changes, including oxidative damages caused by reactive oxygen species (ROS) such as hydrogen peroxide (H2O2) and superoxide radicals (•O2 −) [14]. ROS are highly reactive and cause lipid peroxidation, which could lead to undesirable flavors and odors due to increased lipoxygenase (LOX) activity and malondialdehyde (MDA) content in fruit tissue [15], [16]. Antioxidant enzymes such as ascorbate peroxidase (APX), catalase (CAT) and peroxidase (POD) are applied to degrade ROS to cope with the oxidative challenge during fruit senescence [17]. The strategies to alleviate oxidative senescence of climacteric fruits included treatments with ROS antagonists, nitric oxide (NO) or 1-methylcyclopropene (1-MCP) during postharvest storage of peach (Prunus persica L.) and mango (Mangifera indica L.) [18], [19], [20], [21]. NO could delay ripening and improve the post-cutting quality of banana (Musa balbisiana Colla), peaches and apple (Malus pumila Mill.) [22], [23], [24]. Due to the similar functional characteristics of NO and H2S in animal system and plants [1], [8], we speculate that H2S might also be involved in maturation and senescence regulation in fruit.
NO was found to inhibit the growth of pathogens on postharvest fruit and vegetables [25]. In plants, L-cysteine desulfhydrase expression and activity which are responsible for H2S release are induced upon pathogen attack [26], further suggesting that the released H2S has a role in plant defence. Besides, inorganic and organic sulfur compounds have long being used as fungicides. Considering the similar physiological roles of H2S and NO [1], [8], we hypothesize that H2S has similar inhibitory effect on the growth of microbial pathogens in the background of fruit storage. Therefore, in the present study, we characterized the physiological responses of pears to H2S fumigation during storage and the inhibitory role of H2S on the growth of fungi isolated from pears. We show that H2S alleviated the decay of pear slices probably by an enhanced antioxidant system to eliminate excessive ROS and by inhibiting fungal infection on pears.
Results
H2S alleviates the decay of fresh-cut pears
H2S, released by H2S donor NaHS aqueous solutions, was applied to pear slices. As shown in Figure 1A, H 2S prolonged the shelf life of fresh-cut pears and alleviated flesh mealiness in a dose-dependent manner. Rot index of control tissue treated with water increased significantly after 2 day of storage and reached a maximum value on day 4. In contrast, rot index of pear slices treated with H2S was lower than water controls even after 4 days of storage (Figure 1B). The shelf life of pear slices treated with 2.0 mM NaHS was extended to 8 d, while concentrations higher than 2.0 mM showed no further positive effect. As shown in Figure 1C, pear slices showed a decrease in firmness with storage time in both water controls and NaHS treatment. However, fruits treated with 2 mM NaHS were significantly harder than the control group during the storage period (Figure 1C).
Figure 1. Hydrogen sulfide (H2S) delays the rotting of fresh-cut pear in a dose-dependent manner.

Pear slices were fumigated with H2S released from different concentrations of NaHS as shown in the lower right part of photographs (A), and the photographs were taken on day 0, 2 and 4, and the rot index (B) of pear slices was recorded every two days from day 0 to 10. (C) Change of firmness in fresh-cut pear for water treatment (CK) and 2 mM NaHS treatment (H2S) during storage at 20°C for 0−6 days. Data are presented as means ± SD (n = 6). * and ** in this figure and following ones stand for significant difference between CK and H2S at P<0.05 and P<0.01, respectively.
H2S increases the contents of soluble protein and reducing sugar in pear slices
The data in Figure 2 indicated that H2S treatment sustained higher level of soluble protein and reducing sugar in pear slices during storage compared with water controls. The content of soluble protein in pear slices increased during the first 2 d of storage regardless of treatments. Thereafter the content of soluble protein declined rapidly in water controls, while H2S treatment attenuated such decline (Figure 2A). The levels of reducing sugar in pear slices decreased in the first 3 days in both treatments, then increased gradually (Figure 2B). During the whole storage, H2S treatment sustained higher level of reducing sugar than water controls.
Figure 2. H2S increases (A) soluble protein and (B) reducing sugar in pears.
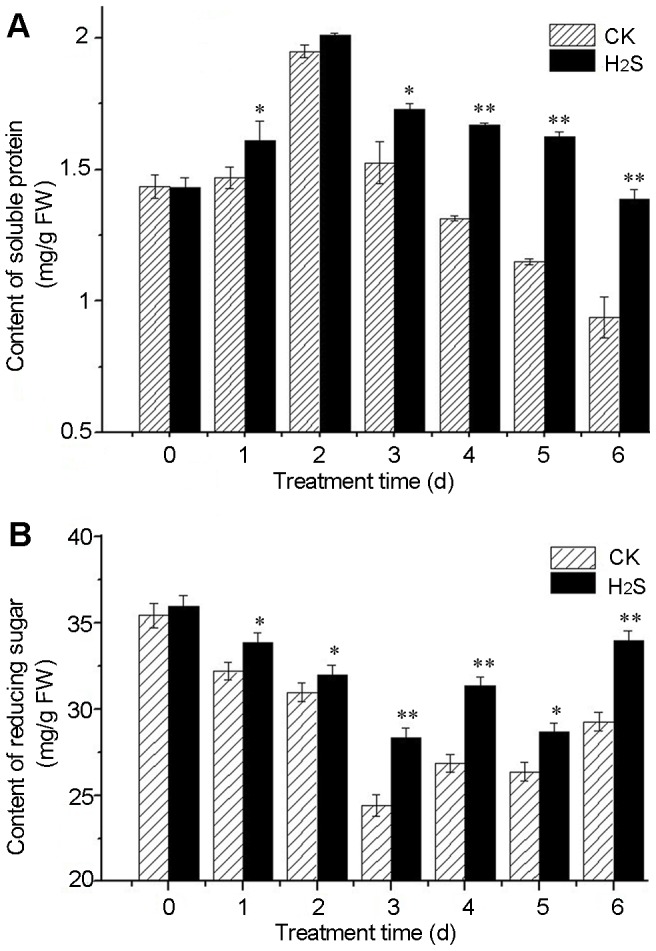
Soluble protein and reducing sugar were measured in slices treated with H2O (CK) or 2.0 mM H2S donor, NaHS (H2S) for 0−6 days. FW: fresh weight.
