

Abstract
The location and morphology of astrocytes are known to contribute to their diversity, and this diversity is often associated with their selective functions. However, molecular markers for astrocyte subtypes are largely unknown. In this study, we found that the immunoreactivity for glycoprotein GPM6B (M6B-IR) is preferentially expressed in the astrocytes associated with ventricles or neurogenic regions of the adult mouse brain. In particular, M6B-IR in the neurogenic niche was confined to glial fibrillary acidic protein- or nestin-expressing neural stem cells. Furthermore, in the injury penumbra, reactive astrocytes expressing nestin also exhibited strong M6B-IR. These results reveal that GPM6B is a potential molecular marker for a subset of astrocytes, as well as for the injury-dependent activation of astrocytes.
Keywords: astrocytes, injury, M6B, nestin, neurogenesis, stem cells
INTRODUCTION
Astrocytes are essential cells for regulating various brain functions including synapse development, neurotransmission and energy metabolism [1-3]. These diverse functions appear to be mediated by different subtypes of astrocytes, and thus astrocytes are believed to be highly heterogeneous depending on their location and the status of the brain [4]. For example, astrocytes in the neurogenic niche have unique functions to promote neuronal differentiation of neural stem cells [5]. On the other hand, astrocytes in the non-neurogenic cerebral cortex regions appear to have glial-promoting activity to the neural stem cells. In fact, adult neural stem cells (NSCs) themselves express markers for astrocytes, and appear to exhibit glia-like characteristics [6]. This heterogeneity of astrocytes, however, is poorly defined and primarily classified by their morphology and/or developmental origin [7], and only few molecular markers have been identified [8, 9]. The identification of selective molecular or cytochemical markers for astrocyte subtypes may provide a valuable tool for studying astrocyte heterogeneity and their selective functions.
Glycoprotein M6B, a transmembrane protein related to the proteolipid protein family, is enriched in the myelin of the central nervous system (CNS) [10]. M6B has been reported to be expressed in neurons and oligodendrocytes [11], but its expression in astrocytes has not been examined. In general, the functions of these proteolipids are unclear, but they appear to aid in the modulation of neurotransmission [12]. For instance M6A, which also belongs to the proteolipid protein family, interacts with µ-opioid receptors, while M6B interacts with serotonin transporters [13]; however, the functional significance of these interactions is not entirely known.
In this study, we found that the immunoreactivity for GPM6B (M6B-IR) is enriched in a subset of astrocytes, mainly those associated with the neurogenic niche in adult mice. Furthermore, astrocytes activated by traumatic brain injury also exhibited strong M6B-IR, which may be a useful molecular marker revealing the heterogeneity of astrocytes in the mouse brain.
MATERIALS AND METHODS
Animals and traumatic brain injury
Adult male C57BL/6 mice (3-5 months old) were obtained from Orient Bio (Sungnam, Korea). For traumatic brain injury, metal probe (3-mm diameter) was pre-chilled in the liquid nitrogen, and placed on the skull (2 mm lateral and 1 mm anterior from the bregma) for 60 seconds. The cut skin was sutured, and the animals returned to home cage. All experimental procedures were approved by Animal Care and Use Committee of Korea University.
Neural stem cell culture
Adult mouse neural stem cells (NSCs) were obtained and maintained as previously reported [14]. Briefly, the subventricular zone (SVZ) tissues were isolated from adult male mice (4-8 weeks old C57BL/6) and incubated with a mix of 0.8% papain (Worthington, Lakewood, NJ, USA) and 0.08% dispase II (Roche Applied Science, USA) in HBSS. Dissociated cells were seeded and maintained as neurosphere in DMEM/F12 media containing 1% N2, 2% B27, penicillin-streptomycin (Gibco BRL, USA), basic fibroblast growth factor (20 ng/ml, Invitrogen, USA), epidermal growth factor (20 ng/ml, Invitrogen, USA), and L-ascorbic acid (20 ng/ml, Sigma-Aldrich, USA). Neurospheres were dissociated with Accutase (Millipore, USA) and seeded on the laminin-coated dish as a monolayer for the differentiation. The differentiation was induced by the deprivation of the EGF/bFGF for 6 days.
