

Graphical abstract
Highlights
► Most current anthelmintics act on the nematode nervous system. ► Nematodes express a large variety of ion channels and receptors. ► Glutamate-gated chloride channels and nicotinic receptors are key drug targets. ► Many other receptor and ion channel families have potential.
Keywords: Avermectin, Levamisole, Anthelmintic resistance, Acetylcholinesterase
Abstract
Many of the anthelmintic drugs in use today act on the nematode nervous system. Ion channel targets have some obvious advantages. They tend to act quickly, which means that they will clear many infections rapidly. They produce very obvious effects on the worms, typically paralyzing them, and these effects are suitable for use in rapid and high-throughput assays. Many of the ion channels and enzymes targeted can also be incorporated into such assays. The macrocyclic lactones bind to an allosteric site on glutamate-gated chloride channels, either directly activating the channel or enhancing the effect of the normal agonist, glutamate. Many old and new anthelmintics, including tribendimidine and the amino-acetonitrile derivatives, act as agonists at nicotinic acetylcholine receptors; derquantel is an antagonist at these receptors. Nematodes express many different types of nicotinic receptor and this diversity means that they are likely to remain important targets for the foreseeable future. Emodepside may have multiple effects, affecting both a potassium channel and a pre-synaptic G protein-coupled receptor; although few other current drugs act at such targets, this example indicates that they may be more important in the future. The nematode nervous system contains many other ion channels and receptors that have not so far been exploited in worm control but which should be explored in the development of effective new compounds.
1. Introduction
Parasitic nematodes remain serious pathogens of humans (Brooker et al., 2010), their animals (both domestic and livestock) (McKellar and Jackson, 2004) and plants (Sasser, 1980; Davis and Mitchum, 2005). They infect a very large proportion of the people on the planet, with serious and frequently underestimated effects on their physical, intellectual and economic development, and ‘steal’ an enormous amount of our food by parasitizing the plants and animals we use to feed ourselves. For these reasons, their control, and possible eventual elimination, remains a major priority in international public health and agriculture. In the continued absence of effective vaccines and major improvements in human sanitation, chemotherapy and chemoprophylaxis are the only viable options for large-scale control and elimination efforts.
Many of the drugs currently used for these purposes target the nematode nervous system, and in many ways this is not surprising. The nervous system makes up about 30% of all of the cells in nematodes, based on work with Caenorhabditis elegans (White et al., 1986), and many worms need to move continuously to maintain their position in the host. In the case of parasites of the gastro-intestinal tract, paralysis will result in their rapid removal. In addition, the presence or absence of observable locomotion is frequently used as a quick and easy method of telling whether nematodes are alive or dead – many species move vigorously and continuously – and so makes a useful screen for anthelmintic activity. This has been further developed into assays such as the larval migration inhibition assay (Wagland et al., 1992; Kotze et al., 2006) and others which rely on inhibition of locomotion to measure susceptibility and resistance to anthelmintic compounds (Kotze et al., 2004); these may have the potential to be at least partially automated (Smout et al., 2010; Carr et al., 2011). However, nematodes do more than just move; they also feed, mate, and detect and respond to environmental cues. All of their behaviors are, of course, controlled by the nervous system and may have potential as anthelmintic targets. The nervous system of nematodes is anatomically simple, generally containing about 300 neurons (White et al., 1986), and is well conserved between species (Goldschmidt, 1909), but is neurochemically extremely complex, using many transmitter and receptor combinations that are not present in mammals. This complexity opens a rich seam of possibilities as anthelmintic targets, but also makes elucidating their exact mechanisms of action a real challenge. The situation is further complicated because the neurochemistry, unlike the anatomy, also varies considerably between nematode species, especially in terms of the size and complexity of some of the receptor and ion channel families (Williamson et al., 2007; Wolstenholme et al., 2011).
In this review, I will discuss the neuroactive drugs currently used to treat and prevent parasitic nematode infections, and our current understanding of how they work. There have been some major advances in this area in the last few years, both in the introduction of new compounds (Kaminsky et al., 2008a; Little et al., 2011) and new information about the way that some of the existing ones, especially the macrocyclic lactones (MLs), work (Lynagh and Lynch, 2010b; Hibbs and Gouaux, 2011). These advances have not only increased our knowledge of the mechanisms of action of these compounds, but may also suggest novel ways in which they can be used to reduce the problems caused by drug resistance (Wolstenholme et al., 2004; Kaplan and Nielsen, 2010; Osei-Atweneboana et al., 2011; Sutherland and Leathwick, 2011; Vercruysse et al., 2011). I will also briefly discuss how other parts of the nematode nervous system might make attractive drug targets, and how they may be affected by our current compounds. I will concentrate on human and animal parasites, as those are the ones targeted by the drugs under discussion; however, several aspects may also be applicable to the plant parasitic species.
