

Abstract
Macrophages are involved in many essential immune functions. Their role in cell-autonomous innate immunity is reinforced by interferon-γ (IFN-γ), which is mainly secreted by proliferating type 1 T helper cells and natural killer cells. Previously, we showed that IFN-γ activates autophagy via p38 mitogen-activated protein kinase (p38 MAPK), but the biological importance of this signalling pathway has not been clear. Here, we found that macrophage bactericidal activity increased by 4 hr after IFN-γ stimulation. Inducible nitric oxide synthase (NOS2) is a major downstream effector of the Janus kinase–signal transducer and activator of transcription 1 signalling pathway that contributes to macrophage bactericidal activity via nitric oxide (NO) generation. However, no NO generation was observed after 4 hr of IFN-γ stimulation, and macrophage bactericidal activity at early stages after IFN-γ stimulation was not affected by the NOS inhibitors, NG-methyl-l-arginine acetate salt and diphenyleneiodonium chloride. These results suggest that an NOS2-independent signalling pathway is involved in IFN-γ-mediated bactericidal activity. We also found that this macrophage activity was attenuated by the addition of the p38 MAPK inhibitors, PD 169316, SB 202190, and SB 203580, or by the expression of short hairpin RNA against p38α or the essential factors for autophagy, Atg5 and Atg7. Collectively, our results suggest that the IFN-γ-mediated autophagy via p38 MAPK, without the involvement of NOS2, also contributes to the ability of macrophages to kill intracellular bacteria. These observations provide direct evidence that p38 MAPK-mediated autophagy can support IFN-γ-mediated cell-autonomous innate immunity.
Keywords: autophagy, cell-autonomous innate immunity, interferon-γ, macrophage
Introduction
Macrophage participates in many essential functions such as tissue homeostasis and clearance of infection.1,2 The type II interferon (IFN), IFN-γ, a cytokine that is mainly secreted by activated T helper type 1 (Th1) T lymphocytes and natural killer (NK) cells, is a powerful macrophage activator.3,4 Interferon-γ induces tyrosine phosphorylation of the signal transducer and activator of transcription 1 (STAT1) protein via Janus kinase 1 (JAK1) and JAK2.5–7 Subsequently, STAT1 dimers bind to IFN-stimulated response elements and induce the transcription of many IFN-stimulated genes.5,6 In addition to the JAK–STAT1 pathway, expression of another gene is mediated by IFN-γ via STAT1-independent pathways and modulates various signalling cascades, including those involving myeloid differentiating factor 88 (MyD88), p38 mitogen-activated protein kinase (MAPK), phosphatidylinositol 3-kinase (PI3K), and protein kinase C.7–14 The major immune factors in the signalling pathway downstream of IFN-γ are MHC class I and II; inflammatory and pyrogenic cytokines; chemokines; and antimicrobial proteins, such as inducible nitric oxide synthase (NOS2; also known as iNOS), phagocyte oxidase, and immune guanosine triphosphatases (GTPases).5–7,15–18
Notably, IFN-γ is critical for cell-autonomous innate immunity against intracellular bacteria, such as Listeria, Mycobacteria and Salmonella.17,19–21 The antimicrobial enzyme NOS2 is largely considered responsible for the bactericidal activity, via nitric oxide (NO) generation, of IFN-γ-activated macrophages against intracellular bacteria.22,23 However, NOS2-knockout (NOS2−/−) mice that had been infected with virulent Mycobacterium tuberculosis have been found to survive significantly longer and exhibit some control of lung M. tuberculosis growth when compared with mice lacking IFN-γ or IFN-γ receptor.24 This observation suggested that IFN-γ-dependent, NOS2-independent immunity against intracellular bacteria exists. Recently, it has been shown that, in addition to NOS2, IFN-γ-inducible immune GTPases, including p47 immunity-related GTPases (p47 IRGs) and p65 guanylate-binding proteins (p65 GBPs), regulate autophagy and contribute to the disposal of intracellular pathogens.17,18,20,24–28
Autophagy has emerged as a major immune defence pathway and this cascade can be provoked by host-derived cytokines, IFN-γ, or pattern recognition receptors, including Toll-like receptors and nucleotide-binding oligomerization domain-like receptors.25,26,29–35 It has been shown that IFN-γ controls autophagy via several types of IFN-γ-inducible immune GTPases belonging to the IRG family and the GBP family.18,25–28,36,37 More recently, we have shown that, in addition to the IFN-inducible GTPase pathway, the p38 MAPK pathway contributes to autophagy activation in the IFN-γ-stimulating cells.38 Interferon-γ is able to activate autophagy through at least two different pathways, the conventional STAT1- and Irgm1-dependent pathway and an alternative p38 MAPK-dependent, STAT1-independent pathway. However, the biological role of IFN-γ-induced autophagy via p38 MAPK remains unclear.
