

Abstract
A new crocodyliform, Sahitisuchus fluminensis gen. et sp. nov., is described based on a complete skull, lower jaw and anterior cervical vertebrae collected in the São José de Itaboraí Basin of Rio de Janeiro, Brazil. The specimen is one of the best preserved crocodyliforms from Paleocene deposits recovered so far and represents a sebecosuchian, one of the few clades that survived the Cretaceous-Paleogene biotic crisis. The new taxon is found in the same deposit as an alligatoroid, a group that experienced large diversification in the Paleogene. The sebecosuchian record suggests that after the Cretaceous-Paleogene biotic crisis, the less specialized members of this clade characterized by a higher number of teeth compared to the baurusuchid sebecosuchians survived, some having terrestrial habits while others developed a semi-aquatic life style (e.g., Lorosuchus). Starting in the Eocene, sebecid sebecosuchians became specialized with a more accentuated oreinirostry as observed in Sebecus and in Langstonia, but not showing the typical reduced dentition developed by the Cretaceous baurusuchid sebecosuchians. The basal position of Barinasuchus arveloi, a high-snouted Miocene sebecid, indicates the occurrence of an independent lineage sometime after the K-Pg biotic crisis that developed accentuated oreinirostry, suggesting a more complex history of the post-K-Pg crocodyliform radiation.
Introduction
Currently crocodyliforms are worldwide distributed in tropical and subtropical regions in relative low numbers and diversity, consisting of 24 to 30 species (e.g. [1]). All are considered semiaquatic ambushers but their fossil record reveals a much richer evolutionary history in terms of anatomy and ecomorphospaces [2], [3]. Particularly during the Cretaceous, the diversity of those reptiles was much higher and they occupied several distinct niches.
As a natural question, researchers tried to understand the crocodyliform decrease in diversity after the Cretaceous-Paleocene (K-Pg) extinction crisis but this discussion is hampered by the scarce nature of their remains in Paleocene deposits (e.g. [4]). Besides the marine dyrosaurids that have survived the K-Pg boundary and diversified during the Paleocene (e.g. [5], [6]), there are only a limited number of Paleocene specimens described so far, most of which are fragmentary and poorly preserved (e.g. [4], [7]–[9]). This contrasts with the high abundance of Late Cretaceous crocodyliforms, particularly in Brazil, which is even higher than in other Gondwanan areas.
During the exploration of the São José de Itaboraí Basin (Rio de Janeiro State, Southeast Brazil) that lasted for about five decades and ended in 1984 [10], hundreds of fossil vertebrates were collected (Figure 1). The vast majority is housed at the Earth Science Museum (now at the Companhia de Pesquisa de Recursos Minerais – CPRM), in Rio de Janeiro, and consists of fragmentary remains representing mainly mammals (e.g. [11], [12]). However, some reptiles have also been collected, including the remains of crocodyliforms [13], [14], with only one species formally proposed so far [15].
Figure 1. Itaboraí Basin location (A); (B) Itaboraí Basin lithochronoestratigraphic column.
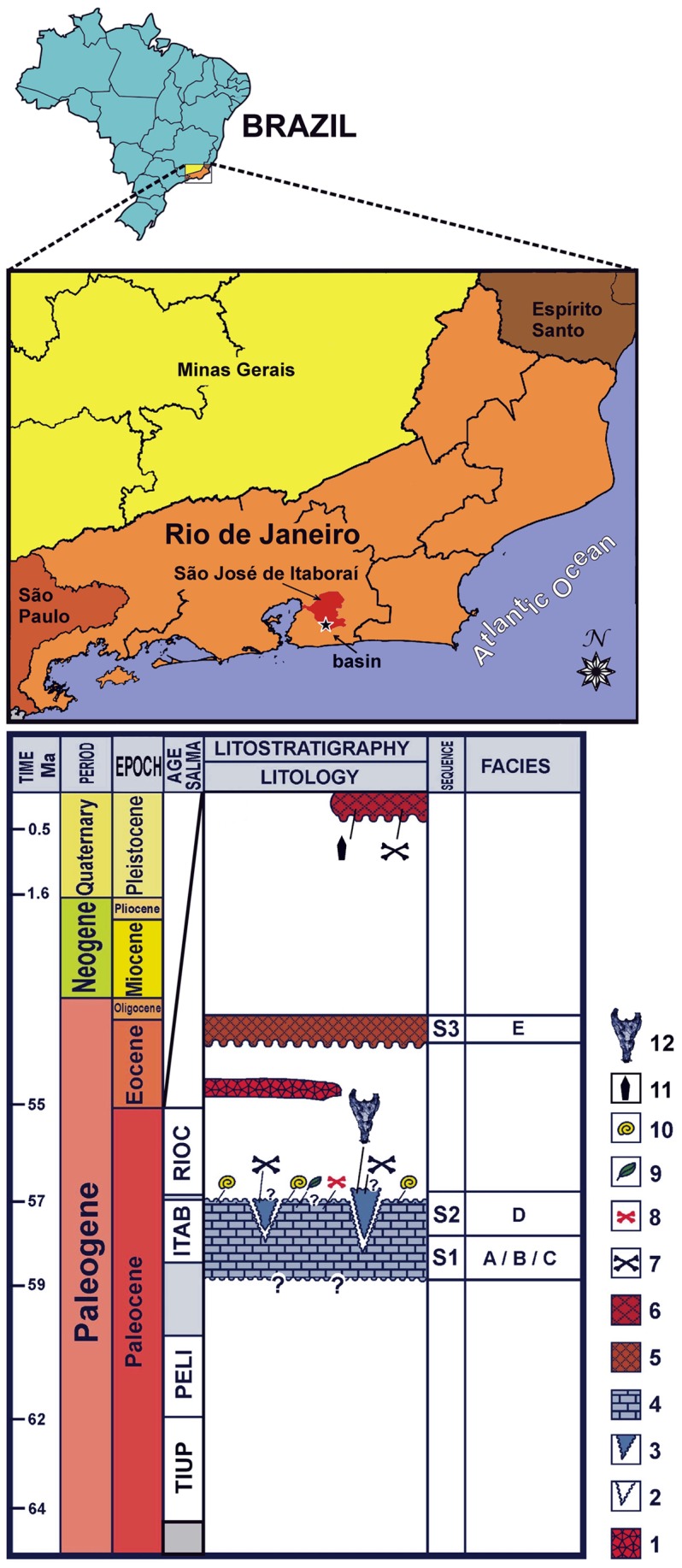
1, ankaramite layer; 2, karst channels and fissures; 3, infilling fissures (Sequence S2, Facies D); 4, main calcareous deposit (Sequence S1, Facies A, B and C); 5, alluvial deposits (Sequence S3, Facies E); 6, clastic sediments; 7, abundant vertebrate fossils; 8, rare vertebrate fossils; 9, plant remains; 10, terrestrial gastropod fossils; 11, archaeological artifacts; 12, Sahitysuchus fluminensis gen. et sp. nov. (MCT 1730-R). Stratigraphic column modified after [12], [21].