Changes of free amino acids in pears during storage
Table 1 shows that total amino acids in pear slices on 3 d of storage increased by ∼44% in water controls relative to fresh tissue. In contrast, H2S treatment for 3 days maintained comparable level of total free amino acids as in fresh pear slices. There were pronounced differences in the amino acid profiles in pear slices treated with H2S compared with water control. Cysteine, glutamic acid, histidine and alanine were all higher in H2S-treated tissue than those of water control, whereas threonine, methionine, isoleucine, tyrosine, phenylalanine and arginine were not detected in pear slices regardless of treatments.
Table 1. Effects of H2S treatment on the content of free amino acids (nmol/g FW) in pear slices determined after 3 days of storage at 20°C.
| Lys | His | Ala | Leu | Val | Asp | Ser | Glu | Gly | Cys | Thr | Met | Ile | Tyr | Phe | Arg | total | |
| 0 | 34.2 | ND | 215.4 | 77.9 | 129.5 | 1041.1 | 3225.5 | 10.2 | 32.9 | ND | ND | ND | ND | ND | ND | ND | 4766.8 |
| CK | 37.9 | 65.2 | 235.9 | 69.5 | 97.4 | 1609.4 | 4707.1 | ND | 19.5 | ND | ND | ND | ND | ND | ND | ND | 6841.8 |
| H2S | ND | 66.2 | 489 | 56.7 | 96.9 | 817.9 | 2760.7 | 73.3 | 14 | 1.3 | ND | ND | ND | ND | ND | ND | 4414 |
0 stands for fresh-cut pear slices; pear slices were treated with water (CK) or 2.0 mM H2S donor NaHS (H2S) for 3 days.
H2S down-regulates •O2 −, H2O2 and MDA contents in pears
Figure 3 shows that H2S fumigation sustained significantly lower levels of •O2 − (A), H2O2 (B) and MDA (C) than water controls. The •O2 − content of pear slices in water controls increased rapidly during the first 3 d of storage, followed by a slight decrease. In contrast, •O2 − levels in H2S-treated pear slices decreased gradually from 0.03 µg/g in fresh tissues to 0.01 µg/g on day 5 and 6 (Figure 3A).
Figure 3. H2S down-regulates superoxide anion (•O2 −) (A), hydrogen peroxide (H2O2) (B) and malonaldehyde (MDA) (C) in pears.
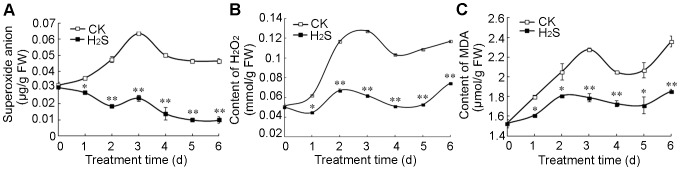
Pear slices were fumigated with 22S donor NaHS aqueous solution (H2S) or with water as the control groups (CK) for 0−6 d. Data are presented as means ± SD (n = 3). FW: fresh weight.
Similar change variations of H2O2 were observed in water controls and H2S treatment (Figure 3B). H2O2 in water controls accumulated quickly during the first 3 days of storage then kept a high stable level until the end. However H2S treatment maintained significantly lower level of H2O2. For instance, H2O2 content in water controls was about two-fold that of pear slices treated with H2S on day 3, 4 and 5 days (Figure 3B).
H2S treatment also kept a significantly lower level of MDA compared with water controls (Figure 3C). MDA content in water controls increased rapidly during the first 3 d of storage, while in H2S-treated pear slices, MDA accumulated much more slowly. Thereafter, MDA content of pear slices showed little variation with time in the presence or absence of H2S (Figure 3C).
H2S up-regulates the activities of APX, CAT, POD, and down-regulates the activities of LOX, PAL and PPO in pears
To further study the antioxidant role of H2S, the activities of enzymes involved in oxidative metabolism in plants, such as ascorbate peroxidase (APX), catalase (CAT), guaiacol peroxidase (POD), lipoxygenase (LOX), polyphenol oxidase (PPO) and phenylalanine ammonia lyase (PAL), are determined in pear slices. In water controls, APX activity increased rapidly and peaked on day 1 of storage and then declined to a level below that of fresh pears (Figure 4A). On the other hand, APX activity in H2S treatment increased steadily during the first 4 days of storage, and then decreased gradually (Figure 4A). After 3 days of storage, H2S treatment maintained significantly higher level of APX than water controls.