In this study, we demonstrated that macrophage bactericidal activity increased at 4 hr after IFN-γ stimulation in an STAT1- and NOS2-independent manner. Furthermore, this macrophage bactericidal activity that occurred early after IFN-γ stimulation was attenuated by the inhibition of p38 MAPK or autophagic function. These results suggest that the autophagy mediated by p38 MAPK, without the influence of NOS2, also contributes to the ability of macrophages to kill intracellular bacteria. To our knowledge, this study is the first to document that p38 MAPK-mediated autophagy can activate IFN-γ-mediated cell-autonomous innate immunity.
Materials and methods
Reagents
Recombinant mouse IFN-γ was purchased from R&D Systems (Minneapolis, MN) and used at a concentration of 200 U/ml. NG-methyl-l-arginine acetate salt (l-NMMA) and diphenyleneiodonium chloride (DPI) were obtained from Sigma (St Louis, MO) and used at a concentration of 500 or 10 μm. PD 169316 and SB 202190 were obtained from Cayman (Ann Arbor, MI) and used at a concentration of 10 μm. SB 203580 was bought from Calbiochem (Darmstadt, Germany) and used at a concentration of 5 μm.
Mammalian cell culture
RAW 264.7 cells were obtained from the American Type Culture Collection (Manassas, VA) and maintained in RPMI-1640 medium containing 10% fetal bovine serum, 10 mm HEPES and 1 mm sodium pyruvate. The primary bone-marrow derived macrophages (BMMs) were generated from C57BL/6 mice, as reported previously.38 The lentiviral vectors used for expressing short hairpin (sh)RNA against IFN-γR1, STAT1 and Atg7 have been described previously.38 The plasmids for expressing shRNA as a non-target control and for expressing shRNA against Atg5 or p38α were constructed using pLKD.neo38 and the Addgene pLKO.1 protocol (http://www.addgene.org). The RNAi sequences were as follows: for the non-target shRNA control, 5′-CAACAAGATGAAGAGCACCAA-3′; for Atg5, 5′-GCAGAACCATACTATTTGCTT-3′; for p38α, 5′-CCTCTTGTTGAAAGATTCCTT-3′. The ViraPower Lentiviral Expression system (Invitrogen, Carlsbad, CA) was used to co-transfect the viral vector into 293FT (Invitrogen) to produce lentiviruses. The resulting viral supernatant was used for the transfection of RAW 264.7 cells or BMMs, and then stable knockdown (KD) cells were selected with G418 (BD Clontech, Palo Alto, CA).
Bacterial culture
Listeria monocytogenes EGD (serovar 1/2a) was a generous gift from Dr Masao Mitsuyama (Kyoto University Graduate School of Medicine, Kyoto, Japan). It was grown overnight in brain–heart infusion broth (BD Biosciences, Sparks, MD) at 37° and shaken. Listeria monocytogenes cells were washed with RPMI-1640 medium once and used in an infection assay. Salmonella enterica serovar Typhimurium (RIMD1985009) was provided by the Research Institute for Microbial Diseases, Osaka University (Osaka, Japan), and was grown overnight in Luria–Bertani broth (Sigma).
Measurement of bacterial growth in the macrophages
RAW 264.7 cells or BMMs were infected with L. monocytogenes for 1 hr at a multiplicity of infection (MOI) of 5 or with S. typhimurium for 10 min at an MOI of 10, and then the cells were washed three times with PBS. Following this, the cell culture medium was changed to new RPMI-1640 containing 50 μg/ml of gentamicin (Sigma) to exclude the bacteria, which were not taken up by the macrophages. After 0 or 4 hr of Listeria infection, or after 1 or 4 hr of Salmonella infection, the macrophages were lysed with 0·1% Triton X-100, and living bacteria were quantified by the colony-forming unit (CFU) method. The growth rate of bacteria in the cells over a 4-hr period was calculated. For each time-point, counts were obtained from three independent experiments.
Measurement of NO production
Levels of NO were measured as the accumulation of nitrite in the cell culture medium. The nitrite level was determined spectrophotometrically with Griess reagent (Sigma). RAW264.7 cells or BMMs were treated with or without 200 U/ml IFN-γ for 4 or 24 hr. Briefly, 250 μl of cell culture supernatant was mixed with an equal volume of Griess reagent. Following incubation for 10 min, the absorbance at 550 nm was measured, and values were quantified against a standard curve of sodium nitrite.