Among the few well preserved and more complete crocodyliform material is an almost complete skull and lower jaw (MCT 1730-R) that was briefly mentioned (but never figured) in the literature [8], [13], [16] and remained undescribed until now. This specimen represents a new taxon, Sahitisuchus fluminensis gen. et sp. nov., and shows that during the Paleocene the São José de Itaboraí crocodyliform fauna was composed by rather primitive (i.e., Sebecosuchia) and more derived (Alligatoridae) post-K-Pg taxa. Such a combination of sebecosuchians and eusuchians has not been previously reported in any deposit so far.
Materials and Methods
Phylogenetic Analysis
In order to access the phylogenetic position of Sahitisuchus fluminensis gen. et sp. nov., a phylogenetic analysis was performed using the data matrix published by Pol et al. (2012) [17]. Regarding Sebecus, we have followed Paollilo & Linares [9], who have restricted this genus to the type species (Sebecus icaerohinus) and regarded “S”. huilensis as belonging to the genus Langstonia. A total of 89 crocodyliform taxa including the new species and 347 characters were used. Parsimony analyses using TNT [18] with heuristics search strategy (10.000 replicates of Wagner trees, 15.000 max. tree in memory) by TBR algorithm were performed. The analyses were run using unordered and ordered characters (1, 3, 6, 10, 23, 37, 43, 44, 45, 49, 65, 67, 69, 73, 77, 79, 86, 90, 91, 96, 97, 104, 105, 106, 108, 116, 126, 140, 142, 143, 149, 167, 176, 182, 197, 226, and 339). Information for Sahitisuchus fluminensis gen. et sp. nov. used in the data matrix [17] is as follows:
10[0/1]?????12 ???0??-000 11011[0/1][0/1]00? 0100022110 100011?11? 1101010?10
?0103?[1/2]12- [2/3]10?101?21 ????????1? ?0???????? ??[1/2]01?00?? ?1?????[0/1]10
01??????0? ??0??00?01 0[0/1]2?1??001 010[0/1]11?1?0 ?1?0?0110- 1?01???110
[0/1]?100000?0 0101?10010 0001?[0/1]0001 01?1000010 ???00????? [0/1]0010[0/1]????
??0?0?0000 0100?0?0?? ?10?00?01? -???00?010 0??????[0/1]00 0001010??? ??????????
?????????? ?????????? ?????????? ???????
For more information see Supporting Information (Data S1, Figures S1 and S2).
Nomenclatural Acts
The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature, and hence the new names contained herein are available under that Code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix “http://zoobank.org/”. The LSID for this publication is: urn:lsid:zoobank.org:pub: 322EE489-D9D2-4CE6-9DAF-36E30C03881D. The electronic edition of this work was published in a journal with an ISSN, and has been archived and is available from the following digital repositories: PubMed Central, LOCKSS.
No permits were required for the described study, which complied with all relevant regulations. See appropriate section of Systematic Paleontology for locality, stratigraphic, repository and specimen number.
Results
Systematic Paleontology
Mesoeucrocodylia Whetstone & Whybrow, 1983 [19], sensu Benton & Clark, 1988 [20]
Sebecosuchia Simpson, 1937 [7]
Sebecidae Simpson, 1937 [7]
Sahitisuchus fluminensis gen. et sp. nov.
urn:lsid:zoobank.org:act:10A04487-436F-4509-BFDD-42B4DF6B8177
Derivation of name
Generic name Sahiti comes from the Xavante culture (sahi ti), one of the indigenous Brazilian inhabitants, meaning “to be angry” or “to be brave”, in allusion to warriors; and souchos, refers to the Egyptian crocodile god. Specific name fluminensis is a latinization of fluminense, designation of citizens born in the Rio de Janeiro State.
Type species
Almost complete skull and lower jaw, proatlas, intercentrum, the axis and the 3rd cervical vertebra (MCT 1730-R), housed at the Museu de Ciências da Terra, Companhia de Pesquisas de Recursos Minerais (CPRM), Rio de Janeiro, Brazil (cast at the Museu Nacional/UFRJ - MN 4711-V).
Type locality, and horizon and age
São José Farm, São José de Itaboraí Municipal District, ENE in the Rio de Janeiro Metropolitan Área (SE Brazil; 22°50′20″S and 42°52′30″W). Collected in the S2 sequence [21]; Itaboraian SALMA (South American Land Mammals Age), middle Upper Paleocene, 58,2-56,5 Ma [22].
Diagnosis
Sebecid crocodyliform with the following autapomorphies: mandible lacking external mandibular fenestra; and odontoid process fused to the axis with vertical anterior surface that lacks medial processes. The new species can be further distinguished from other sebecids by the following combination of characters: infraorbital jugal region with shallow ventrolateral depression (shared with Lorosuchus); shallow elliptical depression on the posterior surface of the quadrate close to the craniomandibular articulation (shared with Sebecus icaeorhinus); rough and rugose dorsal edge of supratemporal fossa (shared with Sebecus icaeorhinus); sharp, semilunate exoccipital posterior processes, directed medially (shared with Ayllusuchus); jugal posterior process higher than anterior process and lateral expanded (shared with Bretesuchus); rough longitudinal ridge on the lateroventral edge of angular and dentary, ending close to the mandibular symphysis level (shared with Bretesuchus, Sebecus).
Description and Comparisons
Overall the material of Sahitisuchus fluminensis is well preserved consisting of the skull, lower jaw and cervical elements (Figures 2–7; Tables 1, 2). Although some dorsoventral crushing is observable it was not severe to affect the shape of most cranial elements, including the rostral end that kept most of original anatomy. The most affected area was the more posterior portion of the skull, with some elements, particularly the supraoccipital displaced towards the foramen magnum.
Figure 2. Sahitysuchus fluminensis gen. et sp. nov. (MCT 1730-R), in dorsal view.

