

Abstract
Executive control of motor responses is a psychological construct of the executive system. Several studies have demonstrated the involvement of the cerebral cortex, basal ganglia, and thalamus in the inhibition of actions and monitoring of performance. The involvement of the cerebellum in cognitive function and its functional interaction with basal ganglia have recently been reported. Based on these findings, we examined the hypothesis of cerebellar involvement in executive control by administering a countermanding task in patients with focal cerebellar damage. The countermanding task requires one to make a movement in response to a ‘go’ signal and to halt it when a ‘stop’ signal is presented. The duration of the go process (reaction time; RT), the duration of the stop process (stop signal reaction time; SSRT), and their relationship, expressed by a psychometric function, are recorded as measures of executive control. All patients had longer go process duration in general and in particular, as a proactive control, as demonstrated by the increase in RT after erroneously performed stop trials. Further, they were defective in the slope of the psychometric function indicating a difficulty on triggering the stop process, although the SSRT did not differ from controls. Notably, their performance was worse when lesions affected deep cerebellar nuclei. Our results support the hypothesis that the cerebellum regulates the executive control of voluntary actions. We speculate that its activity is attributed to specific cerebellar influence over the cortico-striatal loop.
Introduction
Executive control (EC) allows one to adapt to continuously changing environments by initiating and halting specific actions or monitoring and redirecting the behavior as needed. Improper interactions with the environment, which are associated with several psychiatric and developmental disorders, have been linked to defective EC [1,2]. Several neuroimaging and lesion studies [3,4] have implicated the prefrontal cortex (PFC) as a key node in the cortico-subcortical network that subtends EC with regard to the ability to inhibit actions and monitor performance [5,6].
Of the subcortical structures, the basal ganglia and thalamus have received the most attention [7–10], whereas the cerebellum has been neglected. Recently, using Granger causality and imaging approaches [11], cerebellar output has been proposed to contribute to post-error processing by influencing the activity of the PFC and anterior cingulate cortex (ACC) through connections with the thalamus [12]. Further, the cerebellum could contribute easily to proactive and flexible control of behavior–for example, during post-error adaptation [13,14]–by implementing a forward model to predict the sensory input [15] and influencing the activity of basal ganglia [12], which are implicated in error processing and corrective behavior [16–19].
The contribution of the cerebellum to inhibition is inferred, based on the poor performance of cerebellar patients on the Stroop task [20] and the go/no-go task [21] and reflected by the cerebellar activation that has been observed in imaging studies during the antisaccade task [22] and stop (countermanding) task (CMT) [23]. However, only the behavioral data obtained with the CMT allows one to examine the speed of the stop process and behavioral flexibility following the stop trials and, thus, to determine the function of the cerebellum in motor inhibition and monitoring [24,25]. On the other hand the Stroop task and the go/no-go task, which consider the performance either as the resolution of a conflict between a target and distractor or as the probability of a “nonresponse” following onset of the stimulus, respectively, do not. In support, different mechanisms in the brain for all these tasks have also been implicated by neuroimaging data [5,26].
The aim of this study was to determine the function of the cerebellum in the inhibition of actions and control of performance monitoring by testing patients with focal cerebellar (FCb) damage on a manual CMT. On average, patients reacted to the stop signal as quickly as controls but showed greater variability in triggering the stop process and had longer times in adjusting their behavior after stop failures, particularly when the lesion involved the deep cerebellar nuclei.
Materials and Methods
Participants
Thirteen FCb subjects were enrolled; their aetiologies and lesion characteristics are listed in Table 1. The inclusion criteria were no clinical or neuroradiological evidence of extracerebellar pathologies, forming subgroups of similar numerosity with regard to involvement (FCb+; n.7) or noninvolvement (FCb−; n.6) of deep cerebellar nuclei (DCN) in the lesions (as determined by magnetic resonance imaging [MRI]; see below)–the significance of DCN in cognition has recently been highlighted, because they facilitate cerebellar-thalamo-cortical connections [27,28]. The control group comprised 22 subjects, matched for sex, age, and education, with no history of neurological or psychiatric illness. The demographics and education of the groups are shown in Table 2.
Table 1. Lesion characteristics in the subjects with focal cerebellar lesions.
| Case Code | Side | Aetiology | PICA | AICA | SCA | DCN | ANT | POST | Hem | Vermis | Peduncles |
| FCb1 | R | Surgical | - | - | - | X | X | X | X | X | - |
| FCb2 | L | Ischemic | X | - | - | - | - | X | X | - | - |
| FCb3 | R | Surgical | - | - | - | - | - | X | - | X | - |
| FCb4 | R | Surgical | - | - | - | - | X | X | X | - | - |
| FCb5 | L | Surgical | - | - | - | X | X | X | X | X | - |
| FCb6 | R | Ischemic | X | - | - | X | - | X | X | X | - |
| FCb7 | L | Ischemic | X | - | X | - | X | X | X | X | - |
| FCb8 | R | Ischemic | - | - | - | - | - | - | - | - | X |
| FCb9 | R | Ischemic | X | X | - | X | - | X | X | X | - |
| FCb10 | L | Ischemic | X | - | - | X | X | X | X | X | - |
| FCb11 | L | Ischemic | - | - | X | X | X | X | X | - | - |
| FCb12 | R | Surgical | - | - | - | - | - | X | X | - | - |
| FCb13 | R | Ischemic | - | - | X | X | X | X | - | X | - |
Abbreviations: AICA = anteroinferior cerebellar artery; ANT = anterior cerebellar lobe; DCN = deep cerebellar nuclei; Hem = cerebellar hemisphere; L = Left, PICA = posteroinferior cerebellar artery; POST = posterior cerebellar lobe; R = right; SCA = superior cerebellar artery.
Table 2. Demographic and intellectual data of cerebellar patients and controls. Means (standard deviation).
| Groups | Groups Code | N° | M/F | Age | Education (years) | PM (cut off 18,9) |
| Control subjects | C | 22 | 10/12 | 49.1 (11.4) | 12.6 (4.7) | 31.3 (2.4) |
| Focal cerebellar lesions | FCb | 13 | 7/6 | 51.4 (12.5) | 13.0 (4.0) | 27.6 (3.5) |
| Lesions involving deep cerebellar nuclei | FCb+ | 7 | 6/1 | 50.8 (12.8) | 11.5 (4.7) | 29.2 (3.5) |
| Lesions not involving deep cerebellar nuclei | FCb− | 6 | 1/5 | 52.1 (13.2) | 14.6 (2.5) | 25.6 (2.3) |
| Right side focal cerebellar lesions | FCb_R | 8 | 3/5 | 47.2 (12.7) | 14.2 (3.5) | 26.9 (3.1) |
| Left side focal cerebellar lesions | FCb_L | 5 | 4/1 | 58.2 (9.6) | 11.0 (4.4) | 28.6 (4.2) |
PM: Raven's 47 (progressive matrices); t-test for independent samples confirmed that FCb patients and controls were well matched for age (p = 0.93) and education (p = 0.73).
Patients' cognitive profiles were evaluated extensively using a battery of tests (Table 3), including WAISr total intelligence quotient values (TIQ), immediate (IR) and delayed (DR) recall of Rey's 15 words [29], immediate visual memory (IVM) [30], forward (FDS) and backward digit span (BDS), forward (FC) and backward (BC) Corsi Test [31], Raven's 47 progressive matrices (PM) [32], freehand copying of drawings (CD) [33], temporal rules induction [34], and word fluency (WF) [35]. FCb patients had generally preserved cognitive patterns. The lack of clinical deficits is consistent with studies that have reported cognitive impairments only under ad hoc test conditions [36–41].