2. Drugs currently available
A variety of compounds is currently used in the treatment and prevention of parasitic nematode infections. The most important are listed in Table 1, which also demonstrates how many of these drugs specifically target molecules found in neurons and muscles. In fact all of them, except for the benzimidazoles, diethylcarbamizine (DEC) and doxycyline, which affect the Wolbachia endosymbiont of filarial species, act at the ion channels found on nematode nerves and muscles. Many of these targets are widely conserved in animals, including mammals, so at first glance the selective toxicity of the drugs for nematodes, and, in some cases, other invertebrates might be surprising, and by some criteria molecules such as nicotinic acetylcholine receptors and potassium channels would not be considered attractive for developing drugs specific for helminths. However, these receptors and ion channels possess a complex pharmacology, with multiple allosteric and regulatory sites (Kaczorowski et al., 1996; Wu et al., 2006; Thompson et al., 2010), and this has led to a large pharmacopeia of useful compounds, often with very selective actions. Many of the genes that code for these proteins are nematode-specific and well-conserved within the phylum (Table 2), which goes some way to explaining the broad spectrum of many of the drugs that affect them. It would be no surprise if these ‘old’ targets continue to supply us with important new drugs for several years to come.
Table 1.
Anthelmintics in current use.
| Drug | Target | Status | References |
|---|---|---|---|
| Benzimidazoles (albendazole, mebendazole) | β-Tubulin | Widely used in human and animal medicine. Part of Global Program for the Elimination of Lymphatic Filariasis (GPELF) | Kohler (2001) and Ottesen et al. (2008) |
| Macrocyclic lactones (ivermectin, moxidectin) | Glutamate-gated chloride channels | IVM is a mainstay of African Program for Onchocerciasis Control, and is used in GPELF. Widely used in animal medicine | Wolstenholme and Rogers (2005) and Amazigo (2008) |
| Piperazine derivatives (diethylcarbamazine) | Piperazine is an agonist at GABA receptors, DEC has a different mode of action, not fully understood | DEC is widely used in humans, as part of GPELF | Martin (1985) and Bockarie and Deb (2010) |
| Imidazothiazoles (levamisole) | Nicotinic acetylcholine receptors | Widely used in animal medicine | Keiser and Utzinger (2008) and Sattelle (2009) |
| Tetrahydropyrimidine derivatives (pyrantel, oxantel, morantel) | Nicotinic acetylcholine receptors | Used to treat some human infections. Widely used in animal medicine. Oxantel is especially effective against Trichuris spp. | Kopp et al. (2008) |
| Amino-acetonitrile derivatives (monepental) | Choline receptors | Recently introduced for use in sheep | Kaminsky et al. (2008a) and Sager et al. (2009) |
| Tribendimidine | Nicotinic acetylcholine receptors | Approved in China for human use | Steinmann et al. (2008) and Hu et al. (2009) |
| Spiroindoles (derquantel) | Nicotinic acetylcholine receptors | Used in combination with abamectin in sheep (NZ) | Robertson et al. (2002) and Little et al. (2011) |
| Cyclic depsipeptides (emodepside) | SLO-1 potassium channels? Latrophilin receptors? | Used in combination with praziquantel in cats and dogs | Welz et al. (2011) |
| Doxycycline | Wolbachia endosymbionts | Hoerauf et al. (2003) |
Table 2.
Ion channel and receptor genes encoding anthelmintic targets in C. elegans and parasitic nematodes. Nematodes contain a plethora of genes encoding receptors and ion channels and it is beyond the scope of this article to provide a full comparison between the species. Shown here are the C. elegans genes that have been shown to encode components of anthelmintic drug targets, along with the equivalent target genes in H. contortus and, where available, selected other parasitic species. Members of the gene families that have not been shown to encode drug targets (e.g. the predicted GluCl gene glc-4) have been excluded. Older gene names, especially for H. contortus, used in the literature are shown in brackets.