Western blotting
The macrophages were lysed in cell lysis buffer (50 mm Tris, pH 7·5, 1% Triton X-100 and 150 mm NaCl) plus protease inhibitor mixture (Roche, Mannheim, Germany), and centrifuged at 16 400 g for 20 min. The supernatants were used as cell lysates, and were subjected to SDS–PAGE before transferring to PVDF membranes. Western blotting was carried out with the following antibodies: anti-IFN-γR polyclonal antibody (pAb) and anti-Atg7 pAb (GeneTex, Irvine, CA), anti-STAT1 pAb (GenScript, Piscataway, NJ), anti-GAPDH monoclonal antibody (mAb) (6C5; Santa Cruz Biotechnology, Santa Cruz, CA), anti-phospho-p38, p38, Atg5 pAb (Cell Signaling Technology, Danvers, MA), ant-LC3 mAb (2G6; Nano Tools, Hamburg, Germany), horseradish peroxidase-conjugated anti-mouse IgG antibody and horseradish peroxidase-conjugated anti-rabbit IgG antibody (Jackson ImmunoResearch Laboratories, West Grove, PA). The expression of GAPDH was used as the internal control. The intensity of bands was quantified using ImageJ software (National Institutes of Health, Bethesda, MD).
Statistical analysis
Statistical analyses for differences between group means were analysed using Student's t-test. All metrics are given as mean ± standard deviation values.
Results
Macrophage bactericidal activity increased at 4 hr after IFN-γ stimulation in an IFN-γ R1-dependent, but STAT1-independent manner
Previously, we have demonstrated that autophagy, which plays key roles in the host defence pathway, is activated by IFN-γ stimulation for 4 hr.38 In this study, we analysed the function of macrophages that had been stimulated with IFN-γ for 4 hr, to elucidate the biological significance of IFN-γ-activated autophagy via p38 MAPK. Macrophage bactericidal activity is potentiated by IFN-γ stimulation and this effect is predominantly guided by NO, which is produced from arginine by IFN-inducible NOS2.22,39 In mouse macrophage-like RAW 264.7 cells, the expression of NOS2 mRNA was observed at 5 hr after IFN-γ (100 U/ml) stimulation, and NO production remained low until 10 hr after IFN-γ treatment.40,41 Moreover, our result also showed that NO was barely produced at 4 hr after IFN-γ stimulation (Fig. 1a). However, an elevation in bactericidal activity against both intracytosolic (L. monocytogenes) and intravacuolar (S. typhimurium) pathogens was observed in the macrophages that had been treated with IFN-γ for 4 hr (Fig. 1b,c). This kind of IFN-γ-mediated bactericidal activity was inhibited in IFN-γ receptor 1 KD cells, but not in STAT1-KD cells (Fig. 2). These results suggested that bactericidal activity is reinforced at an early stage of IFN-γ stimulation in a STAT1- and NO-independent manner. Unlike NO production, autophagy activation has previously been observed in RAW 264.7 cells and BMMs at 4 hr after IFN-γ stimulation,38 we therefore expected that autophagy may contribute to bactericidal activity at an early stage of IFN-γ stimulation.
Figure 1.

Macrophage bactericidal activity increased on interferon-γ (IFN-γ) stimulation for 4 hr. RAW 264.7 cells or the primary bone-marrow derived macrophages (BMMs) were treated with 200 U/ml IFN-γ for 4 or 24 hr. The nitrite level in the cell culture medium was determined by the Griess method (a). The cells were then infected with Listeria monocytogenes (b) or Salmonella typhimurium (c), and bacterial growth in the macrophages was calculated on the basis of colony-forming units (CFU). The values shown are the mean ± standard deviation values obtained from three independent experiments. *P < 0·01; **P < 0·02.
Figure 2.

Macrophage bactericidal activity increased 4 hr after interferon-γ (IFN-γ) stimulation in an IFN-γR-dependent, signal transducer and activator of transcription 1 (STAT1) -independent manner. (a, b) RAW 264.7 cells, bone-marrow-derived macrophages (BMMs), or stable knockdown cells were treated with 200 U/ml IFN-γ for 4 hr. The cells were then infected with Listeria monocytogenes (a) or Salmonella typhimurium (b), and bacterial growth in the macrophages was calculated on the basis of colony-forming units (CFU). The values shown are the mean ± standard deviation values obtained from three independent experiments. *P < 0·01; **P < 0·1; NS, not significant. (c) The cells were lysed in cell lysis buffer containing protease inhibitors, and the cell lysates were then subjected to SDS–PAGE analysis and Western blotting.
