

Abstract
The bone-regenerative potentials of the periosteum have been explored as early as the 17th century. Over the past few years, however, much has been discovered in terms of the molecular and cellular mechanisms that control the periosteal contribution to bone regeneration. Lineage tracing analyses and knock-in transgenic mice have helped define the relative contributions of the periosteum and endosteum to bone regeneration. Additional studies have shed light on the critical roles that BMP, FGF, Hedgehog, Notch, PDGF, Wnt, and inflammation signaling have or may have in periosteal-mediated bone regeneration, fostering the path to novel approaches in bone-regenerative therapy. Thus, by examining the role that each pathway has in periosteal-mediated bone regeneration, in this review we analyze the status of the current research on the regenerative potential of the periosteum. The provided analysis aims to inform both clinician-scientists who may have interest in the current studies about the biology of the periosteum as well as dental surgeons who may find this review useful to perform periosteal-harnessing bone-regenerative procedures.
Keywords: regenerative surgery, stem cells, osteoprogenitor cells, periosteum, tissue engineering, bone healing
The Anatomy of the Periosteum
In 1925, Blaisdell published a classic paper in which he describes the periosteum as a structure comprised of 2 layers, with osteoblast-like cells in the inner layer (Blaisdell, 1925). We now know in more detail that the outer “fibrous” layer consists of fibroblasts, collagen, and elastin along with a nerve and microvascular network (Allen et al., 2004). These components provide mechanical stability to the periosteum. The inner “cambium layer” is highly cellular and presents with cells that influence bone formation and bone repair (Allen et al., 2004). Specifically, this layer has adult mesenchymal skeletal progenitor cells, osteoblasts, smaller and more isodiametric fibroblasts, and sympathetic nerves (Allen et al., 2004). These features allow the periosteum to be considered a structure with regenerative capacity (Fig. 1). As the bone ages, the reduction in osteoblast number leads to a distinctive atrophy and thinning of the cambium layer (Allen et al., 2004).
Figure 1.
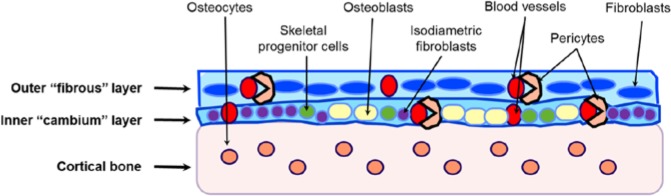
Cell populations present in the outer and inner layers of the periosteum. The outer more “fibrous” layer contains “elongated” fibroblasts and blood vessels. The inner “cambium” layer presents with skeletal progenitor cells, osteoblasts, small and isodiametric fibroblasts, and blood vessels. Because of its high vascularity, the periosteum contains a large number of endothelial pericytes surrounding the blood vessels.
Because of its high vascularity, there are also numerous endothelial pericytes within the periosteum (Diaz-Flores et al., 1992) (Fig. 1). Pericytes are cells in physical contact with capillary endothelial cells. Under specific culture conditions, these cells can differentiate into several cell types, including osteoblasts (Reilly et al., 1998). Thus, the pericytes could be considered a supplemental source of osteoprogenitor cells within the periosteum (Diaz-Flores et al., 1992).
The Role of the Periosteum in Bone Healing
The remarkable regenerative capability of the periosteum was realized centuries ago (Chang and Knothe Tate, 2012). However, only recently have bone biologists been able to understand the precise role of the periosteum in bone healing.
Bone fracture healing has been studied in great detail over the past years. The initial phase of fracture healing is characterized by an acute inflammatory response which includes blood clot formation, inflammatory cell migration, and granular tissue formation. The inflammatory phase is believed to stimulate mesenchymal cell migration and proliferation. Closely following the inflammation period, mesenchymal cells aggregate at the repair site and differentiate into chondrocytes and osteoblasts. Collagen matrix is secreted and subsequently mineralized. These events result in the formation of a soft callus that bridges the 2 fracture ends. With time, the soft callus will continue to ossify, and woven bone will be formed. Eventually, upon remodeling, the healing bone will restore its original shape and structure (Dimitriou et al., 2011; Colnot et al., 2012).
The periosteum makes major contributions to this complex bone-healing process. As early as 24 to 48 hours after the injury, an acute inflammatory reaction can be seen at the periosteum (Colnot et al., 2003; Lu et al., 2005). Subsequently, periosteal cells start to proliferate, and a thickening of periosteum is observed. This process is defined as periosteal activation. By comparing periosteal with endosteal/bone marrow injuries in mice tibia, Colnot found that periosteal injuries heal by endochondral ossification, whereas endosteal/bone marrow injuries heal by intramembranous ossification without chondrogenesis (Colnot, 2009). Live bone grafting in Rosa26 transgenic reporter mice further demonstrated that the expansion of the mesenchymal progenitor cell population from the periosteum largely accounted for the formation of cartilage and bone within the callus (Zhang et al., 2005; Colnot, 2009). Differently, cells of the bone marrow or endosteum remain in the marrow cavity and do not migrate out to form the callus (Colnot, 2009). These results indicate that the contributions of periosteal cells are critical for callus formation. In fact, activation of periosteum-derived progenitor cells induces robust chondrogenesis and osteogenesis, accompanied by marked induction of angiogenesis, which eventually leads to vascularization and remodeling of bone grafts (Zhang et al., 2005; Colnot et al., 2012) (Fig. 2).
Figure 2.