A, photo; B, illustration. 9–12th mlt, eighth to eleven left maxillary teeth; an, angular; anr, angular ridge; afo, mandibular adductor fossa; aof?, antorbital fenestra?; art, articular; bs, basisphenoid; bo, basioccipital; boc, basioccipital middle crest; bot, basioccipital basal tubera; chg, choanal groove; cq, cranio-quadrate passage; d, dentary; dhl, left hypertrophied replacement tooth; dhr, right hypertrophied dentary tooth; ec, ectopterygoid; expp, exoccipital posterior process; f, frontal; fä, foramen aereum; fcp, foramen caroticum posterius; fic, foramen intermandibularis caudalis; fme, median Eustachian foramen; fot, upper temporal fossa; fra, fractured area; fv, foramen vagi; int, intercentrum; j, jugal; jd, jugal latero-ventral depression; l; lachrymal; lptyp, lateral pterygoidal process (flange); ltf, laterotemporal fenestra; m, maxilla; m1l, first left maxillary tooth; m1r, first right maxillary tooth; m3l, third left maxillary tooth; m3r, third right maxillary tooth; m4l, fourth left maxillary tooth; m5l, fifth left maxillary tooth ; nc, nuchal crest; occ, occipital condyle; or, orbit; oti, otic incisure; oto, otoccipital; p, parietal; pl, palatine; pmt, posterior maxillary teeth; po, postorbital; pop, para-occipital process; pfr, prefrontal; pro, proatlas; pty, pterygoid; q, quadrate; qd, quadrate depression; qdc, quadrate dorsal crest; qj, quadratojugal; rap, retroarticular process; san, surangular; sanr, surangular lateral ridge; sf, siphoneal foramen; soc, supraoccipital; sof, suborbital fenestra; spl, splenial; sq, squamosal; sqp, squamosal posterior process (squamosal prong); utf, upper temporal fenestra; XII, twelfth cranial nerve exit. Scale bar: 100 mm.
Figure 7. Sahitisuchus fluminensis gen. et sp. nov. (MCT 1730-R) cervical vertebrae.

A, right lateral view of axis and third cervical vertebra; B, anterior view, showing the odontoid process. cvt cv3, centrum of third cervical vertebra; di ax, axis diapophysis; di cv3, diapophysis of third cervical vertebra; lpsop cv3, postspinal lamina of third cervical vertebra; na ax, axis neural arch; na cv3, neural arch of third cervical vertebra; ncs, neuro-central suture; ns ax, axis neural spine; ns cv3, neural spine of third cervical vertebra; od, odontoid process; pa ax, axis parapophysis; pa cv3, parapophysis of third cervical vertebra; poz ax, axis postzygapophysis; poz cv3, postzygapophysis of third cervical vertebra; prz cv3, prezygapophysis of third cervical vertebra. Scale bar: 10 mm.
Table 1. Measurements in mm of Sahitisuchus fluminensis gen. et sp. nov. (MCT 1730-R), adapted from [25] ∼, estimated measurement.
| 1. greatest width | 179 |
| 2. width of rostrum, posterior | 97 |
| 3. interorbital distance | 60 |
| 4. orbit length | 52.2 |
| 5. skull table width, anterior | 67 |
| 6. skull table length | 96 |
| 7. skull table width, posterior | ∼143.8 |
| 8. occipital condyle width | 23 |
| 9. occipital condyle height | 15 |
| 10. orbit width | 26.6 |
| 11. choana width | 39 |
| 12. choana length | 35.5 |
| 13. skull roof length | 67 |
| 14. quadrate condyle width | 37.7 |
| 15. supratemporal fossa width | 28.4 |
| 16. supratemporal fossa length | 34.4 |
| 17. palatal fenestra length | ∼48.6 |
| 18. pterygoid flanges width | ∼125.5 |
| 19. rostrum width at secondary dental peak | ∼50.3 |
| 20. rostrum width at notch (or fossa) for 4th mandibular tooth | ∼35.4 |
| 21. palatine bar width | 38 |
| 22. mandible length | ∼369 |
| 23. symphysis length | ∼84.2 |
| 24. retroarticular process length | 44 |
| 25. distance between supratemporal fossa/fenestra | 17 |
| 26. distance between medial borders of supratemporal fossae | 15.3 |
| 27. supratemporal fenestra width | 13.8 |
| 28. supratemporal fenestra length | 22 |
| 29. distance between supratemporal fossa and lateral margin of skull roof (at po–sq suture level) | 21 |
| 30. distance between supratemporal fossa and posterior margin of skull roof | 22 |
| 31. quadrate distal body length | 31.5 |
| 32. laterotemporal fenestra length | ∼38.6 |
| 33. occiput height (dorsal skull roof surface to occipital condyle) | 38 |
| 34. occiput height (dorsal skull roof surface to medial exoccipital ventral margin) | 54.7 |
Table 2. Sahitisuchus fluminensis gen. et sp. nov. (MCT 1730-R) teeth measurements in mm.
| Right tooth row | Left tooth row | ||||
| tooth | fabl | tch | tooth | fabl | tch |
| m1 | 4.8 | 14.8 | m1 | 10.9 | 6 |
| m2 | 5.2 | 9.2 | m2 | 14 | 7.5 |
| m3 | 31.5 | 11.7 | m3 | 28.7 | 15.8 |
| m4 | ∼11.7 | ∼5.8 | m4 | 10 | 6.4 |
| lmt | 6.4 | 6.1 | m5 | 8.2 | 5.7 |
| 1std (d4 ?) | ∼26.2 | ∼15.6 | 6thm | – | 9.3 |
| 2ndd | 14.8 | 7.7 | 7thm | 6.14 | 6 |
| 3thd | 8.6 | 5 | 8thm | 6.5 | 7.2 |
| 4thd | – | 6.6 | 9thm | – | 5.7 |
| 10thm | 6.4 | 6.1 | |||
| 1std (d4 ?) s | ∼12.8 | 7.1 | |||
1std (d4 ?), first exposed dentary tooth of the right side; 1std (d4 ?) s, first exposed replacement dentary tooth of the left side; d, dentary tooth; fabl, fore-after basal length; lmt, last exposed maxillary tooth; m, maxillary tooth; tch- tooth crown height; –, lack information; ∼, estimated measurement.
The premaxillae and the region of the external nares were broken off and the skull was slightly compressed dorsoventrally during preservation. The lower jaw is occluded with the skull and some bones lack the external bone cortex. In dorsal view, the skull of Sahitisuchus fluminensis is elongated, showing two slight constrictions at about the level of the 5th and 11th maxillary alveoli (Figures 2A, B). It differs from Sebecus icaeorhinus and Stolokrosuchus by being comparatively shorter [23], [24], but not as short as Lorosuchus [4]. Sahitisuchus does not show the same oreinirostral condition as Sebecus, Barinasuchus, Bretesuchus, Zulmasuchus and Langstonia [8], [9], [16], [23], [25].