NOS2 is not involved in macrophage bactericidal activity in the early stages after IFN-γ stimulation
Next, we used the NOS inhibitors, l-NMMA and DPI, to rule out the possibility of participation of NOS2 and NO in macrophage bactericidal activity immediately after IFN-γ stimulation. The bactericidal activity at 4 hr after IFN-γ stimulation was not affected by either NOS inhibitor (Fig. 3a,b). The effects of these NOS inhibitors were confirmed by NO production in the macrophages that had been treated with IFN-γ for 24 hr, because no NO production was observed immediately after IFN-γ stimulation (Figs 1a and 3c). Both l-NMMA and DPI were effective at these concentrations (500 μm for l-NMMA and 10 μm for DPI). These results suggested that macrophage bactericidal activity in the early stages following IFN-γ stimulation is independent of NOS2 activity.
Figure 3.

Inducible nitric oxide synthase (NOS2) activity is not required for macrophage activation in the early stages after interferon-γ (IFN-γ) stimulation. RAW 264.7 cells or bone-marrow-derived macrophages (BMMs) were treated with 200 U/ml IFN-γ in the presence or absence of 500 μm of l-NMMA or 10 μm of DPI for 4 hr (a, b) or 24 hr (c). (a, b) The cells were then infected with Listeria monocytogenes (a) or Salmonella typhimurium (b), and bacterial growth in the cells was calculated on the basis of colony-forming units (CFU). (c) The nitrite level in the cell culture medium was determined by the Griess method. The values shown are the mean ± standard deviation values obtained from three independent experiments. *P < 0·01; **P < 0·02; ***P < 0·05; ****P < 0·1; NS, not significant.
The p38 MAPK signalling pathway is employed for IFN-γ-mediated pathogen clearance
Interferon-γ activates various signalling cascades, including MyD88, p38 MAPK, PI3K, and protein kinase C.8,9,12–14 In previous studies, we have demonstrated that 4 hr after IFN-γ stimulation, autophagy may be activated in macrophages.38 Therefore, we next investigated whether p38 MAPK-mediated autophagy contributes to bactericidal activity (Figs 4 and 5). We found that IFN-γ-mediated autophagy activation and bactericidal activity were inhibited in the presence of p38 MAPK inhibitors (PD 169316, SB 202190 and SB 203580) (Fig. 4) or in the p38α KD cells (Fig. 5), suggesting that macrophage activation at an early stage following IFN-γ stimulation depends on p38 MAPK.
Figure 4.
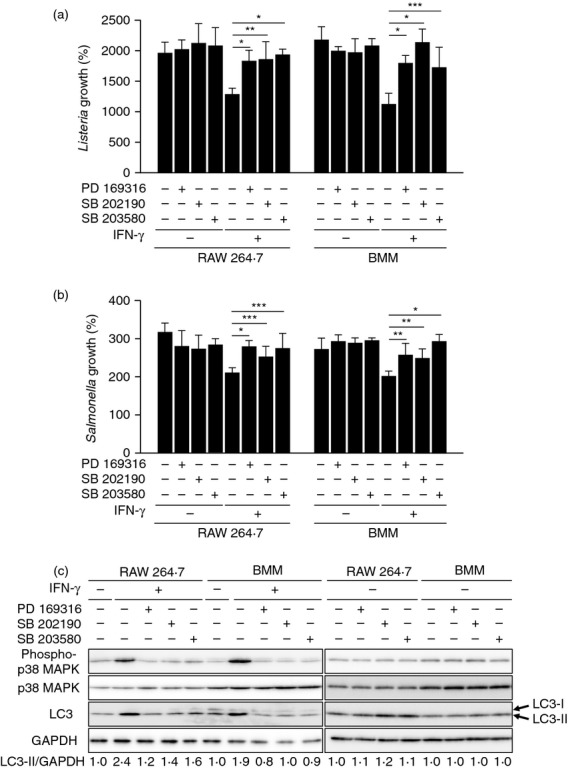
Macrophage activation in the early stages after interferon-γ (IFN-γ) stimulation is reduced by the p38 mitogen-activated protein kinase (MAPK) inhibitors. RAW 264.7 cells or bone marrow-derived macrophages (BMMs) were treated with 200 U/ml IFN-γ in the absence or presence of p38 MAPK inhibitors (10 μm of PD 169316, 10 μm of SB202190, or 5 μm of SB 203580) for 4 hr. (a, b) The cells were then infected with Listeria monocytogenes (a) or Salmonella typhimurium (b), and bacterial growth in the cells was calculated on the basis of colony-forming units (CFU). The values shown are the mean ± standard deviation values obtained from three independent experiments. *P < 0·01; **P < 0·05; ***P < 0·1. (c) The cells were lysed in cell lysis buffer containing protease inhibitors, and the cell lysates were then subjected to SDS–PAGE analysis and Western blotting. Densitometric LC3-II/GAPDH ratios are shown underneath the blot.