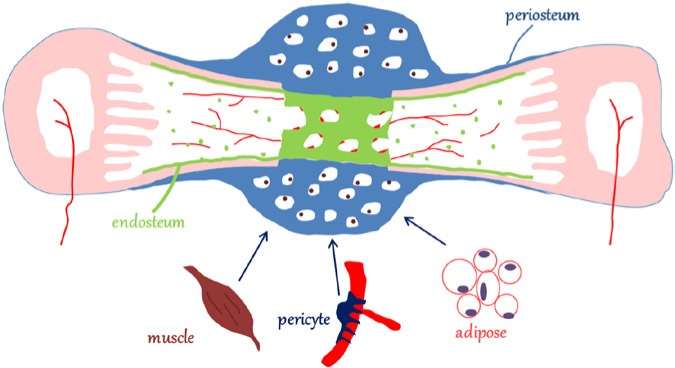
During the fracture healing, cells from the periosteum contribute significantly to callus formation. Cells from other sources, such as stem cells from muscle and adipose tissue, and pericytes from the surrounding blood vessels, are involved in callus formation as well. Cells derived from the endosteum and the bone marrow are primarily responsible for the regeneration of the new endosteum and bone marrow. Endosteal and bone marrow cells do not participate in callus formation (adapted from Colnot et al., 2012).
Interestingly, the size and the cellular content of the callus depend on how much the periosteum is stimulated during healing. For example, when the fracture ends are stabilized with fixation, callus formation is relatively minimal, and bone regeneration occurs by means of intramembranous ossification. However, in the absence of fixation and stabilization, a larger callus is formed by means of endochondral ossification, which occurs as the result of a stronger periosteal response (Thompson et al., 2002; Colnot et al., 2003).
Other sources of mesenchymal progenitor cells have also been proposed to participate in bone-fracture healing, including local bone marrow, adjacent muscle tissue, endothelial cells, and circulating mesenchymal stem cells (Medici et al., 2010; Colnot et al., 2012; Park et al., 2012) (Fig. 2). However, the relative contribution to healing by each cellular phenotype is still unclear. Although cells of endothelial origin have been shown to transdifferentiate into osteoblasts and chondrocytes during heterotopic ossification (Medici et al., 2010), they were sparsely found in the cartilage and bone in the fracture healing site when lineage tracing analysis was used to track the contributions of these cells to fracture repair (Lu et al., 2006). Pericytes, however, are heavily involved in fracture healing. Grcevic et al. (2012) showed that the majority of the callus cells, including chondrocytes and osteoblasts, are derived from SMA9 (Smooth Muscle Actin 9)-expressing cells. Although the origin of SMA9+ cells is still unknown, they are most likely cells derived from the local vascular system adjacent to the fracture site (Grcevic et al., 2012).
Molecular Mechanisms Underlying the Effects of Periosteal Cells in Bone Healing
Until recently, little was known about the molecular signaling that controls the periosteal responses during callus formation. With the development of genetically modified mouse models, studies have probed several signaling pathways and their effects on periosteal cells. BMP, FGF, Hedgehog, Notch, PDGF, Wnt, and inflammatory mediators have all been shown to contribute to the healing process, and therapies that target these pathways are currently under development for bone metabolic diseases such as osteoporosis. Herein, we provide a background of information on the involvement of these pathways in fracture repair, with the ultimate goal of informing future periosteal-targeted therapies for intra-oral bone augmentation procedures.
BMP Signaling
Bone morphogenetic protein-2 (BMP2) is well-known for its powerful effect on bone development and regeneration (Lowery et al., 2011). It has been suspected for years that BMP2 is an intrinsic initiator of bone healing. However, studies to support this theory were limited, since global BMP2-deletion results in embryonic lethality (Lowery et al., 2011). Tsuji et al. (2006) developed a mouse model in which the Bmp2 gene is conditionally inactivated in the mesoderm by means of a Prx-1-Cre deleter mouse. A delayed periosteal activation response and, most strikingly, a complete absence of bridging callus upon fracture were found in these Bmp2 conditional knockout mice, underscoring the important role of BMP2 in the initiation of fracture healing. Similarly, Wang et al. (2011) deleted Bmp2 post-natally at the initiation stage of fracture healing using a Tamoxifen-inducible CreER mouse model and found that Bmp2 inactivation at the onset of healing abolished the bone and cartilage callus formation derived from the periosteum. Further analysis of the periosteal callus showed that most of the Bmp2 conditional knockout progenitors remained undifferentiated (Wang et al., 2011). Recently, Mi et al. (2013) conditionally knocked out the Bmp2 gene in chondrocytes and in osteoblasts by means of Col2-Cre and Col1-Cre deleter mice. They showed that the lack of Bmp2 expression in chondrocytes led to prolonged cartilage callus formation and delayed osteogenesis initiation. However, when the Bmp2 gene was inactivated in osteoblasts by the Col1-Cre deleter, the mice showed no significant difference in fracture healing. These results indicate that, during fracture repair, expression of BMP2 is required in undifferentiated pre-osteoblasts and not in mature osteoblasts.
Interestingly, the skeletal-specific inactivation of Bmp4 and Bmp7 by means of a Prx1-Cre deleter shows no effects on fracture repair (Tsuji et al., 2006), suggesting that, in vivo, under physiological conditions, BMP2, not BMP4 or BMP7, represents a primary modulator of periosteum-mediated fracture repair. Although these results do not exclude local artificial delivery of BMP4 and BMP7 as still promoting fracture healing, they indicate that the use of recombinant BMP2 in bone augmentation procedures may require the presence of the periosteum to achieve maximal therapeutic efficiency. Other bone-regenerative therapies that may target BMP signaling should also take into account the requirement of the periosteum for maximum efficiency.