Table 3. Neuropsychological assessment. Mean values (standard deviation).
| Groups | Tests | |||||||||
| IR | DR | IVM | WF | CD | FDS | BDS | FC | BC | TIQ WAIS-R | |
| FCb | 45.0 (10.3) | 9.8 (2.5) | 19.1 (2.0) | 36.2 (13.8) | 9.7 (1.7) | 6.2 (1.0) | 4.3 (1.2) | 5.3 (1.1) | 4.1 (0.6) | 98.6 (13.0) |
| FCb+ | 50.0 (5.9) | 10.8 (1.6) | 19.1 (2.5) | 37.5 (11.1) | 9.5 (1.8) | 6.4 (1.3) | 4.2 (1.3) | 5.7 (1.3) | 4.2 (0.4) | 99.8 (16.4) |
| FCb− | 39.2 (11.7) | 8.7 (3.1) | 19.0 (1.5) | 34.8 (17.4) | 9.9 (1.6) | 6 (0.6) | 4.5 (1.2) | 4.8 (0.7) | 4.0 (0.8) | 97.3 (8.9) |
| FCb_R | 44.8 (7.8) | 10.1 (2.1) | 18.6 (1.3) | 36.2 (15.9) | 8.9 (1.7) | 6.4 (1.3) | 4.7 (1.4) | 5.2 (1.4) | 4.1 (0.6) | 101.8 (11.9) |
| FCb_L | 45.3 (14.6) | 9.4 (3.4) | 19.8 (2.9) | 36.2 (11.3) | 10.9 (0.8) | 6 (0.7) | 3.8 (0.4) | 5.4 (0.5) | 4.2 (0.8) | 93.6 (12.9) |
| Cut off | 28.5 | 4.6 | 13.8 | 17.3 | 7.1 | 5 | 3 | 5 | 3 | 100 (15) |
IR: Rey's 15 mots short term (immediate recall); DR: Rey's 15 mots long term (delayed recall); IVM: immediate visual memory; WF: word fluency; CD: copying drawings; FDS: forward digit span; BDS: backward digit span; FC: forward Corsi; BC: backwords Corsi; TIQ WAIS – R: Wechsler Adult Intelligence Scale revised.
Motor impairment was quantified in patients using a modified version of the cerebellar motor deficit scale (Motor score) proposed by Apollonio and colleagues [42], which ranges from 0 (absence of any deficit) to 42 (presence of all deficits to the highest degree) and evaluates eight clinical signs (dysarthria, limb tone, postural tremor, upper and lower limb ataxia, standing balance, gait ataxia and ocular movements).
The experimental procedures were approved by the ethical committee of IRCCS Santa Lucia Foundation (CE-PROG.2-AG4-187); written consent for anonymous use of clinical data was obtained from each subject.
MRI data analysis
T2-weighted scans and 3D-T1-weighted magnetization-prepared rapid-acquisition gradient echo (3D MPRAGE; [43]) (TR = 11.0 ms; TE = 4.4 ms, TI = 300 ms; flip angle = 15°) images were obtained from all FCb subjects using a system that was operated at 1.5 T (Siemens Magnetom Vision, Germany).
Cerebellar lesions were first drawn on each patient's 3D MPRAGE images in native space using MRIcro (www.sph.sc.edu/comd/rorden/mricro.html), thus creating a region of interest (ROI) for each lesion and a corresponding lesion mask, smoothed to 8 mm full width at half maximum (FWHM) and a 0.001% threshold. The 3D MPRAGE images were processed using SPM2 (www.fil.ion.ucl.ac.uk/spm). For each FCb subject, images were reoriented manually according to the default template in SPM2 and normalized into the standard proportional stereotaxic space [Montreal Neurological Institute (MNI)] [44].
The macroscopic cerebellar damage in each patient was assessed after spatial normalization. The involvement of specific cerebellar structures was evaluated with the 3D MRI atlas of the human cerebellum [45] and the 3D MRI atlas of human cerebellar nuclei [46]. To detail the involvement of the nuclei, a 3D probabilistic map (51% to 60%) of right and left dentate/interposed cerebellar nuclei (D/I-N) [45] was reconstructed on a standard template using MRIcro, creating a ROI for the nuclei. Then, the patients' lesions and reconstruction of the nuclei were overlapped (Figure 1).
Figure 1. Lesion reconstruction in FCb+ and FCb− patients.

Each individual lesion is presented as an overlay of 2 representative coronal MRI T1-weighted templates in stereotaxic space after spatial normalization. The bottom left of the figure shows lesion overlap between patients and a reconstruction of the 3-D probabilistic map (51% to 60%) of right and left dentate/interposed (D/I-N) cerebellar nuclei. The color code indicates the number of subjects whose lesions overlapped with D/I-N nuclei.
Apparatus and task
Participants were seated in a dimly lit room, with their eyes 45 cm from a standard PC monitor (CRT noninterlaced, refresh rate 85 Hz). The presentation of visual stimuli and data acquisition were controlled using Matlab-based routines (www.mathworks.com). A joystick was fixed to the table, aligned with their body midline, and connected to a USB port.
Subjects performed 2 versions of a joystick reaction time task (Figure 2A). In the GO_only task (GOT), participants moved the joystick in response to a directional go signal (go trials). In the Countermanding task (CMT), subjects responded to the go signal (CMT go trials), as in the GOT, but withheld the movement when, in 25% of the overall trials, a stop signal was presented (stop trials) after varying delays (stop signal delay; SSD). In both GOT and CMT the primary task's demand was of responding as quickly as possible to go signal presentation. If subject correctly reacted to the stop signal, the trial was considered canceled, and if the subject failed to stop the movement, the trial was considered not canceled (Figure 2A). When the RT exceeded 1 s in the go trials, the trial was aborted and the subject received an acoustic error feedback. Successfully performed go trials were signaled using different acoustic feedback. Canceled and not canceled stop trials were signaled with 2 acoustic stimuli. Another acoustic stimulus sounded when the RT exceeded 500 ms as a warning for the participant to respect the primary task's demand of responding as quickly as possible.
Figure 2. Behavioral tasks and variables.

(A) Sequence of events in the go and stop trials for the Go_only (GOT) and countermanding (CMT) tasks. All trials started with the appearance of double vertical bars (fixation point; 2×11 pixels, separated by 2 pixels) in the middle of a screen. After a holding time (800–1200 ms), the central stimulus was replaced by an oriented arrow (Go signal; 12×12 pixels), indicating the direction of the movement. In the go trials (upper part), the subjects had to move the joystick toward the indicated direction as quickly as possible. The time between the Go signal and the onset of movement corresponded to the RT. In the stop trials (lower part), after a variable SSD, a central red square (Stop signal; 200×200 pixels) replaced the arrow, prompting the subjects to withhold the programmed movement and keep the joystick in the resting position. Trials with successfully suppressed movements are indicated as ‘cancelled’; trials with movements executed after a stop signal presentation are indicated as ‘not cancelled’. (B) Relationship among stop signal delay (SSD) duration and probability of error (pE). The longer is the SSD the higher is the pE. By assuming the go process identical in go and stop trials, the reaction time (RT) distribution, and the above relationship could be used to calculate the duration of the stop process (stop signal reaction time; SSRT; see text for details).
Each participant performed 2 sessions; one session with the right and one session with the left arm. Each session included 5 blocks of CMT (80 trials each) and 1 block of GOT (60 trials each). The order in which each arm was used was balanced between subjects, and the GOT always preceded the CMT for each arm. To facilitate the subject's performance on the CMT, the starting SSD for each block was always set to the shortest option (17 ms). A rest period (minimum of 2 minutes) between blocks was allowed.
Behavioral data analysis
A common behavioural finding of the CMT is that the probability of error (pE; i.e., probability of not cancelling the movement) in stop trials increases with the length of the SSD (Figure 2B; white portion of RT distribution). According to the Logan's model [48], the duration of the stop process (stop signal reaction time; SSRT) could be estimated, in each subject, by using the go trials RT distribution and the pE observed for each SSD used. Since is well known that the SSRT estimate is more reliable when obtained with an SSD corresponding to a pE that approximates 0.5 [47], we used a staircase procedure to select the SSD in each stop trial. The SSD was increased by 50 ms after each successfully canceled stop trial and decreased by 50 ms after 2 consecutive not canceled stop trials. The procedure automatically adapts the SSD duration to a subject's performance to obtain a session's pE that approximates 0.5 and is similar between groups (for a similar approach, see [48]).
Figure 3 shows the sequence of consecutive SSDs (red and white diamonds dots) for a CMT session, as adapted by a staircase algorithm to the performance of a control subject in the example. In the right portion of the figure, black squares represent the so-called ‘inhibition function’ (i.e., the proportion of not canceled [pE] stop trials at each SSD). The proportion of various SSDs in the session is shown as a histogram, highlighting those that were presented more than 10 times. These SSDs have been used to compute the session's representative SSD (braked red line in Figure 3) and to compute the session's SSRT (see [49] for a similar approach). According to the Logan's model [50], the duration of the stop process (SSRT) could be estimated by integrating the CMT go trials RT distribution until the integral equals the observed proportion of not canceled trials (pE) corresponding to the representative SSD (integration method; [50]). We also used a second method to estimate the SSRT corresponding to the subtraction of the representative SSD from the CMT RT distribution's average value (mean subtraction method; [50]). The use of at least two methods is common to increase the reliability of SSRT [47,51]. In our study the two methods provided comparable estimates, as confirmed by the analysis reported in the results section. To be consistent with classical approaches, we computed an average value and use it for all comparisons performed.
