

Abstract
Cellulose, the most abundant biopolymer synthesized on land, is made of linear chains of ß (1–4) linked D-glucose. As a major structural component of the cell wall, cellulose is important not only for industrial use but also for plant growth and development. Cellulose microfibrils are tethered by other cell wall polysaccharides such as hemicellulose, pectin, and lignin. In higher plants, cellulose is synthesized by plasma membrane-localized rosette cellulose synthase complexes. Despite the recent advances using a combination of molecular genetics, live cell imaging, and spectroscopic tools, many aspects of the cellulose synthesis remain a mystery. In this chapter, we highlight recent research progress towards understanding the mechanism of cellulose synthesis in Arabidopsis.
1. INTRODUCTION
Every cell in higher plants is encased in an extracellular matrix, the plant cell wall. The plant cell wall is composed of a mixture of polysaccharides (Cosgrove, 2005). Based on structural and functional differences, plant cell walls can be roughly characterized into two types: the primary cell wall and the secondary cell wall. The composition and structural organization of both primary and secondary cell walls has been a major driving force for the evolutionary diversification of plants on earth (Popper, 2008). The primary cell wall contains cellulose, hemicellulose and pectin. Primary cell wall synthesis occurs during the growing phase of the cell when the cell wall expands due to the forces of internal turgor pressure that push outward against the plasma membrane and the cell wall (Cosgrove, 1997). The secondary cell wall is deposited in specific cell types upon the cessation of cell growth. The secondary cell wall contains cellulose, hemicellulose, and lignin and it is usually thicker and more rigid than the primary cell wall (Roberts et al., 2000).
Through intra- and inter-chain hydrogen bonding, parallel linear glucan chains are crystalized to form cellulose microfibrils, which give cellulose high axial stiffness (Gillis et al., 1969). In higher plants, 3-nm elementary cellulose microfibrils can aggregate into larger size cellulose microfibrils that range from 5–10 nm in width in primary cell walls to 30–50 nm in secondary cell walls (Davies et al., 2003; Zhang, 2013) (Figure 1). The length of cellulose microfibrils, which can be referred to as the degree of polymerization (DP), varies significantly among organisms. The DP of cellulose microfibrils is estimated to range from hundreds to thousands of glucose units in primary walls, and up to 15,000 glucose units in secondary walls (Brett, 2000; Somerville, 2006). However, virtually nothing is known about how the DP is regulated in plants.
Figure 1.

Cellulose microfibrils and cellulose synthase complexes (CSCs) in Arabidopsis. (A) Atomic force micrograph of cellulose microfibrils from Arabidopsis callus tissue after cyclohexane-1,2-diaminetetraacetic acid (CDTA) extraction. Image size is 2×2 µm. Adapted from Davies LM & Harris PJ (2003). Copyright by Springer. (B) Electron micrograph of cellulose microfibrils in Arabidopsis. Scale bar = 100 nm. Layers of cellulose microfibrils organized as sheets can be seen. Adapted from Williamson et al. (2002). Copyright by Elsevier. (C) Cellulose synthase complexes (CSCs) in the plasma membrane of Arabidopsis. Scale bar =100 nm. Adapted from Williamson et al. (2002). Copyright by Elsevier.
All cellulose-synthesizing organisms including bacteria, algae, tunicates, and higher plants have cellulose synthase proteins, which catalyze the polymerization of glucan chains (Brett, 2000; Saxena et al., 2005). Although the catalytic domains of cellulose synthases are conserved for all cellulose-synthesizing organisms, the drastic differences in both the lifestyle of the organisms and the structure of the cellulose that they produce suggest that the regulatory proteins and the underlying mechanisms for cellulose synthesis may have evolved independently (Roberts et al., 2007; Lei et al., 2012b). An example of diversity is the variation in the organization of cellulose synthesizing complexes, which were originally named terminal complexes (TCs) due to their association with the ends of cellulose microfibrils (Montezinos et al., 1976). TCs in higher plants adopt a rosette shape with six lobes that have rotational symmetry and span across the plasma membrane with a 25 nm diameter in the transmembrane region and a larger diameter (45–50 nm) in the cytoplasmic region of the complex (Giddings et al., 1980; Mueller et al., 1980; Brown et al., 1996; Bowling et al., 2008). The protein composition of the rosette in higher plants is not well understood, but each rosette contains multiple cellulose synthase proteins to accommodate the synthesis of multiple glucan chains (Taylor et al., 2000; Doblin et al., 2002; Desprez et al., 2007; Persson et al., 2007). It has been postulated that cellulose synthase complexes are involved not only in the polymerization of glucan chains, but also in the crystallization process (Somerville, 2006). It remains unclear how multiple glucan chains are positioned within proximity of one another to accommodate crystallization through hydrogen bonding. It has been proposed that protein other than cellulose synthases may be a part of the complex and aid in the crystallization process in Arabidopsis (Molhoj et al., 2002).
Cellulose biosynthesis in higher plants is a tightly regulated process (Brabham et al., 2012; Li et al., 2012b). For example, the amount of cellulose, the ratio of cellulose to other cell wall polymers, the degree of polymerization, the crystalline cellulose core size, and the orientation of cellulose microfibrils are under tight control. The vast regulation of cellulose biosynthesis is a reflection of the variability in cell wall structure and composition that arises from a diversity of cell types across various developmental stages (Keegstra, 2010). This area has been greatly advanced by genetic studies in Arabidopsis and by the recent development of an in vivo cell imaging system to visualize the dynamics of cellulose synthase complexes (CSCs) in living cells. However, at the biochemical level, the field is still at the beginning stage of understanding the enzymatic mechanism of cellulose synthesis. In that regard, it is appropriate to present information from other species, even distant ones, to use as an introduction, comparison or reference to understand cellulose biosynthesis in Arabidopsis.
2. THE STRUCTURE AND PROPERTIES OF CELLULOSE
2.1. General structure of cellulose
Cellulose, in its simplest form, consists of β-1,4 linked glucan chains. Hydrogen bonds form between hydroxyl groups and oxygen atoms both within a single glucose chain and between neighboring chains. Together with van der Waals forces, hydrogen bonding aggregates glucan chains together side-by-side and promotes parallel stacking of cellulose microfibrils into crystalline cellulose (Brett, 2000; Somerville, 2006). The natural form of crystalline cellulose is cellulose I. Cellulose I can be irreversibly converted into cellulose II, a form that is more stable than the cellulose I (Brown, 1999a; Brett, 2000). Consisting of cellulose la and Iβ, both forms are composed of parallel glucan chains (Kuga et al., 1988; Maurer et al., 1992; Koyama et al., 1997). The parallel glucan chains in natural cellulose are compatible with the idea that glucan chains in an elementary cellulose microfibril are made simultaneously (Brown, 1999a; Brown et al., 2000). In higher plants, the ratio of cellulose la to Iβ varies among different species and types of walls (Atalla et al., 1984; Sturcova et al., 2004). In both forms, each glucose molecule is rotated 180° in relation to its neighboring glucose molecule, forming a flat ribbon in which cellobiose is the repeating unit (Somerville, 2006).
Multiple glucan chains are simultaneously synthesized by a single plasma membrane-localized CSC and immediately assemble to form an elementary cellulose microfibril (Harris et al., 2010). The size of an elementary fibril is physically determined by the number of individual glucan chains synthesized by a CSC (Tsekos, 1999). In Arabidopsis, CSCs exist as rosettes (Figure 1), which contain six subunits arranged in a hexagonal structure. It has been postulated that each of the six rosette subunits contains six cellulose synthase (CESA) proteins (Doblin et al., 2002). Assuming that each CESA protein within a CSC is enzymatically active, this model would suggest that each CSC synthesizes an elementary microfibril that is comprised of 36 glucan chains (Somerville, 2006). The exact number of individual glucan chains in an elementary cellulose microfibril has not been experimentally determined in Arabidopsis. However, studies on cellulose in celery collenchyma (primary wall) and in spruce wood (secondary wall) suggest that an elementary microfibril is more likely to contain 24 glucan chains (Fernandes et al., 2011; Newman et al., 2013; Thomas et al., 2013). It is not clear whether the number of chains in elementary microfibrils is fixed within an organism let alone across species, but based on the current measurements of elementary cellulose microfibril size, it is reasonable to estimate that each of the six subunits of the CSC contains 4–6 enzymatically active CESAs.
Several methods have been developed to measure the crystallinity of cellulose microfibrils. X-ray diffraction (XRD) can provide a rough estimation of crystallinity through measuring the relative crystallinity index, which is based upon the proportions of crystalline and amorphous cell wall material, a measurement that can be influenced by non-cellulosic polysaccharides (L. Segal, 1959; Harris et al., 2010; Park et al., 2010; Harris et al., 2012). Cellulose crystallinity can also be assessed by 13C solid-state NMR spectroscopy, by comparing the relative intensities of peaks that correspond to C4 atoms in the interior of the cellulose versus C4 atoms that are on the surface of the cellulose microfibril, which can be used to estimate crystallite size (Bootten et al., 2011; Dick-Perez et al., 2011). However, this method has trouble differentiating between amorphous cellulose regions and cellulose chains at the surface of crystalline microfibrils. 13C solid-state NMR spectroscopy is best suited for crystallinity analysis of thick cellulose microfibrils (10–25 nm in diameter) from bacteria and certain algae (Brett, 2000). However, cellulose microfibrils from the primary cell walls in higher plants are relatively thin, ranging from 3–10 nm in diameter (Thomas et al., 2013; Zhang, 2013). A model in which regions of crystalline cellulose are interconnected by amorphous cellulose regions has gained popularity in recent years. These amorphous regions can be detected by small-angle neutron scattering and are highly accessible through acid hydrolysis (Fernandes et al., 2011). However, it is important to note that the isolation of cellulose microfibrils in these studies often involves harsh extraction methods that will affect the native eellulose structure (Somerville, 2006). A sum frequency generation (SFG) spectroscopy was recently used to detect the asymmetric distribution of C6H2 and O3H-O5 group in crystalline cellulose microfibrils (Hieu et al., 2011; Barnette et al., 2012). SFG is a desirable method to estimate the content of crystalline cellulose because there is no spectral interference from other cell wall matrix compounds such as hemicellulose and lignin and it does not require any chemical treatment of biological samples (Barnette et al., 2012). SFG can also detect subtle changes in cellulose ordering and packing in secondary cell wall in Arabidopsis inflorescence stem (Park et al., 2013).
2.2. Cellulose microfibril organization
In general, cellulose microfibrils are laid down transversely to the axis of elongation during primary cell wall synthesis (Figure 1). In Arabidopsis, two experimental systems, the epidermal cells of dark grown hypocotyls and the root elongation zone, have been used to investigate the relationship between cellulose biosynthesis and cell elongation of primary cell walls (Fagard et al., 2000a; Williamson et al., 2002). Rapid longitudinal cell elongation rates coupled with minimal lateral expansion make both systems ideal for genetic, physiological and structural analysis (Crowell et al., 2010a). In epidermal cells of the root elongation zone, newly synthesized cellulose is constantly deposited in a transverse orientation to the root axis (Kerstens et al., 2002). During cell elongation, layers of cellulose microfibrils rotate as sheets with angles that increasingly move towards the longitudinal axis (Anderson et al., 2010). The rotation of cellulose microfibril layers in roots is compatible with the multi-net growth hypothesis in which the growth is associated with passive movement of cell wall layers (Roland et al., 1975). Cellulose microfibrils are also arranged in multilayer sheets of varying angles during hypocotyl elongation. In this cell type, cellulose microfibrils in the outer face of the epidermal cell wall are deposited in parallel with the underlying cortical microtubules that undergo continuous rotation (Chan et al., 2007; Lloyd et al., 2008; Chan et al., 2010). It appears that these two systems have co-evolved to adopt a multi-net cellulose microfibril network. How multi-net cellulose microfibril organization relates to the features of the primary cell walls, especially extensibility and rigidity, are not known. Interestingly, secondary cell walls of wood cells also adopt a similar cellulose microfibril organization. During wood formation, cellulose microfibrils are laid down as three consecutive layers (S1, S2, S3). Within each layer the cellulose microfibrils are highly ordered and parallel, but between each layer the angles of the microfibril sheets are oriented differently (Plomion et al., 2001; Barnett et al., 2004). Along with embedded hemicellulose (xylan) and ligin, the multi-layered cellulose microfibril organization makes the wood cell wall an ideal structure for strength and rigidity (Chaffey, 1999; Plomion et al., 2001). Multilayer cellulose microfibril organization may be a convergent evolutionary product that is best fit for providing rigidity to the cell wall.
In both primary and secondary cell walls, cellulose microfibrils often exist as bundles (Anderson et al., 2010; Fernandes et al., 2011; Thomas et al., 2013; Zhang, 2013). The bundling process likely involves the aggregation of closely arranged cellulose microfibrils. Since the geometry and dimensions of CSCs relate to the size of cellulose microfibrils, it is tempting to speculate that the bundling process might also be influenced by how the CSCs are arranged at the plasma membrane. Since cortical microtubules guide the insertion and movement of CSCs at the plasma membrane (Crowell et al., 2009; Gutierrez et al., 2009), it is possible that as multiple CSCs simultaneously extrude cellulose microfibrils along the same cortical microtubule, these cellulose microfibrils interact with one another to form a larger and bundled cellulose microfibril. Therefore, it is reasonable to speculate that the dynamics of cortical microtubules might influence the bundling process. It is not known if the bundling process occurs simultaneously during individual glucan-chain synthesis or occurs sequentially after the termination of synthesis.
Other cell wall polymers that are in close contact with cellulose microfibrils such as hemicellulose and pectin may also affect the bundling of cellulose microfibrils. Xyloglucan is the most dominant hemicellulose in dicot primary walls and it plays a large role in forming the cross-links between cellulose microfibrils (Keegstra et al., 1973). Cellulose microfibrils are also coated with xylogalacturonan, a pectin polysaccharide, during the synthesis of cellulose microfibrils in quince seed mucilage. The xylogalacturonan coating prevents the coalescence of cellulose microfibrils into large bundles (Ha et al., 1998). In primary cell walls of Arabidopsis, pectin directly interacts with cellulose microfibrils. It has been estimated that 25–50% of the cellulose chains are in close contact with pectin polysaccharides (Wang et al., 2012). While numerous documented studies support the direct interaction between hemicellulose/pectin and cellulose microfibrils (Keegstra et al., 1973; Cosgrove, 1997; Keegstra, 2010; Scheller et al., 2010), it is not known how or to what extent the direct contact between these cell wall polymers affects the bundling process.
3. CELLULOSE SYNTHESIS
3.1. CESA genes, CESA proteins and the Cellulose Synthase Complex (CSC)
In 1996, the first higher plant cellulose synthase gene was cloned from cotton (Pear et al., 1996). Since then, CESA genes have been identified in many higher plants including a few model systems such as Arabidopsis, rice, and poplar (Carroll et al., 2011). CESA proteins of higher plants share relatively low amino acid sequence identity with bacterial CESA proteins (Doblin et al., 2002), in which the CESA was first identified. In bacteria, the CESA gene is in an operon that also contains other genes that are directly related to cellulose synthesis and cellulose microfibril assembly (Ross et al., 1991). However, in higher plants no functionally linked genes are present in the proximity of the CESA genes. CESAs belong to family 2 glycosyltransferases (Richmond et al., 2000; Somerville, 2006). The predicted CESA protein structure suggests that some aspects of CESA structure are conserved between prokaryotes and eukaryotes (Lei et al., 2012b; Morgan et al., 2013; Sethaphong et al., 2013). CESAs are integral plasma membrane proteins with multiple transmembrane domains and a central catalytic domain (Richmond, 2000) (Figure 2). In higher plants, CESAs have eight transmembrane domains: two on the N-terminal side of the central domain and six on the C terminal side (Somerville, 2006). These transmembrane domains are predicted to form a pore in the plasma membrane, which might accommodate the extrusion of newly synthesized glucan chains through the membrane (Morgan et al., 2013; Slabaugh et al., 2013). In higher plants, CESAs contain a cytoplasmic amino terminal zinc finger domain, which is believed to play a role in protein-protein interactions and might be responsible for the dimerization of CESA proteins (Kurek et al., 2002). Following the zinc finger domain is a hyper-variable region that is rich in acidic amino acids. In between the second and third transmembrane domains is the central domain or the globular domain, which extends into the cytoplasm. The central domain contains several signature residues of glycosyltransferases that are conserved in all species (Richmond, 2000; Lei et al., 2012b). Three conserved aspartic acid residues (D1, D2, and D3) are distributed throughout the cytoplasmic domain and a QXXRW motif resides near the C-terminal end of the central domain (Figure 2). These residues are often referred to collectively as the D,D,D,QXXRW motif and are thought to be involved in substrate binding, acceptor binding, and catalysis (Saxena et al., 2001). In higher plants, CESAs have two extra plant-specific protein domains: a plant-conserved region (P-CR) that resides between D1 and D2and a class-specific region (CSR) that resides between D2 and D3 The function of these two plant-specific domains is unclear, but it has been proposed that they might be involved in plant specific regulation of cellulose synthesis (Sethaphong et al., 2013; Slabaugh et al., 2013).