Figure 4. Role of H2S in the regulation of enzyme activities.

The activities of ascorbate peroxidase (APX) (A), catalase (CAT) (B), guaiacol peroxidase (POD) (C), lipoxygenase (LOX) (D), phenylalanine ammonia lyase (PAL) (E) and polyphenol oxidase (PPO) (F) were determined in pears. Pear slices were fumigated with 2.0 mM H2S donor NaHS aqueous solution (H2S) or with water as the control groups (CK) for 0−6 d. Data are presented as means ± SD (n = 3). FW: fresh weight.
H2S treatment sustained significantly higher level of CAT compared with water controls (Figure 4B). Similar change of CAT was observed in water controls and H2S treatment. CAT activity in water controls increased and peaked on d 2, and then decreased dramatically till day 6. In contrast, in H2S-treated tissue it increased steadily and peaked at 3 d, followed by a decline.
Similar change pattern of POD activity were observed in water controls and H2S treatment (Figure 4C). POD activity increased rapidly during the first 2 d of storage regardless of treatment and then decreased gradually in both treatments (Figure 4C). However, exposure to H2S maintained significantly higher level of POD activity in pear slices than water treatment.
LOX, which catalyzes the hydroperoxidation of polyunsaturated fatty acids and leads to the production of MDA, was detected in pear slices (Figure 4D). LOX activity increased rapidly in water-treated slices reaching a maximum of ∼90 U/g FW after 3 d of storage followed by a steep decline to levels relative to that of fresh tissues (Figure 4D). In contrast, the rise of LOX activity in H2S treatment was less pronounced and peaked to about 45 U/g FW on 3 d of storage followed by a decline. In general, the H2S treatment maintained significantly lower level of LOX activity compared with water controls during the whole storage.
Phenylalanine ammonia lyase (PAL) and polyphenol oxidase (PPO), which participate in the synthesis of free phenolics and catalyze oxidation of phenolics into brown-colored pigments [27], [28], were also examined in pear slices. Changes in PAL activity, an enzyme that catalyzes the first step in the phenylpropanoid pathway, are shown in Figure 4E. PAL activity increased gradually during the first 3 days of storage in both treatments. Thereafter, a burst PAL activity was observed on d 4 in water controls, whereas the activity fluctuated at a lower level in H2S treatment. Then PAL activity in water controls decreased but was always significantly higher than that of H2S treatment. Changes in the activities of PPO are illustrated in Figure 4F and show that H2S fumigation maintained significantly lower PPO activities compared with controls during the entire storage. PPO activities increased gradually in control pear during the first 2 days of storage, and then its activity decreased steadily until d 5 with an increase on d 6. In contrast, PPO activities in H2S treated pear slices decreased dramatically up to d 4, and then remained stable.
Fungicidal effects of H2S
To understand the phenomenon of reduced fungal growth under H2S fumigation as shown in Figure 1A, we isolated two pathogens A. niger and P. expansum from pears and investigated the effect of H2S on fungi growth in vitro and in vivo on pear fruits. Figure 5A and B show mycelium growth of A. niger and P. Expansum on medium, respectively. With increased concentrations of NaHS, the mycelium diameter of both pathogens grown on potato dextrose agar decreased and mycelium growth was severely inhibited even at 0.1 mM NaHS treatment. At 2 mM NaHS, mycelium growth of both fungi was completely inhibited suggesting a strong fungicidal effect of H2S on the two pathogens. The effects of H2S fumigation on fungal infection of pears are shown in Figure 5C and D. Higher concentrations of NaHS treatment severely inhibited the fungal propagation and reduced the lesion diameters on pears (lower part of Figure 5C and D). At 2 mM NaHS treatment, fungal infection of pears by A. niger and P. expansum was completely inhibited, suggesting an effective fungicidal role of H2S.
Figure 5. H2S inhibits the growth of Aspergillus niger and Penicillium expansum.

A. niger (A) and P. expansum (B), isolated from pear slices, were cultured on medium and subjected to the fumigation of different concentrations of NaHS solutions for 3 and 5 days respectively. The upper photographs of (A) and (B) indicate the growth of fungi subjected to different concentrations from left to right, upper to lower 0, 0.01, 0.05, 0.1, 0.5 and 2 mM NaHS, and the lower part of figure shows the diameters of fungi clones. The upper photographs of (C) and (D) show the pears infected by A. niger and P. expansum for 5 days and 8 days, respectively. The pears were subjected to different concentrations from left to right, upper to lower 0, 0.01, 0.05, 0.1, 0.5 and 2 mM NaHS, and the lower part of (C) and (D) shows the diameters of wounds caused by fungi. Data are presented as means ± SD (n = 4). Different letters indicate significant differences (p<0.05) between the treatments.
Discussion
Fruits are perishable during storage at room temperature due to rapid ripening [29]. An important goal for pear producers is to delay fruit spoilage and enhance storability to deliver benefits to consumers [30]. In this paper, we demonstrate that the application of H2S via the donor NaHS prolongs the storage of pears at 20°C, which is a temperature to mimic the lack of cold storage in developing countries, implying that H2S acts as a regulator in the senescence of postharvest fruit.
