

Abstract
Background
Adolescents with conduct disorder (CD) and substance use disorders (SUD) experience difficulty evaluating and regulating their behavior in anticipation of future consequences. Given the role of the brain's default mode network (DMN) in self-reflection and future thought, this study investigates whether DMN is altered in adolescents with CD and SUD, relative to controls.
Methods
Twenty adolescent males with CD and SUD and 20 male controls of similar ages underwent functional magnetic resonance imaging as they completed a risk-taking decision task. We used independent component analysis as a data-driven approach to identify the DMN spatial component in individual subjects. DMN activity was then compared between groups.
Results
Compared to controls, patients showed reduced activity in superior, medial and middle frontal gyrus (Brodmann area (BA) 10), retrosplenial cortex (BA 30) and lingual gyrus (BA 18), and bilateral middle temporal gryus (BA 21/22) - DMN regions thought to support self-referential evaluation, memory, foresight, and perspective taking. Furthermore, this pattern of reduced activity in patients remained robust after adjusting for the effects of depression and attention-deficit hyperactivity disorder (ADHD). Conversely, when not adjusting for effects of depression and ADHD, patients demonstrated greater DMN activity than controls solely in the cuneus (BA 19).
Conclusions
Collectively, these results suggest that comorbid CD and SUD in adolescents is characterized by atypical activity in brain regions thought to play an important role in introspective processing. These functional imbalances in brain networks may provide further insight into the neural underpinnings of conduct and substance use disorders.
Keywords: Default Mode Network, Conduct Disorder, Substance Use Disorder, Independent Component Analysis, Functional MRI
1. INTRODUCTION
1.1 Conduct and substance use disorder vulnerability
Conduct disorder (CD) and substance use disorder (SUD; DSM-IV, APA 2000) are strongly comorbid (Disney et al., 1999) and prevalent in youth (Nock et al., 2006), with characteristics that reflect a failure of restraint and inhibition (Crowley et al., 2006, 2010b). That failure could result from an inability to engage effectively in introspective processing; this trait may be identifiable by evaluating brain activation in the default mode network (DMN; Whitfied-Gabrieli and Ford, 2012). Therefore, we studied male adolescents in treatment for CD and SUD, seeking functional abnormalities in regions within DMN.
CD and SUD are comorbid manifestations of an underlying liability known as behavioral disinhibition (Kendler et al., 2011) that is highly heritable (i.e., heritability =~0.8; Young et al., 2000). Adolescents with CD/SUD have functional and structural deficits in several critical regions of the decision-making network including those important for monitoring conflicting choices, emotional decision-making, and inhibition (Crowley et al., 2010a; Dalwani et al., 2011). In addition to impairments in executive function, youth with CD/SUD and adults with antisocial phenotype, demonstrate atypical self-reflective, and self-evaluative behavior (Sharp, 2008; Fonagy and Target, 1997). However, we find no functional MRI data on this vulnerable CD/SUD population evaluating the specific DMN.
Children with CD, who lack self-reflection, may experience difficulties in adjusting their behavior based on past outcomes (Delfos, 2004). Individuals who lack the ability to reflect on the negative consequences of immoral actions may become predisposed to rule-breaking antisocial behavior (Raine and Yang, 2006). Adolescents with CD lack in episodic memory (Fairchild et al., 2011), an attribute needed to guide future thoughtful behavior by remembering one's past actions (Schacter et al., 2012).
The antisocial behavior characteristic of CD suggests impairments in social cognition or “theory of mind” (i.e., thinking about others or what others are thinking; Sharp, 2008). Research suggests that theory of mind is an important contributor to the development of antisocial behavior as it is a prerequisite to empathic responding, which facilitates the inhibition of antisocial behavior (Happé and Frith, 1996; Sharp, 2006, 2008). In sum, lack of self-reflection, introspection, internal mentation, theory of mind, and episodic memory are attributes that may lead to lack of remorse or guilt, callousness, and antisocial behavior - traits highly characteristic of CD/SUD youth.
1.2 Role of the Default Mode Network (DMN)
The brain's DMN plays an important role in self-evaluative processing, social perspective-taking, episodic memory, internal mentation and future thought/foresight (Whitfield-Gabrieli and Ford, 2012; Andrews-Hanna, 2012). Though these processes can differentiate youths with CD/SUD from the general youth population, the DMN has yet to be explored in this patient population.
The DMN is a “functionally connected” network and is comprised of the following regions: medial prefrontal cortex (MPFC), medial and lateral parietal cortex, and temporal lobe (Greicius et al., 2003). DMN can be studied in fMRI paradigms that combine periods of rest and active stimuli (Sharp et al., 2011). Cognitively demanding tasks with short periods of rest or periods with low cognitive load, strongly recruit DMN, a high-level network, which shows increased blood oxygenated level dependent (BOLD) signal activation during periods of rest and reduced activation during periods of high cognitive demand (Buckner et al., 2008).
Relevant to SUD, atypical DMN has been reported in heroin-addicted adults (Ma et al., 2011), in prenatally cocaine-exposed adolescents (Li et al., 2011) and in alcoholic adults (Chanraud et al., 2011) in either resting-state functional MRI (fMRI), or in tasks needing cognitive involvement or during tasks with both. DMN alterations have been observed in individuals with major depression, attention-deficit hyperactivity disorder (ADHD), and other psychiatric disorders (Broyd et al., 2009), some of which are comorbid with CD/ SUD. DMN has a strong overlap with brain areas involved in social cognition (Andrews-Hanna, 2012; Schilbach et al., 2008). Tang et al., 2013 recently showed decreased functional connectivity between regions of DMN and attention networks in antisocial personality disorder (ASPD) adults. The same study using data-driven classifier based on machine learning showed that the DMN made a sizeable contribution in discriminating ASPD from control subjects.
By showing improvement in aberrant functional connectivity, changes in DMN may help establish efficacy for treatment of psychiatric disorders (Tregellas et al., 2011; Tanabe et al., 2011). The DMN is a robust non-invasive biomarker, and a potential phenotype for molecular genetic studies and for brain pathology (Biswal et al., 2010; Glahn et al., 2010), distinguishes patients from controls (Broyd et al., 2009), and detects neuropathophysiological diseases (e.g., Koch et al., 2012), all of which could be applied to CD/SUD.
1.3 Study Objectives
Given the relevance of the DMN for CD/SUD, we sought to evaluate DMN activity in male CD/SUD adolescents compared to non-affected controls on a rapid-event fMRI decision-outcome risk-taking paradigm. To identify the DMN, we used independent component analysis (ICA), a multivariate-based (i.e., all voxels analyzed at the same time) data-driven approach that separates independent components (brain networks) from a mixture (fMRI signal; McKeown et al., 1998). We predicted that DMN activity in male CD/SUD adolescents compared to controls would differ in the main DMN areas (medial prefrontal and parietal cortex, lateral parietal and temporal cortex).
2. METHODS
We utilized this dataset previously to examine patient-control differences in fMRI signal change during risky and cautious decisions and their consequences and showed widespread hypoactivation in male CD/SUD adolescents compared to controls in several critical regions of the decision-making network (see Crowley et al., 2010a).