Figure 5.

Macrophage activation in the early stages after interferon-γ (IFN-γ) stimulation was reduced in p38α knockdown cells. (a, b) RAW 264.7 cells, bone-marrow-derived macrophages (BMMs), or stable knockdown cells were treated with 200 U/ml IFN-γ for 4 hr. The cells were then infected with Listeria monocytogenes (a) or Salmonella typhimurium (b), and bacterial growth in the macrophages was calculated on the basis of colony-forming units (CFU). The values shown are the mean ± standard deviation values obtained from three independent experiments. *P < 0·01; **P < 0·02; ***P < 0·05. (c) The cells were lysed in cell lysis buffer containing protease inhibitors, and the cell lysates were then subjected to SDS–PAGE analysis and Western blotting. Densitometric LC3-II/GAPDH ratios are shown underneath the blot.
Autophagy plays an important role in pathogen elimination by IFN-γ-stimulated macrophages
Next, to confirm whether p38 MAPK-mediated autophagy is involved in the function of IFN-γ-activated macrophages, autophagy-deficient Atg5 or Atg7 KD macrophage cells were used (Fig. 6). The antibacterial effect observed at 4 hr after IFN-γ treatment was blocked in autophagy-deficient cells, suggesting that macrophage bactericidal activity in the early stages following IFN-γ stimulation is mediated by autophagy. Our findings collectively suggested that, in IFN-γ-activated macrophages, bactericidal activity is quickly strengthened by p38 MAPK-mediated autophagy, rather than by NO. This action may be advantageous in macrophages before full activation.
Figure 6.

Autophagy plays a critical role in macrophage activation in the early stages after interferon-γ (IFN-γ) stimulation. RAW 264.7 cells, bone-marrow-derived macrophages (BMMs), and stable knockdown cells were treated with 200 U/ml IFN-γ for 4 hr. (a, b) The cells were infected with Listeria monocytogenes (a) or Salmonella typhimurium (b). The bacterial growth in the cells was calculated on the basis of colony-forming units (CFU). The values shown are the mean ± standard deviation values obtained from three independent experiments. *P < 0·01; **P < 0·05; ***P < 0·1; NS, not significant. (c) The cells were lysed in cell lysis buffer containing protease inhibitors, and the cell lysates were then subjected to SDS–PAGE analysis and Western blotting. Densitometric LC3-II/GAPDH ratios are shown underneath the blot.
Discussion
Autophagy has emerged as a major immune defence pathway; it also contributes to inflammation by facilitating an IFN-γ response and signal transduction.34,42 We have previously shown that the p38 MAPK signalling cascade also activates autophagy in IFN-γ-stimulated macrophages, but the effect of p38 MAPK-mediated autophagy on macrophage function was unknown.38 In this study, we found that macrophage bactericidal activity increased at 4 hr after IFN-γ stimulation in a STAT1- and NOS2-independent, p38 MAPK- and autophagy-dependent manner. Furthermore, this macrophage action was effective against both intracytosolic and intravacuolar pathogens. These findings suggest that p38 MAPK-mediated autophagy can help stimulate IFN-γ-mediated cell-autonomous immunity, with implications for understanding how IFN-γ-induced autophagy is mobilized within macrophages for inflammation and host defence.
Both type I and type II IFNs up-regulate gene transcription via the JAK–STAT, p38 MAPK, and PI3K signalling pathways, but slight differences exist in the regulation enacted by each type of IFN.7 Studies using a pharmacological inhibitor of p38 MAPK, over-expression of kinase-inactive p38α mutant, or mouse embryonic fibroblasts from p38α knockout mice, showed that p38 MAPK is required for gene transcription via the IFN-stimulated response element and IFN-γ-activated site in response to type I IFNs, such as IFN-α and IFN-β.43,44 However, p38 MAPK plays no role in type II IFN or IFN-γ-dependent gene transcription.44 In our previous study, phosphorylation of p38 MAPK by IFN-γ stimulation was observed after STAT1 knockdown in RAW 264.7 cells and in BMMs from STAT1-knockout mice.38 Collectively, these data demonstrate that STAT1- and p38 MAPK-dependent pathways operate separately from each other in IFN-γ signalling.