Figure 3. Stop trial sequence and inhibition function.

Each block of CMT (for both arms) started with an SSD of 17 ms (1 unit of refresh rate); this delay (stop signal delay; SSD) was increased by 50 ms after each successfully canceled stop trial and decreased by 50 ms after 2 consecutive not canceled stop trials. For each block, red diamonds represent SSDs presented more than 10 times in the session. These SSDs have been used to compute the representative SSD (red interrupted line). Rightmost panel shows the subject's inhibition function (black squares and black solid lines); i.e., the relationship among SSD duration and probability of error (pE). The number of SSDs presented is reported as a histogram. Filled bars indicate SSDs presented more than 10 times.
Before comparing between groups or conditions, the inhibition function obtained for each subject has been normalized as previously described [50] to remove influences due to subject's differences in RT, SSD and SSRT. Thus, for each participant we computed the so-called standardized (z-score) relative finish time (ZRFT) [per the equation: ZRFT = (RTmean−SSD−SSRT)/RTSD, where RTmean and RTSD are the mean and standard deviation, respectively, of the RT in the go trials]. After this normalization, the slope of the inhibition function is believed to measure one's ability to trigger the stop process, regardless of the variability in RT in the go trials, the different values of SSDs used and the estimated value of SSRT [50]. In general, the slope of the inhibition function will be steeper when the inhibitory mechanism is more efficient.
We also computed a measure of the proactive control [52] and the monitoring of the response after each stop signal appearance in the trial sequence or not canceled (error) stop trial. For each subject, we computed post-stopcorrect and post-stoperror slowing by subtracting the mean RT in the go trials after canceled and not canceled stop trials, respectively, from the mean RT in the go trials that were preceded and followed by a go trial (go-go-go sequence).
In FCb patients, we considered the functional arm to be the arm that was contralateral to the compromised cerebellar hemisphere and the hypofunctional arm as the one that was ipsilateral to it. In controls, the dominant and nondominant arms were considered equivalent to the functional and hypofunctional arms, respectively. This approach allows reducing the possibility that patients' impaired performances result statistically significant because of the comparison between controls' dominant arm and patients' affected arm (for a similar approach see [53]). Multifactorial analysis of variance (ANOVA) was used to test the significance of comparisons in RT and SSRT between patients and controls (or between tasks and arm used). Analysis of covariance (ANCOVA) was used to examine differences in the slope of the linear portion (0.15<pE<0.85) of the normalized inhibition function (ZRFT) between groups [54]. Post hoc contrast was conducted using Fisher LSD (for ANOVA) and Tukey-Kramer test (for ANCOVA) with Matlab's multicompare function.
We used an alpha level of 0.05 for all statistical tests. Thus, p values are reported in the text as exact values. However, p = 0.001 also include values of p<0.001. Similarly, when the software does not provide an exact p value, we indicate p<0.05.
Results
Tables 4 and 5 show the performance of the FCb and Control groups. No significant differences were observed in any measure between arms used. For this reason, the results are reported in the following sections with the arms pooled. We stress, however, that the arm factor was included in all statistical analyses. We will first describe the results for FCb as a homogeneous group and then for the FCb+ and FCb− subgroups separately. Additional analyses were performed with regard to the side of the lesion as the main factor to compare patients with lesions in the right (FCb_R) and left (FCb_L) cerebellum (Table 2).
Table 4. Behavioral results. Mean values (standard deviation) of RT in the GOT CMT and post-stop trials slowing.
| Groups | RT (GOT) | RT (CMT) | PostC slowing | PostE slowing | ||||
| HF/ND | F/D | HF/ND | F/D | HF/ND | F/D | HF/ND | F/D | |
| Controls | 419 (44) | 413 (55) | 457 (49) | 446 (37) | 16 (35) | 26 (23) | 41 (40) | 43 (54) |
| FCb | 462 (41) | 458 (54) | 545 (71) | 528 (60) | 55 (54) | 48 (40) | 85 (53) | 94 (67) |
| FCb+ | 460 (52) | 453 (61) | 560 (69) | 544 (62) | 76 (62) | 54 (53) | 90 (63) | 109 (81) |
| FCb− | 466 (41) | 458 (54) | 545 (71) | 528 (60) | 30 (37) | 41 (41) | 79 (43) | 76 (47) |
| FCb_R | 460 (48) | 454 (58) | 461 (79) | 539 (68) | 64 (66) | 61 (44) | 93 (83) | 84 (84) |
| FCb_L | 467 (46) | 464 (51) | 521 (54) | 510 (46) | 40 (30) | 29 (25) | 94 (35) | 87 (42) |
| Controls (males) | 419 (46) | 400 (57) | 450 (61) | 439 (46) | 16 (38) | 29 (28) | 29 (44) | 20 (56) |
| Controls (females) | 418 (45) | 423 (53) | 463 (38) | 452 (26) | 17 (32) | 25 (25) | 51 (35) | 62 (45) |
HF/ND: hypofunctional/nondominant arm; F/D: functional/dominant arm Controls group was sorted for gender differences. Three-way ANOVA (factors: gender, task, arm) revealed no gender effect on GOT or CMT RT for controls (main effect gender: F(1,20) = 1.69, p = 0.21; interaction between task and gender: F(1,20) = 0.002, p = 0.97). Significant difference in arm used was not detected (main effect arm: F(1,20) = 1.75, p = 0.20). Regardless of gender, all control subjects performed with longer RTs on the CMT than on the GOT (main effect task: F(1,20) = 5.0, p = 0.04). Analysis of post-stop go trials in CMT did not reveal gender to be a main effect (F(1,20) = 1.44, p = 0.24) or differences between arms (main effect arm: F(1,20) = 1.61, p = 0.43). PostE was increased in both groups (main effect trial type: F(1,20) = 7.15, p = 0.01) with a significant increase in females (interaction between trial type and gender: F(1,20) = 5.39, p = 0.03 and post hoc test: p = 0.02).
Table 5. SSRT estimate.
| Group | Estimate | HF/ND | Ttest | F/D | ttest |
| Control | SSRT Integration | 310 (50) | 304 (41) | - | |
| SSRT mean subtracion | 294 (53) | t (42) = 1.02; p = 0.31 | 294 (41) | t (42) = 0.77; p = 0.45 | |
| SSRT Average | 302 (51) | 299 (41) | |||
| FCb | SSRT Integration | 316 (74) | - | 315 (74) | - |
| SSRT mean subtracion | 302 (75) | t (24) = 0.49; p = 0.63 | 306 (65) | t (24) = 0.30; p = 0.76 | |
| SSRT Average | 309 (74) | 311 (69) | |||
| FCb − | SSRT Integration | 333 (85) | - | 354 (38) | - |
| SSRT mean subtracion | 321 (77) | t (10) = 0.26; p = 0.78 | 340 (24) | t (10) = 0.73; p = 0.48 | |
| SSRT Average | 327 (81) | 347 (31) | |||
| FCb + | SSRT Integration | 302 (67) | - | 281(83) | |
| SSRT mean subtracion | 286 (75) | t (12) = 0.42; p = 0.68 | 277 (77) | t (12) = 0.09; p = 0.93 | |
| SSRT Average | 293 (71) | 279 (79) | |||
| FCb_L | SSRT Integration | 350 (50) | - | 356 (49) | - |
| SSRT mean subtracion | 335(54) | t (8) = 0.46; p = 0.66 | 339 (46) | t (8) = 0.57; p = 0.59 | |
| SSRT Average | 343 (51) | 347 (47) | |||
| FCb_R | SSRT Integration | 295 (82) | - | 289 (78) | - |
| SSRT mean subtracion | 281 (82) | t (14) = 0.34; p = 0.74 | 286 (70) | t (14) = 0.08; p = 0.94 | |
| SSRT Average | 288 (81) | 287 (73) | |||
| Controls (males) | SSRT Integration | 291 (55) | - | 295 (26) | - |
| SSRT mean subtracion | 273 (64) | t (18) = 0.69; p = 0.50 | 282 (30) | t (18) = 1.03; p = 0.31 | |
| SSRT Average | 283 (59) | 288 (27) | |||
| Controls (females) | SSRT Integration | 325 (41) | - | 311(52) | - |
| SSRT mean subtracion | 312 (35) | t (22) = 0.87; p = 0.40 | 304 (47) | t (22) = 0.34; p = 0.74 | |
| SSRT Average | 319 (37) | 308 (49) |
Mean values (standard deviation). HF/ND: hypofunctional/nondominant arm; F/D: functional/dominant arm. The two methods did not differ in estimating the SSRT for each used arm in each experimental group, and their average was used for SSRT comparisons between groups and arms. T-tests comparisons between the methods are reported. Two-way ANOVA (factors: gender, arm) revealed no gender differences between female and male controls (main effect gender: F(1,20) = 2.56, p = 0.13; main effect arm F(1,20) = 0.10, p = 0.76; interaction between gender and arm: F(1,20) = 1.03, p = 0.37).