Figure 2.

Domain structure of cellulose synthase (CESA) protein and the positions of CESA mutations.
The direct proof that CESA is a component of the rosette TC came from an immunolabelling experiment in which freeze-fractured replicas from vascular plant Vigna angularis were labeled with CESA antibodies (Kimura et al., 1999). Although the equivalent experiment has not been performed in Arabidopsis, a lesion in Arabidopsis CESA1 (AT4G32410) resulted in a deficiency in cellulose synthesis and caused a disintegration of rosettes, suggesting that CESA1 is a component of the rosette TC in Arabidopsis (Arioli et al., 1998). The Arabidopsis genome encodes ten CESA genes (Figure 2 and Table 1). Based on genetic analysis, CESAs are roughly classified into two groups that correspond to CESAs that are involved in cellulose synthesis in primary cell walls (primary CESAs) and CESAs that are involved in cellulose synthesis in secondary cell walls (secondary CESAs). Among primary CESAs, CESA1 and CESA3 (AT5G05170) are essential while CESA6 (AT5G64740) is redundant with CESA2 (AT4G39350), CESA5 (AT5G09870), and CESA9 (AT2G21770). CESA4 (AT5G44030), CESA7 (AT5G17420), and CESA8 (AT4G18780) are non-redundantly required for cellulose synthesis in secondary cell walls. The functional separation might reflect a need for distinct structural properties of CESA proteins to assemble different CSCs for primary and secondary cell wall synthesis. However, studies also indicate that the involvement of CESA genes in cellulose synthesis of primary and secondary cell walls could be flexible. For example, loss of function of CESA5 or CESA9 resulted in defects of seed coat formation, a process thought to be dependent upon cellulose synthesis in secondary walls (Stork et al., 2010; Harpaz-Saad et al., 2011; Mendu et al., 2011b; Mendu et al., 2011a; Harpaz-Saad et al., 2012). Furthermore, ectopic overexpression of fra5, a dominant mutant of CESA7 affects cell wall formation in both types of walls (Zhong et al., 2003). In a genetic swap experiment, CESA7, under the expression of the CESA3 promoter, can partially complement the growth defect of cesa3 je5, indicating that CESA7 may have structural properties allowing its incorporation into primary CSCs (Carroll et al., 2012). On the other hand, CESA1, under a promoter of a secondary CESA gene, partially complemented cesa8 irx1 null mutant phenotypes, including defects in cellulose content, plant stature and collapsed xylem vessels (Carroll et al., 2012; Li et al., 2013). Interestingly, a phylogenetic analysis revealed that the separation of primary and secondary CESAs may have occurred before the appearance of vascular plants (Carroll et al., 2011).
Table 1.
Genes and mutations implicated in cellulose synthesis
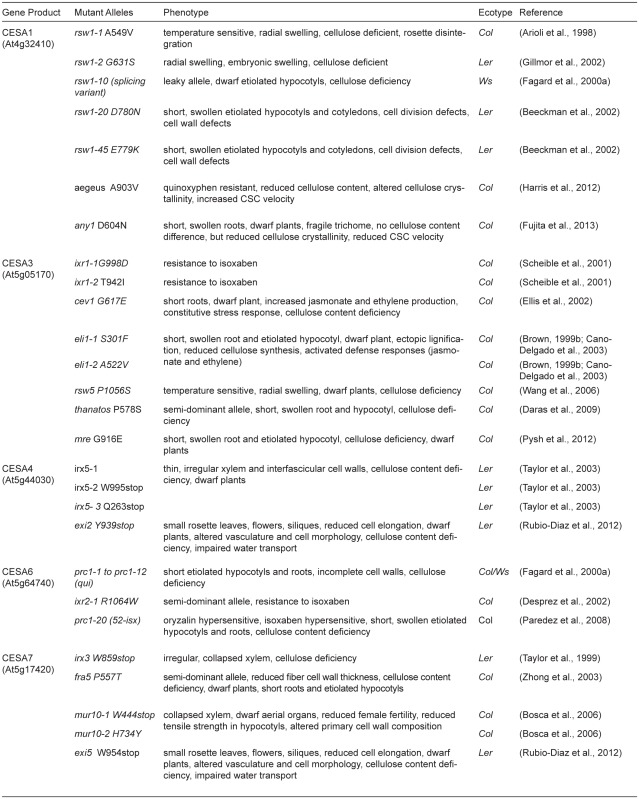
Continued.
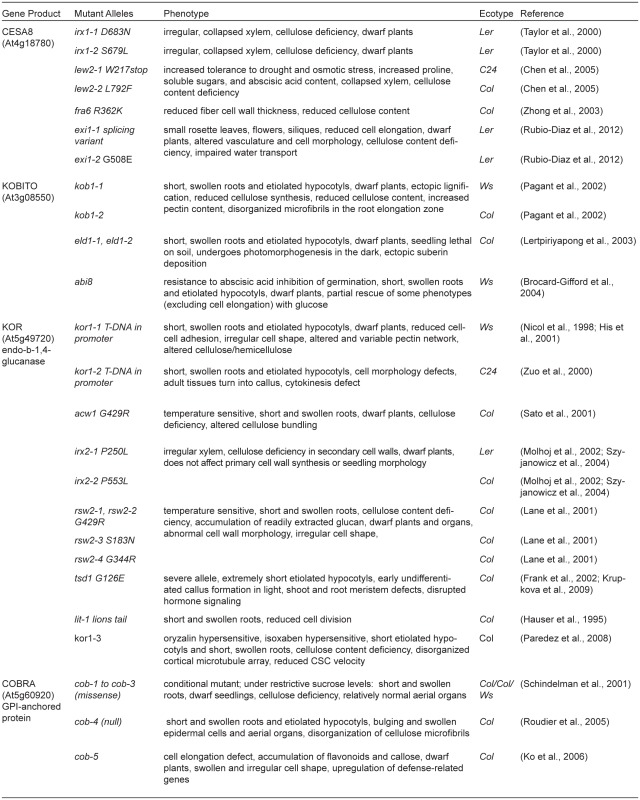
Continued.
3.2. Rosette assembly
Based on the genetic analysis of CESA proteins in Arabidopsis, an early model suggested that each of the six subunits of a rosette contains six CESA proteins, and that each rosette therefore has a total of 36 CESA proteins (Scheible et al., 2001; Doblin et al., 2002). This model explains the formation of a rosette through the distinct binding properties among three different CESA proteins. However, this model has not been experimentally tested. The exact composition and stoichiometry of CESAs in a rosette is unclear. Without such information, the mechanism by which the rosette is assembled from individual CESA proteins remains a mystery. Transmission electron microscopy studies have visualized fully assembled rosettes in the Golgi apparatus (Haigler et al., 1986). This is consistent with the observation that fluorescent protein-labeled CESAs accumulate in the Golgi apparatus, and suggests that rosettes are assembled prior to being delivered to the plasma membrane. Consistent with this idea, rosettes have also been observed in vesicles budding from the Golgi in Micrasterias denticulate (Giddings et al., 1980). Because cellulose synthases are proteins with multiple transmembrane domains, it is reasonable to speculate that rosette assembly could occur at any stage in secretion as early as the endoplasmic reticulum or as late as the Golgi apparatus (Wightman et al., 2010b). In Arabidopsis, genetic studies of both primary and secondary cell walls indicated that at least three different CESAs are required for the formation of an intact CSC (Doblin et al., 2002; Gardiner et al., 2003; Taylor et al., 2003). For secondary cell wall synthesis, deletion of CESA4, CESA7, or CESA8 resulted in a loss of rosette assembly and the remaining CESAs were not trafficked to cell wall deposition sites (Taylor et al., 1999; Taylor et al., 2000; Gardiner et al., 2003; Taylor et al., 2003). Similarly, the temperature sensitive mutant, radial swelling root 1 (rsw1) caused the rosette to disintegrate, revealing the importance of CESA1 in the formation of an intact CSC during the synthesis of primary cell walls (Arioli et al., 1998). Genetic data suggest that primary CSCs are composed of CESA1, CESA3, CESA6 or CESA6-like proteins. Since CESA1 and CESA3 are essential components in the CSC and CESA1 and CESA3 share the highest amino acid similarity, it is reasonable to speculate that CESA1 and/or CESA3 may reside in a special position in the CSC. A chimeric study of CESA1 and CESA3 indicated that these two CESAs have specific positions within the CSC (Wang et al., 2006).
It has been speculated that the dimerization of CESA proteins is important for the formation of the rosette. Attempts to affinity purify an intact CSC using epitope-tagged CESAs led to the isolation of CESA oligomers (Atanassov et al., 2009). The oligomers might be intermediates in the assembly of the complex. The zinc finger domain, the P-CR, and/or CSR domains are only present in rosette forming organisms (Delmer, 1999), therefore, these domains are the obvious candidates for involvement in rosette assembly. An in vitro assay revealed that the zinc-binding domains of GhCESA1 and GhCESA2 were capable of forming homo-dimers and hetero-dimers (Kurek et al., 2002). Interactions between CESAs have been demonstrated by co-immunoprecipitation experiments and yeast two-hybrid assays (Taylor et al., 2003; Desprez et al., 2007; Timmers et al., 2009; Carroll et al., 2012; Li et al., 2013). A recent 3-D model of the central domain of CESA indicates that the P-CR and CSR domains may play a role in rosette assembly (Sethaphong et al., 2013), however, it remains to be determined how the P-CR and/or CSR domains are involved in the interaction (Carroll et al., 2012; Li et al., 2013).
3.3. Functions of the rosette
Formation of crystalline cellulose requires a simultaneous synthesis of multiple glucan chains through the plasma membrane rosettes. The rosettes are thought to have two distinct functions: intracellular polymerization of glucan chains and extracellular assembly of glucan chains into crystalline microfibrils (Arioli et al., 1998). The exact number of glucan chains in an elementary cellulose microfibril remains controversial. The most popular model predicts that 36 glucan chains are synthesized simultaneously by one single rosette (Scheible et al., 2001; Doblin et al., 2002). This prediction implies that 36 enzymatically active CESAs comprise each six-lobed rosette with each lobe being comprised of a hexamer of CESAs. It remains a challenge to express CESAs in vitro and test whether all of the ten CESAs in Arabidopsis are indeed enzymatically active. If some of the CESAs are not enzymatically active but rather acting as a scaffold, it poses an interesting situation in which the spacing of individual glucan chains may vary and may affect the crystallization process.
As of yet, a crystal structure of a CESA protein from higher plants has not been obtained. However, two recent studies have provided significant progress in revealing the structure of cellulose synthases. The first crystal structure of a cellulose synthase has been solved from the bacterium, Rhodobacter spaeroides, and the first predicted tertiary model of the central cytosolic domain of a plant CESA has been revealed (Morgan et al., 2013; Sethaphong et al., 2013). The crystal structure of bacterial CESA, BcsA, supports the involvement of the D,D,D,QXXRW in catalysis and the idea that newly synthesized glucan chains are transported across the plasma membrane through a pore composed of CESA transmembrane domains. The bacterial crystal structure also provides important implications on CESA function in higher plants. In support of this claim, the predicted structure of the central domain of a cotton CESA is superimposable with the crystal structure of the bacterial CESA, BcsA. Consistent with these structural analyses, the Arabidopsis dominant negative mutant, thanatos (Pro578Ser), maps to the invariant QTPH sequence of bacterial BcsA, indicating that the catalytic base function of this amino acid might be conserved (Daras et al., 2009; Morgan et al., 2013). The predicted tertiary structure of the cotton CESA suggests that the P-CR and the CSR domains of cotton CESA fold into distinct subdomains on the periphery of the catalytic region (Morgan et al., 2013; Sethaphong et al., 2013). The putative functions of the P-CR and the CSR may include CSC subunit assembly and interactions with factors involved in the regulation of cellulose synthesis (Sethaphong et al., 2013). In comparing the model of cotton CESA to Arabidopsis sequences, it was revealed that many of the existing Arabidopsis CESA mutation sites are located near the predicted catalytic region. For example, CESA1 E779K (rsw1-45), CESA8 D683N (irx1-1), CESA1 D780N (rsw1-20), and CESA7 D524N reside in the catalytic core (Taylor et al., 2000; Beeckman et al., 2002; Liang et al., 2010; Sethaphong et al., 2013)(Figure 2). While the validation of the model requires additional experiments, the model provides valuable insights into the mechanism of cellulose synthesis in higher plants.
Calcofluor disrupts the crystallization of cellulose in Acetobacter xylinum presumably by interfering with inter-chain hydrogen bonding. This disruption of cellulose microfibril assembly was accompanied by a four-fold increase in the rate of glucose polymerization (Benziman et al., 1980; Haigler et al., 1980). These observations form the basis of a hypothesis that polymerization and crystallization are coupled processes during cellulose biosynthesis. It was proposed that crystallization occurs after the glucan chains have exited the cellulose synthase pores. The structure of BcsA-BcsB from Rhodobacter sphaeroides suggests that glucan synthesis and translocation is a coupled process in which the nascent glucan chain is extended by one glucose molecule at a time (Morgan et al., 2013). If this structure holds true for Acetobacter xylinum, the rate of polymerization will be limited not only by the time required for glucan chains to bundle and crystallize but also by the rate of glucan translocation. Unlike bacteria in which the linear CSCs are stationary with respect to cell surface, rosette CSCs in plants and algae move within the plane of plasma membrane. In Arabidopsis, fluorescently tagged CESAs were observed to move through the plasma membrane at an average speed of 300–350 nm/min. The velocity of CSC movement through the plasma membrane provides a parameter by which the polymerization rate of glucan chains can be measured (Paredez et al., 2006). Recently it was shown that CESA1A903V and CESA3T942I mutants displayed reduced crystalline cellulose content and crystallite size, accompanied by 16% and 8 % increase of velocity of CSCs, respectively (Harris et al., 2012). Although the increase of reaction rate in plants might not be comparable to that of bacteria, these results provide evidence that plants may share a conserved mechanism to couple the polymerization rate with crystallization. Furthermore, overexpression of a cellulose-binding domain (CBD) in poplar resulted an increase in cellulose production, indicating that the crystallization process might also influence the polymerization rate in higher plants (Levy et al., 2002).
Interestingly, the quinoxyphen-resistant mutant, CESA1A903V, aligns with the Tyr455 residue in transmembrane domain 6 of BcsA, which forms a hydrogen bond to the translocating glucan, suggesting that quinoxyphen might affect the translocation of the glucan rather than the synthesis of the glucan (Morgan et al., 2013). It is possible that this mutation affects the configuration of the nascent chain before crystallization, which leads to an affect on the crystallinity of the elementary microfibril and the size of the microfibril. Even if crystallization is a rate-limiting factor for glucan polymerization in higher plants, the difference in magnitude of the influence of crystallization on polymerization rate in bacteria compared to Arabidopsis suggests that regulatory mechanisms of the cellulose synthesis complex differ in the two evolutionarily distant species. Cellulose crystallization in bacteria occurs after glucan chains have been transported out of the cell (Ross et al., 1991), whereas the crystallization of glucan chains in higher plants might be a function of the CSC. If polymerization and crystallization are indeed tightly coupled in cellulose biosynthesis in plants, the time required for glucan chains to bundle and crystallize may limit the rate of polymerization.