Cranial bones
The skull-roof is flat and rectangular, being wider than long. The supratemporal fossa is much larger than the supratemporal fenestra (Figures 2A, B). This fossa is about three times smaller than the orbits. The distance between the supratemporal fossae is half that of the frontal inter-orbital width. The dorsal border of this fossa is surrounded by rugosities, forming an elevation that is more developed in the medial and lateral borders. This condition is similar to Sebecus and the peirosaurid Hamadasuchus, differing from any other mesoeucrocodyliforms (sensu Benton & Clark 1988 [20]) known to date. The palpebral bones, which are present as anterior and posterior elements in other sebecids (e.g. Sebecus, Lumbrera form [4], [26]) and specially in “peirosaurids” (e.g. Lomasuchus, Uberabasuchus, Montealtosuchus) [27]–[29], are not preserved. Even so, the orbit is placed rather laterally, a typical terrestrial sebecid feature, and not laterodorsally as in semi-aquatic crocodyliform morphotypes (e.g., Stolokrosuchus, Lorosuchus [4], [24], and extant species). The frontal is broad and triangular, with a low and smooth longitudinal crest running from the middle part to the posterior portion this bone. A longitudinal frontal crest is a common characteristic for many basal mesoeucrocodylian species, which includes some sebecosuchian taxa (e.g. Sebecus; Zulmasuchus; Iberosuchus macrodon; Pepesuchus, Lorosuchus [4], [16], [23], [30]. The frontal ornamentation is similar to that on the maxilla, with the wrinkles starting at the frontal longitudinal crest directed to the lateral margins. The jugal is very large and ornamented like most other cranial bones. The posterior ramus is laterally expanded and higher than the anterior one, an unusual feature within sebecosuchians only previously observed in Bretesuchus [8]. The ventrolateral margin is concave, a unique feature among basal mesoeucrocodylians. The quadratojugal takes part in the cranio-mandibular articulation forming the “double articulation” (sensu Buffetaut 1975 [31]), a feature absent in extant eusuchians but observed in all sebecid species and some other not closely related taxa (e.g. Trematochampsa, Libysocushus, Dyrosauridae). The squamosal shows a developed sculptured dorsal posteriorly pointed lobe (the squamosal posterior process or the squamosal prong [32] that is directed posteriorly and does not form a horn, similar to Hamadasuchus and Lomasuchus [27], [32]. Sebecus also shows such a developed process, but differs from Sahitisuchus by a more squared-shape posterior end [23]. The quadrates are massive and mostly unsculptured. The portion of the tympanic cavity formed by the quadrate is not multifenestrated like the one found in protosuchians, notosuchians [33] and baurusuchids (e.g. [34]). It also lacks the oblong concavity, which is characteristic of the Baurusuchidae [34]–[36] but, like Sebecus, Hamadasuchus and recent species, shows only two openings: the small, anterior preotic siphonial foramen, followed by the larger, oval otic incisure. The ventral portion of the tympanic membrane was supported by a low and sharp semicircular crest. The quadrate distal body extends beyond the occipital limits and bears a well-developed sharp crest that runs from the lateral region of the cranioquadrate passage to the end of this bone. A semi-elliptical shallow concavity in the most distal portion of the quadrate body, just medial to the quadratojugal-quadrate suture and anterior to the cranial-mandibular articulation is very conspicuous in the new species (Figures 2, 5). In ventral view the quadrate exhibits pronounced crests A and A′ [37] for M. adductor mandibulae posterior [38].
Figure 5. Sahitisuchus fluminensis gen. et sp. nov. (MCT 1730-R), in occipital view.

A, photo; B, illustration. For abbreviations see Figure 1. Scale bar: 100 mm.
Palatal region
The anterior process of the palatine projects over the maxillary palatal shelf with a “U-shaped” anterior margin [39], which extends well forward from the anterior margin of the suborbital fenestrae (Figure 3). The pterygoids are broad wing-shaped elements, similar to those found in Sebecus (MMP 235), being distinct from the broad quadrangular pterygoid of Zulmasuchus, peirosaurids (e.g. Montealtosuchus, Hamadasuchus) and derived eusuchians. The lateral border of the pterygoid flanges are arched and curved inwards similar to Zulmasuchus [9], [16]. In Bretesuchus this curvature is even more accentuated than in the latter species [8]. The pterygoid plate is slightly concave, very large and broad. The basicranium is not verticalized as found in Eusuchia but more verticalized than some basal crocodyliforms forms (e.g. baurusuchids and sphagesaurids) (Figures 3, 5). The choanae are positioned between the palatine and pterygoid, having a low and laminar choanal septum. The choanal groove (or fossa) is circular as the one of Sebecus and Barinasuchus but comparatively smaller than in these taxa.
Figure 3. Sahitisuchus fluminensis gen. et sp. nov. (MCT 1730-R), in ventral view.

A, photo; B, illustration. For abbreviations see Figure 1. Scale bar: 100 mm.
Occipital region
The occiput is about four times wider than high, which is partially attributed to crushing of the specimen (Figure 5). On the skull roof, the supraoccipital, this bone is as a relatively small forward pointed triangle which is inserted between the parietals. In occipital view this bone is relatively large and exhibits a prominent nuchal crest. The latter comprises the insertion point for M. spinalis capitis [40]. The posttemporal fenestrae are not well preserved due to compression, but the preserved part is very reduced with no postoccipital process. The supraoccipital descending portion is acute and reaches the foramen magnum. However, this seems more the product of the compression than to an autapomorphic feature of the new species. The exoccipital has a very distinct mediodorsal process similar to Ayllusuchus. This process is sharp and has a semilunate shape, comprising the insertion point for the M. rectus capitis sublimus and M. spinalis capitis. The basioccipital is trapezoidal and positioned oblique (∼45°) relative to the horizontal plane. This bone possesses a median elevated crest (insertion point for the M. rectus capitis anterior). The basisphenoid is short and completely verticalized, being little exposed both in occipital and palatal views. Similar inclination of basisphenoid and basioccipital is also found in some sebecids (e.g. Zulmasuchus and Bretesuchus) and peirosaurids.
Mandible
In ventral view the mandible shows an inverted “Y-shape” (Figures 3, 4). The robust mandibular symphysis is formed by the dentaries and splenials and reaches to opposite the fourth maxillary teeth while the dentary teeth are not visible, occupying about 21% of the mandibular length. In lateral view the anterior mandible portion is not as high as those of Bretesuchus and baurusuchids. The splenial forms about one-fourth of the mandibular symphysis and medially covers the Meckelian channel as a vertical and thick bone lamina. The mandibular lateral fenestrae is closed, an unusual feature in crocodyliforms and differing from all other sebecosuchian taxa. The angular exhibits a robust well-developed ventrolateral ridge that runs over almost the entire angular length, probably corresponding to the insertion area for the strongest component of the mandibular adductory musculature (M. adductor mandibulae internus pterygoideus ventralis [38]). The surangular takes part in the glenoid fossa, is stout and slightly ornamented. In lateral view, the suture with the dentary is gently convex. The dorsal margin of this bone is arched. A developed ridge with a smooth dorsal and rugose ventral surface is present below the glenoid fossa (Figure 3A, B). Except for Bretesuchus, in all other sebecosuchids where this region is preserved this crest is only incipient or poorly developed. With the retroarticular process, this structure is regarded as the probable insertion point for the components of the M. depressor mandibulae component [38], [41]. The articular forms about 60% of the glenoid fossa, like in other sebecids but unlike some other sebecosuchians, as in baurusuchids. The retroarticular process shows an elevated lateromedially crest just posterior to the glenoid fossa, a structure that does not allow palinal-propalinal jaw movements. The retroarticular process, formed by the articular and the surangular, is arched with a concave dorsal margin. The most medial posterior part, formed by the articular, is “tongue shaped”. Seen from posterior view, the posterior margin is inclined ventromedially forming an angle of about 40° relative the horizontal plane. A blunt crest runs longitudinally in the articular portion of this process. The retroarticular foramen aëreum is small and opens close to the medial margin of the retroarticular process, right after the glenoid fossa.