FGF Signaling
The role of FGFs and their receptors in fracture healing has been extensively investigated over the years (Du et al., 2012). Fgf2, 5, and 6 are markedly up-regulated during early callus formation and maintain high levels of expression throughout the healing process (Du et al., 2012). Other FGFs, such as Fgf16 and Fgf18, reached their peak expression at the chondrogenesis stage of callus formation (Du et al., 2012), whereas FGF receptors such as Fgfr1 and 2 were shown to be expressed in the proliferating periosteal mesenchyme (Du et al., 2012).
Many studies have explored the use of FGFs, especially FGF2, for promoting fracture healing. In the mouse, rat, rabbit, and non-human primates, FGF2 has been found to stimulate the proliferation of periosteal cells, osteoprogenitors, and chondrogenitors, enhancing callus formation (Du et al., 2012). However, some studies showed that FGF2 treatment did not lead to increased bone mineral density and increased mechanical strength of the callus (Nakajima et al., 2007). To explain this discrepancy, additional studies suggested that the effect of FGF on bone formation is biphasic, with inhibitory effects at high doses (Du et al., 2012). Recently, a randomized, placebo-controlled clinical trial in Japan demonstrated that local application of rhFGF2 hydrogel accelerates healing of tibial shaft fracture, with a higher percentage of radiographic bone union (Kawaguchi et al., 2012).
These studies indicate that a precise, time-controlled modulation of FGF signaling may be useful in bone-regenerative procedures, including those of the oral cavity.
Hedgehog Signaling
The Indian hedgehog (Ihh) pathway has been known to be a master regulator in chondrocyte differentiation, proliferation, and maturation, especially during endochondral ossification (Lai and Mitchell, 2005). Loss of hedgehog signaling in post-natal chondrocytes, either genetically or pharmacologically, leads to loss of epiphyseal growth plates, with premature fusion and shortened stature. However, Ihh also plays critical roles in periosteum-mediated fracture healing. For instance, it has been shown that Ihh is highly expressed during callus formation of both non-stabilized and stabilized fractures (Le et al., 2001). More specifically, Ihh appears to be expressed within the nascent cartilaginous tissues, in proximity to the bone surface of the callus (Wang et al., 2010). Smoothened (Smo) is a transmembrane protein that transduces all Hh signals. By means of a tamoxifen-inducible cre-mediated gene inactivation, Wang et al. (2010) showed that deletion of Smo at the onset of bone autograft repair resulted in a nearly 50% reduction in periosteal bone callus formation at the cortical bone junction. From the healing periosteum, they also isolated a population of early periosteum-callus-derived mesenchymal stem cells; gene knockout of Smo markedly reduced osteogenic differentiation of these progenitor cells (Wang et al., 2010). Thus, targeting the hedgehog pathway may represent an approach to bone anabolism as well as an approach to periosteal-mediated bone regeneration.
Notch Signaling
Notch signaling has been shown to regulate skeletal development (Tao et al., 2010). Gain-of-function mutation in the mesenchyme leads to enhanced proliferation and decreased differentiation, inhibiting the maturation of endochondral and intramembranous bone formation. In contrast, disruption of Notch signaling markedly increased osteoprogenitor differentiation and trabecular bone mass (Tao et al., 2010). However, this phenotype diminished rapidly during aging, due to depletion of stem cell pools in bone marrow, indicating that Notch may be implicated in the maintenance of progenitor cells within their niche. Notch signaling is up-regulated during bone fracture healing as Jag1 and Notch2 expression is localized in the undifferentiated mesenchymal cells of the healing periosteum (Dishowitz et al., 2012). Interestingly, it has recently been shown that expression of Jag1 is sufficient to induce osteogenic differentiation of human mesenchymal stem cells (Dishowitz et al., 2013). Thus, targeting Notch signaling by up-regulating Jag1 expression may represent a potential bone-/periosteal-regenerative therapy.
PDGF Signaling
Platelet-derived growth factors (PDGFs) are involved in the wound-healing processes of various tissues, including bone. PDGF ligands and receptors have been shown to be expressed in osteoblasts, chondrocytes, and mesenchymal stem cells (Kaigler et al., 2011). Deletion of PDGF receptors α and β in mice showed that the primary effects of PDGF signaling in bone healing are proliferation and migration responses (Tokunaga et al., 2008). However, to the best of our knowledge, studies that directly evaluate the role of PDGFs in periosteal activation do not exist. Although recombinant PDGF-BB is currently utilized for periodontal regenerative therapy (Kaigler et al., 2011), whether or not it can be utilized for periosteal-mediated bone regeneration has yet to be proven.
Wnt Signaling
Many members of the Wnt family are expressed in the injured periosteum (Chen et al., 2007; Kim et al., 2007). It was observed that fracture repair was inhibited in mice expressing osteoblast-specific beta-catenin null alleles (Chen et al., 2007). When the Wnt/β-catenin signaling pathway was inactivated in chondrocytes by interruption of the β-catenin/TCF complex formation, bone repair was also compromised due to reduced and delayed cartilage formation (Huang et al., 2012). Inhibition of Wnt signaling in bone healing through adenoviral expression of Dkk1, a Wnt inhibitor, reduced bone regeneration by 84% (Kim et al., 2007). Further studies showed that the addition of recombinant DKK1 or Sclerostin to primary periosteal cells reduces proliferation and increases apoptosis (Kim et al., 2013). Therefore, Wnt signaling contributes significantly to fracture healing. In confirmation of these observations, gain-of-function of Wnt signaling seems to promote bone fracture healing. In fact, expression of an activated form of beta-catenin in osteoblasts dramatically enhanced bone healing in mice (Chen et al., 2007). Additionally, gene knockout of Axin2, a negative regulator of Wnt signaling, resulted in accelerated bone healing after injury, which was related to more robust proliferation and earlier differentiation of skeletal stem and progenitor cells (Minear et al., 2010). Wnt3a injection into the periosteum induced faster bone regeneration by stimulating the proliferation of skeletal progenitor cells and accelerating their differentiation into osteoblasts (Minear et al., 2010). Also, the administration of DKK1 neutralizing antibody in bone healing improved bone repair and regeneration in mice (Agholme et al., 2011; Li et al., 2011), and oral administration of the potent GSK3 inhibitor AZD2858, which activates Wnt signaling, led to rapid fracture healing (Sisask et al., 2013). In vivo deletion of the secreted Frizzled Related Protein 1 (sFRP1), one of several frizzled related Wnt antagonists, improves fracture repair by promoting early bone union (Gaur et al., 2009).