2.1 Sample and inclusion/exclusion criteria
This study includes 20 patients (18 right-handed) and 20 controls (19 right-handed). All subjects were males, ages 14-18 years, with IQ≥80 as estimated from the Wechsler Abbreviated Scale of Intelligence (Wechsler, 1999). Inclusion criteria for controls were no CD nor SUD (DSM-IV, APA 2000) except nicotine, no court convictions, no substance related arrests or treatment or school-expulsions, and no obvious psychosis or physical illness. Inclusion criteria for patients were enrollment in our university-based treatment program for serious CD and SUD; presence of serious conduct problems including symptoms of CD (e.g., theft, weapon fights) and at least one non-nicotine SUD diagnosis. Exclusion criteria for patients included psychosis, current high risk of suicide, violence, or fire-setting.
Both patients and controls were excluded if they or their parents lacked sufficient English skills for assenting or consenting or if they had non-prescribed substances present in urine (see Supplementary Material Section S1.a†) or saliva tested about 7 days before, and immediately before, scanning. Other exclusion criteria include marked claustrophobia, orthodontic braces, color blindness, contraindications to MR scanning (embedded metal, pacemakers, cochlear implants, etc.), history of head injury with loss of consciousness for more than 15 minutes, history of significant neurological illness or neurosurgery or a serious general medical disorder.
2.2 Functional MRI paradigm: “Colorado Balloon Game”
While in the MRI, subjects played a rapid-event fMRI paradigm known as the “Colorado Balloon Game”, consisting of trials with an active choice (risky/cautious) called “decision trials” with outcomes (wins/loss if risky; minimal compensation if cautious) and trials where the game directs the subject's response called “directed trials”. The task included brief periods of rest and directed trials provided longer periods of minimal cognitive load (see Crowley et al., 2010a for detailed description of the task, its timing and payoff pattern).
2.3 Image acquisition
After training in a mock scanner, subjects performed the fMRI task in a 3T General Electric MRI scanner. The task consists of 3 sessions of 30 paired decision and directed trials. The echo-planar imaging (EPI) acquisition parameters were: TR=2000ms, TE=26ms, flip angle=70°, FOV=220mm2, 642 matrix, 36 slices, 4mm thick, no gap, angled parallel to the planum sphenoidale. More details on image acquisition are provided in Supplementary Material Section S2‡
2.4 ICA analyses
The EPI data were preprocessed using motion correction, co-registration to respective high-resolution structural image, normalization to the pediatric template in MNI space and smoothing with a 6mm full-width-half-maximum gaussian kernel. Group ICA (using GIFT; http://mialab.mrn.org/software/gift/index.html) was then conducted on the preprocessed data separately on each group (Calhoun et al., 2008). Using the infomax algorithm (Bell and Sejnowski, 1995), we derived independent spatial networks activated during the task e.g. decision network, DMN, etc. with their respective temporal information (time course). Each subject's DMN component was identified as the component with the highest spatial correlation to a standard DMN mask (Pickatlas; http://fmri.wfubmc.edu/software/PickAtlas). Previous studies have utilized similar procedures (e.g., Tregellas et al., 2011). Supplementary Material Section S3§ provides a detailed description of the ICA method.
2.5 Group comparisons and covariates
The z-score for every voxel was estimated by normalizing each voxel's intensity with respect to intensity of all the voxels in the DMN component. We use “DMN activity” to refer to the intensity (z-score) of the DMN signal. We first examined the pattern of DMN activation by conducting a whole-brain voxel-wise analysis using one-sample t-tests on the DMN component (spatial maps) for controls and patients separately. The DMN maps were compared between groups on a voxel-wise basis with a second-level ANCOVA in SPM5 adjusting for age and IQ. Statistical maps were set at a cluster-level threshold of p < 0.05 (97 voxels), corrected for multiple comparisons with family-wise error using AlphaSim Monte-Carlo simulations (Ward, 2000) and a voxel-level threshold of p<0.005. Inclusive masking of the main effect (p<0.05) was applied to ensure results reflected only DMN differences.
Rationale for using Age and IQ as covariates
Age-dependent effects on DMN have been demonstrated before (Pyka et al., 2009) and in large meta-analytic studies (Biswal et al., 2010; Allen et al., 2011). Also, given the age-related brain development in these adolescent years (14-18 years; Lenroot et al., 2007; Giedd et al., 2009) we co-varied age. Intelligence may be associated with strength of connectivity in DMN (Ming et al., 2009), hence we also controlled for IQ.
2.6 Steps to validate DMN
We validated whether we correctly identified the DMN component. First, given the event-related design, we verified that DMN was active during periods of rest by evaluating the pattern of temporal activation of our DMN component (see Supplementary Figure 3**). Second, parameterizing DMN time courses using multiple regression analyses we obtained association estimates (beta weights) between the DMN time course and the phases of the fMRI paradigm [i.e., risky decision, cautious decision, reward, loss, collect (after active decision), risky directed, cautious directed, reward, loss, collect (after directed instructions)]. These beta weights represented the degree of synchrony between DMN time course and the canonical hemodynamic response model, indicating whether or not the network represented in the component was engaged during that task phase (Supplementary Figure 4a and 4b††). Third, we used spectral analyses to demonstrate that our DMN component was activated at low frequency (note: typical signature of DMN activity represents low-frequency BOLD fMRI signal changes (0.012-0.1 Hz) in the resting brain, (see Supplementary Figure 5‡‡). Details of these analyses are included in Supplementary Material Section S4§§.
2.7 Examining patient DMN activity with abstinence and task performance
In exploratory analyses, we examined within patients the association between DMN activity with length of abstinence (days) from substances (see Supplementary Figure 6***) and separately the relationship between task performance (risk-taking, i.e., number of risky presses) and DMN activity using multiple regression analyses after adjusting for age and IQ (see Supplementary Figure 7 and Supplementary Material Section S5†††).
2.8 Secondary analyses
We conducted whole–brain analyses to examine group differences controlling for ADHD and depression. These disorders are more common in youths with CD/SUD than the general population (Crowley et al., 2001). We also separately examined three potential confounds by re-analyzing our data after excluding 3 left-handed subjects (2 patients, 1 control), and excluding 7 recent tobacco smokers (6 patients, 1 control), and excluding subjects taking prescribed medication (6 patients, 4 controls) around the time of scanning. Description of prescribed medications by subjects can be found in Supplemental Section S1.b. Similar to our primary analyses, secondary analyses were adjusted for age and IQ.
3. RESULTS
Demographics, and diagnostic patient-control sample comparisons are included in Table 1. As expected, patients differed significantly on various clinical measures including severity of ADHD, depression, IQ and SES.
Table 1.