3.4. In vitro cellulose synthesis
Attempts to purify enzymatically active CSCs have been unsuccessful. As of yet, in vitro synthesis of cellulose using biochemical approaches has not been established in higher plants. The use of membrane fractions purified from plant tissues has demonstrated limited in vitro cellulose synthesis activities (Okuda et al., 1993; Kudlicka et al., 1995; Kudlicka et al., 1997; Lai-Kee-Him et al., 2002; Ohlsson et al., 2006; Cifuentes et al., 2010). However, rather than cellulose, callose (β-1,3-glucan) appears to be the major product in these limited successful attempts. The disintegration of rosettes in rsw1 mutants indicates that production of crystalline cellulose may require an intact rosette (Arioli et al., 1998). This may explain the difficulties in attempting to purify active complexes for in vitro cellulose synthesis as the purification of intact rosettes is challenging. It has been proposed that structural lipids of the plasma membrane may play an important role in keeping the CSCs intact (Guerriero et al., 2010). The role of lipids in cellulose synthesis in vivo has been supported by the observation that lipids of the sterol family are crucial for cellulose synthesis and cell elongation (Schrick, 2004). It is an attractive idea that the rosette is embedded in a membrane microdomain with a special lipid composition that is required for the integrity of the rosette and the regulation of cellulose synthesis (Guerriero et al., 2010; Schrick et al., 2012). Nonetheless, a comprehensive characterization of all of the components necessary for maintaining the integrity and the activity of the rosette will be required to develop a successful in vitro cellulose synthesis assay in higher plants.
3.5. Genetic studies of CESA genes
Cellulose microfibrils are the major components in both primary and secondary cell walls. Therefore, the content and organization of cellulose microfibrils determine cell wall properties and ultimately control plant development. In Arabidopsis, cellulose deficiency in primary cell walls leads to a reduction in cell expansion (Somerville, 2006; Lei et al., 2012b) (Figure 2 and Table 1). Mutations in either of the two essential CESAs, CESA1 or CESA3, caused abnormal morphogenesis during embryo development (Arioli et al., 1998; Beeckman et al., 2002; Gillmor et al., 2002; Daras et al., 2009). During post-embryonic development, the reduced cell expansion phenotype results in reduced cell elongation and exaggerated radial expansion in epidermal cells of dark grown hypocotyls and in roots of light grown seedlings (Baskin, 2005; Crowell et al., 2010a). For example, the first identified CESA mutation radial swelling root 1 (rswl), a mutation of A589V in Arabidopsis CESA1, showed an enhanced radial expansion defect at the restrictive temperature (Arioli et al., 1998). A null mutant of CESA6 affected the normal cell morphogenesis in both root and etiolated hypocotyls (Desnos et al., 1996; Fagard et al., 2000b). Reverse genetic analyses revealed that the CESA6 like genes, including CESA2, CESA5, and CESA9 are collectively essential for cellulose synthesis (Desprez et al., 2007; Persson et al., 2007). Interestingly, the loss of function mutant of CESA5 or CESA9 also altered seed coat mucilage structure (Stork et al., 2010; Harpaz-Saad et al., 2011; Mendu et al., 2011b; Mendu et al., 2011a; Harpaz-Saad et al., 2012).
Defects in secondary wall cellulose synthesis are represented by collapsed or irregular xylem cells in the vasculature and reduced mechanical strength in fiber cells (Turner et al., 1997). Irregular xylem (irx) mutants such as irx1-1, irx3 and irx5 encode CESA8, CESA7 and CESA4, respectively (Taylor et al., 1999; Taylor et al., 2000; Taylor et al., 2003). Irx1-1 has a mutation in aspartic acid 683 (D683N), which is highly conserved among all plant CESAs. Loss of function of any single secondary CESA causes a complete xylem morphology defect, indicating that each secondary CESA is important for proper function of the whole complex (Taylor et al., 2000). Collapsed xylem vessels in several exigua (exi) mutants, which were mapped to CESA4, CESA7, and CESA8, caused a reduction in water transport capacity and reduced the cell expansion of cell types that do not form secondary cell walls, presumably due to turgor pressure defects (Rubio-Diaz et al., 2012). Fragile fiber 5 (frab), a dominant mutant of CESA7, caused a reduction in cellulose content and thickness of the secondary wall of fiber cells (Zhong et al., 2003). Cellulose synthesis inhibitors such as isoxaben and quinoxyphen have been used in genetic screens for resistant proteins (Brabham et al., 2012). Two CESA mutants, ixr1 and ixr2 conferred resistance to isoxaben (Scheible et al., 2001; Desprez et al., 2002) and a CESA3 mutant was resistant to quinoxyphen (Harris et al., 2012).
Altered cellulose synthesis affects the physical and structural properties of the cell wall. Plant cells can sense the changes in the wall through cell wall integrity sensing mechanisms (Wolf et al., 2012). For example, deficiency in cellulose synthesis in primary cesa mutants is often accompanied by excessive deposition of lignin in non-lignified cells (Cano-Delgado et al., 2003; Hematy et al., 2007). Excessive lignification is also seen in etl1 (At1g05850), a non-CESA mutant with defects in cellulose synthesis (Zhong et al., 2002). Although the molecular mechanism by which a reduction in cellulose biosynthesis triggers the lignification is unknown, it has been speculated that it is part of a response to cell wall damage. Consistent with the cell wall damage hypothesis, lesions in primary CESAs induced accumulation of jasmonic acid and ethylene, two plant hormones that have a major role in plant defense, indicating that cellulose deficiency mimics the effect of physical damage or certain biotic stresses (Ellis et al., 2001; Ellis et al., 2002; Cano-Delgado et al., 2003).
Inhibition of cellulose synthesis also alters plant hormone biosynthesis or signaling. A mutation in CESA3 (repp3) has been associated with altered PIN polarity that is essential for auxin flow in plants (Feraru et al., 2011). Given the importance of auxin trafficking during embryogenesis, it is possible that the defects in embryogenesis seen in CESA1 mutants might, to some degree, result from altered auxin transport (Beeckman et al., 2002). Mutations in all three secondary CESAs confer enhanced resistance to the bacterium, Ralstonia solanacearum, and the necrotrophic fungus, Plectosphaerella cucumerina, presumably through an abscisic acid (ABA) dependent pathway (Chen et al., 2005; Hernandez-Blanco et al., 2007). Consistent with altered ABA synthesis, mutations in CESA8 (lew2) are more tolerant to drought stress and accumulate ABA (Chen et al., 2005).
3.6. Non-CESA genes involved in cellulose synthesis
Through forward genetic screens, many non-CESA encoding genes have also been identified to be involved in cellulose synthesis in primary cell walls (Hauser et al., 1995; Reiter et al., 1997; Nicol et al., 1998; Lane et al., 2001; Lukowitz et al., 2001; Burn et al., 2002; Gillmor et al., 2002; Pagant et al., 2002; Williamson et al., 2002; Scheible et al., 2003; Gillmor et al., 2005) (Table 1). It remains to be determined whether any of the corresponding non-CESA proteins are directly or indirectly associated with CSCs. Nevertheless, these mutants indicate that cellulose synthesis in higher plants is an intricate and regulated process.
Some well-characterized cellulose deficient mutants include COBRA (AT5G60920), COBL4 (AT5G15630), POM-POM1 (AT1G05850), KOBITO1 (AT3G08550), and KORRIGAN1 (AT5G49720). COBRA (COB1) encodes a GPI-anchored extracellular protein (Schindelman et al., 2001). cob1 disrupts cellulose microfibril orientation and COB1 localizes to microtubule-like structures. Therefore, COB1 has been proposed to be involved in the organization of cellulose microfibrils through a microtubule-related function (Roudier et al., 2005). A COB1 paralog (COBL4/irx6) is involved in cellulose synthesis of secondary cell walls (Brown et al., 2005). Mutations in POM-POM1/CTL1, which encode a putative chitinase, cause dwarfism and cellulose deficiency (Hauser et al., 1995; Zhong et al., 2002). CTL1 might affect cellulose synthesis through an interaction with cellulose (Sanchez-Rodriguez et al., 2012). The KOBITO1/ELD1/ABI8 gene was isolated from screens for mutants with cellulose deficiency and dwarfism, and in screens for mutants with ABA insensitivity (Pagant et al., 2002; Lertpiriyapong et al., 2003; Brocard-Gifford et al., 2004). Similar to the cob1 mutant, the cellulose microfibrils of kobito1 were mis-oreinted in epidermal cells of root elongation zone (Pagant et al., 2002).
Lesions in KORRIGAN1 (KOR1) resulted in defects in cellulose synthesis in both primary and secondary cell walls, including defects in cytokinesis and unrestricted cell proliferation, root radial swelling, dwarfism and collapsed xylem (Nicol et al., 1998; Zuo et al., 2000; Lane et al., 2001 ; Sato et al., 2001 ; Szyjanowicz et al., 2004; Paredez etal., 2008). KOR1 encodes a putative membrane bound β-1,4 endoglucanase (Nicol et al., 1998; Rudsander et al., 2008; Liebminger et al., 2013). KOR1 from Brassica napus that was heterologously expressed and purified in Pichia pastoris was able to digest non-crystalline cellulose but not crystalline cellulose or xyloglucan (Molhoj et al., 2001 ). Similar enzymatic activity has been observed in a KOR1 ortholog from poplar (Master et al., 2004). GFP-KOR1 was localized to the Golgi apparatus, endosomal compartments and the plasma membrane in Arabidopsis (Robert et al., 2005; Crowell et al., 2010b). In bacteria and Rhizobium, a cellulase gene is part of the cellulose synthase operon and plays an essential role in cellulose synthesis (Hayashi et al., 2005). KOR1 has several proposed functions including: cleaving of a sitosterol-glucoside, a putative primer for cellulose synthesis initiation (Molhoj et al., 2002; Peng et al., 2002); removing or modifying erroneous glucan chains; and releasing the CSCs from the growing glucan chains at the end of cellulose microfibrils (Nicol et al., 1998; Delmer, 1999). These proposed functions might suggest a direct association exists between KOR1 and CSCs. However, localization studies indicate that KOR1 and CSCs lack a tight association (Szyjanowicz et al., 2004). Immunoprecipitation experiments have also failed to prove a direct interaction between KOR1 and CESAs (Desprez et al., 2007). When the KOR1 ortholog, PttCel9A, was superimposed onto the crystal structure of a homologous bacterial enzyme, it revealed key differences in the active sites (Master et al., 2004). The lack of a few key determinants for substrate binding may explain why the poplar ortholog has such low catalytic activity. The Arabidopsis root-swelling mutant, rsw2, which is a KOR1 allele, has a defect in crystalline cellulose production at restrictive temperatures, a phenotype that is similar to that of rswl (Lane et al., 2001). Together with the Golgi localization of GFP-KOR1, it might be possible that KOR1 is important for maintaining an intact rosette. Overall, KOR1 might be a multi-functional protein in regulating cellulose synthesis.
Similar to cesal and cesa3 mutants, lesions in enzymes involved in N-glycosylation show embryo defects and cellulose deficiency (Lukowitz et al., 2001; Gillmor et al., 2002). Although CESA proteins have not been reported to be N-glycosylated, it is possible that N-glycan modification is required for proteins that are functionally associated with CSCs. For example, Arabidopsis KOR1 is subjected to N-glycosylation at several positions and evidence from in vitro studies with KOR1 and its orthologs from Brassica napus have shown that N-glycan modification of KOR1 is important for its cellulase activity (Molhoj et al., 2001; Liebminger et al., 2013).
Mutants with changes in sterol content also have cellulose content deficiencies (Schrick, 2004). It has been proposed that sterol-glucoside acts as a primer that is required for cellulose synthesis (Peng et al., 2002). In addition to being a primer, sterols may also be involved in the targeting of CSCs to the plasma membrane and in maintaining rosette integrity and activity (Schrick et al., 2012).
4. REGULATION OF CELLULOSE SYNTHESIS
4.1. Regulation at the transcriptional level
CESAs that are essential for primary cell wall synthesis are considered housekeeping genes since every plant cell is surrounded by a primary cell wall. Consistent with the this idea, CESA1, CESA3 and CESA6 transcripts are accumulated at high levels ubiquitously (Hamann et al., 2004). The co-expression of primary CESAs and secondary CESAs are consistent with the hypothesis that CSCs consist of at least three isoforms for cellulose synthesis in primary and secondary cell wall (Brown et al., 2005; Persson et al., 2005). A transgenic line in which the CESA6 promoter drives the expression of CESA2 cDNA complements a cesa6prc mutant phenotype, suggesting that CESA2 is capable of replacing CESA6 in the CSC. Further supporting the transcriptional coregulation of CESAs, the transcription of CESA2 and CESA6 was regulated by ethylene in a similar fashion (Hamann etal., 2004).
Hormones can have various effects on the expression of CESAs. For example, brassinosteroid (BRs) biosynthesis or perception deficiency is accompanied with defects in cellulose synthesis (Schrick et al., 2012). Exogenous application of BR recovers the deficiency of CESA expression in the BR biosynthesis mutatnt, det2-1 (AT2G38050), but not in the BR receptor mutant, bri1-301 (AT4G39400), suggesting that BR signaling is important for transcriptional regulation of cellulose biosynthesis (Xie et al., 2011). Consistent with this hypothesis, BES1 (AT1G19350), a BR-activated transcription factor was shown to interact with upstream elements of many CESA genes (Xie et al., 2011 ).
Transcriptional and genetic analyses have identified a cascade of transcriptional events that are critical for secondary wall synthesis initiation (Zhong et al., 2010). NAC domain transcription factors such as SECONDARY WALL-ASSOCIATED NAC DOMAIN PROTEIN (SND1 ) (AT1G32770) and NAC SECONDARY WALL THICKENING PROMOTING FACTOR (NST1) (AT2G46770) are key regulators of the transcriptional cascade (Zhong et al., 2006; Mitsuda et al., 2007). Double mutants of SND1 and NST1 resulted in a loss of secondary wall formation in the fibers of stems, which included a lack of cellulose (Zhong et al., 2007a). MYB46 (AT5G12870) and MYB83 (AT3G08500) transcription factors are direct targets of NAC regulators (Zhong et al., 2007b; Zhong et al., 2008; McCarthy et al., 2009). Over-expression of MYB46 or MYB83 increased the expression of secondary CESA genes and induced secondary cell wall deposition (Ko et al., 2009; McCarthy et al., 2009). MYB46 can directly bind to promoters of the three secondary CESA genes as demonstrated by in vitro and in vivo assays (Kim etal., 2012b). Two studies identified consensus sequences for MYB binding, named the secondary wall MYB-responsive element (SMRE) or the MYB46-responsive cis-regulatory element (M46RE), which have been shown to be present in the promoter regions of many secondary cell wall related genes (Zhong et al., 2012) (Kim et al., 2012a). The presence of multiple copies of M46RE in the three secondary CESA genes suggests that there is a direct transcriptional regulation of CESAs through MYB46. Genetic complementation experiments confirmed that M46REs are critical for cellulose synthesis in secondary cell walls (Kim et al., 2013).
4.2. Regulation at the post-transcriptional level
The regulation of CESA at the post-transcriptional level may include the regulation of CSC abundance, the lifetime of the CSC at the plasma membrane, and the post-transcriptional modification of the CSC. The abundance and lifetime of CSCs in the plasma membrane may affect the amount and structural properties of cellulose microfibrils. Studies suggest that CESA proteins may have a short lifetime in the plasma membrane. For example, it was estimated that the lifetime of cotton CESA was less than 30 minutes (Jacob-Wilk et al., 2006). CESA dimerization via the N-terminal zinc-binding domain is regulated by redox state and may affect the degradability of CESAs. Cotton GhCESA1 forms a dimer under oxidative conditions and is more resistant to degradation than the reduced monomeric form of GhCESA1, suggesting the N-terminal cytosolic portion of higher plant CESA proteins may be important for the stability of CESAs (Kurek et al., 2002). Even though the zinc-binding domain is conserved among all CESAs from higher plants, it remains to be tested whether other CESAs, especially primary CESAs, are regulated in this manner.