H2S is produced in small amounts by some cells of the mammalian body and has a number of biological signaling functions [2]. Since H2S occurs naturally in the body, the environment and the gut, enzymes existed in the body are capable of detoxifying it by oxidation to harmless sulfate [2]. Besides, several enzymes are responsible for the production of H2S in plants, such as L-cysteine desulfhydrase, sulfite reductase and β-cyanoalanine synthase [8]. Hence, low levels of H2S could be tolerated to human beings [2]. In this paper, the application concentration of NaHS solution is 0.5–2.5 mM, which could release about 0.05–0.5 ppm H2S gas in a sealed container (data not shown). In our previous work, the endogenous H2S level in H2S-treated strawberry was about 10%–20% higher than that of water control. Thus we propose that fumigation with trace H2S gas released from NaHS solution on pear slices could be safe.
Our data show that overproduction of reactive oxygen species (ROS) such as H2O2 and •O2 − contributed to accelerated fresh-cut fruit senescence, an observation that is consistent with other studies showing that ROS increased during senescence [31]. Therefore, maintaining high levels of antioxidants in pear slices is important in delaying fresh-cut fruit spoilage. Fruit tissues have evolved a complex antioxidant system to alleviate and repair oxidative damage [32]. Edible coatings with antibrowning agents can effectively improve the accumulationg of antioxidants vitamin C and total phenols, and finnally alleviate the senescence of fresh-cut pears during storage [32]. Overproduction of ROS up-regulates various antioxidant enzymes such as POD, CAT and APX to degrade ROS and prevent damages induced directly or indirectly by ROS [33]. Then we found that H2S treatment in pear slices are able to increase the activities of APX, CAT and POD (Figure 4A, B and C), thereby alleviating the accumulation of H2O2 and •O2 − (Figure 3A and B). The H2S-induced increase in antioxidant enzyme activities suggests that H2S could maintain the antioxidant potential to ameliorate ROS production during fruit storage. Fumigation of plants with H2S increased the content of cysteine and glutathione as a way to detoxify atmospheric H2S [34]. Consistently, we observed that H2S treatment increased the content of cysteine in pear slice (Table 1). Glutathione, which a major cellular thiol, plays important roles in ROS metabolism [35]. Whether H2S also acts through the increase of glutathione to alleviate senescence of postharvest fruits, still needs further research.
The senescence of fresh-cut pear is a complex and highly regulated process that involves lipid peroxidation that can be quantitated by measuring the content of MDA, which is a good indicator of the structural integrity of plant membranes [36]. Lipid peroxidation in plant tissues is regulated in part by lipoxygenase (LOX) and increased LOX activity is found to be associated with enhanced lipid peroxidation in fruit tissue [37]. LOX can catalyze the oxygenation of polyunsaturated fatty acids into lipidhydroperoxides (LOOHs) and result in the formation of hydroperoxides [38]. In our work, H2S treatment reduced the activities of LOX and a concomitant lowering of MDA content was observed. We speculate that the lowered MDA content and LOX activity in H2S treatment maintains the membrane integrity, and thus a delay in fruit tissue senescence.
Surface browning is frequently observed in fresh-cut fruits. Phenylalanine ammonia lyase (PAL) and polyphenol oxidase (PPO) are involved in the synthesis of free phenolics and catalyse the oxidation of phenolics into brown-colored pigments, and finally contribute to surface browning of fresh-cut fruits [27], [28]. The decreased PAL and PPO activities in H2S fumigated pear slices might alleviate the browning of fresh-cut pears.
In plants, stress conditions generally result in enhanced proteolytic activity to mobilize nutrients out of dying tissues into actively growing tissues in plants [39]. Compared with water controls, H2S treatment of pear slices decreased the accumulation of free amino acids, suggesting a protective role of H2S in delaying proteolysis during storage.
In plants, H2S is released by the activity of L-cysteine desulfhydrase during pathogen attack [26], suggesting that the evolution of H2S could be an important strategy of the plant to combat with fungal attack. In the present study, we isolated two pathogens A. niger and P. expansum from pears and found that 0.5 mM NaHS severely inhibited the growth of two pathogens on medium or on pears. Sulfur compounds have being used as fungicide from the time of antiquity. Marsh reported that H2S was toxic to germinating spores [40]. Then Haneklaus et al. calculated that a minimum uptake of 10 µM H2S/h by the pathogen could generate a fungicidal effect [41]. Besides inhibition of germinating spores, the fungicide mechanism of H2S towards postharvest fungi still needs further research.
In conclusion, the experimental data lend support to the hypothesis that H2S plays as an antioxidant and fungicide in pear slices, leading to a delay in senescence and decay. Our study provides an alternative strategy instead of synthetic fungicides in control of postharvest pathogens. However, the mechanisms whereby H2S functions in plants and fungi are largely unknown and further work is required to elucidate the possible mechanism of senescence alleviation and fungicide.