Figure 4. Sahitisuchus fluminensis gen. et sp. nov. (MCT 1730-R), in left lateral view.

A, photo; B, illustration. For abbreviations see Figure 1. Scale bar: 100 mm.
Dentition
The premaxillary teeth are not preserved but at least twelve maxillary teeth must have been present (Figures 4, 6). Due to the fact that the upper and lower jaws are occluded, most of the mandibular teeth are not visible.
Figure 6. Fourth mandibular tooth from the left side of Sahitisuchus fluminensis gen. et sp. nov. (MCT 1730-R), showing the serrations.

A, labial surface; B, detail for apex carina; C, detail for middle carina; D, detail for basal carina. Scale bar in A: 10 mm; B, C and D: 1 mm.
The new species has the crocodyliform plesiomorphic pattern of occlusion [3], with the hypertrophied dentary caniniform exposed laterally, occluding in the premaxillary-maxillary fossa, and the following maxillary teeth occluding buccally relative to the mandibular tooth row. The upper dentition is heterodont with three morphological arrangements, all showing serrated carinae formed by true denticles (sensu Langston 1975 [42]) (Figure 6). The first three maxillary teeth are ziphodont, curved posteriorly with pointed and buccolingually compressed crowns. The second dental morphotype is formed by the following two (perhaps three) teeth that are lanceolate and bear straight (i.e., not posteriorly curved) crowns. They are followed by progressively shorter teeth with blunt apices and a marked constriction between root and crown. The large caniform tooth shows about 3–3,5 denticles per mm (Figure 6).
Cervical elements
Several of the most anterior cervical elements such as the pro-atlas, intercentrum and odontoid process are described for first time in Sebecidae (Figures 3, 7). The pro-atlas was displaced over the left pterygoid flange. It is a small and laminar V-shaped bone with a low dorsal crest. Compared to modern crocodilians, this bone is rather conservative differing mainly by being narrower (Figure 3). The intercentrum, the only part identifiable of the atlas, was also displaced, being preserved over the left suborbital fenestra. This bone is robust, not laminar, with two blunt posteriorly directed processes for the first cervical ribs. The axis is preserved associated with a well-developed odontoid process that is similar to the one found in recent taxa by being massive and showing on each side blunt anterior tuberous processes (Figure 7). In the new species the anterior region is vertical and lacks the small medial processes found in at least some recent taxa. Furthermore, the odontoid process is fused with the axis with no visible suture indicating that this is most likely a very old individual [43]. The axis is well developed with a low, blade-like neural spine. The third cervical vertebra is amphicoelous, with a tall spike-like neural spine, inclined posteriorly, with a postspinal lamina that is bifurcated at the base (Figure 7). As in Sebecus (the only other sebecid where this part of the skeleton was described [17]), the diapophysis is divided by the neurocentral suture and in lateral view, the centrum shows a medial constriction and a trapezoidal shape, with anterior and posterior articulations inclined anteriorly. Among the differences with Sebecus, Sahitisuchus has more robust and broader diapophyses, and the length of the third cervical centrum is subequal compared to the axis.
Discussion and Conclusions
In order to investigate the phylogenetic position of Sahitisuchus fluminensis, we used the data matrix published by Pol et al. 2012 [17], who have considered all well-known sebecid taxa (see Data S1). The analysis was run through TNT, with characters either unordered or ordered and both results show that Sahitisuchus is a member of the Sebecidae (Figure 8). Furthermore, the addition of the new Brazilian taxon collapses the monophyletic genus Sebecus recovered in previous studies [17] and suggests that Barinasuchus occupies a basal position within the Sebecidae relative to Lorosuchus.
Figure 8. Biochronology of the Crocodylomorpha based on the strict consensus tree obtained by the phylogenetic analysis (see text for details) and recorded temporal range.

The overall crocodyliform record in number of specimens and taxa from Paleocene deposits is rather slim. This low diversity contrasts with the expressive crocodyliform record of the Cretaceous, where a great number of species thrived in a variety of ecological niches, particularly in the Gondwana, such as notosuchians (e.g. Uruguaysuchidae, Sphagesauridae), baurusuchids (e.g. Baurusuchus, Stratiotosuchus, Pabweshi), peirosaurids (e.g. Uberabasuchus, Pepesuchus, Hamadasuchus), mahajangasuchids (e.g. Mahajangasuchus and Kaprosuchus), and other taxa whose phylogenetic position is more controversial [e.g. 44].
Only three crocodyliform lineages are represented either before and after the K-Pg boundary: the marine Dyrosauridae, particularly abundant in coastal deposits of Africa [45], [46], the semiaquatic Crocodylia, recovered specially by alligatoroids from North and South America [15], [47]–[50]; and the terrestrial sebecosuchians (e.g. [17]). Dyrosaurids are very specialized and became quite diversified after the K-Pg boundary, becoming along with sharks the main marine predators after the demise of mosasaurs [6]. Although the Alligatoroidea were already present in the Late Cretaceous this group only diversified after the K-Pg biotic crises (e.g. [39], [49]), contrary to what happened with the sebecosuchians that became less diverse (with baurusuchids restricted to Upper Cretaceous [e.g. 34]).
So far, the only Paleocene deposit where members of the Alligatoroidea (Eocaiman itaboraiensis [15]) and Sebecosuchia (represented by Sahitisuchus) were recovered is the São José de Itaboraí Basin. Having its origin related to the separation of South America and Africa, resulting in the opening of the South Atlantic Ocean [51], this tectonic feature consists of a small half-graben with a NE-SW major axis of 1.400 m and a sedimentary sequence that reaches a maximum thickness of 125 m [52]. The main fossils described so far are mammals that record one of the earliest phases of the mammalian radiation in South America after the K-Pg biotic crisis (e.g. [53]). The inferred age of the Itaboraí Basin based on the mammalian fauna has been the matter of a recent debate [21], [53]–[55]. Despite this uncertainty, the new crocodyliform is part of the so-called S2 paleofauna, whose age (Itaboraian SALMA [55]) is considered middle Upper Paleocene varying in absolute terms between 61.8 million to 58.5 million years [12] or 58.5 million to 56.5 million years [22].