| Drug class | Target | C. elegans genes | H. contortus genes | Other parasites |
|---|---|---|---|---|
| Macrocyclic lactones | GluCl | glc-1 | Hco-glc-2b (HG4) | avr-14 is present in all species examined. glc-2 is present in clades 3, 4 and 5. glc-3 is present in clades 4 and 5 |
| glc-2 (when co-expressed with glc-1) | Hco-glc-3b | |||
| glc-3 | Hco-avr-14b (HG3, GluClα3B) | |||
| avr-14ba (gbr-2b) | Hco-glc-5 (GluCla, HG5) | |||
| avr-15 | Hco-glc-6b | |||
| Piperazine | GABA receptors | unc-49 | Hco-unc-49 | unc-49 is widely conserved in nematodes |
| Imidazothiazoles, tetrahydropyrimidine, tribendimidine, spiroindoles | nAChR | lev-1 | Hco-acr-8 | acr-8, unc-29, unc-38 and unc-63 are widely conserved |
| lev-8 | Hco-lev-1c | |||
| unc-29 | Hco-unc-29d | |||
| unc-38 | Hco-unc-38 | |||
| unc-63 | Hco-unc-63 | |||
| AADs | Choline receptors | acr-23 | Hco-mptl-1e | acr-23/mptl-1is absent from clade 1, 3 and 4 parasites |
| deg-3 | Hco-deg-3 | |||
| des-2 | Hco-deg-2 | |||
| Cyclic depsipeptides | Potassium channels | slo-1 | Hco-slo-1 | Widely conserved |
| Cyclic depsipeptides | Latrophilin receptors | lat-1 | Hco-lat-1 (Hc-110R) | Widely conserved |
The avr-14 gene is alternatively spliced to give two subunits, AVR-14A and AVR-14B. To date, only AVR-14B has been shown to form a ML target.
These genes have not been formally shown to encode components of an ML-sensitive receptor.
Hco-lev-1 encodes a nAChR subunit that lacks an N-terminal signal peptide and has not been shown to contribute to a functional channel or drug target.
Hco-unc-29 has undergone at least two rounds of gene duplication, resulting in four genes, Hco-unc-29.1 to Hco-unc-29.4.
Hco-mptl-1 has not been formally shown to encode part of a monepantel-sensitive receptor.
3. Cys-loop ligand-gated ion channels as anthelmintic targets
One of the reasons that this family of receptors, which includes the glutamate-gated chloride channels (GluCl) and the nicotinic acetylcholine receptors (nAChR), has been so useful to those who wish to kill parasites is that it is large and diverse. Though the family is conserved between nematode species, and even between nematodes and mammals, its size and composition varies greatly, which has resulted in the identification of highly specific and potent drugs. All of the members of this family possess the same basic architecture, with five similar subunits surrounding a central pore; the channels open on binding of a specific ligand to the extracellular part of the receptor, allowing ions to pass down electro-chemical gradients into or out of the cell (Thompson et al., 2010). The ions that are allowed to pass through can be cations (Na+, K+ and Ca++) or anions (Cl−) and the ligands that gate the channels vary considerably, though many are, or are derived from, amino-acids (e.g. glutamate and GABA) and biogenic amines (e.g. 5-HT (serotonin), dopamine and tyramine).
A very important family of receptors in all nervous systems, the ionotropic glutamate receptors (iGluR), is not the target for any current anthelmintic drugs, despite the vital roles of this family of receptors in the nematode central nervous system (Aronoff et al., 2004; Kano et al., 2008). These receptors have quite a different architecture to the GluCl and nAChR (Mayer, 2011); perhaps this restricts the flexibility of the agonist and allosteric binding sites to permit the development of nematode-specific compounds. If this is not the case, then iGluR from parasitic nematodes may have considerable potential as potential drug targets.
3.1. Glutamate-gated chloride channels
The glutamate-gated chloride channels (GluCls) form a branch of the ligand-gated ion channel family that is confined to invertebrates (Cleland, 1996). They are most closely related to mammalian glycine receptors and, more distantly, to the GABA-gated chloride channels found throughout the animal kingdom (Vassilatis et al., 1997b; Dent, 2006). These channels are inhibitory under normal ionic conditions, with Cl− ions flowing into the cell, hyperpolarizing it. They are the targets of the ML group of anthelmintics, the biggest selling class of drugs in veterinary medicine, which includes ivermectin and moxidectin. These drugs are extremely potent anthelmintics, insecticides and acaricides (Campbell et al., 1983), and bind nematode GluCl with extremely high affinity (Rohrer et al., 1994; Gill and Lacey, 1998; Dent et al., 2000; Cheeseman et al., 2001). Recently, Hibbs and Gouaux (2011) published the structure of a C. elegans GluCl complexed with ivermectin (Fig. 1), showing the drug bound in the pore region of the channel, between adjacent subunits. This confirmed earlier results, obtained from site-directed mutagenesis, which had suggested that ivermectin binds to this site, which is analogous to the anesthetic-binding site found on mammalian GABAA and glycine receptors (Lynagh and Lynch, 2010a,b). It has long been known that although ivermectin can directly activate some GluCl, it does not do so via binding to the normal agonist binding site; activation by ivermectin is very slow but essentially irreversible (Cully et al., 1994; Vassilatis et al., 1997a; Horoszok et al., 2001; Forrester et al., 2003; Yates and Wolstenholme, 2004; McCavera et al., 2009). The MLs also potentiate the activation of the channels by the normal ligand, glutamate (Cully et al., 1994; Forrester et al., 2002, 2004), and this may be the important effect of the drug in vivo. The high-resolution structure of the GluCl provides a clear explanation for these physiological and pharmacological findings; the hydrophobic MLs have to dissolve in the plasma membrane of the cell expressing the GluCl and then bind to the pore region of the receptor. It is not clear whether they can bind to the closed state of the GluCl or must wait until it switches to the open state, whether this is a spontaneous opening or caused by agonist binding, but once bound the drug holds the channel in the open state, preventing the α-helixes that form the pore region (Unwin, 2005) from rotating back to close the ion channel. The very slow off-rate of ligand-binding results in a long-lasting, almost permanent, channel-opening and hyperpolarization of the cell – rendering it no longer excitable.