Go process duration and stop process triggering are impaired in patients affected by focal cerebellar lesions
By 3-way (factors: group, task, arm) ANOVA, on the GOT and CMT, FCb patients [factor group; F(1,33) = 28.70; p = 0.001; post hoc: FCb GOT vs control GOT, p = 0.013; FCb CMT vs control CMT, p = 0.001] needed significantly more time to respond (RT) to the go signal than controls (Table 4 and Figure 4 A1). FCb patients, as controls, had a longer RT on the CMT than on the GOT [factor task; F(1,33) = 23.50; p = 0.001; post hoc: FCb GOT vs FCb CMT, p = 0.001; Control GOT vs Control CMT, p = 0.016), suggesting that they adapt their behavior to the increased cognitive demand (presence of stop trials in the block] of the CMT. A significant main effect of the arm factor [F(1,33) = 3.80; p = 0.06] or interactions between it and other factors were not detected [interactions arm × task: F(1,33) = 0.32, p = 0.58; arm × group: F(1,33) = 0.82, p = 0.78; arm × task × group: F(1,33) = 0.31, p = 0.58]). In comparisons with controls, FCb patients slowed their RT in the CMT more than in the GOT. Statistical analysis revealed significant differences in the comparison of RT between groups [post hoc on mean RT and t-test on differences in RT: t(33) = 1.74, p = 0.04; Figure 4 A1, lower part]. We also observed (3-way ANOVA; factors group, task, arm) that the decrease in RT corresponded to a rise in RT variability [RT standard deviation (RT_SD) Figure 4 B1] from the GOT to CMT [main effect task: F(1,33) = 14.52, p = 0.001] in FCb patients but not controls [main effect group: F(1,33) = 4.25, p = 0.04; interaction task × group F(1,33) = 5.59, p = 0.02; post hoc FCb GOT vs FCb CMT: p = 0.001; post hoc control GOT vs control CMT: p = 0.57]. Accordingly, the differences in RT_SD was higher for FCb subjects than controls [Figure 4 B1, lower part; t-test: t(33) = 2.37, p = 0.01].
Figure 4. Go process analysis for groups of FCb patients and controls.
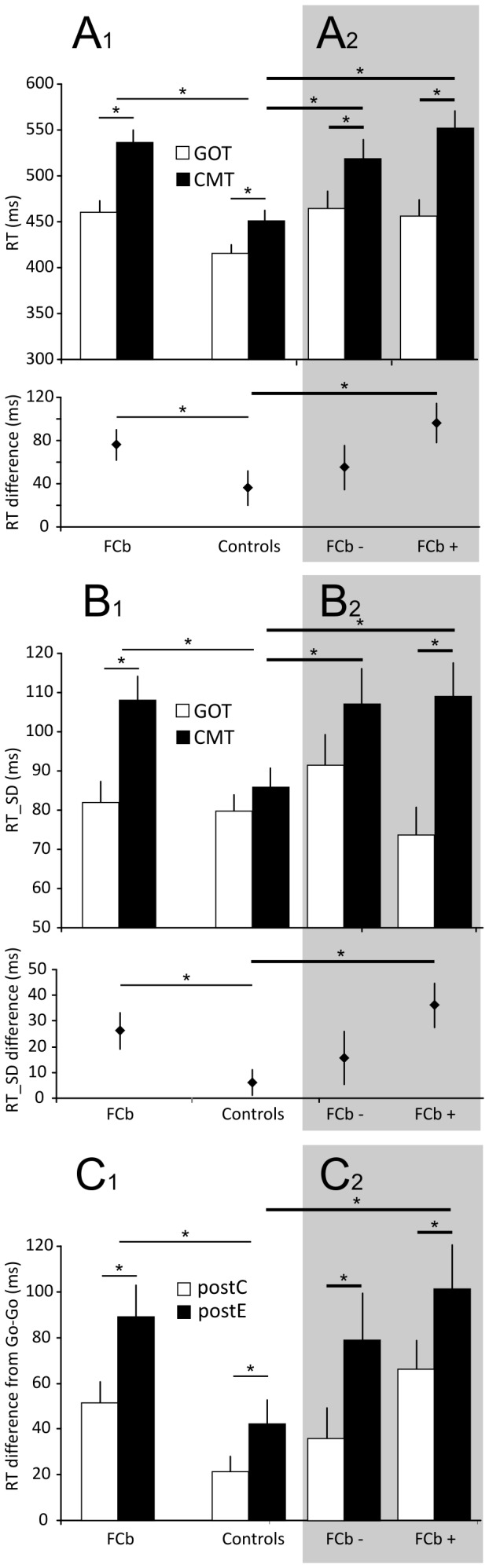
The left portion of each panel shows data from FCb groups and controls; the right portion of each panel (highlighted by the grey area) shows FCb− and FCb+ patient data. (A) Reaction time in the go trials of the GOT (white bars) and the CMT (black bars) task for FCb patients and controls (A1) and after sorting for DCN involvement in the cerebellar lesion (A2). The lower part of panel A1 and A2 shows the average difference in RT (CMT – GOT) for each of the groups indicated. (B) RT standard deviations (RT_SD) for the same groups as in A. The lower part of panel B1 and B2 shows the average difference in RT_SD (CMT – GOT) for each of the groups indicated. (C) Difference in RT of go trials following stop canceled trials (postC) and go trials following stop not canceled (error) trials (postE) from go trials preceded and followed by a go trial (go-go-go sequence) for the same groups in A and B. Significant differences (p<0.05) between groups and tasks are indicated (*). Thick lines indicate significant comparisons when considering FCb−, FCb+, and controls.
In our analysis of trial sequences (Figure 4 C1), after a stop trial, RT-slowing was evident in controls and FCb patients. In both groups, the RT in the post-stopcorrect (postC) and post-stoperror (postE) trials increased significantly (higher than 0; t-test all ps<0.05). By 3-way ANOVA (factors: group, trial type, arm), controls and FCb patients slowed their RT after erroneous stop trials more than after correct stop trials [main effect trial: F(1,33) = 18.90, p = 0.001; post hoc Control post-stopcorrect vs Control post-stoperror: p = 0.01; post hoc FCb post-stopcorrect vs FCb post-stoperror: p = 0.001]. FCb patients had a longer and significant post-stoperror slowing than controls [main factor group: F(1,33) = 8.73, p = 0.006; post hoc FCb post-stoperror vs Control post-stoperror: p = 0.02]. The difference in RT for post-stopcorrect trials did not differ between groups (post hoc FCb post-stopcorrect vs Control post-stopcorrect: p = 0.24).
The duration of the stop process (SSRT; see Methods) in the FCb group did not differ from that of controls [Table 5; two-way ANOVA; factors: group, arm; factor group: F (1,33) = 0.25, p = 0.62]. Between arms, SSRT did not differ in any group [main factor arm: F(1,33) = 0.02, p = 0.89; interaction F(1,33) = 0.11, p = 0.75]. Conversely, the slope of the regression line of the normalized inhibition function (ZRFT) in FCb subjects was significantly lower compared with controls [ANCOVA and post hoc comparisons: F(1,219) = 4.23; p = 0.04; see Figure 5 for details].
Figure 5. Normalized inhibition function.