A Comparison of characteristics of patients and controls
| Controls (n=20) | Patients (n=20) | test statistic | p-value | |
|---|---|---|---|---|
| Demographics: | ||||
| Age (mean(sd)) | 16.5 (1.6) | 16.5 (1.0) | t=0.003 | 0.99 |
| Race: | ||||
| Caucasian (n) | 15 | 12 | χ2 =1.03 | 0.31 |
| Non-Caucasian (n) | 5 | 8 | ||
| Education: | ||||
| Highest-grade comp. (mean(sd)) | 9.70 (1.69) | 9.15 (0.98) | M-W-U=158.5 | 0.25 |
| Ever repeated grade (n) | 1 | 6 | Fisher's Exact | 0.09 |
| Clinical Measures: | ||||
| IQ full-scale (sd) | 104.9 (9.0) | 97.3 (8.9) | t=2.67 | 0.01 |
| Carroll Rating Scale (sd) | 4.1 (3.8) | 8.5 (6.7) | M-W-U=106.5 | 0.01 |
| Attention Deficit Hyperactivity Problems t-scoreB | 53.5 (4.5) | 58.3 (8.2) | t=2.28 | 0.002 |
| Impulsivity, Aggression and Conduct Disorder: | ||||
| Eysenck Impulsivity Scale (sd) | 6.7 (4.5) | 11.9 (6.0) | t=6.84 | 0.004 |
| Aggression score | 0.5 (1.1) | 5.7 (3.2) | M-W-U=40.5 | <0.001 |
| Lifetime CD diagnosis (n) | 1 | 19 | χ2 =36.1 | <0.001 |
| Substance Use Disorders (SUD): | ||||
| SUD Symptoms Across Drugs (sd) | 0.2 (0.67) | 12.4 (7.2) | M-W-U=2.0 | <0.001 |
| Alcohol Abuse | 0 | 8 | Fisher's Exact | <0.004 |
| Alcohol Dependence | 8 | Fisher's Exact | <0.004 | |
| Amphetamine Dependence | 0 | 2 | Fisher's Exact | NS |
| Cannabis Abuse | 0 | 7 | Fisher's Exact | <0.009 |
| Cannabis Dependence | 0 | 10 | χ2 =13.33 | 0.0003 |
| Club Drugs Abuse | 0 | 3 | Fisher's Exact | NS |
| Club Drugs Dependence | 0 | 4 | Fisher's Exact | NS |
| Cocaine Abuse | 0 | 2 | Fisher's Exact | NS |
| Cocaine Dependence | 0 | 2 | Fisher's Exact | NS |
| Hallucinogen Abuse | 0 | 2 | Fisher's Exact | NS |
| Hallucinogen Dependence | 0 | 1 | Fisher's Exact | NS |
| Tobacco Dependence | 1 | 13 | χ2 =15.82 | <0.0001 |
Groups were compared based on appropriate statistical tests based on normality.
For attention deficit hyperactivity problem t-score missing Child Behavioral CheckList (Achenbach, 1991a) score for one patient was replaced by Youth Self Report score (Achenbach, 1991b)
IQ reported in Crowley et al. (2010) has two typos that have been addressed in this manuscript i) Patient IQ Instead of 97.1(9.3) is 97.3(8.9) and ii) the IQ full-scale t-score is IQ full-scale
Abbreviations:
sd=standard deviation; IQ=intelligence quotient measured by the two subtests from the Wechsler Abbreviated Scale of Intelligence;
Carroll Rating Scale score of >=10 is suggestive of clinical depression;
CD diagnosis=at least three lifetime DSM-IV conduct disorder symptoms; n=count (total number); sd=Standard Deviation; SUD=Substance Use Disorder;
t=t-value for the t-test;
χ2= Chi-square test (without the Yates continuity correction);
M-W-U=Mann-Whitney-U
3.1 Single group analyses on DMN
DMN activity was observed in both controls and patients in the [x,y,z] co-ordinates reported in Supplemental Table 1, Supplementary Figures 1 and 2‡‡‡). Group results were identical before and after running a bootstrapping procedure that runs group ICA repeatedly for fixed number of iterations (ICASSO, see Supplementary Material Section S3§§§), suggesting reliability in component selection (Average stability index for all the components: controls: mean±stdev=.977±.007; patients: mean±stddev =.978±.005).
3.2 Group comparison on DMN, adjusting for age and IQ
3.2.1 Contrast: controls > patients
Whole-brain voxel-wise group comparison showed significantly less activation in patients in frontal regions within the DMN, including bilateral superior, middle and medial frontal gyrus (BA10) and in other DMN regions such as bilateral middle temporal gyrus (BA21) and the retrosplenial cortex (BA30; Figure 1 and Table 2).
Figure 1. Control>Patient DMN Activity.
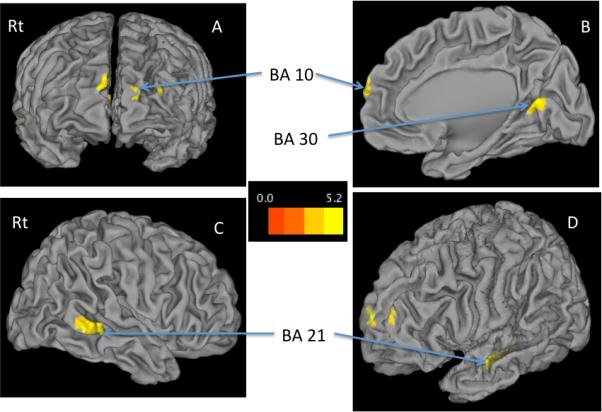
Statistical map set at a family-wise error corrected cluster-level threshold of p < 0.05 and overlaid on 3-D SPM template using CARET. A: Frontal View in Radiological Convention (Rt=Right); B: Right Medial Surface; C: Right Lateral Surface; D: Left Lateral Surface; BA= Brodmann Area; Controls showed more activity than patients in superior and middle frontal gyrus (BA10), retrosplenial cortex (BA30), lingual gyrus (BA18) and middle temporal gyrus (BA21,22)
Table 2.
| Controls>Patients: | Patients>Controls: | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Structure | BA or SideC | Cluster size | Maximum ActivationD | Structure | BA or SideC | Cluster size | Maximum ActivationD | ||||||
| x | y | z | t | x | y | z | t | ||||||
| Sup. Fr. Gy.E | R,L 10 | 191E | −10 | 68 | 14 | 5.4 | Cuneus | L 19 | 128 | −18 | −90 | 36 | 4.6 |
| Mid. Fr. Gy.E | L 10 | −30 | 60 | 12 | 2.8 | Cuneus | R 19 | 171 | 14 | −88 | 34 | 3.9 | |
| Med. Fr. Gy.E | L 10 | −6 | 66 | 10 | 2.9 | ||||||||
| Retrospl. CortexF | R,L 30 | 262F | −2 | −68 | 6 | 4.0 | |||||||
| Lingual Gy.F | R,L 18 | 8 | −64 | 4 | 4.2 | ||||||||
| Mid.Temp. Gy.G | R 21 | 178G | 58 | −46 | −2 | 6.0 | |||||||
| Mid. Temp. Gy.G | R 22 | 66 | −46 | 4 | 3.8 | ||||||||
| Mid. Temp. Gy. | L 21 | 129 | −54 | 6 | −18 | 5.1 | |||||||
| Total Activated Voxels | 760 | 299 | |||||||||||
Procedure for determining significance: whole brain cluster-wise family-wise error correction (pcorr<0.05)
Analyses procedure examined: ANCOVA (adjusted for age and IQ)
If bilateral, the larger maximum is shown
Montreal Neurological Institute coordinates, mm from anterior commissure
Regions bearing the same superscript comprise one activated cluster
Regions bearing the same superscript comprise one activated cluster
Regions bearing the same superscript comprise one activated cluster
Abbreviations: BA: Brodmann Area; Fr: Frontal; Gy: Gyrus; Med: Medial; Mid: Middle; Retrospl: Retrosplenial; Sup: Superior; Temp: Temporal
3.2.2 Contrast: patients > controls
As shown in Table 2 and Figure 2, patients showed significantly increased activation in the bilateral cuneus near BA19, extending into the posterior precuneus.
Figure 2. Patient>Control DMN Activity.