Fig. 1.
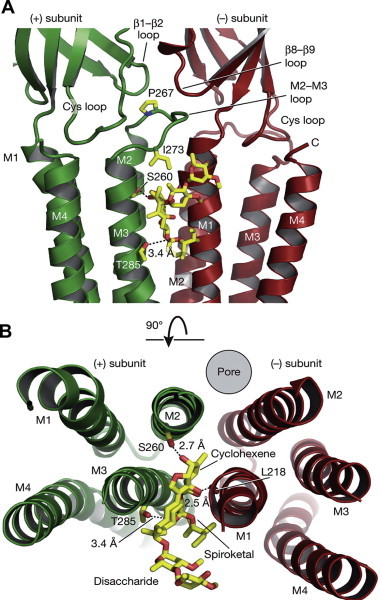
Structure of ivermectin bound to a glutamate-gated chloride channel. The GluCl shown is formed of GLC-1 subunits (Cully et al., 1994). Two views are shown. (A) The view is from the side of the receptor, looking parallel with the membrane. (B) Looking down on the binding site from the outside of the cell, with the extracellular part of the GluCl removed. The binding site is between the M3 helix of the (+) subunit and M1 of the (−) subunit.
© 2011 Reprinted by permission from Macmillan Publishers Ltd.: Nature 274, 54. Copyright 2011
The effects on the worms of this irreversible activation of inhibitory receptors will depend on where the channels are expressed. Studies on C. elegans and parasitic species showed that the GluCl were expressed on pharyngeal muscle and motor neurons, among other cells (Dent et al., 1997, 2000; Laughton et al., 1997a; Portillo et al., 2003; Glendinning et al., 2011), and effects on both locomotion and feeding were observed on parasites, especially GI parasites, though some differences between species were also observed (Kass et al., 1980; Geary et al., 1993; Sheriff et al., 2002, 2005; Holden-Dye and Walker, 2006; Tompkins et al., 2010). However, this does not explain all of the drug’s effects, for example, the long-lasting sterilization of Onchocerca volvulus adults that is observed in vivo (Awadzi et al., 1985). The in vitro concentrations of the drugs that are required to directly inhibit nematode movement or other processes in vitro are frequently much higher than those that kill parasites in vivo; for example, the ML are effective heartworm preventatives at nanomolar concentrations in vivo, yet in vitro micromolar concentrations are required to inhibit locomotion (Evans et al., 2011). Recently, a novel explanation for the effect of MLs on Brugia malayi microfilariae has been proposed, inhibition of protein secretion due to activation of GluCl expressed on the muscle controlling the excretory/secretory pore (Moreno et al., 2010). Such a mechanism could render the parasites more susceptible to attack by components of the innate immune system (Zahner et al., 1997), and at least partially explain the large gap between the in vivo and in vitro drug concentrations needed for anthelmintic action. The GluCl subunit detected in the ES muscle, AVR-14, is conserved in all nematodes studied so far (Laughton et al., 1997b; Jagannathan et al., 1999; Njue et al., 2004; Yates and Wolstenholme, 2004; Tandon et al., 2006; Williamson et al., 2007; El-Abdellati et al., 2011), so this finding may have wider implications for way the drug works in other parasites. AVR-14 is also expressed on motor neurons (Dent et al., 2000; Portillo et al., 2003; Glendinning et al., 2011) and is therefore involved in locomotion, which might be why surrogate assays for resistance which involve movement, such as the LMIA, are effective in some species.
The GluCl are also expressed on sensory neurons, in amphids and elsewhere (Rolfe et al., 2001; Portillo et al., 2003; Glendinning et al., 2011), and it is also possible that the ML might affect sensory processes, though this has been little studied. It is intriguing that amphid derangement has been linked to ML-resistance in both Haemonchus contortus and C. elegans (Freeman et al., 2003); whether this reflects a direct drug effect on sensory processes or perhaps its route on entry into the worm is not known.