The NOS2 is responsible for bactericidal activity in IFN-γ-stimulated macrophages that combat intracellular bacteria via NO generation.22,23 In addition to NOS2, Irgm1 – a member of the IFN-γ-inducible p47 IRG family – activates autophagy, and this contributes to the defence mechanism of macrophages acting against M. tuberculosis.25,26 The IFN-γ-induced macrophage bactericidal activity via NOS2 and Irgm1 depend on STAT1, because this molecule mediates the expression of NOS2 and Irgm1.24,45
In this study, we showed that macrophage bactericidal activity increased at 4 hr following IFN-γ stimulation. This phenomenon was not attenuated in STAT1-KD cells, suggesting that the macrophage bactericidal activity occurring in the early stages following IFN-γ stimulation is not due to NOS2 or Irgm1 (Fig. 2). In fact, the NOS inhibitors l-NMMA and DPI failed to affect this kind of bactericidal activity (Fig. 3). The synergistic effects that are initiated via NOS2-, Irgm1- and p38 MAPK-dependent signalling cascades are likely to contribute to IFN-γ-mediated host defence.
Acknowledgments
We thank Masao Mitsuyama (Kyoto University Graduate School of Medicine, Kyoto, Japan) for the generous gift of L. monocytogenes strain EGD (serovar 1/2a). This work was supported by JSPS KAKENHI grant numbers 22659088 and 24790422.
Disclosure
The authors have no financial conflicts of interest.
References
- 1.Nathan CF. Secretory products of macrophages. J Clin Invest. 1987;79:319–26. doi: 10.1172/JCI112815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Celada A, Nathan C. Macrophage activation revisited. Immunol Today. 1994;15:100–2. doi: 10.1016/0167-5699(94)90150-3. [DOI] [PubMed] [Google Scholar]
- 3.MacMicking JD. Recognizing macrophage activation and host defense. Cell Host Microbe. 2009;5:405–7. doi: 10.1016/j.chom.2009.05.006. [DOI] [PubMed] [Google Scholar]
- 4.Billiau A, Matthys P. Interferon-γ: a historical perspective. Cytokine Growth Factor Rev. 2009;20:97–113. doi: 10.1016/j.cytogfr.2009.02.004. [DOI] [PubMed] [Google Scholar]
- 5.Stark GR, Kerr IM, Williams BR, Silverman RH, Schreiber RD. How cells respond to interferons. Annu Rev Biochem. 1998;67:227–64. doi: 10.1146/annurev.biochem.67.1.227. [DOI] [PubMed] [Google Scholar]
- 6.Darnell JE., Jr STATs and gene regulation. Science. 1997;277:1630–5. doi: 10.1126/science.277.5332.1630. [DOI] [PubMed] [Google Scholar]
- 7.Platanias LC. Mechanisms of type-I- and type-II-interferon-mediated signalling. Nat Rev Immunol. 2005;5:375–86. doi: 10.1038/nri1604. [DOI] [PubMed] [Google Scholar]
- 8.Ivaska J, Bosca L, Parker PJ. PKCε is a permissive link in integrin-dependent IFN-γ signalling that facilitates JAK phosphorylation of STAT1. Nat Cell Biol. 2003;5:363–9. doi: 10.1038/ncb957. [DOI] [PubMed] [Google Scholar]
- 9.Hardy PO, Diallo TO, Matte C, Descoteaux A. Roles of phosphatidylinositol 3-kinase and p38 mitogen-activated protein kinase in the regulation of protein kinase C-α activation in interferon-γ-stimulated macrophages. Immunology. 2009;128:e652–60. doi: 10.1111/j.1365-2567.2009.03055.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Navarro A, Anand-Apte B, Tanabe Y, Feldman G, Larner AC. A PI-3 kinase-dependent, Stat1-independent signaling pathway regulates interferon-stimulated monocyte adhesion. J Leukoc Biol. 2003;73:540–5. doi: 10.1189/jlb.1002508. [DOI] [PubMed] [Google Scholar]
- 11.Ramana CV, Gil MP, Han Y, Ransohoff RM, Schreiber RD, Stark GR. Stat1-independent regulation of gene expression in response to IFN-γ. Proc Natl Acad Sci USA. 2001;98:6674–9. doi: 10.1073/pnas.111164198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sun D, Ding A. MyD88-mediated stabilization of interferon-γ-induced cytokine and chemokine mRNA. Nat Immunol. 2006;7:375–81. doi: 10.1038/ni1308. [DOI] [PubMed] [Google Scholar]