However, studies have shown that Wnt-responsive cells appear to be located mainly on the endosteal surface of injured bone, in contrast to the BMP target cells resident in the periosteum (Minear et al., 2010), raising the possibility that, during fracture repair, BMP signaling and Wnt signaling act in different compartments within bone. Thus, a Wnt-based therapy may not directly target the perisoteum but may indirectly enhance periosteal-mediated bone regeneration. It is therefore possible that therapies under development that target the Wnt pathway, such as therapies based on DKK1 and SOST neutralizing antibodies, may be beneficial not only for bone endocrinology disorders but for periosteal-mediated bone-regenerative therapy as well.
Inflammatory Mediators
Since inflammatory phase and callus formation are well-coordinated during fracture healing, it is reasonable to speculate that inflammatory signals regulate periosteal response. Indeed, matrix metalloproteinase-9 (Mmp9), strongly expressed by neutrophils and macrophages in the early fracture callus, is involved in bone healing. Mmp9−/− mice have non-unions and delayed unions of their fractures caused by persistent cartilage at the injury site (Colnot et al., 2003). It has been further demonstrated that MMP9 mediates its effects on skeletal cell differentiation by regulating the inflammatory response and the distribution of inflammatory cells, leading to the local regulation of periosteal cell differentiation (Wang et al., 2013).
COX-2 is the inducible isoform of cyclooxygenase, a rate-limiting enzyme in the prostaglandin biosynthesis pathway. It is therefore a critical regulator in inflammation. By in situ hybridization, Cox-2 was found in chondroprogenitors and mesenchymal cells along the periosteal surface, and its expression was correlated with the early induction of chondrogenesis and early expansion of the cartilaginous callus (Xie et al., 2009). Administration of a COX-2 inhibitor in the early phase of healing compromised fracture repair (Simon et al., 2002). Furthermore, genetic ablation of Cox-2 impaired periosteal progenitor cell proliferation and delayed subsequent endochondral and intramembranous repair, leading to reduction of bone formation, persistence of cartilaginous tissue, and high incidence of non-unions. Contrarily, overexpressing Cox-2 locally accelerated fracture healing and bone unions (Zhang et al., 2002).
Although further studies are needed, modulation of the inflammatory process may represent the new frontier in periosteal-mediated bone-regenerative therapy.
Periosteum in Craniofacial Bone Regeneration
Craniofacial Periosteum
Little is known about the craniofacial periosteum. Periosteum has been shown to vary among regions and change with age. For example, the human femoral neck has less cellular but more mineralized periosteum than femoral diaphysis (Allen and Burr, 2005). The craniofacial periosteum may have distinct biological characters compared with the periosteum from other locations. In pig models, Ochareon and Herring (2011) showed that the palate presents with a relatively thin periosteum that becomes thinner with age. Interestingly, it is the inner layer of the periosteum that tends to decrease in thickness, whereas the outer (fibrous) layer barely changes its structure (Ochareon and Herring, 2011). Cells of the craniofacial periosteum appear to act differently when compared with cells of long-bone periosteum. Utilizing a cell-labeling strategy to mark neural crest–derived cells and mesoderm-derived cells, Leucht et al. (2008) investigated the osteogenic potential of periosteal cells obtained from the mandible and tibia. When mandibular periosteal cells were transplanted into a tibial bony defect, intramembranous ossification was seen. However, if tibial periosteal cells were transplanted into a mandibular defect, endochondral ossification was noted. It is therefore possible that the signaling pathways involved in the periosteal-mediated regeneration of the long bone may differ from those involved in craniofacial periosteal-mediated regeneration. However, regardless of the signaling pathways, given its richness in skeletal progenitors, preservation and harnessing of the periosteum may foster bone regeneration in both craniofacial and orthopedic surgical procedures.
Clinical Applications
Harnessing of the periosteum for craniofacial bone-regenerative procedures is not new. For instance, almost half a century ago, the periosteal flap was introduced by Skoog for closure of maxillary cleft defects (Skoog, 1965), and in 1989 the tibial periosteum was successfully utilized in cleft palate procedures (Rintala and Ranta, 1989).
Over the years, periodontists and oral surgeons have developed several clinical procedures to take advantage of the bone-regenerative potentials of the intra-oral periosteum. For instance, in a study to evaluate the post-operative fate of bone regeneration in maxillary sinusotomy, Choi et al. (1996) found that procedures performed with periosteal pedicle flaps resulted in more bone formation than those without periosteal pedicle flaps. Periosteal tissue has been used in periodontal-regenerative procedures as a membrane for guided tissue regeneration as well. Lekovic et al. (1991) first reported that a connective tissue graft comprehensive of the periosteum can be used as a barrier to enhance new attachment and osseous regeneration in Class II furcation defects of mandible molars. In a subsequent study, Lekovic et al. (1998) further compared the clinical effectiveness of the periosteal graft with that of the coronally positioned flap in the treatment of Class II furcation defects. The periosteal barrier group presented with a significantly better gain in vertical and horizontal bone regeneration. Recently, Verma et al. (2011) reported similar outcomes in the regeneration of mandibular buccal Class II furcations with a rotated mucoperiosteal flap. Similarly, periosteal grafts were successfully used as a membrane for bone regeneration after periradicular surgery (Tobon-Arroyave et al., 2004).