3.2. Other ligand-gated chloride channels
The GluCl are famous as the target for the ML anthelmintics, but nematodes possess a large number of ligand-gated chloride channels, with a greater variety of endogenous ligands than mammals. It has long been known that nematodes have inhibitory GABA-gated chloride channels at the neuromuscular junction (Holdendye et al., 1989; Richmond and Jorgensen, 1999). These channels are required because of the worm’s sinusoidal movement, with muscles on either side of the body alternately contracting and relaxing – the GABA receptors mediate the relaxation, so drug-induced activation of them induces a flaccid paralysis. This is the mode of action of piperazine (Martin et al., 1991), a GABA agonist that was introduced as an anthelmintic in the 1950s. In addition, it has been recently demonstrated that nematodes possess acetylcholine-, 5-HT-, dopamine- and tyramine-gated chloride channels (Ranganathan et al., 2000; Putrenko et al., 2005; Pirri et al., 2009; Ringstad et al., 2009), none of which are present in mammals. The original identification of many of these channels was carried out on C. elegans, but many of the genes are conserved in parasitic species and one, the dopamine-gated chloride-channel gene Hco-ggr-3, has been suggested to be associated with ML resistance in H. contortus (Rao et al., 2009, 2010). Agonists and antagonists at these receptors might be worth exploring for potential anthelmintic activity.
3.3. Nicotinic acetylcholine receptors
The nicotinic acetylcholine receptors (nAChR) are the targets of a wide range of anthelmintic compounds (Martin, 1997). In most cases, this is because they affect the nAChR at the nematode neuromuscular junction, causing, in the case of agonists, a rapid spastic paralysis that presumably leads to the worms being expelled. However, nematodes possess a wide diversity of nAChR, not only at the neuromuscular junction but also in the nerve ring and in the pharynx (Jones and Sattelle, 2004). The nAChR have a diverse and fascinating pharmacology and it is this that has allowed the development of so many drugs that target them. It might be assumed that resistance to one such drug would lead to cross-resistance to the others, but the recent experience with monepantel and derquantel has shown that this is not the case (Kaminsky et al., 2008a; Little et al., 2011).
3.3.1. Neuromuscular nAChR
The nAChR found at the neuromuscular junction (nmj) have been the most intensively studied in nematodes, especially in C. elegans and Ascaris suum, largely due to their genetic and physiological accessibility, respectively. Studies on both worms have revealed that there are multiple nAChR at the nematode nmj, though the details vary between species. In C. elegans, there are two clearly defined receptors, one very sensitive to levamisole, an important anthelmintic, and one not; the latter is sensitive to nicotine (Richmond and Jorgensen, 1999). Genetic and in vitro reconstitution studies have shown that the C. elegans levamisole-sensitive receptor is made up of five subunits, LEV-1, LEV-8, UNC-29, UNC-38 and UNC-63 (Fig. 3) (Boulin et al., 2008), whereas the levamisole-insensitive receptor is an ACR-16 homomer (Raymond et al., 2000; Touroutine et al., 2005). However, this is not the same in parasitic species. In H. contortus, there is no detectable lev-8 gene and lev-1 does not seem to encode a functional nAChR subunit; in this species the unc-29 gene has undergone multiple duplications to create four very similar genes, Hco-unc-29.1–Hco-unc-29.4. The H. contortus levamisole-sensitive receptor can be reconstituted in Xenopus oocytes by expressing the ACR-8, UNC-29, UNC-38 and UNC-63 subunits (Fig. 3) (Boulin et al., 2011). Removing the ACR-8 subunit from the H. contortus receptor produces a version which gives a smaller response to levamisole and acetylcholine, but is more sensitive to pyrantel, which is frequently thought to act at the same site at levamisole (Martin et al., 1991; Martin and Robertson, 2007). There are suggestions that increased expression of receptors lacking ACR-8 might be associated with levamisole resistance (Fauvin et al., 2010); if that is the case one might speculate that the reduced efficacy of acetylcholine at these receptors could result in fairly rapid reversion of resistance if the drug pressure is removed and the worms might also be predicted to show an increased sensitivity to pyrantel.
Fig. 3.