The co-occurrence of a remnant of the pre-K-Pg sebecosuchian and a post-K-Pg alligatoroid crocodyliform taxon, here represented by Sahitisuchus and Eocaiman [15], respectively, in the Paleocene deposits of the Itaboraí Basin is quite unusual and somewhat surprising. The taphonomic history of those specimens, as of other fossils found in this basin, has been difficult to retrieve, particularly due to the fact that all material was recovered from fissures and not detailed information about their collecting has been recorded. This raises the valid question if all fossils were synchronous [e.g.], [ 56–57]. In the lack of other information, some authors have used the color of the specimens to establish if they came from the same or distinct fissures that stands as a proxy for being regarded synchronous [56].
Regarding the crocodyliforms, it is clear that the sebecid and the alligatorid species came from distinct environments (terrestrial and semi-aquatic, respectively) and represent animals that lived around a freshwater lake before becoming preserved. Among the specimens attributed to Eocaiman, several show distinct colors suggesting that they come from distinct fissures [15]. The material of Sahitisuchus fluminensis is preserved in a greyish colored limestone, similar to some of the Eocaiman material. Furthermore, despite the questions about the correct absolute age, it has been proposed that the calcareous deposits of the São José de Itaboraí Basin were formed in a time span of 2 million years [57]–[58] or less. Therefore, we can conclude that Sahitisuchus and Eocaiman were either set apart for a comparatively short geological timespan or most likely co-occurred, the last hypothesis favored here.
One possible scenario that could explain the co-occurrence of Sahitisuchus and Eocaiman is that, right after the Cretaceous-Paleogene biotic crisis, only the less specialized crocodyliforms survived (e.g. [59]), except for the marine dyrosaurids that appear not to have been negatively affected by this event (e.g. [6]). Regarding sebecosuchians, this appears to be correct since the Cretaceous forms show high skulls and a marked specialization in the dentition that is quite reduced. The Cretaceous taxa Baurusuchus and Stratiotosuchus, for example, show only five maxillary teeth opposed to the 10 in the Paleogene Lorosuchus, Bretesuchus and Zulmasuchus, and 12 in Sahitisuchus. Furthermore, Paleocene sebecosuchians represented only by the Sebecidae show the posterior teeth blunt and not specialized as in the Cretaceous sebecosuchians. It is conceivable that the Paleocene sebecosuchians adopted a mixture of semi-aquatic and terrestrial lifestyles and therefore might have at least partially shared the same environments as Paleocene alligatoroids. After the Eocene, sebecosuchians became again more specialized, developing a higher and laterally compressed rostrum as observed in the Eocene Sebecus and the Upper Miocene Langstonia. They further show a trend to reduce dentition (e.g. Sebecus exhibiting nine maxillary teeth), although not approaching the reduction observed in the Cretaceous baurusuchid sebecosuchians.
The unexpected result in the phylogenetic study presented here with the addition of Sahitisuchus to the data matrix published by Pol et al. [17], is the basal position of the high-snouted Barinasuchus. Recovered from Miocene deposits, this very large sebecid is known from the anterior portion of the rostrum only [9]. If its phylogenetic position is correct, this species indicates the presence of a yet another independent sebecid lineage that sometime after the K-Pg biotic crisis developed accentuated oreinirostry (sensu [60]) independently from other sebecids, suggesting a more complex history of the post-K-Pg crocodyliform radiation.
Supporting Information
Topology resulted by heuristic analysis of unordered characters states. Bootstrap values above the lines (branches), at left and no-italic; Jacknife values above lines (branches), at right and italic; Bremer decay below the lines (branches). Data matrix from Pol et al., (2012) [17] with Sahitisuchus fluminensis added.
(JPG)
{kind=link}
Topology resulted by heuristic analysis of third seven ordered characters states. Bootstrap values above the lines (branches), at left and no-italic; Jacknife values above lines (branchs), at right and italic; Bremer decay below the lines (branches). Data matrix from Pol et al., (2012) [17] with Sahitisuchus fluminensis added.
(JPG)
{kind=link}
Phylogenetic Analyses.
(DOC)
Acknowledgments
We thank Jéssica Pontes da Silva (MN/UFRJ) for preparing the specimen, Orlando N. Grillo (MN/UFRJ) for helping with the pictures, Carla W. Gabriel (Microscophy Laboratory technician DEGEO/CCMN/UFRJ) for the microscopic images, and Lílian P. Bergqvist (DEGEO/CCMN/UFRJ) for information about the São José de Itaboraí Basin. We also would like to thank Chris Brochu (University of Iowa, EUA), James Clark (Washington University, EUA) and an unknown reviewer for comments and suggestions on that improved this contribution.
Funding Statement
AWAK acknowledges the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ nos. E-26/102.779/2008 and E-26/111.273/2010) and the ConselhoNacional de DesenvolvimentoCientífico e Tecnológico (CNPq no. 307276/2009-0). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Hekkala E, Shirley MH, Amato G, Austin JD, Charter S, et al. (2011) An ancient icon reveals new mysteries: mummy DNA resurrects a cryptic species within the Nile crocodile. Molecular Ecology 20: 4199–3215. doi: 10.1111/j.1365-294X.2011.05245.x. [DOI] [PubMed] [Google Scholar]
- 2. Brochu CA (2001) Crocodylian snouts in space and time: phylogenetic approaches toward adaptative radiation. American Zoologist 41: 564–585. [Google Scholar]
- 3. Brochu CA (2003) Phylogenetic approaches toward crocodylian history. Annual Review of Earth and Planetary Sciences 31: 357–397. [Google Scholar]
- 4. Pol D, Powell JE (2011) A new sebecid mesoeucrocodylian from the Rio Loro Formation (Paleocene) of north-western Argentina. Zoological Journal of the Linnean Society 163: 7–36. [Google Scholar]
- 5. Gasparini Z (1996) Biogeographic Evolution of the American Crocodilians. Münchner Geowissenschaftliche Abhandlungen, Reihe A, Geologie und Paläentologie 30: 159–184. [Google Scholar]
- 6. Barbosa JA, Kellner AWA, Viana MSS (2008) New dyrosaurid crocodylomorph and evidences for faunal turnover at the K-P transition in Brazil. Proceedings of The Royal Society B 275: 1385–1391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Simpson GG (1937) New reptiles from the Eocene of South America. American Museum Novitates 927: 1–3. [Google Scholar]
- 8. Gasparini Z, Fernandez M, Powell J (1993) New Tertiary Sebecosuchians (Crocodylomorpha) from South America: Phylogenetic Implications. Historical Biology 7: 1–19. [Google Scholar]
- 9. Paolillo A, Linares O (2007) Nueveos cocodrilos sebecosuchia del cenozoico suramericano (Mesosucia: Crocodylia). Paleobioloiga Neotropical 3: 1–25. [Google Scholar]
- 10. Klein VC, Bergqvist LP (2002) Excursão à Bacia de São José de Itaboraí, Rio de Janeiro. Arquivos do Museu Nacional 60: 245–256. [Google Scholar]
- 11. Paula-Couto C (1953) A Bacia Calcárea de Itaboraí e a tectônica da costa Sudeste do Brasil. Notas Preliminares e Estudos da Divisão de Geologia e Mineralogia do DNPM, Rio de Janeiro 75: 1–17. [Google Scholar]
- 12.Bergqvist LP, Moreira AL, Pinto DR (2005). Bacia de São José de Itaboraí, 75 Anos de História e Ciência. Companhia de Pesquisa de Recursos Minerais/Serviço Geológico do Brasil (CPRM/SGB). pp. 1–84.