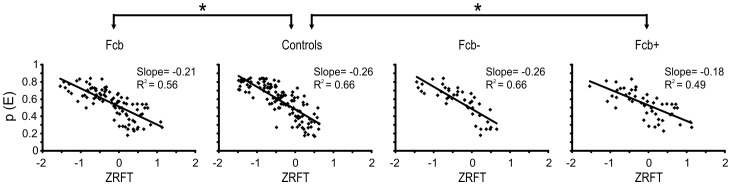
Each plot shows the group's probability to fail in canceling the stop trials (pE) as a function of ZRFT (black dots). The goodness of fit, slopes of the regression lines (black solid lines), and significant differences (*; p<0.05) between groups are indicated.
We also explored the relationship of the motor score (see Methods) with the go process and the stop process duration. The RT of FCb patients in the two tasks significantly correlated with the motor score (GOT: r = 0.83, p = 0.001; CMT: r = 0.57, p = 0.04) suggesting a relationship between RT and motor deficits as measured by the modified version of the cerebellar motor deficit scale. Conversely, patients' SSRT did not correlate with either motor score (r = 0.36; p = 0.23) or post-error slowing (r = 0.52; p = 0.07), indicating a lack of relationship between motor deficits and executive functions as measured by the countermanding task.
Overall, FCb subjects had a longer go process than controls in the GOT and CMT. The go process on the CMT was particularly affected in FCb patients: the slowness of the response, due to changes in the behavioral context, on CMT was accompanied by a significant increase in variability. In the analysis of trial history effects, the increase in RT in FCb patients was longer after errors. FCb patients experienced deficits in responding to the stop signal–i.e., in triggering the stop process (as estimated by ZRFT regression line slope)– although the time to complete it (as estimated by the SSRT), once started, did not differ from controls.
Cerebellar patients are more compromised when the lesion includes the DCN
The shaded grey areas in Figure 4 (panels A2, B2, C2) show the performance characteristics for FCb subjects, grouped by involvement (FCb+) or noninvolvement (FCb−) of DCN (see also Table 4 and 5) by the lesion. Statistical results refer to the analysis of these data and Controls data, as reported in Figure 4 A1, B1, C1.
By 3-way ANOVA (factor: group, task, arm; Figure 4 A2), group [F(2,32) = 14.32, p = 0.001] and task [F(1,32) = 22.25, p = 0.001] were significant main factors with regard to RT. The main effect of arm was not significant [F(1,32) = 3.16, p = 0.08]. Post hoc comparisons demonstrated significant differences in RT between GOT and CMT trials in each group (GOT RT vs CMT RT: Controls, p = 0.001; FCb+, p = 0.02; FCb−, p = 0.049). The between-group post hoc analysis also revealed significant differences in RT on the CMT between FCb+ and FCb− and Controls (FCb+ vs Controls, p = 0.001; FCb− vs Controls, p = 0.006). In contrast, between-group differences in RT on the GOT were not detected (GOT RT: Controls vs FCb+, p = 0.06; Controls vs FCb−, p = 0.05; FCb+ vs FCb−, p = 0.78).
By 3-way ANOVA of RT_SD (Figure 4 B2), we observed a significant effect of the main factor task [F(1,32) = 16.16, p = 0.001] and the group × task interaction [F(2,32) = 4.04, p = 0.03]. Significant differences were noted between tasks in FCb+ (p = 0.001) but not FCb− or Controls (GOT RT_SD vs CMT RT_SD: Controls, p = 0.24; FCb−, p = 0.12). No main effect of group or arm was detected [group: F(2,32) = 2.46, p = 0.10; arm: F(1,32) = 0.24, p = 0.62]. All other interactions between factors were not significant [interaction group × arm: F(2,32) = 2.46, p = 0.10; interaction task × arm: F(1,32) = 0.54, p = 0.47; interaction group × task × arm: F(2,32) = 2.26, p = 0.12].
Overall, our results suggest that when performing a movement implies a cognitive load, such as in CMT trials, cerebellar damage corresponds to a slowdown in readiness and an increase in the variability of responses to the Go signal. The same between-group differences did not appear in the GOT trials. Notably, between the GOT and CMT with regard to the RT and RT_SD, FCb+ patients performed worst. They had the largest increases in RT and RT variability, differing significantly from Controls in GOT when compared to CMT [One-way ANOVA (F (1,32) = 22.25, p = 0.001; post hoc p = 0.04] and in RT_SD difference [One-way ANOVA F (1,32) = 16.16, p = 0.001; post hoc p = 0.008] (Figure 4, bottom part of A2 and B2, respectively, compared with Controls in A1 and B1). The increase in RT and RT_SD between the CMT and GOT did not differ between Controls and FCb− (post hoc RT difference, p = 0.55; post hoc RT_SD difference, p = 0.40).
FCb+ patients also had the largest post-error slowing (Figure 4, panel C2 compared with Controls in C1). By 3-way ANOVA (factors: group, trial type, arm), there was a significant main effect of group [F(2,32) = 5.21, p = 0.01] and trial type [F(1,32) = 17.37, p = 0.001]. In post hoc comparisons, FCb+, FCb−, and controls had significantly longer post-stoperror slowing than post-stopcorrect slowing (post-stoperror vs post-stopcorrect: Controls, p = 0.018; FCb−, p = 0.013; FCb+, p = 0.03), and FCb+ post-stoperror slowing was significantly longer than in controls (p = 0.02). FCb+ and controls did not differ significantly with regard to post-stopcorrect slowing (p = 0.06). FCb− did not differ from controls in post-stoperror (p = 0.46) or post-stopcorrect slowing (LSD Fisher, p = 0.32). Significant differences in post-stoperror (LSD Fisher, p = 0.15) and post-stopcorrect slowing (LSD Fisher, p = 0.56) were not observed between FCb+ and FCb− patients. The main effects of arm [F(1,32) = 0.55, p = 0.46] and its interactions were not significant [interaction arm × trial: F(1,32) = 0.32, p = 0.57; interaction arm × group: F(2,32) = 0.35, p = 0.71; arm × group × trial: F(2,32) = 2.56, p = 0.08].
Stop process duration, measured as the length of the SSRT, did not differ between controls and patients or between the FCb+ and FCb− subgroups [2-way ANOVA; factors: groups, arm; factor group: F(2,32) = 1.73, p = 0.19; factor hand: F(1,32) = 0.01, p = 0.93; interaction: F(2,32) = 1.26, p = 0.30]. Conversely, the slope of the ZRFT, indicative of stop trigger efficiency, resulted shallower in FCb+ but not FCb− subjects [ANCOVA F(2,217) = 3.57, p = 0.03 and post hoc comparisons: p<0.05; see Figure 5 for details].
Finally, we controlled for the effect of the side of the cerebellar lesion for all behavioral parameters. During the recruitment of patients with and without DCN involvement, FCb subjects were not well distributed with regard to the side of the lesion (Table 1; 8 and 5 patients had lesions primarily in the right [FCb_R] and left [FCb_L]– cerebellum, respectively). Nevertheless, we extended the analyses above to include lesion side as a factor. By three-way ANOVA (factors: lesion side, arm, task), there were no differences between FCb_L and FCb_R in performance on the GOT and CMT. Both groups of patients were slower than controls in performing both tasks [main effect lesion side: F(2,32) = 14.30, p = 0.001] and performed the CMT with a longer RT than on the GOT [main effect task: F(1,32) = 19.83, p = 0.001; interaction between task and lesion: F(2,32) = 2.15, p = 0.13]. No between-arm differences were observed [main effect arm: F(1,32) = 2.78, p = 0.13]. The only significant effect was an increase in the variability of RT in FCb patients with right lesions compared with controls (p = 0.03 post hoc; 3-way ANOVA; factors: group, task, arm) by contrasting RT on the GOT versus CMT [main effect lesion side: F(2,32) = 2.49, p = 0.10; main effect task: F(1,32) = 13.92, p = 0.001; main effect arm: F(1,32) = 0.31, p = 0.59].
No differences between FCb_L and FCb_R were detected in post-stop performance. By three-way ANOVA (factors: lesion side, hand, trial type), there was a general increase [main effect trial type: F(1,32) = 19.95, p = 0.001] and a significant difference between controls and patients [main effect lesion: F(2,32) = 4.45, p = 0.02] in post-stoperror slowing. No significant effects of arm [F(1,32) = 0.35, p = 0.35] or the interaction between trial type and lesion side on post-stop slowing were detected [F(2,32) = 1.77, p = 0.19]. By two-way ANOVA (factors: lesion side, arm), there were no significant differences in SSRT between lesion side [main effect lesion side: F(2,32) = 2.11, p = 0.14; main effect arm: F(1,32) = 0.0006, p = 0.98; interaction between lesion side and arm: F(2,32) = 0.07, p = 0.93]. Based on ZRFT, there was no difference in the slope between FCb_L and FCb_R [F(2,217) = 2.35, p = 0.10].