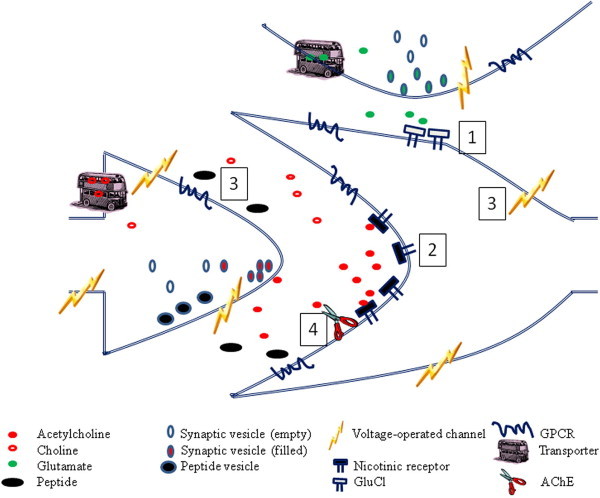
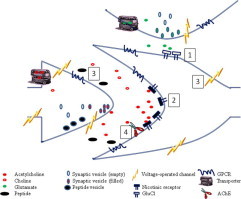
Current and potential anthelmintic targets in the nematode nervous system. A highly diagrammatic representation of a cell receiving two synaptic inputs, one cholinergic, the other glutamatergic. No such arrangement may be present in the nematode nervous system, but the molecules shown do exist, even if not in the relative positions shown here. The targets of four anthelmintic drug families are indicated: (1) The macrocyclic lactones act as glutamate-gated chloride channels; (2) the imidazothiazoles, tetrahydropyrimidine, amino-acetonitrile derivatives, tribendimidine and spiro-indoles all act at nicotinic acetylcholine receptors, though not all to the same form of the nAChR (see Fig. 2); (3) emodepside acts at SLO-1 Ca2+-activated K+ channels and at presynaptic latrophilin receptors; (4) organophosphates are potent inhibitors of acetylcholinesterase. Many other voltage-operated ion channels, transporters and G-protein coupled receptors are present and are potential targets for anthelmintic development.
The nAChR found at the A. suum nmj have been extensively studied. These receptors are amenable to electrophysiological studies because of the large size of the worm and the extrasynaptic distribution of the receptors. These studies have identified three physiological and pharmacological subtypes of nAChR in A. suum – the so-called ‘N’ (for nicotine), ‘L’ (for levamisole) and ‘B’ (for bephenium) sub-types (Martin et al., 2004a,b; Qian et al., 2006). The names reflect those compounds that show selective activity at those receptors. Despite the superficial similarity to the C. elegans receptors implied by these data, the results of in vitro reconstitution experiments were quite different; in this case, receptors with properties similar to those of the ‘N’ and ‘L’ subtypes could be produced by expression of just two subunits, Asu-UNC-29 and Asu-UNC-38 (Williamson et al., 2009). The difference between the two sub-types was the ratio of the two subunits in the pentameric receptor (Fig. 2). Interestingly, a change in the ratio of the two subunits also produced changes in the sensitivity of the resulting receptors to both pyrantel and oxantel, confirming a previous report that oxantel acted at the ‘N’ sub-type of the A. suum nAChR rather than the ‘L’ sub-type, at which pyrantel was more active (Martin et al., 2004b). Of course, such in vitro studies do not necessarily reflect the in vivo composition of the receptors but they provide important information about which subunit combinations may or may not be viable, and what their properties might be. One exciting implication of the studies on both the A. suum and H. contortus receptors is that resistance to levamisole might increase susceptibility to pyrantel, and vice versa, so that it would be difficult for worms to become resistant to both drugs, despite the fact that they apparently have the same target.
Fig. 2.
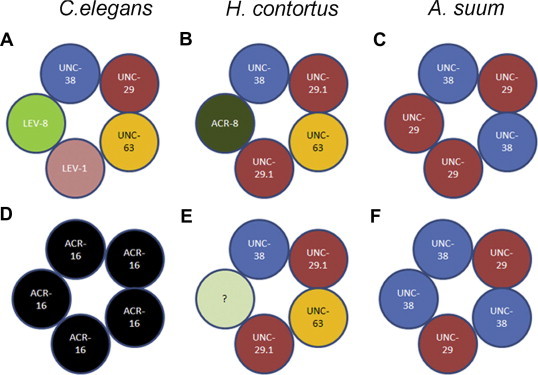
The inferred subunit composition of some nematode neuromuscular junction nAChR. Different forms of the receptor are found elsewhere in the nervous system, and are activated by monepental and related drugs. (A) A levamisole-sensitive, but nicotine-insensitive nAChR was reconstituted in the Xenopus oocyte by co-expression of the C. elegans LEV-1, LEV_8, UNC-29, UNC-38 and UNC-63 subunits (Boulin et al., 2008). (B) A levamisole-sensitive nAChR reconstituted by co-expression of the indicated subunits from H. contortus (Boulin et al., 2011). (C) A levamisole- and oxantel-sensitive, but pyrantel-insensitive, nAChR reconstituted by co-expression of the indicated subunits from A. suum (Williamson et al., 2009). (D) A nicotine-sensitive, but levamisole-insensitive receptor is formed by expression of the C. elegans ACR-16 subunit (Raymond et al., 2000). (E) Omission of Hco-ACR-8 from the receptor shown in panel B results in a nAChR with reduced sensitivity to levamisole, but increased sensitivity to pyrantel (Boulin et al., 2011). (F). Changing the ratio of the Asu-UNC-29 and -UNC-38 subunits results in a receptor with increased sensitivity to nicotine and pyrantel, reduced sensitivity to levamisole and which is oxantel insensitive (Williamson et al., 2009).