Reverse transcription-polymerase chain reaction (RT-PCR)
Total RNAs (2 µg) were isolated from cultured NSCs or adult (8 weeks old) hippocampal tissues using the RNeasy mini kit (Qiagen, Cat# 74104, Germany), and were reverse-transcribed with reverse transcriptase (Bioneer, Cat# E-3100) and oligo (dT) primer. An aliquot of RT products was then subjected to PCR amplification as follows; initial denaturation at 95℃ for 10 min and 40 cycles of 95℃ for 15 s, 60℃ for 30 s, and 72℃ for 30 s. with specific primers. Forward primer, 4F (cac cgt ggc aat tct tga gc) was designed in exon 4 (Genbank # NM_001177955.1). Two reverse primers, 8R (ata aac actg gca ctg cgg a) and 10R (agc ctg cat ttt gct tgc at), were designed in exon 8 and exon 10, respectively. The combination of 4F and 8R (4F8R) yields 260 bp PCR products, whereas 4F and 10R (4F10R) yields two different PCR products from isoform-1 (563 bp) and isoform-2 (654 bp). PCR-amplified cDNA fragments were then separated onto 3% agarose gel containing of etidium bromide.
Histology
Animals were cardiac-perfused with 4% paraformaldehyde and the brains were isolated. Brains were then post-fixed in the same fixative overnight, and transferred to 30% sucrose solution until sinking. Following freezing, brains were cut (40 µm) coronal on a cryostat, and stored at -20℃ in the storage solution (50% phosphate buffered saline (PBS) and 50% glycerol). Following blocking with 3% bovine serum albumin (BSA) and 0.1% Triton-X100 in the PBS, primary antibodies were applied to the sections for 24 hours at room temperature. Primary antibodies used in this study are as follows: anti-GPM6B (Abcam, Cat# ab92988, 1:200), anti-glial fibrillary acidic protein (GFAP: SantaCruz, 1: 1,000), anti-Nestin (Abcam, 1:200), and anti-doublecortin (DCX: SantaCruz, 1: 200). After washes, appropriate secondary antibodies were incubated with sections for 30 min at room temperature. The signals were then observed under the confocal microscope (Zeiss, LSM510, Goettingen, Germany).
RESULTS
Distribution of M6B-IR in the adult mouse brain
We examined the distribution of M6B-IR in the adult brain and found that it was strongly localized in neurogenic regions, including the subventricular zone (SVZ; Fig. 1A~C) and rostral migratory stream (RMS; Fig. 1D~F). Double labeling with GFAP, an astrocyte marker, revealed that cells expressing M6B-IR in these regions were astrocytes. In addition to the astrocytes in the neurogenic regions, the astrocytes associated with ventricles or with white matter showed strong M6B-IR. Examples of M6B-IR in the third ventricle (V III; Fig. 1G~I) and corpus callosum (Corp Cal; Fig. 1J~L) are shown. On the other hand, astrocytes in the grey matter or associated with blood vessels did not exhibit substantial M6B-IR (Fig. 1M~O). Some NeuN-expressing neurons also exhibited M6B-IR, as reported previously (Fig. 1M~O, insets).
Fig. 1.