In the past 3 yrs, several techniques aimed at harnessing the intra-oral periosteum for alveolar bone augmentation have been proposed and described in detail.
Hur et al. (2010) introduced a double-flap incision design for guided bone regeneration (GBR) in the posterior mandible. It consists of 2 layers, a superficial mucosal layer and a deep periosteal layer. They recommend separating the mucosal layer first by dissecting in a partial thickness manner; the periosteum layer is reflected subsequently (Fig. 3). Although the primary objective of this technique is to obtain a tension-free primary closure, based on the evidence provided above, one could say that covering the surgical area with an intact periosteum provides undisturbed blood supply and osteoprogenitor cells to the regeneration site.
Figure 3.

Double-flap incision design, consisting of 2 layers, a superficial mucosal layer and a deep periosteal layer. The authors recommend separating the mucosal layer first by dissecting the tissue in a partial thickness manner; the periosteum layer is reflected subsequently. (A) Crestal incision on the edentulous ridge and one vertical releasing incision are outlined. (B) The double-flap incision design is made, leaving the periosteum on the edentulous ridge. (C, D) The mucosal layer of the double flap is elevated, leaving the periosteal layer. (E, F) The periosteal layer of the double flap is elevated, exposing the alveolar bone. (G) Occlusal view of the double flap. (H) The periosteal layer of the double flap is sutured to stabilize the grafted site. (I) Buccal view after final suturing (from Hur et al., 2010; with permission from the American Academy of Periodontology).
A vascularized interpositional periosteal-connective tissue (VIP-CT) flap that allows for the rotation of a vascularized periosteal pedicle to the anterior maxillary was first proposed by Sclar (2003) to take advantage of the periosteum and its associated blood supply in procedures aimed at reconstructing ridge defects of the maxilla. In 2012, this flap design was slightly modified by Kim et al. (2012) to combine a papilla preservation technique with the periosteal-connective tissue pedicle flap and achieve primary closure as well as bone and soft tissue augmentation (Fig. 4).
Figure 4.

Modified vascularized interpositional periosteal-connective tissue technique. (A, B) The flap is elevated by a papilla preservation technique. (C) The palatal donor site preparation begins by extending the incision horizontally to the first molar, and a subepithelial split-thickness dissection is carried anteriorly toward the palatal area of the missing site. (D) A vertical incision is made internally through the connective tissue and periosteum distal to the palatal incision, and a pediculated graft is elevated carefully from the bone, exposing the periosteum. Then, the connective tissue pedicle is rotated over the surgical site (from Kim et al., 2012; with permission).
A group of investigators from the University of Michigan (Steigmann et al., 2012) recently proposed a periosteal pocket flap for horizontal bone augmentation. In their novel flap design, the first incision is made at the most coronal aspect of the bone crest at a 45-degree angle to the buccal wall. Subsequently, a split-thickness flap on the buccal side separates the mucosa from the underlying periosteum. Vertical releasing is performed only within the mucosa, on both sides of the flap. A periosteal second incision follows, to create the periosteal pocket flap. This flap design generates a periosteal pocket between the buccal bone and the periosteum which can be filled with a bone-grafting material, covered by a resorbable collagen membrane, and closed by the mucosal flap generated with the first incision (Fig. 5).
Figure 5.
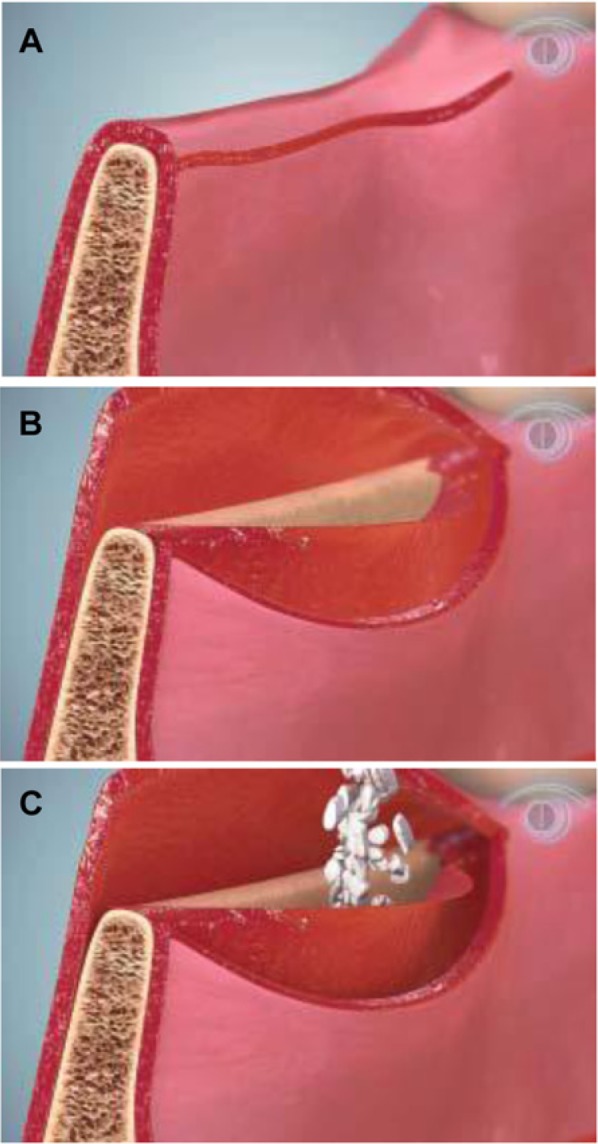
Periosteal pocket flap. (A) The first incision is made at the most coronal aspect of the bone crest at a 45-degree angle to the buccal wall. (B) Subsequently, a split-thickness flap on the buccal side separates the mucosa from the underlying periosteum. Vertical releasing is performed only within the mucosa, on both sides of the flap. A periosteal second incision follows, to create the periosteal pocket flap. (C) Bone-grafting material is inserted into the pocket flap to foster bone regeneration (from Steigmann et al., 2012; with permission).