Post-transcriptional modification may affect the stability and/or activity of CESAs. The phosphorylation of CESAs has attracted much attention because of the recent development in phosphoproteomic studies and genetic manipulation of CESAs. Phosphoproteomic studies in Arabidopsis and maize discovered that CESA proteins including CESA1, CESA3, and CESA5 are phosphorylated in conserved residues of the hypervariable region at the N-terminus (Nuhse et al., 2004; Facette et al., 2013). The role of phosphorylation of CESA1 was tested by complementing the rsw1 mutant with mutated forms of CESA1 that prevent or mimic phosphorylation. Six phosphorylation sites were mutated either to alanine (A) to prevent phosphorylation or to glutamic acid (E) to mimic phosphorylation. The complementation was assessed by rescue of cell elongation, of cellulose content, and of CSC dynamics (Chen et al., 2010). This study suggested that phosphorylation at T166, S686, and S688 is important for normal cellulose content and proper CSC velocity. Interestingly, a few of the single amino acid CESA1 phosphorylation-mutants caused asymmetric motility of CSCs in rsw1 mutant cells. The asymmetric movement appears to rely on an intact cortical microtubule structure since treatment with oryzalin, a microtubule depolymerizing drug, abolished the asymmetric movement (Chen et al., 2010). The role of CESA5 phosphorylation has also been investigated. CESA5 is presumably redundant with CESA6. While the native form of CESA5 did not rescue cesa6 prc1-1, a phosphorylation-mimicking CESA5 mutant with four serine residue substitutions was able to partially complement cesa6prc1-1(Bischoff et al., 2011). Phosphorylation of CESA7 has been documented in vivo at two serine residues within a non-conserved region. These two serine residues were phosphorylated by plant extracts, resulting in protein degradation via a proteasome dependent pathway (Taylor, 2007). While the molecular mechanism of CESA phosphorylation remains to be characterized, it is clear that phosphorylation of CESAs plays multiple roles in regulating the stability and activity of the CSC as well as the relationship between the CSC and microtubules.
4.3. Localization and trafficking of the CSC
CSCs were observed in various locations including the plasma membrane, Golgi-derived vesicles, and Golgi cisternae via transmission electron microscopy of freeze-fracture replicas (Haigler et al., 1986). Observing the dynamic behavior of CSCs in living cells was not possible until the recent development of live imaging and fluorescent protein tagging of individual CESAs. Fusions between GFP variants and several CESA subunits, namely CESA3, 5, 6, and 7, did not interfere with the protein function as demonstrated by complementation of the respective mutant phenotypes with the fluorescent fusions (Gardiner et al., 2003; Paredez et al., 2006; Desprez et al., 2007; Bischoff et al., 2011 ). Presumably, multiple copies of each fluorescent CESA isoform reside within a single CSC, allowing for the detection of individual or multiple CSCs within each diffraction-limited particle at the plasma membrane. Consistent with earlier freeze fracture studies in which rosettes were detected within the periphery of the trans face of the Golgi, fluorescent protein tagged CESAs also localize to the periphery of the Golgi apparatus (Haigler et al., 1986; Crowell et al., 2009; Gutierrez et al., 2009). Immunogold labeling of a GFP-CESA3 fusion showed GFP-CESA3 localized to the ends of medial and trans-Golgi cisternae (Crowell et al., 2009). The presence of CSC containing vesicles moving beneath the sites of secondary wall deposition has been inferred from fluorescence loss in photobleaching experiments (Wightman et al., 2009). For primary cell wall synthesis, CSC-containing vesicles may reside in small subcellular compartments named small CESA compartments (SmaCCs) or microtubule associated cellulose synthase compartments (MASCs) (Crowell et al., 2009; Gutierrez et al., 2009). Observations of SmaCCs/MASCs during cell expansion and under certain stress treatments indicate that they may function as storage vesicles (Gutierrez et al., 2009). SmaCCs/MASCs accumulate in epidermal cells at the base of hypocotyl where cell expansion has ceased and CSC density is low (Crowell et al., 2009). Osmotic stress and certain drug treatments deplete CSCs from the plasma membrane, which may help to reveal SmaCCs/ MASCs and/or induce SmaCC/MASC formation. The function of SmaCCs/MASCs has been interpreted in two different ways. Either SmaCCs/MASCs function as delivery compartments that appear before the insertion of CSCs into the plasma membrane or SmaCCs/MASCs function as intracellular storage vesicles of internalized CSCs. Upon drug treatment, rapid depletion of CSCs from the plasma membrane corresponds with an apparent accumulation of SmaCCs/MASCs, suggesting that SmaCCs/MASCs might result from the internalization of CSCs (Crowell et al., 2009; Gutierrez et al., 2009; Wightman et al., 2010a). SmaCCs/MASCs have been observed to be associated to cortical microtubules and track depolymerizing microtubule ends, which might be consistent with SmaCC/MASC involvement in assisting the delivery of CSCs along the cortical microtubules (Paredez et al., 2006; Gu et al., 2010b; Gu et al., 2010a; Lei et al., 2012a; Li et al., 2012c). Alternatively, the association of SmaCCs/MASCs with microtubules might be related to the active transport of vesicles via microtubule motors (Gutierrez et al., 2009).
In primary cell wall synthesizing epidermal cells, disruption of actin using Latrunculin B, an actin-destabilizing drug, results in reduced velocity of Golgi movement and aggregation of CSC containing Golgi bodies (Gutierrez et al., 2009). As a result, a much higher density of plasma membrane-localized CSCs accumulates in regions directly above the aggregated Golgi bodies than in regions lacking underlying Golgi bodies (Gutierrez et al., 2009). This suggests that actin is not crucial for the delivery of CSCs from the Golgi to the plasma membrane but that actin is important for the global distribution of CSCs. Although cortical microtubules appear to mark the sites of CSC delivery in primary cell walls, the depletion of microtubules did not affect the delivery rate of CSCs, suggesting that the delivery of CSCs is independent of microtubule function (Gutierrez et al., 2009). In developing xylem cells, patterning of secondary cell wall deposition is characterized by bands of fluorescently labeled CSCs beneath the sites of cellulose deposition (Wightman et al., 2008). In this system, YFP-CESA7 labeled Golgi bodies exhibit rapid movement along thick actin cables and a loss of actin bundles results in the cessation of Golgi movement and the loss of CSCs bands, suggesting that the actin cytoskeleton is required for the delivery of CSCs during secondary cell wall formation (Wightman et al., 2008). It appears that the removal of microtubules does not affect the pausing of Golgi bodies, an event that is presumably associated with the delivery of CSCs at secondary cell wall thickening sites (Wightman et al., 2008). It remains to be addressed whether Goklgi-mediated CSC insertion is responsible for the delivery of de novo CSCs, recycled CSCs or both (Wightman et al., 2010b; Lei et al., 2012b).
Clathrin-mediated endocytosis (CME) is a well-characterized process by which eukaryotes internalize material from the plasma membrane and has recently been implicated in the internalization of CESAs from the plasma membrane (Bashline et al., 2013). In CME, the adaptor protein complex 2 (AP2) acts in recruiting plasma membrane protein cargo and clathrin triskelia of the clathrin coat to the sites of endocytic vesicle formation (Chen et al., 2011). µ2 (At5g46630), also referred to as AP2M, is a subunit of the AP2 complex in Arabidopsis (L. Segal, 1959; Li et al., 2012a; Bashline et al., 2013; Yamaoka et al., 2013). Live cell imaging analysis confirmed that the dynamics of µ2 is associated with CME in Arabidopsis (Bashline et al., 2013). This study also demonstrated that CESA is a cargo protein of CME by displaying a direct interaction between CESAs and µ2 in vitro, by observing the participation of µ2-YFP in the endocytosis of CESA6-mCherry in planta, and by revealing that the abundance of YFP-CESA6 labeled CSCs at the plasma membrane is dependent on µ2. This study suggests that only a small fraction of CSCs are undergoing internalization at any given time (Bashline et al., 2013).
The idea that CSCs are internalized from the plasma membrane through CME was challenged by the sizes of cytoplasmic portion of CSCs compared to the size of CME-derived vesicles (Bowling et al., 2008; Crowell et al., 2009; Wightman et al., 2010a; Bashline et al., 2011). The cytoplasmic portion of the CSC was estimated to be 45–50 nm in width (Bowling et al., 2008) while the diameter of CME-derived vesicles without the clathrin coat was estimated to be an average of 30 nm in Arabidopsis (Dhonukshe et al., 2007; Li et al., 2012a). However, it is quite possible that CSCs may become structurally defective before being internalized and therefore CSCs that are undergoing internalization may contain a reduced number of subunits and have a reduced overall size. Alternatively, the measurements of the cytoplasmic portion of the CSC via TEM may be skewed by the association of accessory or regulatory proteins that bind to the CSC when it is functioning, but that dissociate from the CSC prior to the internalization of the CSC (Bowling et al., 2008).
4.4. Microtubule cytoskeleton and CSC
The microtubule cytoskeleton is essential for controlling cell morphogenesis in higher plants (Fisher et al., 1998; Burk et al., 2001; Burk et al., 2002; Paradez et al., 2006; Hamant et al., 2008; Boudaoud et al., 2009). The anisotropic growth of most post mitotic plant cells is determined by the transverse orientation (perpendicular to the axis of elongation) of cellulose microfibrils. An intimate relationship between microtubules and cellulose microfibrils was postulated at the time of the discovery of the microtubule cytoskeleton. In 1962, Green hypothesized that cytoplasmic elements, later named microtubules, at the cell periphery determined the orientation of newly synthesized cellulose microfibrils, and thereby determined the final shape of plant cells (Green, 1962; Ledbetter et al., 1963). The parallel alignment between the orientation of cellulose microfibrils and the underlying cortical microtubules forms the basis of the “alignment hypothesis” (Ledbetter et al., 1963). Many cytological data support the alignment hypothesis, although the microtubules do not predict the orientation of cellulose microfibrils in some cases (Hepler et al., 1964; Baskin, 2001; Himmelspach et al., 2003; Sugimoto et al., 2003; Wasteneys, 2004; Wasteneys et al., 2004). Two molecular mechanisms have been put forward to explain the alignment hypothesis. In the “direct hypothesis”, CSCs are thought to be attached to cortical microtubules via a direct or indirect interaction, which causes the movement of CSCs and the synthesis of cellulose to be guided by microtubules (Heath, 1974). In an alternative model, the “bumper hypothesis,” cellulose synthesis is constrained spatially by the presence of cortical microtubules that define channels within which CSCs can move at the plasma membrane without a physical link between the CSC and the microtubules (Giddings et al., 1991). Recent live imaging provides convincing evidence to support the direct hypothesis. By dual labeling microtubules and CSCs using different fluorescent proteins, CSCs were observed to move along the underlying track provided by the cortical microtubules (Paredez et al., 2006) (Figure 3). The association between CSCs and the cortical microtubules was further demonstrated by the concomitant reorientation of both CSC trajectories and cortical microtubules upon blue light excitation and oryzalin treatment (Paredez et al., 2006). While the live imaging of the intimate association between CSCs and microtubules rules out the bumper model, it does not differentiate between CSCs interacting with microtubules directly or through an indirect mechanism via other microtubule associated proteins (MAPs).
Figure 3.

Cellulose synthase complexes (CSCs) and microtubules. Confocal images of epidermal cells of dark grown Arabidopsis seedlings expressing both YFP-CESA6 and RFP-TUA5. The coalignment of CSC trajactories and microtubules is evident in merged image. Time averaged image (duration 5 min, 5 sec interval) Scale bar = 10 mm.
To identify putative MAPs that facilitate the association between CSCs and microtubules, a yeast two-hybrid screen was performed using the central domain of primary CESAs (Gu et al., 2010b; Gu et al., 2010a). Cellulose synthase interacting protein 1 (CSI1) (AT2G22125) was identified as an interaction partner of the central domain of CESA6. CSI1 was shown to fit the criteria for a CSC-microtubule linker protein in that CSI1 directly interacted with CESAs in a yeast two-hybrid assay and CSI1 directly interacted with microtubules in vitro (Baskin et al., 2012). In addition, fluorescent protein-tagged CSI1 co-localized with CSCs in planta and exhibited bidirectional motility that was identical to that of CSCs, indicating that CSI1 is associated with CSCs in vivo (Bringmann et al., 2012; Li et al., 2012c). Also supporting the role of CSI1 as a linker protein, csi1 null mutants disrupted the association between CSCs and cortical microtubules in vivo (Lei et al., 2012a, b; Li et al., 2012c).
In addition to mediating the interaction between CSCs and microtubules, CSI1 also is required to maintain the normal velocity of CSCs. Microtubules have been shown to modulate the velocity of CSCs. The velocity of fluorescently labeled CESA5 in cesa6 prc1-1 background increases after the depletion of cortical microtubules upon oryzalin treatment (Bischoff et al., 2011). Prolonged treatment with oryzalin reduced the velocity of GFP-CESA6 labeled CSCs (Li et al., 2012c). It has also been reported a shorter oryzalin treatment at a low concentration did not significantly affect the velocity of CSCs containing GFP-CESA3 or YFP-CESA6, indicating that cortical microtubule structure is not solely responsible for maintaining normal activity of CSCs (DeBolt et al., 2007; Chan et al., 2010). Loss of CSI1 resulted in reduced velocity of CSCs, which is comparable to the loss of microtubules upon prolonged treatment of oryzalin (Brown, 1999b; Gu et al., 2010b; Gu et al., 2010a). It remains to be determined whether CSI1-mediated activity of CSCs is a microtubule dependent process.
The alignment hypothesis might be a simplified model of a more complex association between cortical microtubules and cellulose synthesis. It is more likely that microtubules have additional functions in regulating cellulose biosynthesis. For example, microtubules may regulate exocytosis via direct transport of vesicles containing CSCs and their associated proteins such as KOR1 (Robert et al., 2005; Fujita et al., 2012). Cortical microtubules have been proposed to change the composition of lipids in the plasma membrane, which may influence cellulose synthesis (Fujita et al., 2012). It has also been observed that cortical microtubules compartmentalize endocytosis sites, a process capable of changing the physiological status of the plasma membrane (Kakar et al., 2013). Overall, cortical microtubules most likely act as a platform for regulating many aspects of CSC related functions.
5. CONCLUDING REMARKS AND FUTURE DIRECTIONS
An understanding of the mechanism of cellulose synthesis in higher plants is important not only due to the vast everyday use of products that are made from cellulosic plant material, but also due to the importance of cellulose biosynthesis in plant development. Decoding the mechanism of cellulose synthesis in higher plants should be a pivotal part of understanding plant evolution. Cellulosic biofuel, an environmentally sustainable energy source, has the potential to significantly change the global energy supply and to reduce our dependency on fossil fuels. The future for economically adopting such biofuel is dependent on how well the cellulose biosynthesis process is understood (Somerville, 2007; Carroll et al., 2009; Pauly et al., 2010). Arabidopsis has provided its wealth of genetic capability for understanding this process, however many fundamental questions remain to be addressed. For example, how is the rosette CSC assembled? Is there a primer for cellulose synthesis? How is CSC activated in the plasma membrane? What determines the lifetime of a CSC in the plasma membrane? What is the force/energy to power the movement of CSCs and how does it relate to mutants with defects in cellulose synthesis? How is crystallization controlled considering that the CSC is oriented perpendicular to the orientation of nascent microfibrils? How do cellulose elementary microfibrils form bundles? Many of these questions need innovations in the areas of cell biology, biochemistry, biophysics and computational modeling.
ACKNOWLEDGEMENTS
This work is supported by grant from National Science Foundation (1121375).