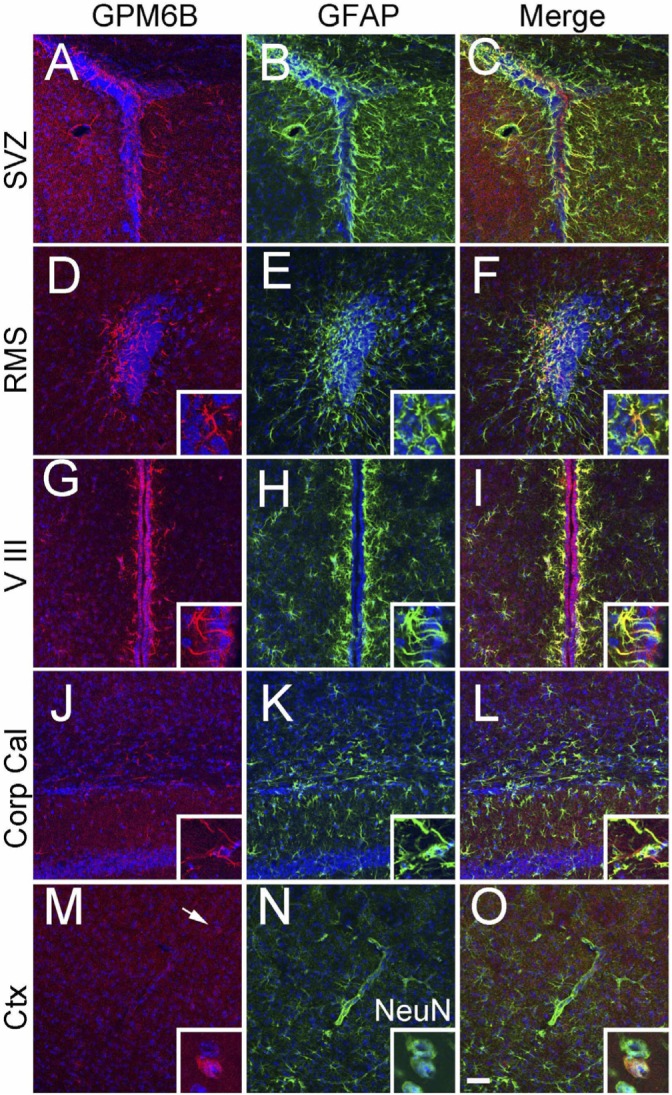
Distribution of M6B-IR in the astrocytes in several regions of adult mouse brain, including subventricular zone (SVZ, A~C), rostral migratory stream (RMS, D~F), third ventricle (V III, G~I), corpus callosum (Corp Cal, J~L), and cerebral cortex (Ctx, M~O). M6B-IR was visualized in red (A, D, G, J, M), GFAP was double-labeled in green (B, E, H, K, N), and merged images were shown in C, F, I, L, O. Nuclei were counter-stained with Hoechst33342 in blue. Insets in A~O show large magnification images of M6B and GFAP double-labeled cells. Arrow in M indicates the M6B-IR in neuron-like cells. Insets in M~O show M6B and NeuN double-labeled cell. At least 6 sections from 3 different animals were examined, and the typical images are shown. Scale Bar: 50 µm.
Expression of GPM6B in neural stem cells
Because neural stem cells (NSCs) in the SVZ are known to express GFAP, we examined whether M6B-IR was found in NSCs both in vivo and in vitro (Figs. 2 and 3). GFAP-expressing cells in the SVZ of the lateral ventricle expressed M6B-IR (Fig. 2A~C); these cells also expressed the NSC marker, nestin (Fig. 2D~F). Although the majority of GFAP or nestin-expressing cells in the SVZ also exhibited M6B-IR, we found that a small subset (<5%) of nesting-expressing NSCs did not exhibit M6B-IR (Fig. 2D~F, arrows). On the other hand, neuroblasts expressing DCX in the SVZ did not exhibit noticeable M6B-IR (Fig. 2G~I), suggesting that GPM6B expression was rapidly down-regulated by the differentiation of NSCs into neuroblasts.
Fig. 2.