Materials and Methods
Materials
Pears (Pyrus pyrifolia cv. Dangshan), harvested from commercial orchards in Dangshan, China, were purchased from supermarket in Hefei, China. The pears are available broadly in the markets in Hefei, China. Thus no specific permissions were required for these activities. Thirty pears of uniform size were selected for experiments (weight: 300±5 g, diameter: 8.0×10.0 cm). Each pear was cut into six slices longitudinally and each slice from different pears was placed in the middle part of a sealed 3 L container. Aqueous solutions of sodium hydrosulfide (NaHS) were used as an H2S donor. 200 mL NaHS solutions at concentrations of 0.0, 0.5, 1.0, 1.5, 2.0, 2.5, 3 and 3.5 mM were placed in the bottom of containers to fumigate the slices for 0–6 days at 20°C, which is a temperature to mimic the lack of cold storage in developing countries. The relative humidity in the container is 90–95%. After treatment, pear slices were peeled, cored and stored at −20°C for further analysis. Each experiment was repeated three times.
Rot index of pears
Rot index was measured by Zhu and Zhou [42] and Ayala-Zavala et al. [43] with modifications. In each treatment, six pear slices were selected for investigating the number of decay fruits. All slices were classified in four ranks by the extent of rot. 0, no rot; 1, rot surface less than 10%; 2, rot surface between 10% and 30%; 3, rot surface between 30% and 50%; 4, rot surface more than 50%. The rot index was expressed by the following equation: Rot index = [∑(rank×quantity)/10×6]×100%. The rot classification was recorded every two days. The experiment was repeated three times.
Fruit firmness evaluation
Fruit firmness was measured at the equatorial part of each pear slice by a 5 mm diameter flat probe with a texture analyzer (Model TA XT plus, SMS) [44]. The penetration depth was 5 mm and the cross-head speed was 5 mm/s. Fruit firmness values were an average of 6 slices of pear. Data are presented as means ± SD (n = 6).
Measurement of soluble protein and reducing sugars
Soluble protein and reducing sugar contents were measured according to Bradford [45] and Miller [46], respectively. Fruit samples (5.00±0.05 g) were ground in 5 mL of phosphate buffer (pH 7.0, 200 mM). The homogenate was centrifuged at 10,000 g for 30 min, and the supernatant was used for determination of soluble protein and reducing sugar content. For soluble protein, 0.1 mL of supernatant was mixed with 0.9 mL of dH2O and 5 mL of Coomassie brilliant blue. Absorbance was recorded at 595 nm after 5 min. The results are expressed as mg/g FW (fresh weight). Each experiment was repeated three times.
Reducing sugar was measured via the dinitrosalicylic acid method. The supernatant (0.2 mL) was mixed with 1.5 mL of 3,5-dinitrosalicylic acid and 1.8 mL of distilled water; the mixture was heated at 100°C for 5 min, cooled, and added to 25 mL with distilled water. Reducing sugars were determined spectrophotometrically at 540 nm and the results are expressed as mg/g FW (fresh weight). Each experiment was repeated three times.
Measurement of free amino acids
For amino acid measurement, 5 g pear tissue was ground in 5 mL ethanol. The homogenate was centrifuged at 10,000 g for 10 min and the supernatant was heated at 80°C to removal ethanol. The residue was then diluted in water and filtered through a membrane (0.45 µm pore) and used for free amino acid assay. Free amino acids were analyzed with a Beckman Coulter S433D amino acid analyzer [47].
Determination of •O2 −, H2O2 and MDA
Contents of •O2 −, H2O2 and MDA were determined according to the methods described by Hu et al [11].
Activity assays of LOX, CAT, APX, POD, PAL and PPO
Lipoxygenase (LOX, EC 1.13.11.12) activity was determined following the description by Surrey [48], and catalase (CAT, EC 1.11.1.6) and ascorbate peroxidase (APX, EC 1.11.1.11) were assayed according to Garcia-Limones et al [49]. Pear tissues (5 g) were homogenized in 2.5 mL ice-cold 0.15 M phosphate buffer (pH 7.6) containing 1.0 mM ethylenediaminetetraacetic acid and 1% polyvinyl pyrrolidone. The homogenate was centrifuged at 10,000 g at 4°C for 30 min. The supernatant was used for activity measurement. For CAT activities, H2O2 was used as substrate. CAT activities were assayed by monitoring the decrease in absorbance at 240 nm. APX was determined in the presence 2.0 mM ascorbic acid and 2.0 mM ethylenediaminetetraacetic acid by measuring the decrease in absorbance at 290 nm. Analysis of POD was based on the oxidation of guaiacol using hydrogen peroxide by monitoring an increase in absorbance at 470 nm [11]. The reaction mixture contained 5.0 mL of 50 mM potassium phosphate buffer (pH 6.0), 1 mM of 1% hydrogen peroxide, 1 mM of 1% guaiacol and 10–20 µL of enzyme extract. One unit of CAT, APX or POD was defined as an increase or decrease of 0.01 in absorbance per minute under the assay conditions. The activities of antioxidant enzymes were expressed as U/g FW (fresh weight). Each experiment was repeated three times.
Activity of phenylalanine ammonia lyase (PAL) was determined by procedures described by Beaudoin-Eagan and Thorpe [50]. One unit of PAL activity was defined as a change of 0.01 OD value in absorbance at 290 nm per minute. The results were expressed as U/g FW (fresh weight). Each experiment was repeated three times.