- 13. Price LI, Paula-Couto C (1946) Vertebrados Terrestres do Eoceno na Bacia Calcárea de Itaboraí, Brasil. Bulletin, II Congresso Pan-Americano de Engenharia de Minas e Geologia, Rio de Janeiro 3: 149–173. [Google Scholar]
- 14. Kellner AWA, Campos DA (1999) Vertebrate paleontology in Brasil–a review. Episodes 22: 238–251. [Google Scholar]
- 15. Pinheiro AEP, Fortier DC, Pol D, Campos DA, Bergqvist LP (2012) A new Eocaiman (Alligatoridae, Crocodylia) from Itaboraí Basin, Paleogene of Rio de Janeiro, Brazil. Historical Biology 1–12. doi: 10.1080/08912963.2012.705838. [DOI] [Google Scholar]
- 16.Buffetaut E, Marshall LG (1991). A new crocodylian, Sebecus querejazus nov.sp. (Mesosuchia, Sebecidae) from the Santa Lucía Formation (early Paleocene) at Vila Vila, South-Central Bolivia. In: Suarez-Soruco R ed. Fósiles y Facies de Bolivia - Vol I Vertebrados. Revista Técnica de YPFB 12: : 545–557. [Google Scholar]
- 17. Pol D, Leardi JM, Lecuona A, Krause M (2012) Postcranial anatomy of Sebecus icaeorhinus (Crocodyliformes, Sebecidae) from the Eocene of Patagonia. Journal of Vertebrate Paleontology 32: 328–354. [Google Scholar]
- 18.Goloboff PA, Farris JS, Nixon KC (2008). TNT: Tree analysis using New Technologies. Program and documentation available from the authors and http://www.zmuc.dk/public/phylogeny.
- 19. Whetstone KN, Whybrow P (1983) A Cursorial Crocodilian from the Triassic of Lesotho (Basutiland, South Africa). Occasional Papers of the Museum of Natural History, University Kansas 106: 1–37. [Google Scholar]
- 20.Benton M, Clark JM (1988) Archosaur phylogeny and the relationships of the Crocodylia. In: Benton M, ed. The phylogeny and classification of the tetrapods. Oxford: Clarendon Press. pp 295–338. [Google Scholar]
- 21. Medeiros RA, Bergqvist LP (1999) Paleocene of the São José de Itaboraí Basin, Rio de Janeiro, Brazil: lithostratigraphy and biostratigraphy. Acta Geológica Leopoldensia 22: 3–22. [Google Scholar]
- 22.Marshall LG (1985). Geochronology and Land-Mammal biochronology of the transamerican faunal interchange. In: Steli FG, Webb SD eds. The Great American biotic interchange. New York Plenum Press. pp. 49–85. [Google Scholar]
- 23. Colbert EW (1946) Sebecus, Representative of a Peculiar Suborder of Fossil crocodilia from Patagonia. Bulletin of The American Museum of Natural History 87: 217–270. [Google Scholar]
- 24. Larsson HCE, Gado B (2000) A new Early Cretaceous crocodyliform from Niger. N Jb Geol Paläont Abh 217: 131–141. [Google Scholar]
- 25.Langston W, Gasparini Z (1997). Crocodilians, Gryposuchus, and the South American Gavials. In: Kay RF, Madden RH, Cifelli RL, Flynn J eds. Vertebrate Paleontology in the Neotropics, The Miocene Fauna of la Venta, Colombia. Smithsonian Institute Press, Washington and London.592 p. [Google Scholar]
- 26. Powell JE, Babot MJ, López DAG, Deraco MV, Herrera C (2011) Eocene vertebrates of nothwestern Argentina: annotated list. Cenozoic Geology of the Central Andes of Argentina 49–370. [Google Scholar]
- 27. Gasparini Z, Chiappe LM, Fernandez M (1991) A new Senonian peirosaurid (Crocodylomorpha) from Argentina and synopsis of the South American Cretaceous crocodylians. Journal of Vertebrate Paleontology 11: 316–333. [Google Scholar]
- 28. Carvalho IS, Ribeiro LCB, Avilla L (2004) Uberabasuchus terrificus sp. nov., a new Crocodylomorpha from the Bauru Basin (Upper Cretaceous), Brazil. Gondwana Research 7: 975–1002. [Google Scholar]
- 29. Carvalho IS, Vasconcellos FM, Tavares SAS (2007) Montealtosuchus arrudacamposi, a new peirosaurid crocodile (Mesoeucrocodylia) from the Late Cretaceous Adamantina Formation Brazil. Zootaxa 1607: 35–46. [Google Scholar]
- 30. Campos DA, Oliveira GR, Figueiredo RG, Riff D, Azevedo AK, et al. (2011) On a new peirosaurid crocodyliform from the Upper Cretaceous, Bauru Group, southeastern Brazil. Anais da Academia Brasileira de Ciências 83: 317–327. [DOI] [PubMed] [Google Scholar]
- 31. Buffetaut E (1975) Sur l'articulation entre le quadratojugal et le surangulaire de certains Crocodiliens fossiles. Comptes Rendus Acad Sc Paris, D 280: 2741–2743. [Google Scholar]
- 32. Larsson HCE, Sues H-D (2007) Cranial Osteology and Phytlogenetic Relationships of Hamadasuchus rebouli (Crocodyliformes: Mesoeucrocodylia) From The Cretaceous of Morocco. Zoological Journal of the Linnean Society 149: 533–567. [Google Scholar]
- 33. Hecht MK, Tarsitano SF (1983) On the cranial morphology of Protosuchia, Notosuchia and Eusuchia. Konstruktionsmorphologie 152: 657–668. [Google Scholar]
- 34. Montefeltro FC, Larsson HCE, Langer MC (2011) A New Baurusuchid (Crocodyliformes, Mesoeucrocodylia) from the Late Cretaceous of Brazil and the Phylogeny of Baurusuchidae. Plos One 6. e21916. doi: 10.1371/journal.pone.0021916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Riff D, Kellner AWA (2011) Baurusuchid crocodyliforms as theropod mimics: clues from the skull and appendicular morphology of Stratiotosuchus maxhechti (Upper Cretaceous of Brazil). Zoological Journal of the Linnean Society 163: 37–56. [Google Scholar]
- 36. Pinheiro AEP, Bertini RJ, Andrade MB, Martins Neto RG (2008) A new specimen of Stratiotosuchus maxhechti (Baurusuchidae, Crocodyliformes) from the Adamantina Formation (Upper Cretaceous), Southeastern Brazil. Revista Brasileira de Paleontologia 11: 37–50. [Google Scholar]