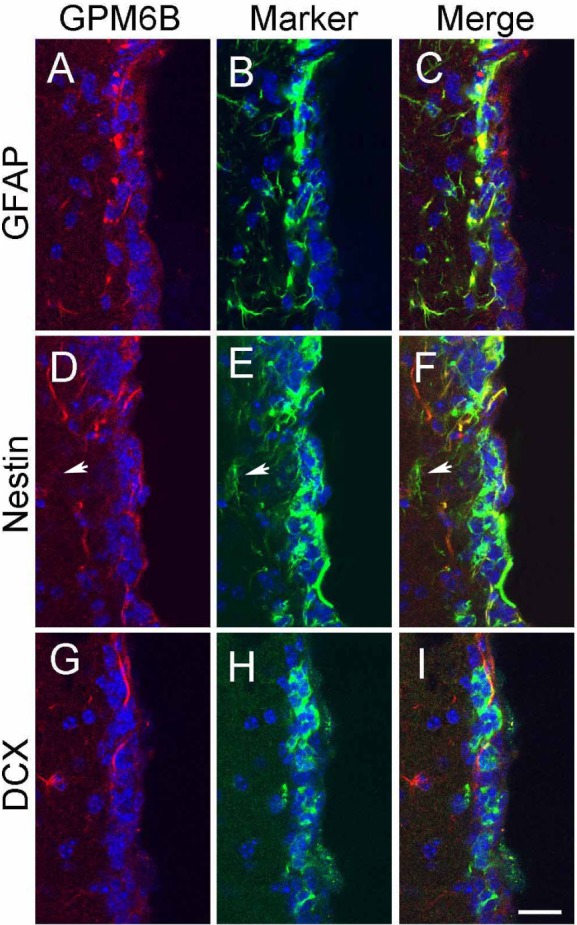
Distribution of M6B-IR in the astrocytes in the subventricular zone. GPM6B (A, D, G) was double-labeled with GFAP (B), Nestin (E), and DCX (H). Merged images are shown in C, F, I. Arrows indicate the cell expressing nestin but not M6B-IR. Nuclei were counter-stained with Hoechst33342 in blue. At least 6 sections from 3 different animals were examined, and the typical images are shown. Scale Bar: 20 µm.
Fig. 3.

Expression of GPM6B in the cultured neural stem cells. GPM6B (A, D) was double-labeled with Nestin (B) or GFAP (E). Merged images are shown in C and F. Nuclei were counter-stained with Hoechst33342 in blue. Scale Bar: 20 µm. (G) Expression of Gmp6b isoforms by RT-PCR. We used two different sets of primers which amplify all isoforms with the same size (4F8R, 260 bp, arrow), or two different sizes (4F10R, 563 bp for isoform-1, and 654 bp for isoform-2), respectively. Abbreviations are N, NSCs; H, adult hippocampus tissue.
In cultured NSCs, M6B-IR was found in proliferating nestin-expressing cells (Fig. 3A~C). When growth factors were removed from the culture media, NSCs spontaneously differentiate into neurons and glial cells. After spontaneous differentiation we also found that M6B-IR was maintained in the GFAP-expressing astrocytes, as expected (Fig. 3D~F). On the other hand, neuron-like cells containing multiple processes also exhibited M6B-IR (arrows, Fig. 3D~F); this finding is not similar to our in vivo results. Multiple isoforms of GPM6B have been identified [11, 15]. Because the antibody we used recognize all isoforms of GPM6B, we further confirmed the expression of GPM6B isoforms in the cultured NSCs at mRNA level by RT-PCR (Fig. 3G). Both cDNAs from culture NSCs and adult hippocampal tissues yielded the same size of PCR products by 4F8R primer set which amplifies common region (exon4-exon8). Interestingly, 4F10R primer set demonstrated that isoform-2 is more abundant in the NSCs comparing to the adult hippocampus, which is consistent with the previous finding that the expression of isoform-2 is down-regulated during the development [15].
Induction of M6B-IR in reactive astrocytes after traumatic brain injury
Traumatic brain injury is known to promote the activation of quiescent astrocytes, resulting in the proliferation and hypertrophy of astrocytes in the injury penumbra region. Therefore, we investigated whether reactive astrocytes also exhibit the enhanced M6B-IR (Fig. 4). Double labeling of GPM6B and nestin, which is known to be expressed in immature reactive astrocytes, revealed that a subset of reactive astrocytes expressed GPM6B on 3 (Fig. 4D~F) or 7 days (Fig. 4G~I) after traumatic brain injury. Double labeling with GFAP and GPM6B also indicated that these cells were astrocytes (insets, Fig. 4D~F).
Fig. 4.