- 13.Valledor AF, Sanchez-Tillo E, Arpa L, Park JM, Caelles C, Lloberas J, Celada A. Selective roles of MAPKs during the macrophage response to IFN-γ. J Immunol. 2008;180:4523–9. doi: 10.4049/jimmunol.180.7.4523. [DOI] [PubMed] [Google Scholar]
- 14.Choudhury GG. A linear signal transduction pathway involving phosphatidylinositol 3-kinase, protein kinase Cε, and MAPK in mesangial cells regulates interferon-γ-induced STAT1α transcriptional activation. J Biol Chem. 2004;279:27399–409. doi: 10.1074/jbc.M403530200. [DOI] [PubMed] [Google Scholar]
- 15.Boehm U, Klamp T, Groot M, Howard JC. Cellular responses to interferon-γ. Annu Rev Immunol. 1997;15:749–95. doi: 10.1146/annurev.immunol.15.1.749. [DOI] [PubMed] [Google Scholar]
- 16.MacMicking JD, Nathan C, Hom G, et al. Altered responses to bacterial infection and endotoxic shock in mice lacking inducible nitric oxide synthase. Cell. 1995;81:641–50. doi: 10.1016/0092-8674(95)90085-3. [DOI] [PubMed] [Google Scholar]
- 17.Shenoy AR, Kim BH, Choi HP, Matsuzawa T, Tiwari S, MacMicking JD. Emerging themes in IFN-γ-induced macrophage immunity by the p47 and p65 GTPase families. Immunobiology. 2007;212:771–84. doi: 10.1016/j.imbio.2007.09.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.MacMicking JD. Interferon-inducible effector mechanisms in cell-autonomous immunity. Nat Rev Immunol. 2012;12:367–82. doi: 10.1038/nri3210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.MacMicking JD. IFN-inducible GTPases and immunity to intracellular pathogens. Trends Immunol. 2004;25:601–9. doi: 10.1016/j.it.2004.08.010. [DOI] [PubMed] [Google Scholar]
- 20.Howard J. The IRG proteins: a function in search of a mechanism. Immunobiology. 2008;213:367–75. doi: 10.1016/j.imbio.2007.11.005. [DOI] [PubMed] [Google Scholar]
- 21.Taylor GA, Feng CG, Sher A. p47 GTPases: regulators of immunity to intracellular pathogens. Nat Rev Immunol. 2004;4:100–9. doi: 10.1038/nri1270. [DOI] [PubMed] [Google Scholar]
- 22.MacMicking J, Xie QW, Nathan C. Nitric oxide and macrophage function. Annu Rev Immunol. 1997;15:323–50. doi: 10.1146/annurev.immunol.15.1.323. [DOI] [PubMed] [Google Scholar]
- 23.MacMicking JD, North RJ, LaCourse R, Mudgett JS, Shah SK, Nathan CF. Identification of nitric oxide synthase as a protective locus against tuberculosis. Proc Natl Acad Sci USA. 1997;94:5243–8. doi: 10.1073/pnas.94.10.5243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.MacMicking JD, Taylor GA, McKinney JD. Immune control of tuberculosis by IFN-γ-inducible LRG-47. Science. 2003;302:654–9. doi: 10.1126/science.1088063. [DOI] [PubMed] [Google Scholar]
- 25.Gutierrez MG, Master SS, Singh SB, Taylor GA, Colombo MI, Deretic V. Autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages. Cell. 2004;119:753–66. doi: 10.1016/j.cell.2004.11.038. [DOI] [PubMed] [Google Scholar]
- 26.Singh SB, Davis AS, Taylor GA, Deretic V. Human IRGM induces autophagy to eliminate intracellular mycobacteria. Science. 2006;313:1438–41. doi: 10.1126/science.1129577. [DOI] [PubMed] [Google Scholar]
- 27.Kim BH, Shenoy AR, Kumar P, Das R, Tiwari S, MacMicking JD. A family of IFN-γ-inducible 65-kD GTPases protects against bacterial infection. Science. 2011;332:717–21. doi: 10.1126/science.1201711. [DOI] [PubMed] [Google Scholar]
- 28.Shenoy AR, Wellington DA, Kumar P, Kassa H, Booth CJ, Cresswell P, MacMicking JD. GBP5 promotes NLRP3 inflammasome assembly and immunity in mammals. Science. 2012;336:481–5. doi: 10.1126/science.1217141. [DOI] [PubMed] [Google Scholar]