In conclusion, our cohort of focal cerebellar patients (FCb) have significant differences from controls in the go and stop processes when lesions involve the DCN. Although the FCb+ and FCb− subgroups had longer RTs overall than controls, the go process in the CMT was affected only in FCb+ patients, as evidenced by the significant increase in variability when the go signal was followed by a stop signal. In our analysis of the trial history effect, the increase in RT after errors was more evident in FCb+ subjects. Despite having comparable SSRTs as controls, FCb+ subjects, but not FCb−, were impaired in their ability to trigger the stop process.
Discussion
We report the performance of FCb subjects on a CMT and provide evidence that the cerebellum has a specific function in the cognitive control of movement suppression and performance monitoring, two core properties of EC [2,25].
Cerebellar function in executive motor control
According to evidence in humans and animals [3,4,25], various areas of the frontal cortex and subcortical structures contribute to the voluntary control of movement. Among subcortical structures, the subthalamic nucleus (STN) is often proposed as a significant area in the basal ganglia that supports the cortical control of the generation of movements by gating the inhibitory control of the internal segment of the globus pallidus (GPi) over the thalamic drive [7,10].
The STN is linked functionally to the inferior frontal cortex (IFC), presupplementary motor areas (preSMAs), anterior cingulate cortex (ACC), dorsal premotor cortex (PMd), and frontal eye field (FEF)–areas that regulate the executive control of movement generation 10,25,55–58. With regard to motor control, basal ganglia models [59–62] suggest that movement can be suppressed by activation of the STN through the ‘hyperdirect’ (in which the STN receives a direct excitatory projection from the cortex) or ‘indirect’ (the STN is activated by contrasting–via the striatum–the inhibition that is exerted by the external segment of the globus pallidus [GPe]) pathway.
The use of neurotrophic viruses to track the connections between areas of the brain has provided anatomical evidence for the reciprocal influence of the cerebellum and cortical-basal ganglia-cortical loops, wherein DCN, the output of the cerebellum, influence the ‘indirect’ pathway by specifically targeting striatal neurons that project to the GPe; in turn, the STN projects to pontine nuclei and thus governs the activity of the cerebellar cortex [12]. The emerging model contends that frontal cortical areas (e.g., the PFc, SMA, and PMd) send movement-related decisions to the cerebellum and basal ganglia. Both nodes send signals back to the primary motor cortex via the thalamus that are sufficient to permit or facilitate execution of the programmed movement or, conversely, countermand it by activating the indirect or hyperdirect pathway [62]. The interplay between the cerebellum and basal ganglia could thus contribute to fine executive control of motor generation.
Our results show that FCb patients do not experience deficits in the duration of the stop process. The SSRT was similar between cerebellar and control subjects. Grouping the patients by DCN involvement did not yield further information. However, the comparison of ZRFT slopes suggests that FCb patients have specific inhibitory deficits, failing more often than controls in triggering the stop process. In our group of patients, the impairment was evident when the lesions involved the DCN. In FCb+ patients, the slope of the inhibition function was the shallowest measured out of the slope measured in the other groups and the only one that differed significantly from slope in controls.
In our analysis of the go process, patients took longer than controls to react to the go signal. Prolonged RT is a hallmark of cerebellar patients [63]. Luciani [64] proposed that DCN exert a tonic facilitatory influence over the motor cortex. The activity of DCN is in turn modified by the inhibitory control of the cerebellar cortex [65]. We observed that this effect correlates with the cognitive load of the motor task that is performed–e.g., when a sudden stop signal is expected in a sequence of standard reaction time trials, requiring cancellation of the action. This form of control is commonly termed proactive control [52].
We propose that the cerebellum receives a copy of the cortically originated signal to suppress a movement via the STN [10,12] and contributes to executive control by modulating the tonic inhibition that is exerted by the cerebellar cortex on the DCN. The results of this process are normally relayed back to the basal ganglia through the indirect pathway, influencing its contribution to inhibitory action. Our results suggest that focal lesions preserve the contribution of the cerebellum in determining the speed of the stop process. Action interruption occurs with lower probability in patients, but when it does occur, it takes the same amount of time as controls; this was particularly evident after DCN damage.
However, our results could have been influenced by the small number of subjects in the FCb subgroups after sorting patients by DNC involvement, which could have introduced effects of other factors that we did not control for with regard to the behavioral performance of FCb patients. For example, as shown in Table 2, FCb+ and FCb− did not match controls for the gender and had a nonhomogeneous distribution of it. Recent evidence has highlighted a disparate pattern of activation of cerebral areas between males and females during the execution of stop trials in the CMT [66,67]. However, this gender effect was not affecting the reported performance on the task. We observed only a significantly longer post-stoperror slowing in female than male controls (see Tables 4 and 5; statistical details are described in the caption), contrasting what we observed on grouping DCN lesions, because the FCb+ group comprised primarily males (Table 2).
Similarly, no main effect of the lesion side was detected, as described in other motor paradigms that are focused on procedural learning and time elaboration [68,69] and in neuroimaging studies [70,71]. However, due to the difference in the number of subjects in the FCb_R and FCb_L groups (see Table 2) we cannot exclude the possibility that the absence of a lesion side effect was related to the difference in sample size. These aspects require further investigation in future studies.
Cerebellar function in the proactive control of behavior and error monitoring
In the CMT, go trials always have the potential to turn into stop trials, thus requiring a continuous update (lengthening) of the RT to maximize stop success [2,50]. We observed that the RT of go trials in the CMT was significantly longer than in the GOT for the patient and control groups, more so in the former. Cerebellar patients were strongly affected by the trial history effect. Their increase in go trial RT, following a stop trial, was larger than in controls, especially after having failed to stop (i.e., they experienced notable post-error slowing). This deceleration became significant if the lesion involved the DCN, indicating that in performing a basic task (go trials), these patients experienced greater perturbations by an infrequent and unpredictable event (stop trial) and by errors than other subjects.
This evidence also supports the hypothesis that the anatomical link between the basal ganglia and cerebellum has a significant function in executive control. Our data suggest that the contribution of the basal ganglia to error-related cognitive control, as reported [16–19,72], requires efficient interaction with the cerebellum.
The cerebellum has been hypothesized to mediate the correction of errors in the motor and cognitive domains [73–77]. The cerebellum, in a network that comprises the PFC, the dorsal ACC, thalamus, and SMA, has also been proposed to affect behavioral adjustments after presentation of a stop signal in the CMT [11]. Our data provide evidence that a lesion in the cerebellum impairs the processing of errors and that this effect is present when DCN are involved.
Notably, the effects of DCN damage that we observed are consistent with previous studies on various cognitive domains [28,78,79]. However, our findings must be confirmed in future studies using advanced methods to better characterize the site of the lesion in the DCN [80]. Such steps will allow us to determine the various functions of the interposite and dentate nuclei in cerebellar-cortical loops.
Moreover, future studies should focus on assessing the CMT in patients who are affected by degenerative cerebellar atrophy, in which the Purkinje cells are more selectively affected and thus influence the inhibitory control of DCN.
In conclusion, our study has provided evidence of impaired efficiency of execution of actions and performance monitoring in the presence of cerebellar damage, increasing our understanding of motor and cognitive cerebellar symptoms and the consequent behavioural impairments. We propose that the cerebellar function in executive control is mediated by its reciprocal interaction with the basal ganglia, although the possibility of a direct cerebello-prefrontal interaction cannot be excluded in controlling the evaluated functions.
Acknowledgments
The editing support of Blue Pencil Science is acknowledged.