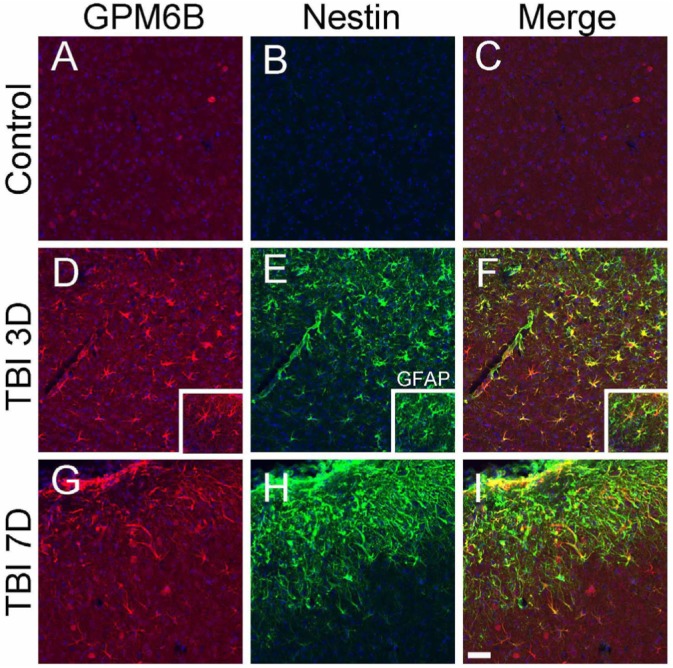
Distribution of M6B-IR in reactive astrocytes after traumatic brain injury. In control cerebral cortex (A~C), or penumbra regions 3 days (D~F), and 7 days (G~I) after traumatic brain injury, GPM6B (A, D, G) and Nestin (B, E, H) were double-labeled. Nuclei were counter-stained with Hoechst33342. Merged images were shown in C, F, I. Insets in D~F show the double-labeled cells of M6B-IR and GFAP. At least 6 sections from 3 different animals were examined, and the typical images are shown. Scale Bar: 50 µm.
DISCUSSION
In this study, we identified a subset of astrocytes that strongly exhibited M6B-IR. GPM6B has been reported to be expressed in neurons and oligodendrocytes, but its expression in astrocytes has yet to be addressed [11]. Interestingly, astrocytes that have thick processes and express high levels of GFAP are frequently associated with the ventricles; this astrocyte subtype was found to be immunoreactive for GPM6B. The role of astrocytes in the ventricle is highly specialized. Ventricles contain cerebral spinal fluid, which selectively exchanges material with the brain parenchyma. Because ependymal cells that constitute the ventricular wall do not form tight junctions [16], the selective transport of materials has to be controlled by specialized cells such as ventricular astrocytes. Thus, ventricular astrocytes should be functionally distinct from astrocytes in other brain regions, and the presence of M6B-IR in these astrocytes may be related to their molecular heterogeneity and specialized function. In this respect, it should be noted that astrocytes associated with blood vessels-which are a component of the blood-brain-barrier-do not strongly exhibit M6B-IR. Although both astrocyte subpopulations contribute to the selective transport of extracellular materials in the brain parenchyma, they differentially exhibit M6B-IR and are, therefore, molecularly distinct.
Further, we found that in addition to ventricular astrocytes, cell types such as neurons and other astrocyte subpopulations in the white matter or glia tube in the RMS express M6B-IR. Moreover, NSCs localized to the SVZ also exhibited M6B-IR. Considering that NSCs also express GFAP, expression of M6B-IR in both astrocytes and NSCs also indicate the molecular similarity of these two cell populations. Strong M6B-IR in non-astrocytic cells suggests that M6B-IR cannot be a unique cell-type marker for the subset of astrocytes, and also expressed in other cell types including NSCs. In fact, reactive astroglia situated near the traumatic brain injury penumbra in the cerebral cortex also exhibited up-regulation of M6B-IR, although cortical astrocytes did not strongly exhibit M6B-IR under normal conditions. During reactive astrogliosis, astrocytes undergo massive hypertrophy with the induction of GFAP fiber bundles [17]. Our results showed that most astrocytes expressing M6B-IR in the normal brain also expressed relatively high-levels of GFAP. Together, these findings suggest that the hypertrophic or "reactive" status of astrocytes under normal or injured conditions could up-regulate the expression of GPM6B.