Footnotes
Citation: Shundai Li, Logan Bashline, Lei Lei and Ying Gu. (2014) Cellulose Synthesis and Its Regulation. The Arabidopsis Book 11:e0169. doi:10.1199/tab.0169
elocation-id: e0169
First published on January 13, 2014: e0169. doi: 10.1199/tab.0169
REFERENCES
- Anderson C.T., Carroll A., Akhmetova L., Somerville C. Real-time imaging of cellulose reorientation during cell wall expansion in Arabidopsis roots. Plant Physiol. 2010;152:787–796. doi: 10.1104/pp.109.150128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arioli T., Peng L., Betzner A.S., Burn J., Wittke W., Herth W., Camilleri C., Hofte H., Plazinski J., Birch R., Cork A., Glover J., Redmond J., Williamson R.E. Molecular analysis of cellulose biosynthesis in Arabidopsis. Science. 1998;279:717–720. doi: 10.1126/science.279.5351.717. [DOI] [PubMed] [Google Scholar]
- Atalla R.H., Vanderhart D.L. Native Cellulose - a Composite of 2 Distinct Crystalline Forms. Science. 1984;223:283–285. doi: 10.1126/science.223.4633.283. [DOI] [PubMed] [Google Scholar]
- Atanassov II, Pittman. J.K. Turner S.R. Elucidating the mechanisms of assembly and subunit interaction of the cellulose synthase complex of Arabidopsis secondary cell walls. J. Biol. Chem. 2009;284:3833–3841. doi: 10.1074/jbc.M807456200. [DOI] [PubMed] [Google Scholar]
- Barnett J.R., Bonham V.A. Cellulose microfibril angle in the cell wall of wood fibres. Biol. Rev. Cam. Philo. Soc. 2004;79:461–472. doi: 10.1017/s1464793103006377. [DOI] [PubMed] [Google Scholar]
- Barnette A.L., Lee C., Bradley L.C., Schreiner E.P., Park Y.B., Shin H., Cosgrove D.J., Park S., Kim S.H. Quantification of crystalline cellulose in lignocellulosic biomass using sum frequency generation (SFG) vibration spectroscopy and comparison with other analytical methods. Carbo. Polymers. 2012;89:802–809. doi: 10.1016/j.carbpol.2012.04.014. [DOI] [PubMed] [Google Scholar]
- Bashline L., DU J., Gu Y. The trafficking and behavior of cellulose synthase and a glimpse of potential cellulose synthesis regulators. Front. Biol. 2011;6:377–383. [Google Scholar]
- Bashline L., Li S., Anderson C.T., Lei L., Gu Y. The endocytosis of cellulose synthase in Arabidopsis is dependent on mu2, a clathrin mediated endocytosis adaptin. Plant Physiol; 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baskin T.I. On the alignment of cellulose microfibrils by cortical microtubules: a review and a model. Protoplasma. 2001;215:150–171. doi: 10.1007/BF01280311. [DOI] [PubMed] [Google Scholar]
- Baskin T.I. Anisotropic expansion of the plant cell wall. Annu. Rev. Cell Dev. Biol. 2005;21:203–222. doi: 10.1146/annurev.cellbio.20.082503.103053. [DOI] [PubMed] [Google Scholar]
- Baskin T.I., Gu Y. Making parallel lines meet: transferring information from microtubules to extracellular matrix. Cell Adhesion Migration. 2012;6:404–408. doi: 10.4161/cam.21121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beeckman T., Przemeck G.K., Stamatiou G., Lau R., Terryn N., De Rycke R., Inze D., Berleth T. Genetic complexity of cellulose synthase a gene function in Arabidopsis embryogenesis. Plant Physiol. 2002;130:1883–1893. doi: 10.1104/pp.102.010603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benziman M., Haigler C.H., Brown R.M., White A.R., Cooper K.M. Cellulose biogenesis: Polymerization and crystallization are coupled processes in Acetobacter xylinum. Proc. Natl. Acad. Sci. USA. 1980;77:6678–6682. doi: 10.1073/pnas.77.11.6678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bichet A., Desnos T., Turner S., Grandjean O., Hofte H. BOTERO1 is required for normal orientation of cortical microtubules and anisotropic cell expansion in Arabidopsis. Plant J. 2001;25:137–148. doi: 10.1046/j.1365-313x.2001.00946.x. [DOI] [PubMed] [Google Scholar]
- Bischoff V., Desprez T., Mouille G., Vernhettes S., Gonneau M., Hofte H. Phytochrome Regulation of Cellulose Synthesis in Arabidopsis. Curr. Biol. 2011;21:1822–1827. doi: 10.1016/j.cub.2011.09.026. [DOI] [PubMed] [Google Scholar]
- Bootten T.J., Harris P.J., Melton L.D., Newman R.H. Using Solid-State C-13 NMR Spectroscopy to Study the Molecular Organisation of Primary Plant Cell Walls. Plant Cell Wall: Methods and Protocols. 2011;715:179–196. doi: 10.1007/978-1-61779-008-9_13. [DOI] [PubMed] [Google Scholar]
- Bosca S., Barton C.J., Taylor N.G., Ryden P., Neumetzler L., Pauly M., Roberts K., Seifert G.J. Interactions between MUR10/CesA7-dependent secondary cellulose biosynthesis and primary cell wall structure. Plant Physiol. 2006;142:1353–1363. doi: 10.1104/pp.106.087700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boudaoud A., Corson F., Hamant O., Bohn S., Traas J., Couder Y. Turning a plant tissue into a living cell froth through isotropic growth. Proc. Natl. Acad. Sci. USA. 2009;106:8453–8458. doi: 10.1073/pnas.0812493106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowling A.J., Brown R.M. The cytoplasmic domain of the cellulose-synthesizing complex in vascular plants. Protoplasma. 2008;233:115–127. doi: 10.1007/s00709-008-0302-2. [DOI] [PubMed] [Google Scholar]
- Brabham C., Debolt S. Chemical genetics to examine cellulose biosynthesis. Front. Plant Sci. 2012;3:309. doi: 10.3389/fpls.2012.00309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brett C.T. Cellulose microfibrils in plants: biosynthesis, deposition, and integration into the cell wall. Intl. Rev. Cytol. 2000;199:161–199. doi: 10.1016/s0074-7696(00)99004-1. [DOI] [PubMed] [Google Scholar]
- Bringmann M., Li E., Sampathkumar A., Kocabek T., Hauser M.T., Persson S. POM-POM2/cellulose synthase interacting1 is essential for the functional association of cellulose synthase and microtubules in Arabidopsis. Plant Cell. 2012;24:163–177. doi: 10.1105/tpc.111.093575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brocard-Gifford I., Lynch T.J., Garcia M.E., Malhotra B., Finkelstein R.R. The Arabidopsis thaliana ABSCISIC ACID-INSENSITIVE8 encodes a novel protein mediating abscisic acid and sugar responses essential for growth. Plant Cell. 2004;16:406–421. doi: 10.1105/tpc.018077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown D.M., Zeef L.A., Ellis J., Goodacre R., Turner S.R. Identification of novel genes in Arabidopsis involved in secondary cell wall formation using expression profiling and reverse genetics. Plant Cell. 2005;17:2281–2295. doi: 10.1105/tpc.105.031542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown R.M. Cellulose structure and biosynthesis. Pure and Applied Chemistry. 1999a;71:767–775. [Google Scholar]
- Brown R.M. Cellulose structure and biosynthesis: Where do we stand today?. Abstracts of Papers of the American Chemical Society. 1999b;218:U214–U214. [Google Scholar]
- Brown R.M., Saxena I.M. Cellulose biosynthesis: A model for understanding the assembly of biopolymers. Plant Physiol. Biochem. 2000;38:57–67. [Google Scholar]
- Brown R.M., Saxena I.M., Kudlicka K. Cellulose biosynthesis in higher plants. Trends Plant Sci. 1996;1:149–156. [Google Scholar]
- Burk D.H., Ye Z.H. Alteration of oriented deposition of cellulose microfibrils by mutation of a katanin-like microtubule-severing protein. Plant Cell. 2002;14:2145–2160. doi: 10.1105/tpc.003947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burk D.H., Liu B., Zhong R., Morrison W.H., Ye Z.H. A katanin-like protein regulates normal cell wall biosynthesis and cell elongation. Plant Cell. 2001;13:807–827. [PMC free article] [PubMed] [Google Scholar]
- Burn J.E., Hurley U.A., Birch R.J., Arioli T., Cork A., Williamson R.E. The cellulose-deficient Arabidopsis mutant rsw3 is defective in a gene encoding a putative glucosidase II, an enzyme processing N-glycans during ER quality control. Plant J. 2002;32:949–960. doi: 10.1046/j.1365-313x.2002.01483.x. [DOI] [PubMed] [Google Scholar]
- Cano-Delgado A., Penfield S., Smith C., Catley M., Bevan M. Reduced cellulose synthesis invokes lignification and defense responses in Arabidopsis thaliana. Plant J. 2003;34:351–362. doi: 10.1046/j.1365-313x.2003.01729.x. [DOI] [PubMed] [Google Scholar]
- Carroll A., Somerville C. Cellulosic Biofuels. Annu. Rev. Plant Biol. 2009;60:165–182. doi: 10.1146/annurev.arplant.043008.092125. [DOI] [PubMed] [Google Scholar]
- Carroll A., Specht C.D. Understanding Plant Cellulose Synthases through a Comprehensive Investigation of the Cellulose Synthase Family Sequences. Front. Plant Sci. 2011;2:5. doi: 10.3389/fpls.2011.00005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carroll A., Mansoori N., Li S.D., Lei L., Vernhettes S., Visser R.G.F., Somerville C., Gu Y., Trindade L.M. Complexes with Mixed Primary and Secondary Cellulose Synthases Are Functional in Arabidopsis Plants. Plant Physiol. 2012;160:726–737. doi: 10.1104/pp.112.199208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaffey N. Wood formation in forest trees: from Arabidopsis to Zinnia. Trends Plant Sci. 1999;4:203–204. doi: 10.1016/s1360-1385(99)01417-x. [DOI] [PubMed] [Google Scholar]
- Chan J., Calder G., Fox S., Lloyd C. Cortical microtubule arrays undergo rotary movements in Arabidopsis hypocotyl epidermal cells. Nat. Cell Biol. 2007;9:171–175. doi: 10.1038/ncb1533. [DOI] [PubMed] [Google Scholar]
- Chan J., Crowell E., Eder M., Calder G., Bunnewell S., Findlay K., Vernhettes S., Hofte H., Lloyd C. The rotation of cellulose synthase trajectories is microtubule dependent and influences the texture of epidermal cell walls in Arabidopsis hypocotyls. J. Cell Sci. 2010;123:3490–3495. doi: 10.1242/jcs.074641. [DOI] [PubMed] [Google Scholar]
- Chen S., Ehrhardt D.W., Somerville C.R. Mutations of cellulose synthase (CESA1) phosphorylation sites modulate anisotropic cell expansion and bidirectional mobility of cellulose synthase. Proc Natl Acad Sci U S A. 2010;107:17188–17193. doi: 10.1073/pnas.1012348107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X., Irani N.G., Friml J. Clathrin-mediated endocytosis: the gateway into plant cells. Curr. Opin. Plant Biol. 2011;14:674–682. doi: 10.1016/j.pbi.2011.08.006. [DOI] [PubMed] [Google Scholar]
- Chen Z., Hong X., Zhang H., Wang Y., Li X., Zhu J.K., Gong Z. Disruption of the cellulose synthase gene, AtCesA8/IRX1, enhances drought and osmotic stress tolerance in Arabidopsis. Plant J. 2005;43:273–283. doi: 10.1111/j.1365-313X.2005.02452.x. [DOI] [PubMed] [Google Scholar]
- Cifuentes C., Bulone V., Emons A.M.C. Biosynthesis of Callose and Cellulose by Detergent Extracts of Tobacco Cell Membranes and Quantification of the Polymers Synthesized in vitro. J. Int. Plant Biol. 2010;52:221–233. doi: 10.1111/j.1744-7909.2010.00919.x. [DOI] [PubMed] [Google Scholar]
- Cosgrove D.J. Assembly and enlargement of the primary cell wall in plants. Annu Rev Cell Dev Biol. 1997;13:171–201. doi: 10.1146/annurev.cellbio.13.1.171. [DOI] [PubMed] [Google Scholar]
- Cosgrove D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005;6:850–861. doi: 10.1038/nrm1746. [DOI] [PubMed] [Google Scholar]
- Crowell E.F., Gonneau M., Vernhettes S., Hofte H. Regulation of anisotropic cell expansion in higher plants. Comptes Rendus Biol. 2010a;333:320–324. doi: 10.1016/j.crvi.2010.01.007. [DOI] [PubMed] [Google Scholar]
- Crowell E.F., Gonneau M., Stierhof Y.D., Hofte H., Vernhettes S. Regulated trafficking of cellulose synthases. Curr. Opin. Plant. Biol. 2010b;13:700–705. doi: 10.1016/j.pbi.2010.07.005. [DOI] [PubMed] [Google Scholar]
- Crowell E.F., Bischoff V., Desprez T., Rolland A., Stierhof Y.D., Schumacher K., Gonneau M., Hofte H., Vernhettes S. Pausing of Golgi bodies on microtubules regulates secretion of cellulose synthase complexes in Arabidopsis. Plant Cell. 2009;21:1141–1154. doi: 10.1105/tpc.108.065334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daras G., Rigas S., Penning B., Milioni D., McCann M.C., Carpita N.C., Fasseas C., Hatzopoulos P. The thanatos mutation in Arabidopsis thaliana cellulose synthase 3 (AtCesA3) has a dominant-negative effect on cellulose synthesis and plant growth. New Phytol. 2009;184:114–126. doi: 10.1111/j.1469-8137.2009.02960.x. [DOI] [PubMed] [Google Scholar]
- Davies L.M., Harris P.J. Atomic force microscopy of microfibrils in primary cell walls. Planta. 2003;217:283–289. doi: 10.1007/s00425-003-0979-6. [DOI] [PubMed] [Google Scholar]
- DeBolt S., Gutierrez R., Ehrhardt D.W., Melo C.V., Ross L., Cutler S.R., Somerville C., Bonetta D. Morlin, an inhibitor of cortical microtubule dynamics and cellulose synthase movement. Proc. Natl. Acad. Sci. USA. 2007;104:5854–5859. doi: 10.1073/pnas.0700789104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delmer D.P. CELLULOSE BIOSYNTHESIS: Exciting Times for A Difficult Field of Study. Annu Rev. Plant Physiol. Plant Mol. Biol. 1999;50:245–276. doi: 10.1146/annurev.arplant.50.1.245. [DOI] [PubMed] [Google Scholar]
- Desnos T., Orbovic V., Bellini C., Kronenberger J., Caboche M., Traas J., Hofte H. Procuste1 mutants identify two distinct genetic pathways controlling hypocotyl cell elongation, respectively in dark- and light-grown Arabidopsis seedlings. Development. 1996;122:683–693. doi: 10.1242/dev.122.2.683. [DOI] [PubMed] [Google Scholar]
- Desprez T., Vernhettes S., Fagard M., Refregier G., Desnos T., Aletti E., Py N., Pelletier S., Hofte H. Resistance against herbicide isoxaben and cellulose deficiency caused by distinct mutations in same cellulose synthase isoform CESA6. Plant Physiol. 2002;128:482–490. doi: 10.1104/pp.010822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desprez T., Juraniec M., Crowell E.F., Jouy H., Pochylova Z., Parcy F., Hofte H., Gonneau M., Vernhettes S. Organization of cellulose synthase complexes involved in primary cell wall synthesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA. 2007;104:15572–15577. doi: 10.1073/pnas.0706569104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhonukshe P., Aniento F., Hwang I., Robinson D.G., Mravec J., Stierhof Y.D., Friml J. Clathrin-mediated constitutive endocytosis of PIN auxin efflux carriers in Arabidopsis. Curr. Biol. 2007;17:520–527. doi: 10.1016/j.cub.2007.01.052. [DOI] [PubMed] [Google Scholar]
- Dick-Perez M., Zhang Y., Hayes J., Salazar A., Zabotina O.A., Hong M. Structure and interactions of plant cell-wall polysaccharides by two- and three-dimensional magic-angle-spinning solid-state NMR. Biochemistry. 2011;50:989–1000. doi: 10.1021/bi101795q. [DOI] [PubMed] [Google Scholar]
- Doblin M.S., Kurek I., Jacob-Wilk D., Delmer D.P. Cellulose biosynthesis in plants: from genes to rosettes. Plant Cell Physiol. 2002;43:1407–1420. doi: 10.1093/pcp/pcf164. [DOI] [PubMed] [Google Scholar]