Abrahamsson et al. (2012) proposed the use of self-inflatable osmotic expanders to be implanted under the periosteum before guided bone-regenerative procedures are performed. By this approach, they gain soft-tissue expansion as well as expansion of the periosteum, with obvious advantages to the subsequent surgical bone augmentation procedures.
Verdugo et al. (2012) showed that a successful GBR of large bone defects can be achieved by means of cortical perforation and no use of membranes but the periosteum alone. The protocol includes the elevation of a full-thickness mucoperiosteal flap by means of a crestal incision and two vertical releasing incisions, followed by decortication and fixation of autologous bone block grafts. The carefully preserved periosteal membrane is then used to fully cover the autografts, providing stem cells, blood supply, and nutrients to the bone regeneration site.
The reported clinical procedures have all been shown to be effective in bone-regenerative procedures. However, because of the limited number of cases reported and the lack of well-controlled clinical trials, further studies are needed to evaluate the effectiveness of these periosteal-harnessing approaches.
Conclusions
In this review, we analyzed the biology of the periosteum and its potential for bone-regenerative procedures. We showed how numerous studies have identified the existence of stem cells within the periosteum and how their presence and activation are fundamental to bone regeneration. Subsequently, we reviewed the molecular signaling that controls the periosteal-mediated bone regeneration and showed how the BMP, FGF, Hedgehog, Notch, PDGF, Wnt, and inflammatory pathways are involved in this process. Finally, we explored in detail the surgical techniques proposed to harness the periosteum in intra-oral bone-regenerative procedures.
Therefore, this review emphasized the importance of the preservation of the periosteum in bone augmentation procedures and proposes that, whenever possible, periosteal harnessing should be considered for the achievement of better therapeutic effects. As new molecular therapies that target the reviewed pathways are emerging for treatment of bone metabolic diseases such as osteoporosis, it would be reasonable to propose the use of these therapies to target the osteogenic potential of the perisoteum during oral regenerative procedures.
Footnotes
Dr. Giuseppe Intini is supported by a grant from the US National Institutes of Health (National Institute of Dental and Craniofacial Research, Grant # DE021069).
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
References
- Abrahamsson P, Walivaara DA, Isaksson S, Andersson G. (2012). Periosteal expansion before local bone reconstruction using a new technique for measuring soft tissue profile stability: a clinical study. J Oral Maxillofac Surg 70:e521-e530. [DOI] [PubMed] [Google Scholar]
- Agholme F, Isaksson H, Kuhstoss S, Aspenberg P. (2011). The effects of Dickkopf-1 antibody on metaphyseal bone and implant fixation under different loading conditions. Bone 48:988-996. [DOI] [PubMed] [Google Scholar]
- Allen MR, Burr DB. (2005). Human femoral neck has less cellular periosteum, and more mineralized periosteum, than femoral diaphyseal bone. Bone 36:311-316. [DOI] [PubMed] [Google Scholar]
- Allen MR, Hock JM, Burr DB. (2004). Periosteum: biology, regulation, and response to osteoporosis therapies. Bone 35:1003-1012. [DOI] [PubMed] [Google Scholar]
- Blaisdell FE. (1925). The osteogenic function of periosteum. Arch Surg 11:933. [Google Scholar]
- Chang H, Knothe Tate ML. (2012). Concise review: the periosteum: tapping into a reservoir of clinically useful progenitor cells. Stem Cells Transl Med 1:480-491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y, Whetstone HC, Lin AC, Nadesan P, Wei Q, Poon R, et al. (2007). Beta-catenin signaling plays a disparate role in different phases of fracture repair: implications for therapy to improve bone healing. PLoS Med 4:e249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi BH, Yoo JH, Sung KJ. (1996). Radiographic comparison of osseous healing after maxillary sinusotomy performed with and without a periosteal pedicle. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 82:375-378. [DOI] [PubMed] [Google Scholar]
- Colnot C. (2009). Skeletal cell fate decisions within periosteum and bone marrow during bone regeneration. J Bone Miner Res 24:274-282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colnot C, Thompson Z, Miclau T, Werb Z, Helms JA. (2003). Altered fracture repair in the absence of MMP9. Development 130:4123-4133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colnot C, Zhang X, Knothe Tate ML. (2012). Current insights on the regenerative potential of the periosteum: molecular, cellular, and endogenous engineering approaches. J Orthop Res 30:1869-78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diaz-Flores L, Gutierrez R, Lopez-Alonso A, Gonzalez R, Varela H. (1992). Pericytes as a supplementary source of osteoblasts in periosteal osteogenesis. Clin Orthop Relat Res 275:280-286. [PubMed] [Google Scholar]