Although the function of GPM6B in astrocytes is unknown, it has been established that chronic stress represses GPM6A, its close family molecule, expression in the hippocampus. Furthermore, overexpression of GPM6A in neurons increases the density of filopodia, suggesting its involvement in actin-based structural remodeling in neurons [18]. The selective expression of GPM6B in astrocytes that show strong cytoskeletal bundling and hypertrophy observed in this study may be indicative of GPM6B's role in the structural remodeling of astrocytes. Both GPM6A and GPM6B contain an extracellular domain that enables their interaction with external ligands, and treatment with GPM6A antibodies in cultured media interferes with neurite extension [19, 20]. Therefore, it appears that GPM6B is localized at the astrocyte cellular membrane and is involved in the modulation of transmembrane receptors related to downstream actin remodeling. This idea is supported by the interaction of GPM6B with serotonin transporters [13]. It is well known that astrocytes in the RMS show high expression levels for GABA transporters, which are essential for the regulation of neuroblast migration [13]. Thus, it is plausible that GPM6B in astrocytes may be involved in the regulation of neuroblast migration via an interaction with transmembrane receptor/transporters. However, M6A-induced membrane deformation has been reported to be not associated with actin [21, 22]. Therefore, it is also possible that GPM6B may exert its role in an actin-independent manner.
ACKNOWLEDGEMENTS
This research was supported by the Brain Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT & Future Planning (2012-0004004 to YKim and 2013R1A1A3011896 to JYK).
References
- 1.Magistretti PJ, Pellerin L. The astrocyte-mediated coupling between synaptic activity and energy metabolism operates through volume transmission. Prog Brain Res. 2000;125:229–240. doi: 10.1016/S0079-6123(00)25013-9. [DOI] [PubMed] [Google Scholar]
- 2.Bélanger M, Allaman I, Magistretti PJ. Brain energy metabolism: focus on astrocyte-neuron metabolic cooperation. Cell Metab. 2011;14:724–738. doi: 10.1016/j.cmet.2011.08.016. [DOI] [PubMed] [Google Scholar]
- 3.Prebil M, Jensen J, Zorec R, Kreft M. Astrocytes and energy metabolism. Arch Physiol Biochem. 2011;117:64–69. doi: 10.3109/13813455.2010.539616. [DOI] [PubMed] [Google Scholar]
- 4.Oberheim NA, Goldman SA, Nedergaard M. Heterogeneity of astrocytic form and function. Methods Mol Biol. 2012;814:23–45. doi: 10.1007/978-1-61779-452-0_3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ma DK, Ming GL, Song H. Glial influences on neural stem cell development: cellular niches for adult neurogenesis. Curr Opin Neurobiol. 2005;15:514–520. doi: 10.1016/j.conb.2005.08.003. [DOI] [PubMed] [Google Scholar]
- 6.Alvarez-Buylla A, Seri B, Doetsch F. Identification of neural stem cells in the adult vertebrate brain. Brain Res Bull. 2002;57:751–758. doi: 10.1016/s0361-9230(01)00770-5. [DOI] [PubMed] [Google Scholar]
- 7.Chaboub LS, Deneen B. Developmental origins of astrocyte heterogeneity: the final frontier of CNS development. Dev Neurosci. 2012;34:379–388. doi: 10.1159/000343723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kim WR, Kim JY, Moon Y, Kim HJ, Kim H, Sun W. Regional difference of reactive astrogliosis following traumatic brain injury revealed by hGFAP-GFP transgenic mice. Neurosci Lett. 2012;513:155–159. doi: 10.1016/j.neulet.2012.02.023. [DOI] [PubMed] [Google Scholar]