The nmj nAChR are also the targets of two newer drugs, tribendimidine and derquantel (Robertson et al., 2002; Hu et al., 2009; Ruiz-Lancheros et al., 2011). Tribendimidine has been reported to act as an agonist at the C. elegans levamisole-sensitive nAChR, and also has important activities against flukes, and derquantel has been shown to antagonize nicotinic responses in both C. elegans and A. suum. More detailed analysis suggests that derquantel is most active against the ‘B’ sub-type of the A. suum nAChR. The appearance of these drugs, and of the amino-acetonitrile derivatives demonstrates that nAChR remain important molecules to consider in anthelmintic drug discovery.
3.3.2. Other nAChR
Given the large number of nAChR subunit genes encoded by at least some nematodes, it is not surprising that these receptors are expressed in a large number of locations throughout the nervous system and, at least in the case of C. elegans, in the pharynx (Treinin and Chalfie, 1995; Jones and Sattelle, 2004; McKay et al., 2004; Jospin et al., 2009). The pharmacology of some of these receptors is significantly different to those found at the nmj and this has been exploited in the development of a novel anthelmintic class, the amino-acetonitrile derivatives (AADs) (Kaminsky et al., 2008a), one of which, AAD1566 or monepantel, has been commercialized (Kaminsky et al., 2008b). These compounds act on neuronal nAChR, and experiments using mutant strains of C. elegans indicated that the target receptor included the ACR-23 and DES-2 subunits (Kaminsky et al., 2008a). These subunits form, in C. elegans, a subfamily of nAChR which seem to be unique to nematodes; other members of the family are DEG-3, ACR-5, ACR-7, ACR-17, ACR-18 and ACR-20. There is evidence to suggest that their normal ligand is in fact choline rather than acetylcholine (Yassin et al., 2001; Rufener et al., 2010a). In H. contortus resistance to monepantel was largely associated with mutations in the Hco-mptl-1 gene, a member of the deg-3 sub-family most similar to acr-20 (Rufener et al., 2009). Members of this family are widely found across parasitic species and at least one member is present in the genome of all parasites sequenced to date; however it has been reported that the efficacy of monepantel correlates with the presence of an acr-23 or mptl-1(acr-20) ortholog (Rufener et al., 2010b). These genes are absent from Pristionchus pacificus, Strongyloides ratti, B. malayi, Trichinella spiralis or A. suum. The normal functions of MPTL-1 and ACR-20 are not known; in C. elegans knockdown of acr-23 expression by RNA interference is embryonic lethal (Sonnichsen et al., 2005). Treatment of nematodes with monepantel causes paralysis, presumably due either to activation of neurons controlling movement or because the choline receptor is also expressed on muscle cells; C. elegans DES-2 is expressed on head muscles (Yassin et al., 2001).
In vitro, monepantel has been shown to act as a positive allosteric modulator, enhancing the response to choline of a reconstituted nAChR containing the H. contortus DEG-3 and DES-2 subunits (Rufener et al., 2010a). However, the addition of MPTL-1 to this receptor had no effect on either its response to choline or to monepantel, which may mean that this subunit combination does not reflect the native monepantel target. The inactive enantiomer of monepantel, AAD-2224, was an antagonist at the Hco-DEG-3/Hco-DES-2 receptor.
The example of monepantel illustrates the potential of this unique sub-family of nematode nAChR as potential drug targets. The fact that this drug is effective against parasites resistant to other nicotinic drugs again shows that we cannot assume that the emergence of resistance to one nicotinic drug, such as levamisole, implies that this will inevitably cause cross-resistance to the others. For the worms, their neurochemical complexity may, in this case, be a two-edged sword; as they adapt to cope with the pressure applied by one drug, they may make themselves more vulnerable to other compounds. The trick for those interested in control may be to keep up with those changes and to respond quickly and effectively to alterations in the parasites resistance/susceptibility spectrum as they arise.
4. Other ion channels
4.1. Potassium channels
In excitable cells such as neurons and muscle, the membrane potential and generation of action potentials is largely controlled by ion channels that open and close in response to changes in the membrane potential and other signals. Three families of voltage-operated cation channels are present in vertebrates, gating sodium, potassium and calcium, respectively. No voltage-operated sodium channels have been detected in nematodes and it is believed that their ‘action potentials’ are calcium-dependent (Mellem et al., 2008; Gao and Zhen, 2011). A considerable number of human pharmaceuticals act as modulators of voltage-operated channels, as do many toxins, so it would not be surprising if compounds could be developed that would specifically activate or block nematode channels. Such compounds would be predicted to seriously affect neuronal signaling, causing the death of the parasite.