- Ellis C., Turner J.G. The Arabidopsis mutant cev1 has constitutively active jasmonate and ethylene signal pathways and enhanced resistance to pathogens. Plant Cell. 2001;13:1025–1033. doi: 10.1105/tpc.13.5.1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis C., Karafyllidis I., Wasternack C., Turner J.G. The Arabidopsis mutant cev1 links cell wall signaling to jasmonate and ethylene responses. Plant Cell. 2002;14:1557–1566. doi: 10.1105/tpc.002022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Facette M.R., Shen Z., Bjornsdottir F.R., Briggs S.P., Smith L.G. Parallel Proteomic and Phosphoproteomic Analyses of Successive Stages of Maize Leaf Development. Plant Cell; 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fagard M., Hofte H., Vernhettes S. Cell wall mutants. Plant Physiol. Biochem. 2000a;38:15–25. [Google Scholar]
- Fagard M., Desnos T., Desprez T., Goubet F., Refregier G., Mouille G., McCann M., Rayon C., Vernhettes S., Hofte H. PROCUSTE1 encodes a cellulose synthase required for normal cell elongation specifically in roots and dark-grown hypocotyls of Arabidopsis. Plant Cell. 2000b;12:2409–2424. doi: 10.1105/tpc.12.12.2409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feraru E., Feraru M.I., Kleine-Vehn J., Martiniere A., Mouille G., Vanneste S., Vernhettes S., Runions J., Friml J. PIN polarity maintenance by the cell wall in Arabidopsis. Curr. Biol. 2011;21:338–343. doi: 10.1016/j.cub.2011.01.036. [DOI] [PubMed] [Google Scholar]
- Fernandes A.N., Thomas L.H., Altaner C.M., Callow P., Forsyth V.T., Apperley D.C., Kennedy C.J., Jarvis M.C. Nanostructure of cellulose microfibrils in spruce wood. Proc. Natl. Acad. Sci. USA. 2011;108:E1195–E1203. doi: 10.1073/pnas.1108942108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher D.D., Cyr R.J. Extending the Microtubule/Microfibril paradigm. Cellulose synthesis is required for normal cortical microtubule alignment in elongating cells. Plant Physiol. 1998;116:1043–1051. doi: 10.1104/pp.116.3.1043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank M., Guivarc'h A., Krupkova E., Lorenz-Meyer I., Chriqui D., Schmulling T. Tumorous shoot development (TSD) genes are required for co-ordinated plant shoot development. Plant J. 2002;29:73–85. doi: 10.1046/j.1365-313x.2002.01197.x. [DOI] [PubMed] [Google Scholar]
- Fujita M., Lechner B., Barton D.A., Overall R.L., Wasteneys G.O. The missing link: do cortical microtubules define plasma membrane nanodomains that modulate cellulose biosynthesis? Protoplasma 249 Suppl. 2012;1:S59–67. doi: 10.1007/s00709-011-0332-z. [DOI] [PubMed] [Google Scholar]
- Fujita M., Himmelspach R., Ward J., Whittington A., Hasenbein N., Liu C., Truong T.T., Galway M.E., Mansfield S.D., Hocart C.H., Wasteneys G.O. The anisotropy1 D604N mutation in the Arabidopsis cellulose synthasel catalytic domain reduces cell wall crystallinity and the velocity of cellulose synthase complexes. Plant Physiol. 2013;162:74–85. doi: 10.1104/pp.112.211565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furumizu C., Komeda Y. A novel mutation in KNOPF uncovers the role of alpha-glucosidase I during post-embryonic development in Arabidopsis thaliana. FEBS Lett. 2008;582:2237–2241. doi: 10.1016/j.febslet.2008.05.019. [DOI] [PubMed] [Google Scholar]
- Gardiner J.C., Taylor N.G., Turner S.R. Control of cellulose synthase complex localization in developing xylem. Plant Cell. 2003;15:1740–1748. doi: 10.1105/tpc.012815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giddings T.H., Staehelin L.A. Microtubule-mediated control of microfibril deposition: a re-examination of the hypothesis. In: Cw L., editor. The cytoskeletal basis of plant growth and form. London: Academic; 1991. pp. 85–99. [Google Scholar]
- Giddings T.H., Jr, Brower D.L., Staehelin L.A. Visualization of particle complexes in the plasma membrane of Micrasterias denticulata associated with the formation of cellulose fibrils in primary and secondary cell walls. J. Cell Biol. 1980;84:327–339. doi: 10.1083/jcb.84.2.327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillis P.P., Mark R.E., Tang R.C. Elastic Stiffness of Crystalline Cellulose in Folded-Chain Solid State. Journal of Materials Science. 1969;4:1003. [Google Scholar]
- Gillmor C.S., Poindexter P., Lorieau J., Palcic M.M., Somerville C. Alpha-glucosidase I is required for cellulose biosynthesis and morphogenesis in Arabidopsis. J. Cell Biol. 2002;156:1003–1013. doi: 10.1083/jcb.200111093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillmor C.S., Lukowitz W., Brininstool G., Sedbrook J.C., Hamann T., Poindexter P., Somerville C. Glycosylphosphati-dylinositol-anchored proteins are required for cell wall synthesis and morphogenesis in Arabidopsis. Plant Cell. 2005;17:1128–1140. doi: 10.1105/tpc.105.031815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green P.B. Mechanism for Plant Cellular Morphogenesis. Science. 1962;138:1404–1405. doi: 10.1126/science.138.3548.1404. [DOI] [PubMed] [Google Scholar]
- Gu Y., Somerville C. Cellulose synthase interacting protein: a new factor in cellulose synthesis. Plant Signal. Behav. 2010a;5:1571–1574. doi: 10.4161/psb.5.12.13621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu Y., Kaplinsky N., Bringmann M., Cobb A., Carroll A., Sampath-kumar A., Baskin T.I., Persson S., Somerville C.R. Identification of a cellulose synthase-associated protein required for cellulose biosynthesis. Proc. Natl. Acad. Sci. USA. 2010b;107:12866–12871. doi: 10.1073/pnas.1007092107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guerriero G., Fugelstad J., Bulone V. What Do We Really Know about Cellulose Biosynthesis in Higher Plants? J. Integr. Plant Biol. 2010;52:161–175. doi: 10.1111/j.1744-7909.2010.00935.x. [DOI] [PubMed] [Google Scholar]
- Gutierrez R., Lindeboom J.J., Paredez A.R., Emons A.M., Ehrhardt D.W. Arabidopsis cortical microtubules position cellulose synthase delivery to the plasma membrane and interact with cellulose synthase trafficking compartments. Nat. Cell Biol. 2009;11:797–806. doi: 10.1038/ncb1886. [DOI] [PubMed] [Google Scholar]
- Ha M.A., Apperley D.C., Evans B.W., Huxham M., Jardine W.G., Vietor R.J., Reis D., Vian B., Jarvis M.C. Fine structure in cellulose microfibrils: NMR evidence from onion and quince. Plant J. 1998;16:183–190. doi: 10.1046/j.1365-313x.1998.00291.x. [DOI] [PubMed] [Google Scholar]
- Haigler C.H., Brown R.M. Transport of Rosettes from the Golgi-Apparatus to the Plasma-Membrane in Isolated Mesophyll-Cells of Zinnia-Elegans during Differentiation to Tracheary Elements in Suspension-Culture. Protoplasma. 1986;134:111–120. [Google Scholar]
- Haigler C.H., Malcolmbrown R., Benziman M. Calcofluor White St Alters the Invivo Assembly of Cellulose Microfibrils. Science. 1980;210:903–906. doi: 10.1126/science.7434003. [DOI] [PubMed] [Google Scholar]
- Hamann T., Osborne E., Youngs H.L., Misson J., Nussaume L., Somerville C. Global expression analysis of CESA and CSL genes in Arabidopsis. Cellulose. 2004;11:279–286. [Google Scholar]
- Hamant O., Heisler M.G., Jonsson H., Krupinski P., Uyttewaal M., Bokov P., Corson F., Sahlin P., Boudaoud A., Meyerowitz E.M., Couder Y., Traas J. Developmental patterning by mechanical signals in Arabidopsis. Science. 2008;322:1650–1655. doi: 10.1126/science.1165594. [DOI] [PubMed] [Google Scholar]
- Harpaz-Saad S., Western T.L., Kieber J.J. The FEI2-SOS5 pathway and CELLULOSE SYNTHASE 5 are required for cellulose biosynthesis in the Arabidopsis seed coat and affect pectin mucilage structure. Plant Signal. Behav. 2012;7:285–288. doi: 10.4161/psb.18819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harpaz-Saad S., McFarlane H.E., Xu S., Divi U.K., Forward B., Western T.L., Kieber J.J. Cellulose synthesis via the FEI2 RLK/SOS5 pathway and cellulose synthase 5 is required for the structure of seed coat mucilage in Arabidopsis. Plant J. 2011;68:941–953. doi: 10.1111/j.1365-313X.2011.04760.x. [DOI] [PubMed] [Google Scholar]
- Harris D., Bulone V., Ding S.Y., DeBolt S. Tools for cellulose analysis in plant cell walls. Plant Physiol. 2010;153:420–426. doi: 10.1104/pp.110.154203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris D.M., Corbin K., Wang T., Gutierrez R., Bertolo A.L., Petti C., Smilgies D.M., Estevez J.M., Bonetta D., Urbanowicz B.R., Ehrhardt D.W., Somerville C.R., Rose J.K., Hong M., Debolt S. Cellulose microfibril crystallinity is reduced by mutating C-terminal transmembrane region residues CESA1A903V and CESA3T942I of cellulose synthase. Proc. Natl. Acad. Sci. USA. 2012;109:4098–4103. doi: 10.1073/pnas.1200352109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauser M.T., Morikami A., Benfey P.N. Conditional root expansion mutants of Arabidopsis. Development. 1995;121:1237–1252. doi: 10.1242/dev.121.4.1237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi T., Yoshida K., Park Y.W., Konishi T., Baba K. Cellulose metabolism in plants. Intl. Rev. Cytol. 2005;247:1–34. doi: 10.1016/S0074-7696(05)47001-1. [DOI] [PubMed] [Google Scholar]
- Heath I.B. A unified hypothesis for the role of membrane bound enzyme complexes and microtubules in plant cell wall synthesis. J Theor. Biol. 1974;48:445–449. doi: 10.1016/s0022-5193(74)80011-1. [DOI] [PubMed] [Google Scholar]
- Hematy K., Sado P.E., Van Tuinen A., Rochange S., Desnos T., Balzergue S., Pelletier S., Renou J.P., Hofte H. A receptor-like kinase mediates the response of Arabidopsis cells to the inhibition of cellulose synthesis. Curr. Biol. 2007;17:922–931. doi: 10.1016/j.cub.2007.05.018. [DOI] [PubMed] [Google Scholar]
- Hepler P.K., Newcomb E.H. Microtubules and Fibrils in the Cytoplasm of Coleus Cells Undergoing Secondary Wall Deposition. J Cell. Biol. 1964;20:529–532. doi: 10.1083/jcb.20.3.529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hermans C., Porco S., Verbruggen N., Bush D.R. Chitinase-like protein CTL1 plays a role in altering root system architecture in response to multiple environmental conditions. Plant Physiol. 2010;152:904–917. doi: 10.1104/pp.109.149849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernandez-Bianco C., Feng D.X., Hu J., Sanchez-Vallet A., Deslandes L., Llorente F., Berrocal-Lobo M., Keller H., Bariet X., Sanchez-Rodriguez C., Anderson L.K., Somerville S., Marco Y., Molina A. Impairment of cellulose synthases required for Arabidopsis secondary cell wall formation enhances disease resistance. Plant Cell. 2007;19:890–903. doi: 10.1105/tpc.106.048058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hieu H.C., Tuan N.A., Li H.Y., Miyauchi Y., Mizutani G. Sum Frequency Generation Microscopy Study of Cellulose Fibers. Applied Spectro. 2011;65:1254–1259. doi: 10.1366/11-06388. [DOI] [PubMed] [Google Scholar]
- Himmelspach R., Williamson R.E., Wasteneys G.O. Cellulose microfibril alignment recovers from DCB-induced disruption despite microtubule disorganization. Plant J. 2003;36:565–575. doi: 10.1046/j.1365-313x.2003.01906.x. [DOI] [PubMed] [Google Scholar]
- His I., Driouich A., Nicol F., Jauneau A., Hofte H. Altered pectin composition in primary cell walls of korrigan, a dwarf mutant of Arabidopsis deficient in a membrane-bound endo-1,4-beta-glucanase. Planta. 2001;212:348–358. doi: 10.1007/s004250000437. [DOI] [PubMed] [Google Scholar]
- Jacob-Wilk D., Kurek I., Hogan P., Delmer D.P. The cotton fiber zinc-binding domain of cellulose synthase A1 from Gossypium hirsutum displays rapid turnover in vitro and in vivo. Proc. Natl. Acad. Sci. USA. 2006;103:12191–12196. doi: 10.1073/pnas.0605098103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kakar K., Zhang H., Scheres B., Dhonukshe P. CLASP-mediated cortical microtubule organization guides PIN polarization axis. Nature. 2013;495:529–533. doi: 10.1038/nature11980. [DOI] [PubMed] [Google Scholar]
- Keegstra K. Plant Cell Walls. Plant Physiol. 2010;154:483–486. doi: 10.1104/pp.110.161240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keegstra K., Talmadge K.W., Bauer W.D., Albersheim P. The Structure of Plant Cell Walls: III. A Model of the Walls of Suspension-cultured Sycamore Cells Based on the Interconnections of the Macromolecular Components. Plant Physiol. 1973;51:188–197. doi: 10.1104/pp.51.1.188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kerstens S., Verbelen J.P. Cellulose orientation in the outer epidermal wall of angiosperm roots: implications for biosystematics. Annals Bot. 2002;90:669–676. doi: 10.1093/aob/mcf237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim W.C., Ko J.H., Han K.H. Identification of a cis-acting regulatory motif recognized by MYB46, a master transcriptional regulator of secondary wall biosynthesis. Plant Mol. Biol. 2012a;78:489–501. doi: 10.1007/s11103-012-9880-7. [DOI] [PubMed] [Google Scholar]
- Kim W.C., Kim J.Y., Ko J.H., Kim J., Han K.H. Transcription factor MYB46 is an obligate component of the transcriptional regulatory complex for functional expression of secondary wall-associated cellulose synthases in Arabidopsis thaliana. Plant Physiol. 2013;170:1374–1378. doi: 10.1016/j.jplph.2013.04.012. [DOI] [PubMed] [Google Scholar]
- Kim W.C., Ko J.H., Kim J.Y., Kim J.M., Bae H.J., Han K.H. Plant J. 2012b. MYB46 directly regulates the gene expression of secondary wall-associated cellulose synthases in Arabidopsis. [DOI] [PubMed] [Google Scholar]
- Kimura S., Laosinchai W., Itoh T., Cui X., Linder C.R., Brown R.M., Jr Immunogold labeling of rosette terminal cellulose-synthesizing complexes in the vascular plant vigna angularis. Plant Cell. 1999;11:2075–2086. doi: 10.1105/tpc.11.11.2075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ko J.H., Kim W.C., Han K.H. Ectopic expression of MYB46 identifies transcriptional regulatory genes involved in secondary wall biosynthesis in Arabidopsis. Plant J. 2009;60:649–665. doi: 10.1111/j.1365-313X.2009.03989.x. [DOI] [PubMed] [Google Scholar]
- Ko J.H., Kim J.H., Jayanty S.S., Howe G.A., Han K.H. Loss of function of COBRA, a determinant of oriented cell expansion, invokes cellular defence responses in Arabidopsis thaliana. J. Exp. Bot. 2006;57:2923–2936. doi: 10.1093/jxb/erl052. [DOI] [PubMed] [Google Scholar]
- Koyama M., Helbert W., Imai T., Sugiyama J., Henrissat B. Parallel-up structure evidences the molecular directionality during biosynthesis of bacterial cellulose. Proc. Natl. Acad. Sci. USA. 1997;94:9091–9095. doi: 10.1073/pnas.94.17.9091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krupkova E., Schmulling T. Developmental consequences of the tumorous shoot development1 mutation, a novel allele of the cellulose-synthesizing KORRIGAN1 gene. Plant Mol Biol. 2009;71:641–655. doi: 10.1007/s11103-009-9546-2. [DOI] [PubMed] [Google Scholar]
- Kudlicka K., Brown Jr R.M. Cellulose and Callose Biosynthesis in Higher Plants (I. Solubilization and Separation of (1->3)-and (1->4)-[beta]-Glucan Synthase Activities from Mung Bean). Plant Physiol. 1997;115:643–656. doi: 10.1104/pp.115.2.643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kudlicka K., Brown R.M., Jr, Li L., Lee J.H., Shin H., Kuga S. [beta]-Glucan Synthesis in the Cotton Fiber (IV. In Vitro Assembly of the Cellulose I Allomorph). Plant Physiol. 1995;107:111–123. doi: 10.1104/pp.107.1.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuga S., Brown R.M. Silver labeling of the reducing ends of bacterial cellulose. Carbo. Research. 1988;180:345–350. [Google Scholar]