- 9.Emsley JG, Macklis JD. Astroglial heterogeneity closely reflects the neuronal-defined anatomy of the adult murine CNS. Neuron Glia Biol. 2006;2:175–186. doi: 10.1017/S1740925X06000202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fernández ME, Alfonso J, Brocco MA, Frasch AC. Conserved cellular function and stress-mediated regulation among members of the proteolipid protein family. J Neurosci Res. 2010;88:1298–1308. doi: 10.1002/jnr.22298. [DOI] [PubMed] [Google Scholar]
- 11.Werner H, Dimou L, Klugmann M, Pfeiffer S, Nave KA. Multiple splice isoforms of proteolipid M6B in neurons and oligodendrocytes. Mol Cell Neurosci. 2001;18:593–605. doi: 10.1006/mcne.2001.1044. [DOI] [PubMed] [Google Scholar]
- 12.Cooper B, Fuchs E, Flügge G. Expression of the axonal membrane glycoprotein M6a is regulated by chronic stress. PLoS One. 2009;4:e3659. doi: 10.1371/journal.pone.0003659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fjorback AW, Müller HK, Wiborg O. Membrane glycoprotein M6B interacts with the human serotonin transporter. J Mol Neurosci. 2009;37:191–200. doi: 10.1007/s12031-008-9092-4. [DOI] [PubMed] [Google Scholar]
- 14.Moon Y, Kim JY, Choi SY, Cho HM, Kim H, Sun W. Expression of ezrin in subventricular zone neural stem cells and their progeny in adult and developing mice. Histochem Cell Biol. 2013;139:403–413. doi: 10.1007/s00418-012-1048-7. [DOI] [PubMed] [Google Scholar]
- 15.Vouyiouklis DA, Werner H, Griffiths IR, Stewart GJ, Armin-Nave K, Thomson CE. Molecular cloning and transfection studies of M6b-2, a novel splice variant of a member of the PLP-DM20/M6 gene family. J Neurosci Res. 1998;52:633–640. doi: 10.1002/(SICI)1097-4547(19980615)52:6<633::AID-JNR2>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- 16.Mack A, Neuhaus J, Wolburg H. Relationship between orthogonal arrays of particles and tight junctions as demonstrated in cells of the ventricular wall of the rat brain. Cell Tissue Res. 1987;248:619–625. doi: 10.1007/BF00216492. [DOI] [PubMed] [Google Scholar]
- 17.Eng LF, Ghirnikar RS. GFAP and astrogliosis. Brain Pathol. 1994;4:229–237. doi: 10.1111/j.1750-3639.1994.tb00838.x. [DOI] [PubMed] [Google Scholar]
- 18.Scorticati C, Formoso K, Frasch AC. Neuronal glycoprotein M6a induces filopodia formation via association with cholesterol-rich lipid rafts. J Neurochem. 2011;119:521–531. doi: 10.1111/j.1471-4159.2011.07252.x. [DOI] [PubMed] [Google Scholar]
- 19.Michibata H, Okuno T, Konishi N, Wakimoto K, Kyono K, Aoki K, Kondo Y, Takata K, Kitamura Y, Taniguchi T. Inhibition of mouse GPM6A expression leads to decreased differentiation of neurons derived from mouse embryonic stem cells. Stem Cells Dev. 2008;17:641–651. doi: 10.1089/scd.2008.0088. [DOI] [PubMed] [Google Scholar]
- 20.Michibata H, Okuno T, Konishi N, Kyono K, Wakimoto K, Aoki K, Kondo Y, Takata K, Kitamura Y, Taniguchi T. Human GPM6A is associated with differentiation and neuronal migration of neurons derived from human embryonic stem cells. Stem Cells Dev. 2009;18:629–639. doi: 10.1089/scd.2008.0215. [DOI] [PubMed] [Google Scholar]
- 21.Sato Y, Watanabe N, Fukushima N, Mita S, Hirata T. Actin-independent behavior and membrane deformation exhibited by the four-transmembrane protein M6a. PLoS One. 2011;6:e26702. doi: 10.1371/journal.pone.0026702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sato Y, Mita S, Fukushima N, Fujisawa H, Saga Y, Hirata T. Induction of axon growth arrest without growth cone collapse through the N-terminal region of four-transmembrane glycoprotein M6a. Dev Neurobiol. 2011;71:733–746. doi: 10.1002/dneu.20941. [DOI] [PubMed] [Google Scholar]