- 37.Iordansky NN (1973). The skull of the Crocodilia. In: Gans C ed. Biology of the Reptilia, Academic Press, London and New York 4. pp. 201–262. [Google Scholar]
- 38. Bona P, Desojo JB (2011) Osteology and Cranial Musculature of Caiman latirostris (Crocodylia: Alligatoridae). Journal of Morphology 1–16. doi: 10.1002/jmor.10894. [DOI] [PubMed] [Google Scholar]
- 39. Brochu CA (1999) Phylogenetics, Taxononomy, and Historical Biogeography of Alligatoroidea. Journal of Vertebrate Paleontology 19: 9–100. [Google Scholar]
- 40.Chiasson RB (1962). Laboratory Anatomy of the Alligator.University of Arizona, WM.C. Brown Company Publishers. pp. 1–36. [Google Scholar]
- 41. Iordansky NN (2000) Jaw Muscles Of The Crocodiles: Structure, Synonymy, and some implications on Homology and Functions. Russian Journal of Herpetology 7: 41–50. [Google Scholar]
- 42. Langston W (1975) Ziphodont crocodyles: Pristichampsus vorax (Troxell), a new combination, from the Eocene of North America. Fieldiana - Geologie, Chicago 33: 291–314. [Google Scholar]
- 43. Brochu CA (1996) Closure of Neurocentral sutures during crocodilian ontogeny: implications for maturity assessment in fossil Archosaurs. Journal of Vertebrate Paleontology 16 1: 49–62. [Google Scholar]
- 44. Kellner AWA (1987) Ocorrência de um novo crocodiliano no Cretáceo Inferior da Bacia do Araripe, Nordeste do Brasil. Anais da Academia Brasileira de Ciências 59 3: 219–232. [Google Scholar]
- 45. Jouve S, Bouya B, Amaghzaz MB (2005) A short-snouted dyrosaurid (Crocodyliformes, Mesoeucrocodylia) from the Paleocene of Morocco. Paläeontology 48: 359–369. [Google Scholar]
- 46. Jouve S, Bouya B, Amaghzaz MB (2008) A long-snouted dyrosaurid (Crocodyliformes, Mesoeucrocodylia) from the Paleocene of Morocco: Phylogenetic and Palaeobiogeographic implications. Palaeontology 51: 281–294. [Google Scholar]
- 47. Bartels WS (1984) Osteology and systematic affinities of the horned alligator Ceratosuchus (Reptilia, Crocodilia). Journal of Paleontology 58: 1347–1353. [Google Scholar]
- 48. Brochu CA (2004) Alligatorine phylogeny and the status of Allognathosuchus Mook, 1921. Journal of Vertebrate Paleontology 24: 857–873. [Google Scholar]
- 49. Brochu CA (2011) Phylogenetic relationschips of Necrosuchus ionensis Simpson, 1937 and the early history of caimanines. Zoological Journal of the Linnean Society 163: 228–256. [Google Scholar]
- 50. Bona P (2007) Una nueva especie de Eocaiman Simpson (Crocodylia, Alligatoridae) del Paleoceno Inferior de Patagonia. Ameghiniana 44: 435–445. [Google Scholar]
- 51.Riccomini C, Sant'Anna LG, Ferrari AL (2004). Evolução geológica do Rift Continental do Sudeste do Brasil. In: Mantesso-Neto V, Bertorelli A, Carneiro CDR, Brito Neves BB eds. Geologia do Continente Sul-Americano: evolução da obra de Fernando Flávio Marques de Almeida. Beca, São Paulo. pp. 393–405.
- 52. Rodrígues-Francisco BH, González BB, Peroba CEN, Guedes SC (1985) Estudo dos testemunhos de sondagem na Bacia de São José, Itaboraí, R. J. I. Furo SJ-2T. Paleontological Actas, VIII Congresso Brasileiro de Paleontologia, DNPM Série Geologia, 27; Secção Paleontologia e Estratigrafia 2, Rio de Janeiro, Brazil 651–652. [Google Scholar]
- 53. Muizon C, Brito IM (1993) Le Basin calcacarie de São José de Itaboraí (Rio de Janeiro, Brésil) ses relations fauniques avec le site de Tiupampa (Cochabamba, Bolivie). Annales Paléontologie 79: 233–269. [Google Scholar]
- 54. Gelfo JN, Goin FJ, Woodburne MO, Muizon C (2009) Biochronological relationships of the earliest south american paleocene mammals faunas. Palaeontolgy 52: 251–269. [Google Scholar]
- 55. Paula-Couto C (1952) Fossil mammals from the beginning of the Cenozoic in Brasil. Marsupialia: Polydolopidae and Borhyaenidae. American Museum Novitates 1559: 1–27. [Google Scholar]
- 56. Bergqvist LP, Almeida EB, Araújo-Júnior HI (2011) Tafonomia da assembléia fossilífera de mamíferos da “Fenda 1968”, Bacia de São José de Itaboraí, Estado do Rio de Janeiro, Brasil. Revista Brasileira de Paleontologia 14 1: 75–86. [Google Scholar]
- 57. Rage J-C (1998) Fossil snakes from the Paleocene of São José de Itaboraí, Brasil. Part I. Madtsoiidae, Aniliidae. Paleovertebrata 27: 109–104. [Google Scholar]
- 58. Marshall LG, Sempere T, Butler RF (1997) Chronostratigraphy of the mammal-bearing Paleocene of South America. Journal of South America Earth Sciences 10 1: 49–70. [Google Scholar]
- 59. Buffetaut E (1990) Vertebrate extinctions and survival across the Cretaceous–Tertiary boundary. Tectonophysics 171: 337–345. [Google Scholar]
- 60.Busbey AB III (1995). The structural consequences of skull flattening in crocodylians. In: Thomason J, ed. Functional morphology in vertebrate paleontology. Londom: Cambridge University Press. pp 171–192. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Topology resulted by heuristic analysis of unordered characters states. Bootstrap values above the lines (branches), at left and no-italic; Jacknife values above lines (branches), at right and italic; Bremer decay below the lines (branches). Data matrix from Pol et al., (2012) [17] with Sahitisuchus fluminensis added.
(JPG)
Topology resulted by heuristic analysis of third seven ordered characters states. Bootstrap values above the lines (branches), at left and no-italic; Jacknife values above lines (branchs), at right and italic; Bremer decay below the lines (branches). Data matrix from Pol et al., (2012) [17] with Sahitisuchus fluminensis added.
(JPG)
Phylogenetic Analyses.
(DOC)