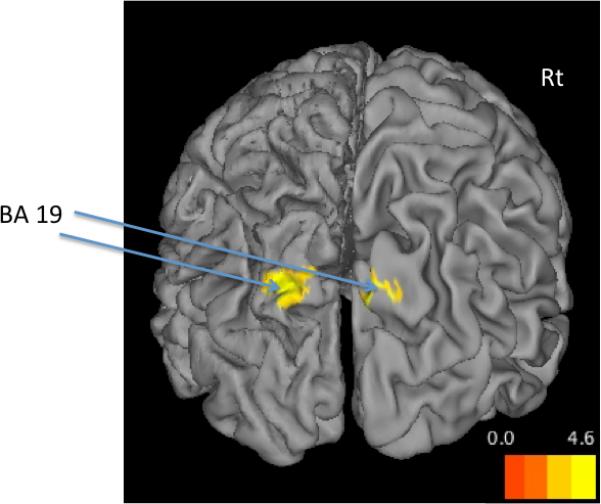
Statistical map set at a family-wise error corrected cluster-level threshold of p < 0.05 and overlaid on 3-D SPM template using CARET. The figure shows the back view of the surface (Rt=Right); BA: Brodmann Area. Patients showed more activity than controls in the cuneus region (BA19).
Figure 3 and 4 illustrate the activity of all subjects in the regions with patient-control difference.
Figure 3. Individual DMN Activity z-scores for Control>Patient.
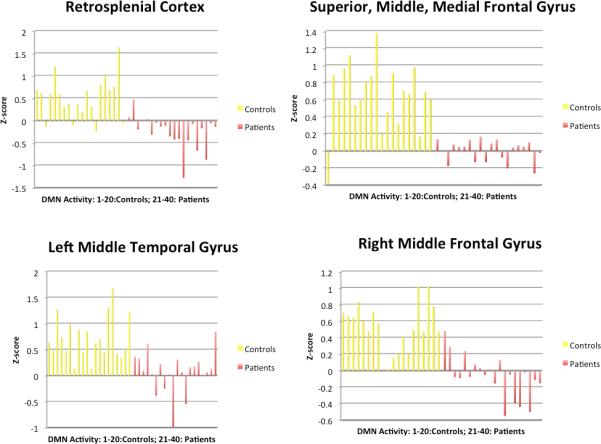
Activity (z-scores) in regions where controls show greater DMN activity than patients. The yellow bars represent DMN activity in controls and the red bars represent DMN activity in patients.
Figure 4. Individual DMN Activity z-scores for Patient>Control.
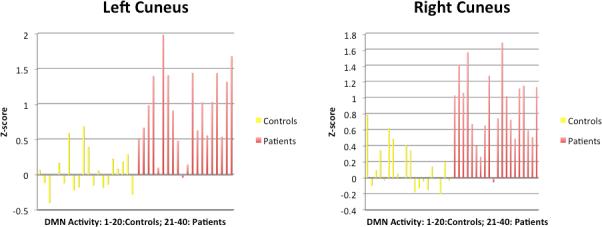
Activity (z-scores) in regions where patients show greater DMN activity than controls. The yellow bars represent DMN activity in controls and the red bars represent DMN activity in patients.
3.3 Within patient DMN activity vs. Abstinence and task performance
Exploratory analyses show that longer period of abstinence was associated with increased DMN activity in two regions (clusters): left superior, middle and medial frontal gyrus (BA9,10) (t=4.8, df=16, r=0.77) and right superior, medial frontal gyrus (BA10) and anterior cingulate (BA32) (t=6.2, df=16, r=0.84). Lower DMN activity was associated with greater risk-taking in one region (cluster): cuneus and lingual gyrus (BA18), precuneus (BA30) and posterior cingulate (BA31) (t=−4.24, df=18, r=−0.71) (see Supplementary Material Section S5 and Supplementary Figures 6 and 7****).
3.4 Examining effect of potential confounders
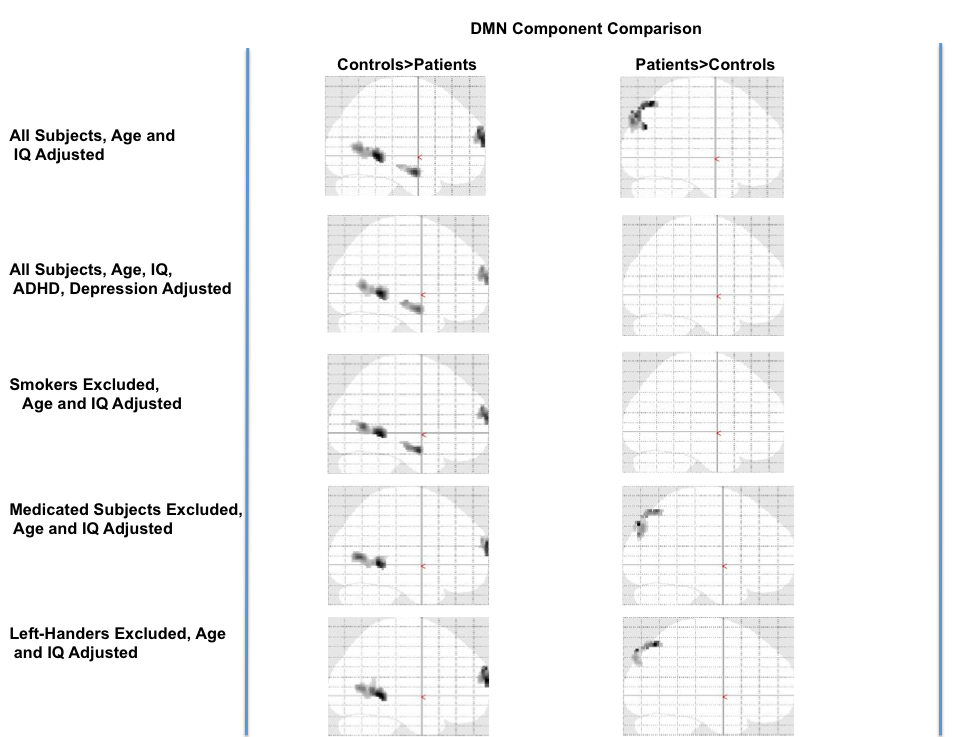
After adjusting for ADHD and depression scores, patients still demonstrated significantly reduced activity compared to controls within a similar set of regions as observed in Figure 1 (see results on SPM's glass brain in Supplementary Figure 8††††). But no regions exhibited significantly increased activation in patients compared to controls. Even after excluding smokers, left handers, and medicated subjects there was still a similar patterns of greater DMN activity in controls vs. patients (see Supplementary Figure 8‡‡‡‡).
4. DISCUSSION
Our primary goal was to identify and compare DMN activity in male CD/SUD adolescents and controls when engaged in a rapid-event fMRI task that involved decision-making, outcomes, short periods of rest (fixations) and relatively longer periods of minimal cognitive load (directed trials).
Both groups exhibited typical DMN activation patterns consistent with prior literature in adults (Buckner et al., 2008) and adolescents (Lagioia et al., 2010). As hypothesized, patients exhibited reduced DMN activity compared to controls in several DMN regions (see Figure 1, Table 2) and this pattern changed minimally (1) after adjusting for ADHD and depression, the common comorbidities in our patients and (2) after separately excluding recent smokers, left handed and medicated subjects (see Supplementary Figure 8§§§§). Conversely, patients only showed more activation than controls in the posterior precuneus/cuneus (Figure 2) and that difference was no longer significant after adjusting for ADHD and depression or after we excluded recent smokers. However, as a caveat, our approach of excluding subjects to assess possible contributions of confounding variables reduces our power to detect such differences.