Activity of polyphenol oxidase (PPO) was determined by procedures described by Benjamin and Montgomery [51]. Pear samples (2.00±0.05 g) were homogenized with 5.0 mL of sodium phosphate buffer (50 mM, pH 6.8). After centrifugation, the supernatant was used for the activity assay using catechol as substrate. One unit of PPO activity was defined as an increase of 0.01 OD value in absorbance at 410 nm per minute. The results were expressed as U/g FW (fresh weight). Each experiment was repeated three times.
Effect of H2S on fungal growth
The fungi Aspergillus niger and Penicillium expansum were isolated from the surface of pear fruits, and cultured on a potato-dextrose agar (PDA) medium (containing the extract from 200 g of boiled potato, 20 g of glucose and 20 g of agar in 1 L of distilled water) at 28°C in the dark [52]. Spore suspensions were prepared by flooding 6-day-old sporulating cultures of A. niger and P. expansum with sterile distilled water. The spore concentrations of the pathogen were determined with a hemacytometer and diluted with sterile distilled water to 1×105 spores/mL. Four aliquots of spore suspension (5 µL) were placed on 7 mm diameter Petri dishes maintained in sealed 3 L containers. 300 mL NaHS solutions at concentrations of 0.0, 0.01, 0.05, 0.1, 0.5, 2 mM were placed in the bottom of the sealed containers at 20°C with a relative humanity of 90−95% to fumigate fungi for the number of days indicated in the figure legends. For fungal infection of pears, pear fruits were wounded, four wounds per pear (5 mm diameter and 5 mm deep), using the tip of a sterile borer [37]. 20 µL spore suspension at 1×105 spores/mL of A. niger and P. expansum were inoculated onto each wound. After air-drying, the pears were stored under the same conditions as for fungi and fumigated with different concentrations of NaHS. Each experiment was repeated three times.
Statistical analysis
Each experiment was repeated three times. Significance was tested by one-way or two-way ANOVA, and the results were expressed as the mean values ± SD of three independent experiments. Fisher's least significant differences (LSD) were calculated following a significant (P<0.01 or P<0.05) t test.
Acknowledgments
We thank Russell Jones, University of California at Berkeley for critical reading of the manuscript.
Funding Statement
This work was financially supported by National Natural Science Foundation of China (http://www.nsfc.gov.cn/Portal0/default152.htm, project number 31271803, 31301820, 31300133), the Scientific Research Foundation for Returned Overseas Chinese Scholars (SRF for ROCS, SEM), the Natural Science Foundations of Anhui Province (11040606M85), and the Anhui Provincial Education Department (2012AJZR0028, ZD200910). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Wang R (2002) Two's company, three's a crowd: can H2S be the third endogenous gaseous transmitter? FASEB J 16: 1792–1798. [DOI] [PubMed] [Google Scholar]
- 2. Wang R (2012) Physiological implications of hydrogen sulfide: a whiff exploration that blossomed. Physiol Rev 92: 791–896. [DOI] [PubMed] [Google Scholar]
- 3. Papenbrock J, Riemenschneider A, Kamp A, Schulz-Vogt H, Schmidt A (2007) Characterization of cysteine-degrading and H2S-releasing enzymes of higher plants - from the field to the test tube and back. Plant Biol 9: 582–588. [DOI] [PubMed] [Google Scholar]
- 4. Zhang H, Hu LY, Hu KD, He YD, Wang SH, et al. (2008) Hydrogen sulfide promotes wheat seed germination and alleviates the oxidative damage against copper stress. J Integr Plant Biol 50: 1518–1529. [DOI] [PubMed] [Google Scholar]
- 5. Zhang H, Tang J, Liu XP, Wang Y, Yu W, et al. (2009) Hydrogen sulfide promotes root organogenesis in Ipomoea batatas, Salix matsudana, and Glycine max . J Integr Plant Biol 51: 1086–1094. [DOI] [PubMed] [Google Scholar]
- 6. García-Mata C, Lamattina L (2010) Hydrogen sulphide, a novel gasotransmitter involved in guard cell signalling. New Phytol 188: 977–984. [DOI] [PubMed] [Google Scholar]
- 7. Álvarez C, García I, Moreno I, Pérez-Pérez ME, Crespo JL, et al. (2012) Cysteine-generated sulfide in the cytosol negatively regulates autophagy and modulates the transcriptional profile in Arabidopsis . Plant Cell 24: 4621–4634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Lisjak M, Teklic T, Wilson ID, Whiteman M, Hancock JT (2013) Hydrogen sulfide: environmental factor or signalling molecule? Plant Cell Environ 36: 1607–1616. [DOI] [PubMed] [Google Scholar]
- 9. Rausch T, Wachter A (2005) Sulfur metabolism: a versatile platform for launching defence operations. Trends Plant Sci 10: 503–509. [DOI] [PubMed] [Google Scholar]