- Kurek I., Kawagoe Y., Jacob-Wilk D., Doblin M., Delmer D. Dimerization of cotton fiber cellulose synthase catalytic subunits occurs via oxidation of the zinc-binding domains. Proc. Natl. Acad. Sci. USA. 2002;99:11109–11114. doi: 10.1073/pnas.162077099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwon Y., Kim S.H., Jung M.S., Kim M.S., Oh J.E., Ju H.W., Kim K.I., Vierling E., Lee H., Hong S.W. Arabidopsis hot2 encodes an endochitinase-like protein that is essential for tolerance to heat, salt and drought stresses. Plant J. 2007;49:184–193. doi: 10.1111/j.1365-313X.2006.02950.x. [DOI] [PubMed] [Google Scholar]
- L. Segal J.J.C., Martin A. E., Conrad C. M. An Empirical Method for Estimating the Degree of Crystallinity of Native Cellulose Using the X-Ray Diffractometer. Text Res. J. 1959;29:786–794. [Google Scholar]
- Lai-Kee-Him J., Chanzy H., Muller M., Putaux J.L., Imai T., Bulone V. In vitro versus in vivo cellulose microfibrils from plant primary wall synthases: structural differences. J. Biol. Chem. 2002;277:36931–36939. doi: 10.1074/jbc.M203530200. [DOI] [PubMed] [Google Scholar]
- Lane D.R., Wiedemeier A., Peng L., Hofte H., Vernhettes S., Desprez T., Hocart C.H., Birch R.J., Baskin T.I., Burn J.E., Arioli T., Betzner A.S., Williamson R.E. Temperature-sensitive alleles of RSW2 link the KORRIGAN endo-1,4-beta-glucanase to cellulose synthesis and cytokinesis in Arabidopsis. Plant Physiol. 2001;126:278–288. doi: 10.1104/pp.126.1.278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ledbetter M.C., Porter K.R. A “Microtubule” in Plant Cell Fine Structure. J. Cell Biol. 1963;19:239–250. doi: 10.1083/jcb.19.1.239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lei L., Li S., Gu Y. Cellulose synthase interactive protein 1 (CSI1) mediates the intimate relationship between cellulose microfibrils and cortical microtubules. Plant Signal. Behav. 2012a;7:714–718. doi: 10.4161/psb.20338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lei L., Li S., Gu Y. Cellulose synthase complexes: composition and regulation. Front. Plant Sci. 2012b;3:75. doi: 10.3389/fpls.2012.00075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lertpiriyapong K., Sung Z.R. The elongation defective1 mutant of Arabidopsis is impaired in the gene encoding a serine-rich secreted protein. Plant Mol. Biol. 2003;53:581–595. doi: 10.1023/B:PLAN.0000019067.05185.d6. [DOI] [PubMed] [Google Scholar]
- Levy I., Shani Z., Shoseyov O. Modification of polysaccharides and plant cell wall by endo-1,4-beta-glucanase and cellulose-binding domains. Biomol. Engineer. 2002;19:17–30. doi: 10.1016/s1389-0344(02)00007-2. [DOI] [PubMed] [Google Scholar]
- Li R., Liu P., Wan Y., Chen T., Wang Q., Mettbach U., Baluska F., Samaj J., Fang X., Lucas W.J., Lin J. A membrane microdomain-associated protein, Arabidopsis Flot1, is involved in a clathrin-independent endocytic pathway and is required for seedling development. Plant Cell. 2012a;24:2105–2122. doi: 10.1105/tpc.112.095695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S., Gu Y. Cellulose biosynthesis in higher plants and the role of the cytoskeleton. In: eLS A.M. Hetherington., editor. Chichester: John Wiley and Sons; 2012b. [Google Scholar]
- Li S., Lei L., Gu Y. Functional analysis of complexes with mixed primary and secondary cellulose synthases. Plant Signal. Behav. 2013:8. doi: 10.4161/psb.23179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S., Lei L., Somerville C., Gu Y. Cellulose synthase interacting protein 1 (CSI1) links microtubules and cellulose synthase complexes. Proc. Natl. Acad. Sci. USA. 2012c;109:185–190. doi: 10.1073/pnas.1118560109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang Y.K., Xie X., Lindsay S.E., Wang Y.B., Masle J., Williamson L., Leyser O., Hetherington A.M. Cell wall composition contributes to the control of transpiration efficiency in Arabidopsis thaliana. Plant J. 2010;64:679–686. doi: 10.1111/j.1365-313X.2010.04362.x. [DOI] [PubMed] [Google Scholar]
- Liebminger E., Grass J., Altmann F., Mach L., Strasser R. Characterizing the Link between Glycosylation State and Enzymatic Activity of the Endo-beta1,4-glucanase KORRIGAN1 from Arabidopsis thaliana. J. Biol. Chem. 2013;288:22270–22280. doi: 10.1074/jbc.M113.475558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin D., Cao L., Zhou Z., Zhu L., Ehrhardt D., Yang Z., Fu Y. Rho GTPase signaling activates microtubule severing to promote microtubule ordering in Arabidopsis. Curr Biol. 2013;23:290–297. doi: 10.1016/j.cub.2013.01.022. [DOI] [PubMed] [Google Scholar]
- Lloyd C., Chan J. The parallel lives of microtubules and cellulose microfibrils. Curr. Opin. Plant Biol. 2008;11:641–646. doi: 10.1016/j.pbi.2008.10.007. [DOI] [PubMed] [Google Scholar]
- Lukowitz W., Nickle T.C., Meinke D.W., Last R.L., Conklin P.L., Somerville C.R. Arabidopsis cytl mutants are deficient in a mannose-1 -phosphate guanylyltransferase and point to a requirement of N-linked glycosylation for cellulose biosynthesis. Proc. Natl. Acad. Sci. USA. 2001;98:2262–2267. doi: 10.1073/pnas.051625798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Master E.R., Rudsander U.J., Zhou W., Henriksson H., Divne C., Denman S., Wilson D.B., Teeri T.T. Recombinant expression and enzymatic characterization of PttCel9A, a KOR homologue from Populus tremula x tremuloides. Biochemistry. 2004;43:10080–10089. doi: 10.1021/bi049453x. [DOI] [PubMed] [Google Scholar]
- Maurer A., Fengel D. Parallel Orientation of the Molecular Chains in Cellulose-I and Cellulose-Ii Deriving from Higher-Plants. Holz Als Roh-Und Werkstoff. 1992;50:493–493. [Google Scholar]
- McCarthy R.L., Zhong R., Ye Z.H. MYB83 is a direct target of SND1 and acts redundantly with MYB46 in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell Physiol. 2009;50:1950–1964. doi: 10.1093/pcp/pcp139. [DOI] [PubMed] [Google Scholar]
- Mendu V., Stork J., Harris D., DeBolt S. Cellulose synthesis in two secondary cell wall processes in a single cell type. Plant Signal. Behav. 2011a;6:1638–1643. doi: 10.4161/psb.6.11.17709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mendu V., Griffiths J.S., Persson S., Stork J., Downie A.B., Voiniciuc C., Haughn G.W., DeBolt S. Subfunctionalization of Cellulose Synthases in Seed Coat Epidermal Cells Mediates Secondary Radial Wall Synthesis and Mucilage Attachment. Plant Physiol. 2011b;157:441–453. doi: 10.1104/pp.111.179069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitsuda N., Iwase A., Yamamoto H., Yoshida M., Seki M., Shinozaki K., Ohme-Takagi M. NAC transcription factors, NST1 and NST3, are key regulators of the formation of secondary walls in woody tissues of Arabidopsis. Plant Cell. 2007;19:270–280. doi: 10.1105/tpc.106.047043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molhoj M., Ulvskov P., Dal Degan F. Characterization of a functional soluble form of a Brassica napus membrane-anchored endo-1,4-beta-glucanase heterologously expressed in Pichia pastoris. Plant Physiol. 2001;127:674–684. doi: 10.1104/pp.010269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molhoj M., Pagant S., Hofte H. Towards understanding the role of membrane-bound endo-beta-1,4-glucanases in cellulose biosynthesis. Plant Cell Physiol. 2002;43:1399–1406. doi: 10.1093/pcp/pcf163. [DOI] [PubMed] [Google Scholar]
- Montezinos D., Brown M., Jr Surface architecture of the plant cell: biogenesis of the cell wall, with special emphasis on the role of the plasma membrane in cellulose biosynthesis. J. Supramol. Struct. 1976;5:277–290. doi: 10.1002/jss.400050303. [DOI] [PubMed] [Google Scholar]
- Morgan J.L., Strumillo J., Zimmer J. Crystallographic snapshot of cellulose synthesis and membrane translocation. Nature. 2013;493:181–186. doi: 10.1038/nature11744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mueller S.C., Brown R.M., Jr Evidence for an intramem-brane component associated with a cellulose microfibril-synthesizing complex in higher plants. J. Cell Biol. 1980;84:315–326. doi: 10.1083/jcb.84.2.315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura M., Hashimoto T. A mutation in the Arabidopsis gamma-tubulin-containing complex causes helical growth and abnormal microtubule branching. J Cell Sci. 2009;122:2208–2217. doi: 10.1242/jcs.044131. [DOI] [PubMed] [Google Scholar]
- Newman R.H., Hill S.J., Harris P.J. Wide-angle x-ray scattering and solid-state nuclear magnetic resonance data combined to test models for cellulose microfibrils in mung bean cell walls. Plant Physiol. 2013;163:1558–1567. doi: 10.1104/pp.113.228262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nickle T.C., Meinke D.W. A cytokinesis-defective mutant of Arabidopsis (cyt1) characterized by embryonic lethality, incomplete cell walls, and excessive callose accumulation. Plant J. 1998;15:321–332. doi: 10.1046/j.1365-313x.1998.00212.x. [DOI] [PubMed] [Google Scholar]
- Nicol F., His I., Jauneau A., Vernhettes S., Canut H., Hofte H. A plasma membrane-bound putative endo-1,4-beta-D-glucanase is required for normal wall assembly and cell elongation in Arabidopsis. EMBO J. 1998;17:5563–5576. doi: 10.1093/emboj/17.19.5563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nuhse T.S., Stensballe A., Jensen O.N., Peck S.C. Phosphoproteomics of the Arabidopsis plasma membrane and a new phosphorylation site database. Plant Cell. 2004;16:2394–2405. doi: 10.1105/tpc.104.023150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohlsson A.B., Djerbi S., Winzell A., Bessueille L., Staldal V., Li X.G., Blomqvist K., Bulone V., Teeri T.T., Berglund T. Cell suspension cultures of Populus tremula x P-tremuloides exhibit a high level of cellulose synthase gene expression that coincides with increased in vitro cellulose synthase activity. Protoplasma. 2006;228:221–229. doi: 10.1007/s00709-006-0156-4. [DOI] [PubMed] [Google Scholar]
- Okuda K., Li L., Kudlicka K., Kuga S., Brown R.M., Jr [beta]-Glucan Synthesis in the Cotton Fiber (I. Identification of [beta]-1,4-and [beta]-1,3-Glucans Synthesized in Vitro). Plant Physiol. 1993;101:1131–1142. doi: 10.1104/pp.101.4.1131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pagant S., Bichet A., Sugimoto K., Lerouxel O., Desprez T., Mc-Cann M., Lerouge P., Vernhettes S., Hofte H. KOBITO1 encodes a novel plasma membrane protein necessary for normal synthesis of cellulose during cell expansion in Arabidopsis. Plant Cell. 2002;14:2001–2013. doi: 10.1105/tpc.002873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paradez A., Wright A., Ehrhardt D.W. Microtubule cortical array organization and plant cell morphogenesis. Curr. Opin. Plant Biol. 2006;9:571–578. doi: 10.1016/j.pbi.2006.09.005. [DOI] [PubMed] [Google Scholar]
- Paredez A.R., Somerville C.R., Ehrhardt D.W. Visualization of cellulose synthase demonstrates functional association with microtubules. Science. 2006;312:1491–1495. doi: 10.1126/science.1126551. [DOI] [PubMed] [Google Scholar]
- Paredez A.R., Persson S., Ehrhardt D.W., Somerville C.R. Genetic evidence that cellulose synthase activity influences microtubule cortical array organization. Plant Physiol. 2008;147:1723–1734. doi: 10.1104/pp.108.120196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park S., Baker J.O., Himmel M.E., Parilla P.A., Johnson D.K. Cellulose crystallinity index: measurement techniques and their impact on interpreting cellulase performance. Biotechnol. Biofuels. 2010;3:10. doi: 10.1186/1754-6834-3-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park Y.B., Lee C.M., Koo B.W., Park S., Cosgrove D.J., Kim S.H. Monitoring meso-scale ordering of cellulose in intact plant cell walls using sum frequency generation spectroscopy. Plant Physiol. 2013;163:907–913. doi: 10.1104/pp.113.225235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pauly M., Keegstra K. Plant cell wall polymers as precursors for biofuels. Curr. Opin. Plant Biol. 2010;13:305–312. doi: 10.1016/j.pbi.2009.12.009. [DOI] [PubMed] [Google Scholar]
- Pear J.R., Kawagoe Y., Schreckengost W.E., Delmer D.P., Stalker D.M. Higher plants contain homologs of the bacterial celA genes encoding the catalytic subunit of cellulose synthase. Proc. Natl. Acad. Sci. USA. 1996;93:12637–12642. doi: 10.1073/pnas.93.22.12637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peng L.C., Kawagoe Y., Hogan P., Delmer D. Sitosterol-beta-glucoside as primer for cellulose synthesis in plants. Science. 2002;295:147–150. doi: 10.1126/science.1064281. [DOI] [PubMed] [Google Scholar]
- Persson S., Wei H., Milne J., Page G.P., Somerville C.R. Identification of genes required for cellulose synthesis by regression analysis of public microarray data sets. Proc. Natl. Acad. Sci. USA. 2005;102:8633–8638. doi: 10.1073/pnas.0503392102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Persson S., Paredez A., Carroll A., Palsdottir H., Doblin M., Poindexter P., Khitrov N., Auer M., Somerville C.R. Genetic evidence for three unique components in primary cell-wall cellulose synthase complexes in Arabidopsis. Proc. Natl. Acad. Sci. USA. 2007;104:15566–15571. doi: 10.1073/pnas.0706592104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plomion C., Leprovost G., Stokes A. Wood formation in trees. Plant Physiol. 2001;127:1513–1523. [PMC free article] [PubMed] [Google Scholar]
- Popper Z.A. Evolution and diversity of green plant cell walls. Curr. Opin. Plant Biol. 2008;11:286–292. doi: 10.1016/j.pbi.2008.02.012. [DOI] [PubMed] [Google Scholar]
- Pysh L., Alexander N., Swatzyna L., Harbert R. Four alleles of AtCESA3 form an allelic series with respect to root phenotype in Arabidopsis thaliana. Physiol. Planta. 2012;144:369–381. doi: 10.1111/j.1399-3054.2012.01575.x. [DOI] [PubMed] [Google Scholar]
- Reiter W.D., Chapple C., Somerville C.R. Mutants of Arabidopsis thaliana with altered cell wall polysaccharide composition. Plant J. 1997;12:335–345. doi: 10.1046/j.1365-313x.1997.12020335.x. [DOI] [PubMed] [Google Scholar]
- Richmond T. Higher plant cellulose synthases. Genome Biol. 2000;1:REVIEWS3001. doi: 10.1186/gb-2000-1-4-reviews3001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richmond T.A., Somerville C.R. The cellulose synthase superfamily. Plant Physiol. 2000;124:495–498. doi: 10.1104/pp.124.2.495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robert S., Bichet A., Grandjean O., Kierzkowski D., Satiat-Jeunemaitre B., Pelletier S., Hauser M.T., Hofte H., Vernhettes S. An Arabidopsis endo-1,4-beta-D-glucanase involved in cellulose synthesis undergoes regulated intracellular cycling. Plant Cell. 2005;17:3378–3389. doi: 10.1105/tpc.105.036228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts A., Roberts E. Evolution of the cellulose synthase (CesA) gene family: insights from green algae and seedless plants. In: Brown R.M., Saxena I.M., editors. Cellulose: Molecular and Structural Biology. Dordrecht: Springer; 2007. pp. 17–34. In. [Google Scholar]