4.1. Lower DMN activity in patients vs. controls
4.1.1 Medial Prefrontal Cortex (MPFC)
The DMN plays an important role in internally-directed thought (Buckner et al., 2008), becomes engaged when an individual's mind wanders about thoughts unrelated to the task at hand (Christoff et al., 2009), and activates during experimentally-directed tasks that are of an internally-focused, introspective nature (Andrews-Hanna, 2012). Furthermore, the DMN demonstrates reciprocal relationships with task difficulty during externally-directed tasks (Mason et al., 2007). Pyka et al. (2009) explanation that increased DMN activity following increased cognitive load (in a working-memory paradigm) is due to increased subsequent self-evaluation and reflection of preceding events, supports our proposal that patients may be less likely to engage in self-reflective activity in a conflicting decision paradigm requiring regular cognitive load than their non-affected peers. The anterior MPFC may play a particular role in self-reflective and evaluative thought, as it becomes engaged across a variety of tasks requiring individuals to reference information to one's self or close others (e.g. Northoff et al., 2006; Andrews-Hanna, 2012).
4.1.2 Retrosplenial Cortex
The retrosplenial cortex is part of a “medial temporal lobe (MTL) subsystem” within the DMN, which is thought to play a role in retrieving information from the past to construct mental models of what the future might hold (Buckner et al., 2008). This subsystem activates when individuals are instructed to remember their past and imagine their future using imagery-based simulation (Andrews-Hanna et al., 2010b). Furthermore, resting-state functional connectivity between regions within the MTL subsystem correlates with individual differences in spontaneous episodic memories and plans (Andrews-Hanna et al., 2010a). These findings might reflect patients being less able to utilize recollections about past behaviors and consequences when making decisions about future behaviors. This inability to use the past to guide future decisions may reduce their ability to think about the consequences of current drug use, especially in regards to past negative outcomes.
4.1.3 Lateral temporal cortex
DMN regions have also been activated in social cognition (Schilbach et al., 2008). The lateral temporal cortex is part of a “dorsal MPFC (dMPFC) subsystem” that becomes engaged during tasks involving mentalizing or theory of mind (Andrews-Hanna, 2012). The dMPFC subsystem is altered in psychological disorders associated with aberrations in social functions, including autism (Kennedy et al., 2006), schizophrenia and social phobia (Gentili et al., 2009). Alterations in these regions could therefore contribute to antisocial behaviors or tendencies in patients.
4.2 Greater DMN activity in patients vs. controls
When not adjusting for ADHD and depression, patients showed more DMN activity than controls near the bilateral cuneus, with activation overlapping the posterior zone of the precuneus and extending along the visual prestriate cortex into the dorsal lateral occipital region. Though these regions are included in the DMN mask, resting-state functional connectivity and anatomical tracing studies suggest these regions exhibit different patterns of connectivity than MPFC, posterior cingulate and lateral parietal regions (Andrews-Hanna et al., 2010a). Specifically, the posterior precuneus/cuneus connects to extrastriate regions, posterior fusiform, and lateral prestriate cortex (Margulies et al., 2009). Consistent with their pattern of projections, these regions activate during tasks involving visuo-spatial imagery, broad visual attention, and mental imagery associated with episodic memory retrieval (Andrews-Hanna et al., 2010b). After adjusting for ADHD and depression, there were no regions where patients showed significantly more DMN activity than controls. This change in cuneus (BA19) findings could indicate this DMN characteristic is related to these comorbidities (Grimm et al., 2009). For example, Cao et al. (2006) showed increased regional homogeneity (a measure of functional connectivity) in occipital cortex and the cuneus regions and decreased regional homogeneity during resting -state fMRI in various frontal regions in boys (mean age13.3 years) with ADHD.
4.3 DMN activity in a stimulus-oriented vs. resting-state fMRI paradigm
Group differences may be driven by task-related or task-unrelated alterations in the DMN (Singh and Fawcett, 2008). Other studies have used ICA to extract the DMN component across task periods (Pyka et al., 2009; Li et al., 2011; Esposito et al., 2006). DMN (or intrinsic activity) have been assessed across task performance (Fair et al., 2007; Fox and Raichle, 2007; Garrity et al., 2007; Meda et al., 2009). Smith et al. (2009) compared ICA-defined resting-state networks with fMRI networks showing consistent co-activation during task performance (using the activation maps of experiment included in the online BrainMap database (www.brainmap.org)) and showed remarkable similarity between the task-based and resting-state functional connectivity networks.
To investigate this issue, we extracted the spectral power information of the DMN component for every subject (see Supplementary Material Section S4.c*****). Consistent with prior ICA studies that examine DMN activity in the absence of an overt task (i.e., at rest), the DMN component in our study exhibited the highest spectral power in the low-frequency bin (0-0.04 Hz) for both groups (Supplementary Figure 5†††††). However, the average time course of the DMN component (see Supplementary Material Section S4.a‡‡‡‡‡) also suggests higher activity for both groups during fixation trials and during less cognitively demanding periods such as following directions versus making decisions (Supplementary Figure 3§§§§§). We also conducted multiple regression analyses fitting the DMN time course with various phases of the task [i.e., risky decision, cautious decision, reward, loss, collect (after active decision), risky directed, cautious directed, reward, loss, collect (after directed instructions)] and the beta estimates clearly suggest that the DMN activity for both groups and all conditions consistently showed higher DMN activity during directed (or inactive) phase, where subjects are mostly following simple instructions, compared to the decision (or active) phase, where subjects are more cognitively involved (see Supplementary Material Section S4.b and Supplementary Figure 4a and 4b******). Thus, it is possible that group changes in DMN activity are driven by a combination of low-frequency fluctuations unrelated to the task as well as changes induced by the task itself. Future studies examining group differences in DMN activity during a pure resting-state paradigm may help differentiate these possibilities.
4.4 Relation of DMN in patient with abstinence and task performance
DMN activity in frontal cortex (i.e., superior, middle, medial frontal gyrus and anterior cingulate) showed a positive association with abstinence from substances. Improved DMN activity with treatment has been shown before (Tregellas et al., 2011; Tanabe et al., 2011).
DMN activity in posterior cingulate, lingual gyrus, cuneus and precuneus showed a negative association with greater risk-taking. Clearly, these exploratory analyses highlight the importance of studying DMN, which could be a clinical phenotype for patients with CD/SUD. However, these exploratory analyses do not suggest causality and more work needs to be done with a larger sample to understand the intricacies of DMN functioning.
4.5 Relation between DMN and the decision-network
We previously reported in this sample hypoactivation in various critical regions of the decision-making network in patients compared to controls (Crowley et al., 2010a) and in this study we report hypoactivation in the DMN in patients compared to controls. Together, these findings suggest that patients show impaired brain activity during decision-making and inhibition and diminished activity during self-evaluation, introspection and retrieval of memory from past actions. Tang et al. (2013) recently showed poor functional connectivity between DMN and attention networks in subjects with antisocial personality disorder (age=20.52 1.37years) compared with controls (age=21.67 2.54years). More work is needed to address functional connectivity between different networks in adolescent CD/SUD patients.
4.6 Clinical Implications
DMN is a robust non-invasive biomarker, and a potential phenotype for molecular genetic studies and for brain pathology (Biswal et al., 2010; Glahn et al., 2010), distinguishes patients from controls (Broyd et al., 2009), is associated with risk-taking in our task and detects neuropathophysiological diseases (e.g., Koch et al., 2012). DMN may be sensitive to abstinence and showed improvement by therapeutic intervention (Tregellas et al., 2011; Tanabe et al., 2011). Tang et al. (2013) using a data-driven classifier method based on machine learning, recently showed that the DMN made a sizeable contribution in discriminating ASPD from control subjects.