- Dimitriou R, Jones E, McGonagle D, Giannoudis PV. (2011). Bone regeneration: current concepts and future directions. BMC Med 9:66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dishowitz MI, Terkhorn SP, Bostic SA, Hankenson KD. (2012). Notch signaling components are upregulated during both endochondral and intramembranous bone regeneration. J Orthop Res 30:296-303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dishowitz MI, Zhu F, Sundararaghavan HG, Ifkovits JL, Burdick JA, Hankenson KD. (2013). Jagged1 immobilization to an osteoconductive polymer activates the notch signaling pathway and induces osteogenesis. J Biomed Mater Res A [Epub ahead of print 6/14/2013] [in press]. [DOI] [PubMed] [Google Scholar]
- Du X, Xie Y, Xian CJ, Chen L. (2012). Role of FGFs/FGFRs in skeletal development and bone regeneration. J Cell Physiol 227:3731-3743. [DOI] [PubMed] [Google Scholar]
- Gaur T, Wixted JJ, Hussain S, O’Connell SL, Morgan EF, Ayers DC, et al. (2009). Secreted frizzled related protein 1 is a target to improve fracture healing. J Cell Physiol 220:174-181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grcevic D, Pejda S, Matthews BG, Repic D, Wang L, Li H, et al. (2012). In vivo fate mapping identifies mesenchymal progenitor cells. Stem Cells 30:187-196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang Y, Zhang X, Du K, Yang F, Shi Y, Huang J, et al. (2012). Inhibition of beta-catenin signaling in chondrocytes induces delayed fracture healing in mice. J Orthop Res 30:304-310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hur Y, Tsukiyama T, Yoon TH, Griffin T. (2010). Double flap incision design for guided bone regeneration: a novel technique and clinical considerations. J Periodontol 81:945-952. [DOI] [PubMed] [Google Scholar]
- Kaigler D, Avila G, Wisner-Lynch L, Nevins ML, Nevins M, Rasperini G, et al. (2011). Platelet-derived growth factor applications in periodontal and peri-implant bone regeneration. Expert Opin Biol Ther 11:375-385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawaguchi H, Oka H, Jingushi S, Izumi T, Fukunaga M, Sato K, et al. (2012). A local application of recombinant human fibroblast growth factor 2 for tibial shaft fractures: a randomized, placebo-controlled trial. J Bone Miner Res 25:2735-2743. [DOI] [PubMed] [Google Scholar]
- Kim CS, Jang YJ, Choi SH, Cho KS. (2012). Long-term results from soft and hard tissue augmentation by a modified vascularized interpositional periosteal-connective tissue technique in the maxillary anterior region. J Oral Maxillofac Surg 70:484-491. [DOI] [PubMed] [Google Scholar]
- Kim HK, Oxendine I, Kamiya N. (2013). High-concentration of BMP2 reduces cell proliferation and increases apoptosis via DKK1 and SOST in human primary periosteal cells. Bone 54:141-150. [DOI] [PubMed] [Google Scholar]
- Kim JB, Leucht P, Lam K, Luppen C, Ten Berge D, Nusse R, et al. (2007). Bone regeneration is regulated by wnt signaling. J Bone Miner Res 22:1913-1923. [DOI] [PubMed] [Google Scholar]
- Lai LP, Mitchell J. (2005). Indian hedgehog: its roles and regulation in endochondral bone development. J Cell Biochem 96:1163-1173. [DOI] [PubMed] [Google Scholar]
- Le AX, Miclau T, Hu D, Helms JA. (2001). Molecular aspects of healing in stabilized and non-stabilized fractures. J Orthop Res 19:78-84. [DOI] [PubMed] [Google Scholar]
- Lekovic V, Kenney EB, Carranza FA, Martignoni M. (1991). The use of autogenous periosteal grafts as barriers for the treatment of Class II furcation involvements in lower molars. J Periodontol 62:775-780. [DOI] [PubMed] [Google Scholar]
- Lekovic V, Klokkevold PR, Camargo PM, Kenney EB, Nedic M, Weinlaender M. (1998). Evaluation of periosteal membranes and coronally positioned flaps in the treatment of Class II furcation defects: a comparative clinical study in humans. J Periodontol 69:1050-1055. [DOI] [PubMed] [Google Scholar]
- Leucht P, Kim JB, Amasha R, James AW, Girod S, Helms JA. (2008). Embryonic origin and Hox status determine progenitor cell fate during adult bone regeneration. Development 135:2845-2854. [DOI] [PubMed] [Google Scholar]
- Li X, Grisanti M, Fan W, Asuncion FJ, Tan HL, Dwyer D, et al. (2011). Dickkopf-1 regulates bone formation in young growing rodents and upon traumatic injury. J Bone Miner Res 26:2610-2621. [DOI] [PubMed] [Google Scholar]
- Lowery JW, Pazin D, Intini G, Kokabu S, Chappuis V, Capelo LP, et al. (2011). The role of BMP2 signaling in the skeleton. Crit Rev Eukaryot Gene Expr 21:177-185. [DOI] [PubMed] [Google Scholar]
- Lu C, Miclau T, Hu D, Hansen E, Tsui K, Puttlitz C, et al. (2005). Cellular basis for age-related changes in fracture repair. J Orthop Res 23:1300-1307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu C, Marcucio R, Miclau T. (2006). Assessing angiogenesis during fracture healing. Iowa Orthop J 26:17-26. [PMC free article] [PubMed] [Google Scholar]
- Medici D, Shore EM, Lounev VY, Kaplan FS, Kalluri R, Olsen BR. (2010). Conversion of vascular endothelial cells into multipotent stem-like cells. Nat Med 16:1400-1406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mi M, Jin H, Wang B, Yukata K, Sheu TJ, Ke QH, et al. (2013). Chondrocyte BMP2 signaling plays an essential role in bone fracture healing. Gene 512:211-218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minear S, Leucht P, Jiang J, Liu B, Zeng A, Fuerer C, et al. (2010). Wnt proteins promote bone regeneration. Sci Transl Med 2:29ra30. [DOI] [PubMed] [Google Scholar]