- 29.Deretic V, Levine B. Autophagy, immunity, and microbial adaptations. Cell Host Microbe. 2009;5:527–49. doi: 10.1016/j.chom.2009.05.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Xu Y, Jagannath C, Liu XD, Sharafkhaneh A, Kolodziejska KE, Eissa NT. Toll-like receptor 4 is a sensor for autophagy associated with innate immunity. Immunity. 2007;27:135–44. doi: 10.1016/j.immuni.2007.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sanjuan MA, Dillon CP, Tait SW, et al. Toll-like receptor signalling in macrophages links the autophagy pathway to phagocytosis. Nature. 2007;450:1253–7. doi: 10.1038/nature06421. [DOI] [PubMed] [Google Scholar]
- 32.Cooney R, Baker J, Brain O, et al. NOD2 stimulation induces autophagy in dendritic cells influencing bacterial handling and antigen presentation. Nat Med. 2010;16:90–7. doi: 10.1038/nm.2069. [DOI] [PubMed] [Google Scholar]
- 33.Travassos LH, Carneiro LA, Ramjeet M, et al. Nod1 and Nod2 direct autophagy by recruiting ATG16L1 to the plasma membrane at the site of bacterial entry. Nat Immunol. 2010;11:55–62. doi: 10.1038/ni.1823. [DOI] [PubMed] [Google Scholar]
- 34.Levine B, Mizushima N, Virgin HW. Autophagy in immunity and inflammation. Nature. 2011;469:323–35. doi: 10.1038/nature09782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Fujiwara E, Washi Y, Matsuzawa T. Observation of autophagosome maturation in the interferon-γ-primed and lipopolysaccharide-activated macrophages using a tandem fluorescent tagged LC3. J Immunol Methods. 2013;394:100–6. doi: 10.1016/j.jim.2013.05.009. [DOI] [PubMed] [Google Scholar]
- 36.Singh SB, Ornatowski W, Vergne I, et al. Human IRGM regulates autophagy and cell-autonomous immunity functions through mitochondria. Nat Cell Biol. 2010;12:1154–65. doi: 10.1038/ncb2119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Deretic V. Autophagy in immunity and cell-autonomous defense against intracellular microbes. Immunol Rev. 2011;240:92–104. doi: 10.1111/j.1600-065X.2010.00995.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Matsuzawa T, Kim BH, Shenoy AR, Kamitani S, Miyake M, Macmicking JD. IFN-γ elicits macrophage autophagy via the p38 MAPK signaling pathway. J Immunol. 2012;189:813–8. doi: 10.4049/jimmunol.1102041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Robbins RA, Grisham MB. Nitric oxide. Int J Biochem Cell Biol. 1997;29:857–60. doi: 10.1016/s1357-2725(96)00167-7. [DOI] [PubMed] [Google Scholar]
- 40.Nussler AK, Billiar TR, Liu ZZ, Morris SM., Jr Coinduction of nitric oxide synthase and argininosuccinate synthetase in a murine macrophage cell line. Implications for regulation of nitric oxide production. J Biol Chem. 1994;269:1257–61. [PubMed] [Google Scholar]
- 41.Weisz A, Oguchi S, Cicatiello L, Esumi H. Dual mechanism for the control of inducible-type NO synthase gene expression in macrophages during activation by interferon-γ and bacterial lipopolysaccharide. Transcriptional and post-transcriptional regulation. J Biol Chem. 1994;269:8324–33. [PubMed] [Google Scholar]
- 42.Chang YP, Chen CL, Chen SO, et al. Autophagy facilitates an IFN-γ response and signal transduction. Microbes Infect. 2011;13:888–94. doi: 10.1016/j.micinf.2011.05.008. [DOI] [PubMed] [Google Scholar]
- 43.Uddin S, Majchrzak B, Woodson J, et al. Activation of the p38 mitogen-activated protein kinase by type I interferons. J Biol Chem. 1999;274:30127–31. doi: 10.1074/jbc.274.42.30127. [DOI] [PubMed] [Google Scholar]
- 44.Li Y, Sassano A, Majchrzak B, et al. Role of p38α MAP kinase in Type I interferon signaling. J Biol Chem. 2004;279:970–9. doi: 10.1074/jbc.M309927200. [DOI] [PubMed] [Google Scholar]
- 45.Meraz MA, White JM, Sheehan KC, et al. Targeted disruption of the Stat1 gene in mice reveals unexpected physiologic specificity in the JAK-STAT signaling pathway. Cell. 1996;84:431–42. doi: 10.1016/s0092-8674(00)81288-x. [DOI] [PubMed] [Google Scholar]