These studies support the clinical importance of studying DMN in CD/SUD adolescents. Furthermore, by studying DMN during a task we answer an important clinical question of whether other brain networks such as DMN also differ along with decision networks (Crowley et al., 2010a) during a decision-making task.
4.7 Strengths and limitations
Results from this male-only study cannot be generalized to female adolescents. Our cross-sectional study cannot determine whether differences in DMN activity between patients and controls predate the onset of substance abuse or resulted from repeated exposure to illicit substances, alcohol and nicotine. However, in our sample no youth had currently used either nicotine or other drugs for at least a week before scanning and in some cases for up to two months. Finally, it is important to understand the nature of DMN alterations in patients and controls during effortful tasks and during the resting-state. DMN interaction with other resting-state networks and other networks responsible for cognitive and executive control such as attention and inhibitory networks could highlight certain other aspects of neural disorganization in patients. These efforts may eventually lead to candidate sites for possible therapeutic intervention.
Supplementary Material
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Acknowledgments
The authors acknowledge Yiping Du and Deb Singel from the Brain Imaging Center at University of Colorado for their technical support, Vince Calhoun and Srinivas Rachakonda from University of New Mexico for useful advise on the GIFT ICA toolbox and Samantha MaWhinney from the department of Biostatistics at University of Colorado for providing useful advice on the method section of the manuscript.
Role of Funding Source: NIDA grants DA009842, DA011015, DA034604 and the Kane Family Foundation. Dr. Sakai is also supported by NIDA grant DA031761. NIDA had no further role in study design, in the collection, analysis and interpretation of data; in writing of the report; or in the decision to submit the paper for publication.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Contributors: Authors Crowley, Banich, and Sakai are responsible for study design and in decisions involving patient sample, inclusion and exclusion criteria; authors Dalwani and Raymond were responsible in overseeing the MRI procedures, and data collection; authors Dalwani and Tregellas were responsible for data analyses procedures and running ICA analyses, author Dalwani wrote the first draft and author Andrews-Hanna assisted in overall manuscript and constructed some sections in the discussion; authors Dalwani and Mikulich-Gilbertson were responsible for statistical analyses. All authors contributed to and have approved the final manuscript.
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Supplementary material can be found by accessing the online version of this paper at http://dx.doi.org and by entering doi:...
Conflict of Interest: Dr. Crowley received travel support from the American Psychiatric Association to participate in revising the Diagnostic and Statistical Manual of Mental Disorders (DSM V). Dr. Sakai received reimbursement in 2012 for completing a policy review for the WellPoint Office of Medical Policy & Technology Assessment (OMPTA), WellPoint, Inc., Thousand Oaks, CA. He also serves as a board member of the ARTS Foundation. The other authors report no conflict of interest.
REFERENCES
- Achenbach TM. Manual for the Child Behavior Checklist/4-18 and 1991 profile. University of Vermont, Department of Psychiatry; Burlington, VT: 1991a. [Google Scholar]
- Achenbach TM. Manual for the Youth Self-Report and 1991 Profile. University of Vermont, Department of Psychiatry; Burlington, VT: 1991b. [Google Scholar]
- Allen EA, Erhardt EB, Damaraju E, Gruner W, Segall JM, Silva RF, Havlicek M, Rachakonda S, Fries J, Kalyanam R, Michael AM, Caprihan A, Turner JA, Eichele T, Adelsheim S, Bryan AD, Bustillo J, Clark VP, Feldstein Ewing SW, Filbey F, Ford CC, Hutchison K, Jung RE, Kiehl KA, Kodituwakku P, Komesu YM, Mayer AR, Pearlson GD, Phillips JP, Sadek JR, Stevens M, Teuscher U, Thoma RJ, Calhoun VD. A baseline for the multivariate comparison of resting-state networks. Front. Syst. Neurosci. 2011;5:2. doi: 10.3389/fnsys.2011.00002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- American Psychiatric Association . Diagnostic and Statistical Manual of Mental Disorders. 4th Edition, Text Revision American Psychiatric Press; Washington, DC: 2000. [Google Scholar]
- Andrews-Hanna JR. The brain's default network and its adaptive role in internal mentation. Neuroscientist. 2012;18:251–270. doi: 10.1177/1073858411403316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andrews-Hanna JR, Reidler JS, Huang C, Buckner RL. Evidence for the default network's role in spontaneous cognition. J. Neurophysiol. 2010a;104:322–335. doi: 10.1152/jn.00830.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andrews-Hanna JR, Reidler JS, Sepulcre J, Poulin R, Buckner RL. Functional-anatomic fractionation of the brain's default network. Neuron. 2010b;65:550–562. doi: 10.1016/j.neuron.2010.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bell AJ, Sejnowski TJ. An information-maximization approach to blind separation and blind deconvolution. Neural Comput. 1995;7:1129–1159. doi: 10.1162/neco.1995.7.6.1129. [DOI] [PubMed] [Google Scholar]
- Biswal BB, Mennes M, Zuo XN, Gohel S, Kelly C, Smith SM, Beckmann CF, Adelstein JS, Buckner RL, Colcombe S, Dogonowski AM, Ernst M, Fair D, Hampson M, Hoptman MJ, Hyde JS, Kiviniemi VJ, Kotter R, Li SJ, Lin CP, Lowe MJ, Mackay C, Madden DJ, Madsen KH, Margulies DS, Mayberg HS, McMahon K, Monk CS, Mostofsky SH, Nagel BJ, Pekar JJ, Peltier SJ, Petersen SE, Riedl V, Rombouts SA, Rypma B, Schlaggar BL, Schmidt S, Seidler RD, Siegle GJ, Sorg C, Teng GJ, Veijola J, Villringer A, Walter M, Wang L, Weng XC, Whitfield-Gabrieli S, Williamson P, Windischberger C, Zang YF, Zhang HY, Castellanos FX, Milham MP. Toward discovery science of human brain function. Proc. Natl. Acad. Sci. U S A. 2010;107:4734–4739. doi: 10.1073/pnas.0911855107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broyd SJ, Demanuele C, Debener S, Helps SK, James CJ, Sonuga-Barke EJ. Default-mode brain dysfunction in mental disorders: a systematic review. Neurosci. Biobehav. Rev. 2009;33:279–296. doi: 10.1016/j.neubiorev.2008.09.002. [DOI] [PubMed] [Google Scholar]
- Buckner RL, Andrews-Hanna JR, Schacter DL. The brain's default network: anatomy, function, and relevance to disease. Ann. N. Y. Acad. Sci. 2008;1124:1–38. doi: 10.1196/annals.1440.011. [DOI] [PubMed] [Google Scholar]