- 10. Zhang H, Hu SL, Zhang ZJ, Hu LY, Jiang CX, et al. (2011) Hydrogen sulfide acts as a regulator of flower senescence in plants. Postharvest Biol Technol 60: 251–257. [Google Scholar]
- 11. Hu LY, Hu SL, Wu J, Li YH, Zheng JL, et al. (2012) Hydrogen sulfide prolongs postharvest shelf life of strawberry and plays an antioxidative role in fruits. J Agric Food Chem 60: 8684–8693. [DOI] [PubMed] [Google Scholar]
- 12. Fonseca S, Hackler L, Zvara Á, Ferreira S, Baldé A, et al. (2004) Monitoring gene expression along pear fruit development, ripening and senescence using cDNA microarrays. Plant Sci 167: 457–469. [Google Scholar]
- 13. Gupta SM, Srivastava S, Sane AP, Nath P (2006) Differential expression of genes during banana fruit development, ripening and 1-MCP treatment: Presence of distinct fruit specific, ethylene induced and ethylene repressed expression. Postharvest Biol Technol 42: 16–22. [Google Scholar]
- 14. Oms-Oliu G, Soliva-Fortuny R, Martín-Belloso O (2008) Edible coatings with antibrowning agents to maintain sensory quality and antioxidant properties of fresh-cut pears. Postharvest Biol Technol 50: 87–94. [Google Scholar]
- 15. Raseetha S, Leong SY, Burritt DJ, Oey I (2013) Understanding the degradation of ascorbic acid and glutathione in relation to the levels of oxidative stress biomarkers in broccoli (Brassica oleracea L. italica cv. Bellstar) during storage and mechanical processing. Food Chem 138: 1360–1369. [DOI] [PubMed] [Google Scholar]
- 16. Noctor G, Foyer CH (1998) Ascorbate and glutathione: keeping active oxygen under control. Annu Rev Plant Physiol Plant Mol Biol 49: 249–279. [DOI] [PubMed] [Google Scholar]
- 17. Vicente AR, Martínez GA, Chaves AR, Civello PM (2006) Effect of heat treatment on strawberry fruit damage and oxidative metabolism during storage. Postharvest Biol Technol 40: 116–122. [Google Scholar]
- 18. Jimenez A, Creissen G, Kular B, Firmin J, Robinson S, et al. (2002) Changes in oxidative processes and components of the antioxidant system during tomato fruit ripening. Planta 214: 751–758. [DOI] [PubMed] [Google Scholar]
- 19. Ding ZS, Tian SP, Zheng XL, Zhou ZW, Xu Y (2007) Responses of reactive oxygen metabolism and quality in mango fruit to exogenous oxalic acid or salicylic acid under chilling temperature stress. Physiol Plant 130: 112–121. [Google Scholar]
- 20. Flores FB, Sánchez-Bel P, Valdenegro M, Romojaro F, Martínez-Madrid MC, et al. (2008) Effects of a pretreatment with nitric oxide on peach (Prunus persica L.) storage at room temperature. Eur Food Res Technol 227: 1599–1611. [Google Scholar]
- 21. Singh R, Dwivedi UN (2008) Effect of ethrel and 1-methylcyclopropene (1-MCP) on antioxidants in mango (Mangifera indica var. Dashehari) during fruit ripening. Food Chem 111: 951–956. [DOI] [PubMed] [Google Scholar]
- 22. Cheng G, Yang E, Lu W, Jia Y, Jiang Y, et al. (2009) Effect of nitric oxide on ethylene synthesis and softening of banana fruit slice during ripening. J Agric Food Chem 57: 5799–5804. [DOI] [PubMed] [Google Scholar]
- 23. Zhu LQ, Zhou J, Zhu SH, Guo LH (2009) Inhibition of browning on the surface of peach slices by short-term exposure to nitric oxide and ascorbic acid. Food Chem 114: 174–179. [Google Scholar]
- 24. Pristijono P, Wills RBH, Golding JB (2006) Inhibition of browning on the surface of apple slices by short term exposure to nitric oxide (NO) gas. Postharvest Biol Technol 42: 256–259. [Google Scholar]
- 25. Lazar EE, Wills RB, Ho BT, Harris AM, Spohr LJ (2008) Antifungal effect of gaseous nitric oxide on mycelium growth, sporulation and spore germination of the postharvest horticulture pathogens, Aspergillus niger, Monilinia fructicola and Penicillium italicum . Lett Appl Microbiol 46: 688–692. [DOI] [PubMed] [Google Scholar]
- 26. Bloem E, Riemenschneider A, Volker J, Papenbrock J, Schmidt A, et al. (2004) Sulphur supply and infection with Pyrenopeziza brassicae influence L-cysteine desulphydrase activity in Brassica napus L. J Exp Bot 55: 2305–2312. [DOI] [PubMed] [Google Scholar]
- 27. Nguyen TBT, Ketsa S, van Doorn WG (2003) Relationship between browning and the activities of polyphenoloxidase and phenylalanine ammonialyase in banana peel during low temperature storage. Postharvest Biol Technol 30: 187–193. [Google Scholar]
- 28. Mishra BB, Gautam S, Sharma A (2013) Free phenolics and polyphenol oxidase (PPO): the factors affecting post-cut browning in eggplant (Solanum melongena). Food Chem 139: 105–114. [DOI] [PubMed] [Google Scholar]
- 29. Bonghi C, Ramina A, Ruperti B, Vidrih R, Tonutti P (1999) Peach fruit ripening and quality in relation to picking time, and hypoxic and high CO2 short-term postharvest treatments. Postharvest Biol Technol 16: 213–222. [Google Scholar]