- Nakajima F, Nakajima A, Ogasawara A, Moriya H, Yamazaki M. (2007). Effects of a single percutaneous injection of basic fibroblast growth factor on the healing of a closed femoral shaft fracture in the rat. Calcif Tissue Int 81:132-138. [DOI] [PubMed] [Google Scholar]
- Ochareon P, Herring SW. (2011). Cell replication in craniofacial periosteum: appositional vs. resorptive sites. J Anat 218:285-297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park D, Spencer JA, Koh BI, Kobayashi T, Fujisaki J, Clemens TL, et al. (2012). Endogenous bone marrow MSCs are dynamic, fate-restricted participants in bone maintenance and regeneration. Cell Stem Cell 10:259-272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reilly TM, Seldes R, Luchetti W, Brighton CT. (1998). Similarities in the phenotypic expression of pericytes and bone cells. Clin Orthop Relat Res 346:95-103. [PubMed] [Google Scholar]
- Rintala AE, Ranta R. (1989). Periosteal flaps and grafts in primary cleft repair: a follow-up study. Plast Reconstr Surg 83:17-24. [DOI] [PubMed] [Google Scholar]
- Sclar AG. (2003). Vascularized interpositional periosteal-connective tissue (VIP-CT) flap. In: Soft tissue and esthetic considerations in implant therapy. Chicago, IL: Quintessence Publishing Co., pp. 163-187. [Google Scholar]
- Simon AM, Manigrasso MB, O’Connor JP. (2002). Cyclo-oxygenase 2 function is essential for bone fracture healing. J Bone Miner Res 17:963-976. [DOI] [PubMed] [Google Scholar]
- Sisask G, Marsell R, Sundgren-Andersson A, Larsson S, Nilsson O, Ljunggren O, et al. (2013). Rats treated with AZD2858, a GSK3 inhibitor, heal fractures rapidly without endochondral bone formation. Bone 54:126-132. [DOI] [PubMed] [Google Scholar]
- Skoog T. (1965). The use of periosteal flaps in the repair of clefts of the primary palate. Cleft Palate J 2:332-339. [PubMed] [Google Scholar]
- Steigmann M, Salama M, Wang HL. (2012). Periosteal pocket flap for horizontal bone regeneration: a case series. Int J Periodontics Restorative Dent 32:311-320. [PubMed] [Google Scholar]
- Tao J, Chen S, Lee B. (2010). Alteration of Notch signaling in skeletal development and disease. Ann NY Acad Sci 1192:257-268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson Z, Miclau T, Hu D, Helms JA. (2002). A model for intramembranous ossification during fracture healing. J Orthop Res 20:1091-1098. [DOI] [PubMed] [Google Scholar]
- Tobon-Arroyave SI, Dominguez-Mejia JS, Florez-Moreno GA. (2004). Periosteal grafts as barriers in periradicular surgery: report of two cases. Int Endod J 37:632-642. [DOI] [PubMed] [Google Scholar]
- Tokunaga A, Oya T, Ishii Y, Motomura H, Nakamura C, Ishizawa S, et al. (2008). PDGF receptor beta is a potent regulator of mesenchymal stromal cell function. J Bone Miner Res 23:1519-1528. [DOI] [PubMed] [Google Scholar]
- Tsuji K, Bandyopadhyay A, Harfe BD, Cox K, Kakar S, Gerstenfeld L, et al. (2006). BMP2 activity, although dispensable for bone formation, is required for the initiation of fracture healing. Nat Genet 38:1424-1429. [DOI] [PubMed] [Google Scholar]
- Verdugo F, D’Addona A, Pontón J. (2012). Clinical, tomographic, and histological assessment of periosteal guided bone regeneration with cortical perforations in advanced human critical size defects. Clin Implant Dent Relat Res 14:112-120. [DOI] [PubMed] [Google Scholar]
- Verma V, Saimbi CS, Khan MA, Goel A. (2011). Use of periosteal membrane as a barrier membrane for the treatment of buccal Grade II furcation defects in lower molars: a novel technique. Indian J Dent Res 22:511-516. [DOI] [PubMed] [Google Scholar]
- Wang Q, Huang C, Zeng F, Xue M, Zhang X. (2010). Activation of the Hh pathway in periosteum-derived mesenchymal stem cells induces bone formation in vivo: implication for postnatal bone repair. Am J Pathol 177:3100-3111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Q, Huang C, Xue M, Zhang X. (2011). Expression of endogenous BMP-2 in periosteal progenitor cells is essential for bone healing. Bone 48:524-532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X, Yu YY, Lieu S, Yang F, Lang J, Lu C, et al. (2013). MMP9 regulates the cellular response to inflammation after skeletal injury. Bone 52:111-119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie C, Liang B, Xue M, Lin AS, Loiselle A, Schwarz EM, et al. (2009). Rescue of impaired fracture healing in COX-2-/- mice via activation of prostaglandin E2 receptor subtype 4. Am J Pathol 175:772-785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X, Schwarz EM, Young DA, Puzas JE, Rosier RN, O’Keefe RJ. (2002). Cyclooxygenase-2 regulates mesenchymal cell differentiation into the osteoblast lineage and is critically involved in bone repair. J Clin Invest 109:1405-1415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X, Xie C, Lin AS, Ito H, Awad H, Lieberman JR, et al. (2005). Periosteal progenitor cell fate in segmental cortical bone graft transplantations: implications for functional tissue engineering. J Bone Miner Res 20:2124-2137. [DOI] [PMC free article] [PubMed] [Google Scholar]