Funding Statement
This study was supported by Italian Ministry of Health grants (RC08G- to MM, and ML, in part) and by the Italian National Institute of Health (ISS; project BRUCOS grant agreement 11US/38A to SF, in part). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Reichenberg A (2010) The assessment of neuropsychological functioning in schizophrenia. Dialogues Clin Neurosci 12: 383–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Verbruggen F, Logan GD (2008) Response inhibition in the stop-signal paradigm. Trends in Cognitive Sciences 12: 418–424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Alvarez JA, Emory E (2006) Executive function and the frontal lobes: A meta-analytic review. Neuropsychology Review 16: 17–42. [DOI] [PubMed] [Google Scholar]
- 4. Aron A, Robbins T, Poldrack R (2004) Inhibition and the right inferior frontal cortex. Trends in Cognitive Science 8: 170–177. [DOI] [PubMed] [Google Scholar]
- 5. Rubia K, Smith AB, Brammer MJ, Taylor E (2003) Right inferior prefrontal cortex mediates response inhibition while mesial prefrontal cortex is responsible for error detection. Neuroimage 20: 351–8. [DOI] [PubMed] [Google Scholar]
- 6. Garavan H, Ross TJ, Murphy K, Roche RA, Stein EA (2002) Dissociable executive functions in the dynamic control of behavior: inhibition, error detection, and correction. Neuroimage 17: 1820–29. [DOI] [PubMed] [Google Scholar]
- 7.Aron AR, Poldrack RA, Wise SP (2009) Cognition: Basal Ganglia Role. In: Squire LR editor. Encyclopedia of Neuroscience, volume 2 . Oxford: Academic Press. pp. 1069–1077. [Google Scholar]
- 8. Li CS, Yan P, Sinha R, Lee TW (2008) Subcortical processes of motor response inhibition during a stop signal task. Neuroimage 41: 1352–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Chevrier AD, Noseworthy MD, Schachar R (2007) Dissociation of response inhibition and performance monitoring in the stop signal task using event-related fMRI. Human Brain Mapping 28: 1347–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Aron AR, Poldrack RA (2006) Cortical and subcortical contributions to Stop signal response inhibition: role of the subthalamic nucleus. The Journal of Neuroscience 26: 2424–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Ide JS, Li CS (2011) A cerebellar thalamic cortical circuit for error-related cognitive control. Neuroimage 54: 455–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Bostan AC, Strick PL (2010) The cerebellum and basal ganglia are interconnected. Neuropsychology Review 20: 261–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Miall RC (1998) The cerebellum, predictive control and motor coordination. Novartis Found Symp 218: 272–84. [DOI] [PubMed] [Google Scholar]
- 14. Schlerf J, Ivry RB, Diedrichsen J (2012) Encoding of sensory prediction errors in the human cerebellum. J Neurosci 32: 4913–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Wolpert DM, Miall RC, Kawato M (1998) Internal models in the cerebellum. Trends Cogn Sci 2: 338–47. [DOI] [PubMed] [Google Scholar]
- 16. Falkenstein M, Hielscher H, Dziobek I, Schwarzenau P, Hoormann J, et al. (2001) Action monitoring, error detection, and the basal ganglia: an ERP study. Neuroreport 12: 157–61. [DOI] [PubMed] [Google Scholar]
- 17. Falkenstein M, Hoormann J, Christ S, Hohnsbein J (2000) ERP components on reaction errors and their functional significance: a tutorial. Biological Psychology 51(2–3): 87–107. [DOI] [PubMed] [Google Scholar]
- 18. Beste C, Saft C, Andrich J, Gold R, Falkenstein M (2006) Error processing in Huntington's disease. PLoS One 1: e86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Ullsperger M, von Cramon DY (2006) The role of intact frontostriatal circuits in error processing. Journal of Cognitive Neuroscience 18: 651–64. [DOI] [PubMed] [Google Scholar]
- 20. Neau JP, Arroyo-Anllo E, Bonnaud V, Ingrand P, Gil R (2000) Neuropsychological disturbances in cerebellar infarcts. Acta Neurol Scand 102: 363–52. [DOI] [PubMed] [Google Scholar]
- 21. Tanaka H, Harada M, Arai M, Hirata K (2003) Cognitive dysfunction in cortical cerebellar atrophy correlates with impairment of the inhibitory system. Neuropsychobiology 47: 206–11. [DOI] [PubMed] [Google Scholar]
- 22. Tu PC, Yang TH, Kuo WJ, Hsieh JC, Su TP (2006) Neural correlates of antisaccade deficits in schizophrenia, an fMRI study. Journal of Psychiatric Research 40: 606–61. [DOI] [PubMed] [Google Scholar]
- 23. Rubia K, Smith AB, Taylor E, Brammer M (2007) Linear age-correlated functional development of right inferior fronto-striato-cerebellar networks during response inhibition and anterior cingulate during error-related processes. Hum Brain Mapp 28: 1163–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Thoma P, Koch B, Heyder K, Schwarz M, Daum I (2008) Subcortical contributions to multitasking and response inhibition. Behavioral Brain Research 194: 214–22. [DOI] [PubMed] [Google Scholar]
- 25. Schall JD, Godlove DC (2012) Current advances and pressing problems in studies of stopping. Curr Opin Neurobiol 22: 1012–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, et al. (1999) Hypofrontality in attention deficit hyperactivity disorder during higher-order motor control: a study with functional MRI. Am J Psychiatry 156: 891–6. [DOI] [PubMed] [Google Scholar]
- 27. Habas C (2010) Functional imaging of deep cerebellar nuclei: a review. Cerebellum 9: 22–8. [DOI] [PubMed] [Google Scholar]
- 28. Tedesco AM, Chiricozzi FR, Clausi S, Lupo M, Molinari M, et al. (2011) The cerebellar cognitive profile. Brain 134: 3669–83. [DOI] [PubMed] [Google Scholar]
- 29.Rey A (1958) L'examen clinique en psychologie. Paris: Presses Universiteries de France. [Google Scholar]
- 30. Carlesimo GA, Marfia GA, Loasses A, Caltagirone C (1996) Perceptual and conceptual components in implicit and explicit stem completion. Neuropsychologia 34: 785–92. [DOI] [PubMed] [Google Scholar]
- 31. Corsi PM (1972) Human memory and the medial temporal regions of the brain. Dissert Abst Int 34: 891B. [Google Scholar]
- 32.Raven JC (1949) Sets A, Ab, B: Board and Book forms. Progressive matrices London: Lewis.