- 30. Villalobos-Acuña M, Mitcham EJ (2008) Ripening of European pears: the chilling dilemma. Postharvest Biol Technol 49: 187–200. [Google Scholar]
- 31. Aguayo E, Requejo-Jackman C, Stanley R, Woolf A (2010) Effects of calcium ascorbate treatments and storage atmosphere on antioxidant activity and quality of fresh-cut apple slices. Postharvest Biol Technol 57: 52–60. [Google Scholar]
- 32. Oms-Oliu G, Odriozola-Serrano I, Soliva-Fortuny R, Martín-Belloso O (2008) The role of peroxidase on the antioxidant potential of fresh-cut ‘Piel de Sapo’ melon packaged under different modified atmospheres. Food Chem 106: 1085–1092. [Google Scholar]
- 33. Reyes LF, Villarreal JE, Cisneros-Zevallos L (2007) The increase in antioxidant capacity after wounding depends on the type of fruit or vegetable tissue. Food Chem 101: 1254–1262. [Google Scholar]
- 34. Riemenschneider A, Nikiforova V, Hoefgen R, De Kok LJ, Papenbrock J (2005) Impact of elevated H2S on metabolite levels, activity of enzymes and expression of genes involved in cysteine metabolism. Plant Physiol Biochem 43: 473–483. [DOI] [PubMed] [Google Scholar]
- 35. Mhamdi A, Hager J, Chaouch S, Queval G, Han Y, et al. (2010) Arabidopsis GLUTATHIONE REDUCTASE1 plays a crucial role in leaf responses to intracellular hydrogen peroxide and in ensuring appropriate gene expression through both salicylic acid and jasmonic acid signaling pathways. Plant Physiol 153: 1144–1160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Lana MM, Tijskens LMM (2006) Effects of cutting and maturity on antioxidant activity of fresh-cut tomatoes. Food Chem 97: 203–211. [Google Scholar]
- 37. Hodges DM, Toivonen P (2008) Quality of fresh-cut fruits and vegetables as affected by exposure to abiotic stress. Postharvest Biol Technol 48: 155–162. [Google Scholar]
- 38. Duan XW, You YL, Su XG, Qu HX, Joyce DC, et al. (2007) Influence of the nitric oxide donor, sodium nitroprusside, on lipid peroxidation and anti-oxidant activity in pericarp tissue of longan fruit. J Hort Sci Biotech 82: 467–473. [Google Scholar]
- 39. Foyer CH, Descourvieres P, Kunert KJ (1994) Protection against oxygen radicals: an important defence mechanism studied in transgenic plants. Plant Cell Environ 17: 507–523. [Google Scholar]
- 40. Mars RW (1929) Investigations on the fungicidal action of sulphur. III. Studies on the toxicity of sulphuretted hydrogen and on the interaction of sulphur with fungi. Jour Pom Hort Sci 7: 237–250. [Google Scholar]
- 41.Haneklaus S, Bleom E, Schnug E (2007) Sulfur and plant disease. In: Datnoff L, Elmer W, Huber D (Eds.), Mineral elements and plant disease. Minneapolis, MN, USA: APS Press: St Paul, MN.
- 42. Zhu SH, Zhou J (2007) Effects of nitric oxide on ethylene production in strawberry fruit during storage. Food Chem 100: 1517–1522. [Google Scholar]
- 43. Ayala-Zavala JF, Wang SY, Wang CY, González-Aguilar GA (2004) Effect of storage temperatures on antioxidant capacity and aroma compounds in strawberry fruit. LWT - Food Sci Technol 37: 687–695. [Google Scholar]
- 44. Hernández-Muñoz P, Almenar E, Valle VD, Velez D, Gavara R (2008) Effect of chitosan coating combined with postharvest calcium treatment on strawberry (Fragaria×ananassa) quality during refrigerated storage. Food Chem 110: 428–435. [DOI] [PubMed] [Google Scholar]
- 45. Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–254. [DOI] [PubMed] [Google Scholar]
- 46. Miller GL (1959) Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Chem 31: 426–428. [Google Scholar]
- 47. Simpson RJ, Neuberger MR, Liu T (1976) Complete amino acid analysis of proteins from a single hydrolysate. J Biol Chem 251: 1936–1940. [PubMed] [Google Scholar]
- 48. Surrey K (1964) Spectrophotometric method for determination of lipoxidase activity. Plant Physiol 39: 65–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Garcia-Limones C, Hervás A, Navas-Cortés JA, Jiménez-Díaz RM, Tena M (2002) Induction of an antioxidant enzyme system and other oxidative stress markers associated with compatible and incompatible interactions between chickpea (Cicer arietinum L.) and Fusarium oxysporum f. sp. Ciceris . Physiol Mol Plant Pathol 61: 325–337. [Google Scholar]
- 50. Beaudoin-Eagan LD, Thorpe TA (1985) Tyrosine and phenylalanine ammonia lyase activities during shoot initiation in tobacco callus cultures. Plant Physiol 78: 438–441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Benjamin ND, Montgomery MW (1973) Polyphenol oxidase of royal ann cherries: purification and characterization. J Food Sci 38: 799–806. [Google Scholar]
- 52. Zheng X, Yu T, Chen R, Huang B, Wu C (2007) Inhibiting Penicillium expansum infection on pear fruit by Cryptococcus laurentii and cytokinin. Postharvest Biol Technol 45: 221–227. [Google Scholar]