- Calhoun VD, Kiehl KA, Pearlson GD. Modulation of temporally coherent brain networks estimated using ICA at rest and during cognitive tasks. Hum. Brain Mapp. 2008;29:828–838. doi: 10.1002/hbm.20581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao Q, Zang Y, Sun L, Sui M, Long X, Zou Q, Wang Y. Abnormal neural activity in children with attention deficit hyperactivity disorder: a resting-state functional magnetic resonance imaging study. Neuroreport. 2006;17:1033–1036. doi: 10.1097/01.wnr.0000224769.92454.5d. [DOI] [PubMed] [Google Scholar]
- Crowley TJ, Dalwani MS, Mikulich-Gilbertson SK, Du YP, Lejuez CW, Raymond KM, Banich MT. Risky decisions and their consequences: neural processing by boys with Antisocial Substance Disorder. PLoS One. 2010a;5:e12835. doi: 10.1371/journal.pone.0012835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crowley TJ, Gelhorn H. Antisocial drug dependence. In: Koob G, Le, Moal M, Thompson R, editors. Encyclopedia of Behavioral Neuroscience. Elsevier Press; Amsterdam: 2010b. p. 1816. [Google Scholar]
- Crowley TJ, Mikulich SK, Ehlers KM, Whitmore EA, MacDonald MJ. Validity of structured clinical evaluations in adolescents with conduct and substance problems. J. Am. Acad. Child Adolesc. Psychiatry. 2001;40:265–273. doi: 10.1097/00004583-200103000-00005. [DOI] [PubMed] [Google Scholar]
- Crowley TJ, Raymond KM, Mikulich-Gilbertson SK, Thompson LL, Lejuez CW. A risk-taking “set” in a novel task among adolescents with serious conduct and substance problems. J. Am. Acad. Child Adolesc. Psychiatry. 2006;45:175–183. doi: 10.1097/01.chi.0000188893.60551.31. [DOI] [PubMed] [Google Scholar]
- Chanraud S, Pitel AL, Pfefferbaum A, Sullivan EV. Disruption of functional connectivity of the default-mode network in alcoholism. Cereb. Cortex. 2011;21:2272–2281. doi: 10.1093/cercor/bhq297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christoff K, Gordon AM, Smallwood J, Smith R, Schooler JW. Experience sampling during fMRI reveals default network and executive system contributions to mind wandering. Proc. Natl. Acad. Sci. U S A. 2009;106:8719–8724. doi: 10.1073/pnas.0900234106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dalwani M, Sakai JT, Mikulich-Gilbertson SK, Tanabe J, Raymond K, McWilliams SK, Thompson LL, Banich MT, Crowley TJ. Reduced cortical gray matter volume in male adolescents with substance and conduct problems. Drug Alcohol Depend. 2011;118:295–305. doi: 10.1016/j.drugalcdep.2011.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delfos MF. Children and Behavioural Problems: Anxiety, Aggression, Depression and ADHD; A Biopsychological Model with Guidelines for Diagnostics and Treatment. Harcourt book publishers; Amsterdam: 2004. pp. 79–100. [Google Scholar]
- Disney ER, Elkins IJ, McGue M, Iacono WG. Effects of ADHD, conduct disorder, and gender on substance use and abuse in adolescence. Am. J. Psychiatry. 1999;156:1515–1521. doi: 10.1176/ajp.156.10.1515. [DOI] [PubMed] [Google Scholar]
- Esposito F, Bertolino A, Scarabino T, Latorre V, Blasi G, Popolizio T, Tedeschi G, Cirillo S, Goebel R, Di Salle F. Independent component model of the default-mode brain function: assessing the impact of active thinking. Brain. Res. Bull. 2006;70:263–269. doi: 10.1016/j.brainresbull.2006.06.012. [DOI] [PubMed] [Google Scholar]
- Fair DA, Schlaggar BL, Cohen AL, Miezin FM, Dosenbach NUF, Wenger KK, Fox MD, Snyder AZ, Raichle ME, Petersen SE. A method for using blocked and event-related fMRI data to study “resting state” functional connectivity. Neuroimage. 2007;35:396–405. doi: 10.1016/j.neuroimage.2006.11.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fairchild G, Passamonti L, Hurford G, Hagan CC, von dem Hagen EA, van Goozen SH, Goodyer IM, Calder AJ. Brain structure abnormalities in early-onset and adolescent-onset conduct disorder. Am. J. Psychiatry. 2011;168:624–633. doi: 10.1176/appi.ajp.2010.10081184. [DOI] [PubMed] [Google Scholar]
- Fonagy P, Target M. Attachment and reflective function: their role in self-organization. Dev. Psychopathol. 1997;9:679–700. doi: 10.1017/s0954579497001399. [DOI] [PubMed] [Google Scholar]
- Fox MD, Raichle ME. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat. Rev. Neurosci. 2007;8:700–711. doi: 10.1038/nrn2201. [DOI] [PubMed] [Google Scholar]
- Garrity AG, Pearlson GD, McKiernan K, Lloyd D, Kiehl KA, Calhoun VD. Aberrant “default mode” functional connectivity in schizophrenia. Am. J. Psychiatry. 2007;164:450–457. doi: 10.1176/ajp.2007.164.3.450. [DOI] [PubMed] [Google Scholar]
- Giedd JN, Lalonde FM, Celano MJ, White SL, Wallace GL, Lee NR, Lenroot RK. Anatomical brain magnetic resonance imaging of typically developing children and adolescents. J. Am. Acad. Child Adolesc. Psychiatry. 2009;48:465–470. doi: 10.1097/CHI.0b013e31819f2715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gentili C, Ricciardi E, Gobbini MI, Santarelli MF, Haxby JV, Pietrini P, Guazzelli M. Beyond amygdala: Default Mode Network activity differs between patients with social phobia and healthy controls. Brain Res. Bull. 2009;79:409–413. doi: 10.1016/j.brainresbull.2009.02.002. [DOI] [PubMed] [Google Scholar]
- Glahn DC, Winkler AM, Kochunov P, Almasy L, Duggirala R, Carless MA, Curran JC, Olvera RL, Laird AR, Smith SM, Beckmann CF, Fox PT, Blangero J. Genetic control over the resting brain. Proc. Natl. Acad. Sci. U S A. 2010;107:1223–1228. doi: 10.1073/pnas.0909969107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greicius MD, Krasnow B, Reiss AL, Menon V. Functional connectivity in the resting brain: a network analysis of the default mode hypothesis. Proc. Natl. Acad. Sci. U S A. 2003;100:253–258. doi: 10.1073/pnas.0135058100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grimm S, Boesiger P, Beck J, Schuepbach D, Bermpohl F, Walter M, Ernst J, Hell D, Boeker H, Northoff G. Altered negative BOLD responses in the default-mode network during emotion processing in depressed subjects. Neuropsychopharmacology. 2009;34:932–843. doi: 10.1038/npp.2008.81. [DOI] [PubMed] [Google Scholar]
- Happé F, Frith U. Theory of mind and social impairment in children with conduct disorder. Br. J. Dev. Psychol. 1996;14:385–398. [Google Scholar]
- Kendler KS, Aggen SH, Knudsen GP, Roysamb E, Neale MC, Reichborn-Kjennerud T. The structure of genetic and environmental risk factors for syndromal and subsyndromal common DSM-IV axis I and all axis II disorders. Am. J. Psychiatry. 2011;168:29–39. doi: 10.1176/appi.ajp.2010.10030340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kennedy DP, Redcay E, Courchesne E. Failing to deactivate: resting functional abnormalities in autism. Proc. Natl. Acad. Sci. U S A. 2006;103:8275–8280. doi: 10.1073/pnas.0600674103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch W, Teipel S, Mueller S, Benninghoff J, Wagner M, Bokde AL, Hampel H, Coates U, Reiser M, Meindl T. Diagnostic power of default mode network resting state fMRI in the detection of Alzheimer's disease. Neurobiol. Aging. 2012;33:466–478. doi: 10.1016/j.neurobiolaging.2010.04.013. [DOI] [PubMed] [Google Scholar]