- Roberts K., McCann M.C. Xylogenesis: the birth of a corpse. Curr. Opin. Plant Biol. 2000;3:517–522. doi: 10.1016/s1369-5266(00)00122-9. [DOI] [PubMed] [Google Scholar]
- Rogers L.A., Dubos C., Surman C., Willment J., Cullis I.F., Mansfield S.D., Campbell M.M. Comparison of lignin deposition in three ectopic lignification mutants. New Phytol. 2005;168:123–140. doi: 10.1111/j.1469-8137.2005.01496.x. [DOI] [PubMed] [Google Scholar]
- Roland J.C., Vian B., Reis D. Observations with cytochemistry and ultracryotomy on the fine structure of the expanding walls in actively elongating plant cells. J. Cell Sci. 1975;19:239–259. doi: 10.1242/jcs.19.2.239. [DOI] [PubMed] [Google Scholar]
- Ross P., Mayer R., Benziman M. Cellulose biosynthesis and function in bacteria. Microbiol. Rev. 1991;55:35–58. doi: 10.1128/mr.55.1.35-58.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roudier F., Fernandez A.G., Fujita M., Himmelspach R., Borner G.H., Schindelman G., Song S., Baskin T.I., Dupree P., Wasteneys G.O., Benfey P.N. COBRA, an Arabidopsis extracellular glycosyl-phosphatidyl inositol-anchored protein, specifically controls highly anisotropic expansion through its involvement in cellulose microfibril orientation. Plant Cell. 2005;17:1749–1763. doi: 10.1105/tpc.105.031732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rubio-Diaz S., Perez-Perez J.M., Gonzalez-Bayon R., Munoz-Viana R., Borrega N., Mouille G., Hernandez-Romero D., Robles P., Hofte H., Ponce M.R., Micol J.L. Cell Expansion-Mediated Organ Growth Is Affected by Mutations in Three EXIGUA Genes. PloS one. 2012:7. doi: 10.1371/journal.pone.0036500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rudsander U.J., Sandstrom C., Piens K., Master E.R., Wilson D.B., Brumer Iii H., Kenne L., Teeri T.T. Comparative NMR analysis of cellooligosaccharide hydrolysis by GH9 bacterial and plant endo-1,4-beta-glucanases. Biochemistry. 2008;47:5235–5241. doi: 10.1021/bi702193e. [DOI] [PubMed] [Google Scholar]
- Sanchez-Rodriguez C., Bauer S., Hematy K., Saxe F., Ibanez A.B., Vodermaier V., Konlechner C., Sampathkumar A., Ruggeberg M., Aichinger E., Neumetzier L., Burgert I., Somerville C., Hauser M.T., Persson S. Chitinase-like1/pom-pom1 and its homolog CTL2 are glucan-interacting proteins important for cellulose biosynthesis in Arabidopsis. Plant Cell. 2012;24:589–607. doi: 10.1105/tpc.111.094672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato S., Kato T., Kakegawa K., Ishii T., Liu Y.G., Awano T., Takabe K., Nishiyama Y., Kuga S., Nakamura Y., Tabata S., Shibata D. Role of the putative membrane-bound endo-1,4-beta-glucanase KORRIGAN in cell elongation and cellulose synthesis in Arabidopsis thaliana. Plant Cell Physiol. 2001;42:251–263. doi: 10.1093/pcp/pce045. [DOI] [PubMed] [Google Scholar]
- Saxena I.M., Brown R.M., Jr Cellulose biosynthesis: current views and evolving concepts. Ann. Bot. 2005;96:9–21. doi: 10.1093/aob/mci155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saxena I.M., Brown R.M., Dandekar T. Structure-function characterization of cellulose synthase: relationship to other glycosyl-transferases. Phytochemistry. 2001;57:1135–1148. doi: 10.1016/s0031-9422(01)00048-6. [DOI] [PubMed] [Google Scholar]
- Scheible W.R., Eshed R., Richmond T., Delmer D., Somerville C. Modifications of cellulose synthase confer resistance to isoxaben and thiazolidinone herbicides in Arabidopsis Ixr1 mutants. Proc. Natl. Acad. Sci. USA. 2001;98:10079–10084. doi: 10.1073/pnas.191361598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheible W.R., Fry B., Kochevenko A., Schindelasch D., Zimmerli L., Somerville S., Loria R., Somerville C.R. An Arabidopsis mutant resistant to thaxtomin A, a cellulose synthesis inhibitor from Streptomyces species. Plant Cell. 2003;15:1781–1794. doi: 10.1105/tpc.013342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheller H.V., Ulvskov P. Hemicelluloses. Annu. Rev. Plant Biol. 2010;61:263–289. doi: 10.1146/annurev-arplant-042809-112315. [DOI] [PubMed] [Google Scholar]
- Schindelman G., Morikami A., Jung J., Baskin T.I., Carpita N.C., Derbyshire P., McCann M.C., Benfey P.N. COBRA encodes a putative GPI-anchored protein, which is polarly localized and necessary for oriented cell expansion in Arabidopsis. Genes Dev. 2001;15:1115–1127. doi: 10.1101/gad.879101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schrick K. A link between sterol biosynthesis, the cell-wall, and cellulose in Arabidopsis (vol 38, pg 227, 2004). Plant J. 2004;38:562–562. doi: 10.1111/j.1365-313X.2004.02039.x. [DOI] [PubMed] [Google Scholar]
- Schrick K., Debolt S., Bulone V. Deciphering the molecular functions of sterols in cellulose biosynthesis. Front. Plant Sci. 2012;3:84. doi: 10.3389/fpls.2012.00084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sethaphong L., Haigler C.H., Kubicki J.D., Zimmer J., Bonetta D., DeBolt S., Yingling Y.G. Tertiary model of a plant cellulose synthase. Proc. Natl. Acad. Sci. USA. 2013;110:7512–7517. doi: 10.1073/pnas.1301027110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slabaugh E., Davis J.K., Haigler C.H., Yingling Y.G., Zimmer J. Cellulose synthases: new insights from crystallography and modeling. Trends Plant Sci; 2013. [DOI] [PubMed] [Google Scholar]
- Somerville C. Cellulose synthesis in higher plants. Annu. Rev. Cell Dev. Biol. 2006;22:53–78. doi: 10.1146/annurev.cellbio.22.022206.160206. [DOI] [PubMed] [Google Scholar]
- Somerville C. Biofuels. Curr. Biol. 2007;17:R115–R119. doi: 10.1016/j.cub.2007.01.010. [DOI] [PubMed] [Google Scholar]
- Stork J., Harris D., Griffiths J., Williams B., Beisson F., Li-Beisson Y., Mendu V., Haughn G., DeBolt S. CELLULOSE SYNTHASE9 Serves a Nonredundant Role in Secondary Cell Wall Synthesis in Arabidopsis Epidermal Testa Cells. Plant Physiol. 2010;153:580–589. doi: 10.1104/pp.110.154062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sturcova A., His I., Apperley D.C., Sugiyama J., Jarvis M.C. Structural details of crystalline cellulose from higher plants. Biomacromolecules. 2004;5:1333–1339. doi: 10.1021/bm034517p. [DOI] [PubMed] [Google Scholar]
- Sugimoto K., Himmelspach R., Williamson R.E., Wasteneys G.O. Mutation or drug-dependent microtubule disruption causes radial swelling without altering parallel cellulose microfibril deposition in Arabidopsis root cells. Plant Cell. 2003;15:1414–1429. doi: 10.1105/tpc.011593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szyjanowicz P.M., McKinnon I., Taylor N.G., Gardiner J., Jarvis M.C., Turner S.R. The irregular xylem 2 mutant is an allele of korrigan that affects the secondary cell wall of Arabidopsis thaliana. Plant J. 2004;37:730–740. doi: 10.1111/j.1365-313x.2003.02000.x. [DOI] [PubMed] [Google Scholar]
- Taylor N.G. Identification of cellulose synthase AtCesA7 (IRX3) in vivo phosphorylation sites-a potential role in regulating protein degradation. Plant Mol. Biol. 2007;64:161–171. doi: 10.1007/s11103-007-9142-2. [DOI] [PubMed] [Google Scholar]
- Taylor N.G., Laurie S., Turner S.R. Multiple cellulose synthase catalytic subunits are required for cellulose synthesis in Arabidopsis. Plant Cell. 2000;12:2529–2540. doi: 10.1105/tpc.12.12.2529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor N.G., Scheible W.R., Cutler S., Somerville C.R., Turner S.R. The irregular xylem3 locus of Arabidopsis encodes a cellulose synthase required for secondary cell wall synthesis. Plant Cell. 1999;11:769–780. doi: 10.1105/tpc.11.5.769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor N.G., Howells R.M., Huttly A.K., Vickers K., Turner S.R. Interactions among three distinct CesA proteins essential for cellulose synthesis. Proc. Natl. Acad. Sci. USA. 2003;100:1450–1455. doi: 10.1073/pnas.0337628100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas L.H., Forsyth V.T., Sturcova A., Kennedy C.J., May R.P., Altaner C.M., Apperley D.C., Wess T.J., Jarvis M.C. Structure of cellulose microfibrils in primary cell walls from collenchyma. Plant Physiol. 2013;161:465–476. doi: 10.1104/pp.112.206359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Timmers J., Vernhettes S., Desprez T., Vincken J.P., Visser R.G., Trindade L.M. Interactions between membrane-bound cellulose synthases involved in the synthesis of the secondary cell wall. FEBS letters. 2009;583:978–982. doi: 10.1016/j.febslet.2009.02.035. [DOI] [PubMed] [Google Scholar]
- Tsekos I. The sites of cellulose synthesis in algae: Diversity and evolution of cellulose-synthesizing enzyme complexes. J. Phycol. 1999;35:635–655. [Google Scholar]
- Turner S.R., Somerville C.R. Collapsed xylem phenotype of Arabidopsis identifies mutants deficient in cellulose deposition in the secondary cell wall. Plant Cell. 1997;9:689–701. doi: 10.1105/tpc.9.5.689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J., Howles P.A., Cork A.H., Birch R.J., Williamson R.E. Chimeric proteins suggest that the catalytic and/or C-terminal domains give CesA1 and CesA3 access to their specific sites in the cellulose synthase of primary walls. Plant Physiol. 2006;142:685–695. doi: 10.1104/pp.106.084004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang T., Zabotina O., Hong M. Pectin-cellulose interactions in the Arabidopsis primary cell wall from two-dimensional magic-angle-spinning solid-state nuclear magnetic resonance. Biochemistry. 2012;51:9846–9856. doi: 10.1021/bi3015532. [DOI] [PubMed] [Google Scholar]
- Wasteneys G.O. Progress in understanding the role of microtubules in plant cells. Curr. Opin. Plant Biol. 2004;7:651–660. doi: 10.1016/j.pbi.2004.09.008. [DOI] [PubMed] [Google Scholar]
- Wasteneys G.O., Collings D.A. Expanding beyong the great divide: the cytoskeleton and axial growth. In: Hussey P.J., editor. The plant sytoskeleton in cell differentiation and development. Blackwell; 2004. pp. 83–115. [Google Scholar]
- Webb M., Jouannic S., Foreman J., Linstead P., Dolan L. Cell specification in the Arabidopsis root epidermis requires the activity of ECTOPIC ROOT HAIR 3--a katanin-p60 protein. Development. 2002;129:123–131. doi: 10.1242/dev.129.1.123. [DOI] [PubMed] [Google Scholar]
- Wightman R., Turner S.R. The roles of the cytoskeleton during cellulose deposition at the secondary cell wall. Plant J. 2008;54:794–805. doi: 10.1111/j.1365-313X.2008.03444.x. [DOI] [PubMed] [Google Scholar]
- Wightman R., Turner S. Trafficking of the plant cellulose synthase complex. Plant Physiol. 2010a;153:427–432. doi: 10.1104/pp.110.154666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wightman R., Turner S. Trafficking of the cellulose synthase complex in developing xylem vessels. Biochemical Society Transactions. 2010b;38:755–760. doi: 10.1042/BST0380755. [DOI] [PubMed] [Google Scholar]
- Wightman R., Marshall R., Turner S.R. A cellulose synthase-containing compartment moves rapidly beneath sites of secondary wall synthesis. Plant Cell Physiol. 2009;50:584–594. doi: 10.1093/pcp/pcp017. [DOI] [PubMed] [Google Scholar]
- Williamson R.E., Burn J.E., Hocart C.H. Towards the mechanism of cellulose synthesis. Trends Plant Sci. 2002;7:461–467. doi: 10.1016/s1360-1385(02)02335-x. [DOI] [PubMed] [Google Scholar]
- Wolf S., Hematy K., Hofte H. Growth control and cell wall signaling in plants. Ann. Rev. Plant Biol. 2012;63:381–407. doi: 10.1146/annurev-arplant-042811-105449. [DOI] [PubMed] [Google Scholar]
- Xie L., Yang C., Wang X. Brassinosteroids can regulate cellulose biosynthesis by controlling the expression of CESA genes in Arabidopsis. J. Exp. Bot. 2011;62:4495–4506. doi: 10.1093/jxb/err164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaoka S., Shimono Y., Shirakawa M., Fukao Y., Kawase T., Hatsugai N., Tamura K., Shimada T., Hara-Nishimura I. Identification and dynamics of Arabidopsis adaptor protein-2 complex and its involvement in floral organ development. Plant Cell. 2013;25:2958–2969. doi: 10.1105/tpc.113.114082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang T.M.-L., S. Tittmann B., Cosgrove DJ. Visualization of the nanoscale pattern of recently-deposited cellulose microfibrils and matrix materials in never-dried primary walls of the onion epidermis. 2013. Cellulose DOI 10.1007/s10570-013-9996-1.
- Zhong R., Ye Z.H. MYB46 and MYB83 bind to the SMRE sites and directly activate a suite of transcription factors and secondary wall biosynthetic genes. Plant Cell Physiol. 2012;53:368–380. doi: 10.1093/pcp/pcr185. [DOI] [PubMed] [Google Scholar]
- Zhong R., Demura T., Ye Z.H. SND1, a NAC domain transcription factor, is a key regulator of secondary wall synthesis in fibers of Arabidopsis. Plant Cell. 2006;18:3158–3170. doi: 10.1105/tpc.106.047399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong R., Richardson E.A., Ye Z.H. Two NAC domain transcription factors, SND1 and NST1, function redundantly in regulation of secondary wall synthesis in fibers of Arabidopsis. Planta. 2007a;225:1603–1611. doi: 10.1007/s00425-007-0498-y. [DOI] [PubMed] [Google Scholar]
- Zhong R., Richardson E.A., Ye Z.H. The MYB46 transcription factor is a direct target of SND1 and regulates secondary wall biosynthesis in Arabidopsis. Plant Cell. 2007b;19:2776–2792. doi: 10.1105/tpc.107.053678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong R., Lee C., Ye Z.H. Evolutionary conservation of the transcriptional network regulating secondary cell wall biosynthesis. Trends Plant Sci. 2010;15:625–632. doi: 10.1016/j.tplants.2010.08.007. [DOI] [PubMed] [Google Scholar]
- Zhong R., Kays S.J., Schroeder B.P., Ye Z.H. Mutation of a chitinase-like gene causes ectopic deposition of lignin, aberrant cell shapes, and overproduction of ethylene. Plant Cell. 2002;14:165–179. doi: 10.1105/tpc.010278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong R., Morrison W.H., 3rd, Freshour G.D., Hahn M.G., Ye Z.H. Expression of a mutant form of cellulose synthase AtCesA7 causes dominant negative effect on cellulose biosynthesis. Plant Physiol. 2003;132:786–795. doi: 10.1104/pp.102.019331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong R., Lee C., Zhou J., McCarthy R.L., Ye Z.H. A battery of transcription factors involved in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell. 2008;20:2763–2782. doi: 10.1105/tpc.108.061325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zuo J., Niu Q.W., Nishizawa N., Wu Y., Kost B., Chua N.H. KORRIGAN, an Arabidopsis endo-1,4-beta-glucanase, localizes to the cell plate by polarized targeting and is essential for cytokinesis. Plant Cell. 2000;12:1137–1152. doi: 10.1105/tpc.12.7.1137. [DOI] [PMC free article] [PubMed] [Google Scholar]