- 33. Gainotti G, Miceli G, Caltagirone C (1977) Constructional apraxia in left brain-damage patients: a planning disorders? Cortex 13: 109–18. [DOI] [PubMed] [Google Scholar]
- 34. Villa G, Gainotti G, De Bonis C, Marra C (2008) Double dissociation between temporal and spatial pattern processing in patients with frontal and parietal damage. Cortex 26: 399–407. [DOI] [PubMed] [Google Scholar]
- 35. Borkowsky JG, Benton AL, Spreen O (1967) Word fluency and brain-damage. Neuropsychologia 5: 135–140. [Google Scholar]
- 36. Leggio MG, Silveri MC, Petrosini L, Molinari M (2000) Phonological grouping is specifically affected in cerebellar patients: a verbal fluency study. Journal of Neurology, Neurosurgery and Psychiatry 69: 102–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Leggio MG, Tedesco AM, Chiricozzi FR, Clausi S, Orsini A, et al. (2008) Cognitive sequencing impairment in patients with focal or atrophic cerebellar damage. Brain 131: 1332–43. [DOI] [PubMed] [Google Scholar]
- 38. Molinari M, Petrosini L, Misciagna S, Leggio MG (2004) Visuospatial abilities in cerebellar disorders. Journal of Neurology Neurosurgery and Psychiatry 75: 235–40. [PMC free article] [PubMed] [Google Scholar]
- 39. Restuccia D, Della Marca G, Valeriani M, Leggio MG, Molinari M (2007) Cerebellar damage impairs detection of somatosensory input changes. A somatosensory mismatch-negativity study. Brain 130: 276–87. [DOI] [PubMed] [Google Scholar]
- 40. Silveri MC, Di Betta AM, Filippini V, Leggio MG, Molinari M (1998) Verbal short-term store-rehearsal system and the cerebellum. Evidence from a patient with a right cerebellar lesion. Brain 121: 2175–87. [DOI] [PubMed] [Google Scholar]
- 41. Justus T The cerebellum and English grammatical morphology: evidence from production, comprehension, and grammaticality judgments. (2004) Journal of Cognitive Neuroscience 16: 1115–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Appollonio IM, Grafman J, Schwartz V, Massaquoi S, Hallett M (1993) Memory in patients with cerebellar degeneration. Neurology 43: 1536–44. [DOI] [PubMed] [Google Scholar]
- 43. Mugler JPD, Brookrman JR (1990) Three-dimensional magnetization-prepared rapid gradient-echo imaging (3D MP RAGE). Magn Reson Med 15: 152–157. [DOI] [PubMed] [Google Scholar]
- 44. Brett M, Leff AP, Rorden C, Ashburner J (2001) Spatial normalization of brain images with focal lesions using cost function masking. Neuroimage 14: 486–500. [DOI] [PubMed] [Google Scholar]
- 45.Schmahmann JD, Dojon J, Toga AW, Petrides M, Evans AC (2000) MRI atlas of the human cerebellum. San Diego: Academic Press. [DOI] [PubMed] [Google Scholar]
- 46. Dimitrova A, Zeljko D, Schwarze F, Maschke M, Gerwig M, et al. (2006) Probabilistic 3D MRI atlas of the human cerebellar dentate/interposed nuclei. Neuroimage 30: 12–25. [DOI] [PubMed] [Google Scholar]
- 47. Band GP, van der Molen MW, Logan GD (2003) Horse-race model simulations of the stop-signal procedure. Acta Psychol (Amst) 112: 105–142. [DOI] [PubMed] [Google Scholar]
- 48. Brunamonti E, Pani P, Papazachariadis O, Onorati P, Albertini G, et al. (2011) Cognitive control of movement in Down syndrome. Research in Developmental Disabililities 32: 1792–7. [DOI] [PubMed] [Google Scholar]
- 49. Mirabella G, Pani P, Ferraina S (2009) The presence of visual gap affects the duration of stopping process. Exp Brain Res 192: 199–209. [DOI] [PubMed] [Google Scholar]
- 50.Logan GD (1984) On the ability to inhibit thought and action: A users' guide to the stop signal paradigm. In: Dagenbach D, Carr TH, editors. Inhibitory Processes in Attention, Memory, and Language. San Diego: Academic Press. p 189–239. [Google Scholar]
- 51. Brunamonti E, Ferraina S, Paré M (2012) Controlled movement processing: Evidence for a common inhibitory control of finger, wrist, and arm movements. Neuroscience 215: 69–78. [DOI] [PubMed] [Google Scholar]
- 52. Braver TS (2012) The variable nature of cognitive control: a dual mechanisms framework. Trends Cogn Sci (Regul. Ed.) 16: 106–113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Duque J, Thonnard JL, Vandermeeren Y, Sébire G, Cosnard G, Olivier E (2003) Correlation between impaired dexterity and corticospinal tract dysgenesis in congenital hemiplegia. Brain 126: 732–47. [DOI] [PubMed] [Google Scholar]
- 54.Zar J (2009) Biostatistical Analysis, 4th edition, Prentice-Hall. [Google Scholar]
- 55. Mirabella G, Pani P, Ferraina S (2011) Neural correlates of cognitive control of reaching movements in the dorsal premotor cortex of rhesus monkeys. Journal of Neurophysiology 106: 1454–1466. [DOI] [PubMed] [Google Scholar]
- 56. Marcos E, Pani P, Brunamonti E, Deco G, Ferraina S, Verschure P (2013) Neural variability in premotor cortex is modulated by trial history and predicts behavioral performance. Neuron 78: 249–255. [DOI] [PubMed] [Google Scholar]
- 57. Duann JR, Ide JS, Luo X, Li CS (2009) Functional connectivity delineates distinct roles of the inferior frontal cortex and presupplementary motor area in stop signal inhibition. The Journal of Neuroscience 29: 10171–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Nambu A, Takada M, Inase M, Tokunoz H (1996) Dual somatotopical representations in the primate subthalamic nucleus: evidence for ordered but reversed body-map transformations from the primary motor cortex and the supplementary motor area. The Journal of Neuroscience 16: 2671–2683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Brunia CHM (2003) How is stopping realized? In: Ullsperger M, Falkenstein M, editors. Errors, Conflicts, and the Brain. Current Opinions on Performance Monitoring. Leipzig: MPI of Cognitive Neuroscience. [Google Scholar]
- 60. Mink JW (1996) The basal ganglia: focused selection and inhibition of competing motor programs. Progress in Neurobiology 50: 381–425. [DOI] [PubMed] [Google Scholar]
- 61. Nambu A, Tokuno H, Takada M (2002) Functional significance of the cortico-subthalamo-pallidal ‘hyperdirect’ pathway. Neuroscience Research 43: 111–7. [DOI] [PubMed] [Google Scholar]
- 62. Alexander GE, Crutcher MD (1990) Functional architecture of basal ganglia circuits: neural substrates of parallel processing. Trends in Neuroscience 13: 266–71. [DOI] [PubMed] [Google Scholar]
- 63. Holmes G (1939) The cerebellum of man. Brain 62: 1–30. [Google Scholar]
- 64.Luciani L (1891) Il cervelletto: nuove studi di fisiologia normale e patologica. Firenze: Le Monnier. [Google Scholar]
- 65.Ito M (1984) The cerebellum and neural control. New York: Raven. [Google Scholar]
- 66. Li CS, Huang C, Constable RT, Sinha R (2006) Gender differences in the neural correlates of response inhibition during a stop signal task. Neuroimage 32: 1918–29. [DOI] [PubMed] [Google Scholar]
- 67. Li CS, Zhang S, Duann JR, Yan P, Sinha R, Mazure CM (2009) Gender Differences in Cognitive Control: an Extended Investigation of the Stop Signal Task. Brain Imaging Behav 3: 262–276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Molinari M, Leggio MG, Solida A, Ciorra R, Misciagna S, et al. (1997) Cerebellum and procedural learning: evidence from focal cerebellar lesions. Brain 120: 1753–62. [DOI] [PubMed] [Google Scholar]
- 69. Molinari M, Leggio MG, Filippini V, Gioia MC, Cerasa A, et al. (2005) Sensorimotor transduction of time information is preserved in subjects with cerebellar damage. Brain Res Bull 67: 448–58. [DOI] [PubMed] [Google Scholar]
- 70. Van Mier HI, Tempel LW, Perlmutter JS, Raichle ME, Petersen SE (1998) Changes in brain activity during motor learning measured with PET: effects of hand of performance and practice. J Neurophysiol 80: 2177–2199. [DOI] [PubMed] [Google Scholar]
- 71. Van Mier HI, Petersen SE (2002) Role of the cerebellum in motor cognition. Ann N Y Acad Sci 978: 334–53. [DOI] [PubMed] [Google Scholar]
- 72. Willemssen R, Müller T, Schwarz M, Hohnsbein J, Falkenstein M (2008) Error processing in patients with Parkinson's disease: the influence of medication state. Journal of Neural Transmission 115: 461–8. [DOI] [PubMed] [Google Scholar]
- 73. Ebner TJ, Pasalar S (2008) Cerebellum predicts the future motor state. Cerebellum 7: 583–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Doya K (2000) Complementary roles of basal ganglia and cerebellum in learning and motor control. Current Opinion in Neurobiology 10: 732–9. [DOI] [PubMed] [Google Scholar]
- 75. Blakemore SJ, Frith CD, Wolpert DM (2001) The cerebellum is involved in predicting the sensory consequences of action. Neuroreport 12: 1879–84. [DOI] [PubMed] [Google Scholar]
- 76. Imamizu H, Miyauchi S, Tamada T, Sasaki Y, Takino R, et al. (2000) Human cerebellar activity reflecting an acquired internal model of a new tool. Nature 403: 192–5. [DOI] [PubMed] [Google Scholar]
- 77. Courchesne E, Allen G (1997) Prediction and preparation, fundamental functions of the cerebellum. Learning and Memory 4: 1–35. [DOI] [PubMed] [Google Scholar]
- 78. Ilg W, Giese MA, Gizewski ER, Schoch B, Timmann D (2008) The influence of focal cerebellar lesions on the control and adaptation of gait. Brain 131: 2913–27. [DOI] [PubMed] [Google Scholar]
- 79. Clausi S, Bozzali M, Leggio MG, Di Paola M, Hagberg GE, et al. (2009) Quantification of gray matter changes in the cerebral cortex after isolated cerebellar damage: a voxel-based morphometry study. Neuroscience 162: 827–35. [DOI] [PubMed] [Google Scholar]
- 80. Diedrichsen J, Maderwald S, Küper M, Thürling M, Rabe K, et al. (2011) Imaging the deep cerebellar nuclei: a probabilistic atlas and normalization procedure. Neuroimage 54: 1786–94. [DOI] [PubMed] [Google Scholar]