- Lagioia A, Van De Ville D, Debbane M, Lazeyras F, Eliez S. Adolescent resting state networks and their associations with schizotypal trait expression. Front. Syst. Neurosci. 2010;4 doi: 10.3389/fnsys.2010.00035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lenroot RK, Gogtay N, Greenstein DK, Wells EM, Wallace GL, Clasen LS, Blumenthal JD, Lerch J, Zijdenbos AP, Evans AC, Thompson PM, Giedd JN. Sexual dimorphism of brain developmental trajectories during childhood and adolescence. Neuroimage. 2007;36:1065–1073. doi: 10.1016/j.neuroimage.2007.03.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Z, Santhanam P, Coles CD, Lynch ME, Hamann S, Peltier S, Hu X. Increased “default mode” activity in adolescents prenatally exposed to cocaine. Hum. Brain Mapp. 2011;32:759–770. doi: 10.1002/hbm.21059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma N, Liu Y, Fu XM, Li N, Wang CX, Zhang H, Qian RB, Xu HS, Hu X, Zhang DR. Abnormal brain default-mode network functional connectivity in drug addicts. PLoS One. 2011;6:e16560. doi: 10.1371/journal.pone.0016560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Margulies DS, Vincent JL, Kelly C, Lohmann G, Uddin LQ, Biswal BB, Villringer A, Castellanos FX, Milham MP, Petrides M. Precuneus shares intrinsic functional architecture in humans and monkeys. Proc. Natl. Acad. Sci. U S A. 2009;106:20069–20074. doi: 10.1073/pnas.0905314106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mason MF, Norton MI, Van Horn JD, Wegner DM, Grafton ST, Macrae CN. Wandering minds: the default network and stimulus-independent thought. Science. 2007;315:393–395. doi: 10.1126/science.1131295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKeown MJ, Makeig S, Brown GG, Jung TP, Kindermann SS, Bell AJ, Sejnowski TJ. Analysis of fMRI data by blind separation into independent spatial components. Hum. Brain Mapp. 1998;6:160–188. doi: 10.1002/(SICI)1097-0193(1998)6:3<160::AID-HBM5>3.0.CO;2-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meda SA, Stevens MC, Folley BS, Calhoun VD, Pearlson GD. Evidence for Anomalous Network Connectivity during Working Memory Encoding in Schizophrenia: An ICA Based Analysis. PLoS One. 2009;4:e7911. doi: 10.1371/journal.pone.0007911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ming S, Yong L, Yuan Z, Kun W, Chunshui Y, Tianzi J. Default Network and Intelligence Difference. IEEE Trans. Auton. Ment. Dev. 2009;1:101–109. [Google Scholar]
- Nock MK, Kazdin AE, Hiripi E, Kessler RC. Prevalence, subtypes, and correlates of DSM-IV conduct disorder in the National Comorbidity Survey Replication. Psychol. Med. 2006;36:699–710. doi: 10.1017/S0033291706007082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Northoff G, Heinzel A, de Greck M, Bermpohl F, Dobrowolny H, Panksepp J. Self-referential processing in our brain--a meta-analysis of imaging studies on the self. Neuroimage. 2006;31:440–457. doi: 10.1016/j.neuroimage.2005.12.002. [DOI] [PubMed] [Google Scholar]
- Pyka M, Beckmann CF, Schoning S, Hauke S, Heider D, Kugel H, Arolt V, Konrad C. Impact of working memory load on FMRI resting state pattern in subsequent resting phases. PLoS One. 2009;4:e7198. doi: 10.1371/journal.pone.0007198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raine A, Yang Y. Neural foundations to moral reasoning and antisocial behavior. Soc. Cogn. Affect. Neurosci. 2006;1:203–213. doi: 10.1093/scan/nsl033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schacter DL, Addis DR, Hassabis D, Martin VC, Spreng RN, Szpunar KK. The future of memory: remembering, imagining, and the brain. Neuron. 2012;76:677–694. doi: 10.1016/j.neuron.2012.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schilbach L, Eickhoff SB, Rotarska-Jagiela A, Fink GR, Vogeley K. Minds at rest? Social cognition as the default mode of cognizing and its putative relationship to the “default system” of the brain. Conscious. Cogn. 2008;17:457–467. doi: 10.1016/j.concog.2008.03.013. [DOI] [PubMed] [Google Scholar]
- Sharp C. Mentalizing problems in childhood disorders. In: Allen JG, Fonagy P, editors. Handbook of Mentalization-based Treatments. Wiley; Chichester: 2006. pp. 101–121. [Google Scholar]
- Sharp C. Theory of mind and conduct problems in children: deficits in reading the “emotions of the eyes.”. Cogn. Emot. 2008;22:1149–1158. [Google Scholar]
- Sharp DJ, Beckmann CF, Greenwood R, Kinnunen KM, Bonnelle V, De Boissezon X, Powell JH, Counsell SJ, Patel MC, Leech R. Default mode network functional and structural connectivity after traumatic brain injury. Brain. 2011;134:2233–2247. doi: 10.1093/brain/awr175. [DOI] [PubMed] [Google Scholar]
- Singh KD, Fawcett IP. Transient and linearly graded deactivation of the human default-mode network by a visual detection task. Neuroimage. 2008;41:100–112. doi: 10.1016/j.neuroimage.2008.01.051. [DOI] [PubMed] [Google Scholar]
- Smith SM, Fox PT, Miller KL, Glahn DC, Fox PM, Mackay CE, Filippini N, Watkins KE, Toro R, Laird AR, Beckmann CF. Correspondence of the brain's functional architecture during activation and rest. Proc. Natl. Acad. Sci. 2009;106:13040–13045. doi: 10.1073/pnas.0905267106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanabe J, Nyberg E, Martin LF, Martin J, Cordes D, Kronberg E, Tregellas JR. Nicotine effects on default mode network during resting state. Psychopharmacology (Berl.) 2011;216:287–295. doi: 10.1007/s00213-011-2221-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang Y, Jiang W, Liao J, Wang W, Luo A. Identifying individuals with antisocial personality disorder using resting-state fMRI. PLoS One. 2013;8:e60652. doi: 10.1371/journal.pone.0060652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tregellas JR, Tanabe J, Rojas DC, Shatti S, Olincy A, Johnson L, Martin LF, Soti F, Kem WR, Leonard S, Freedman R. Effects of an alpha 7-nicotinic agonist on default network activity in schizophrenia. Biol. Psychiatry. 2011;69:7–11. doi: 10.1016/j.biopsych.2010.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ward BD. [2009 June 1];Simultaneous inference for fMRI data. 2000 Jun; Available: http://afni.nimh.nih.gov/pub/dist/doc/manual/AlphaSim.pdf.
- Wechsler D. Wechsler Abbreviated Scale of Intelligence. Psychological Corp.; San Antonio, TX: 1999. [Google Scholar]
- Whitfield-Gabrieli S, Ford JM. Default mode network activity and connectivity in psychopathology. Annu. Rev. Clin. Psychol. 2012;8:49–76. doi: 10.1146/annurev-clinpsy-032511-143049. [DOI] [PubMed] [Google Scholar]
- Young SE, Stallings MC, Corley RP, Krauter KS, Hewitt JK. Genetic and environmental influences on behavioral disinhibition. Am. J. Med. Genet. 2000;96:684–695. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.