To date, only one anthelmintic, emodepside, has been shown to require a voltage-gated ion channel for its activity (Guest et al., 2007; Welz et al., 2011). Emodepside inhibits locomotion, feeding, egg-laying and slows development of C. elegans and parasitic worms (Harder et al., 2003; Willson et al., 2003; Bull et al., 2007; Welz et al., 2011), and these multiple effects may be due to activation of more than one target. Following a screen to identify C. elegans mutants resistant to emodepside, it was found that worms with loss-of-function mutations in slo-1, a gene that encodes a voltage- and calcium-activated potassium, or BK, channel (Atkinson et al., 1991), are resistant to emodepside inhibition of locomotion and pharyngeal pumping whereas gain-of-function mutants in the same gene phenocopy the action of the drug (Guest et al., 2007). Evidence that this mode of action was conserved in parasites was recently provided by the successful expression of SLO-1 from Ancylostoma caninum and Cooperia oncophora in a slo-1 mutant of C. elegans; this restored susceptibility to the drug (Welz et al., 2011), whereas expression of the human form of the protein did not (Crisford et al., 2011). SLO-1 channels are highly conserved, regulating neurosecretion, hormone release, muscle contraction, and neuronal excitability (Salkoff et al., 2006), which is consistent with the multiple effects of the drug seen in vivo. A very recent paper (Buxton et al., 2011) has confirmed that SLO-1 is conserved in A. suum, along with LAT-1 (see Section 5) and that both are expressed in muscle flaps. Application of emodepside to this preparation produced a slow increase in voltage-activated K+ currents, consistent with activation of SLO-1 channels, but this increase was enhanced by pharmacological stimulation of the protein kinase C (PKC) and nitric oxide (NO) signaling pathways. One interpretation of these findings is that, although the anthelmintic effects of emodepside are manifested through SLO-1 activation, the actual target of the drug is upstream of the channel, potentially activating it via PKC and/or NO.
To date, none of the other nematocidal drugs in current use have effects on voltage-operated ion channels, though praziquantel, the mainstay of current schistosomiasis chemotherapy, has been shown to activate schistosome calcium channels in vitro (Kohn et al., 2001; Greenberg, 2005). Kwok et al. (2006) demonstrated that C. elegans genetic screens could be used to identify novel small molecule inhibitors of voltage-operated calcium channels, but these do not seem to have been developed for any anthelmintic activity.
4.2. Other ion channels
All animals, including nematodes, possess several other families of ion channels that are expressed in the nervous system. To date, none of these have been exploited in anthelmintic drug development, though the intracellular ryanodine receptors are the targets of the phthalic acid diamide and anthranilic diamide classes of insecticides (Sattelle et al., 2008); these receptors are conserved in nematodes, where they are encoded by the unc-68 gene (Maryon et al., 1996). Parasitic nematodes possess multiple TRP channels (Wolstenholme et al., 2011), which in C. elegans and other phyla are involved in multiple sensory processes (Chalfie, 2009; Garrity et al., 2010) and might be interesting potential targets. Another potential ion channel target is the DEG/ENaC (named after the C. elegans degenerins and the mammalian epithelial amiloride-sensitive Na+ channel) family of sodium channels. In the worm, the genes encoding these channels were originally identified because dominant mutant alleles caused inappropriate cell death (Driscoll, 1992), later found to be due to their constitutive activation (Hong and Driscoll, 1994). This property would suggest that specific agonists of these receptors might make useful anthelmintics, though they may not as fast-acting as those that induce paralysis.
5. G protein-coupled receptors
The G protein-coupled receptors (GPCRs) make up the largest family of proteins in the nervous system, with about 900 genes encoding these receptors in the C. elegans genome (Bargmann, 1998). However, a large proportion of these are olfactory receptors which are not well conserved in parasitic species and may not be considered to be good drug targets. The ligands for many others have not yet been identified; many of these may be peptide receptors (see Section 6) or ‘hormone’-like receptors not expressed solely in the nervous system and therefore not strictly within the scope of this review. However, this still leaves many GPCRs that are expressed within the nervous system that might be viable targets for anthelmintic drugs. Table 1 shows that, if this is the case, they have not been exploited so far.
There is one possible exception to this. Emodepside, in addition to its action at SLO-1 potassium channels, binds to Hc-110R, a GPCR from H. contortus that also acts as a receptor for latrotoxin, a toxin from the black widow spider (Saeger et al., 2001). This receptor, also called latrophilin or depsiphilin (Kruger et al., 2009), is encoded by the lat-1 gene in C. elegans. Activation of Hc-110R or LAT-1 inhibits pharyngeal pumping in both the parasite and C. elegans (Willson et al., 2004). However, LAT-1 does not mediate the effects of emodepside on C. elegans locomotion (Bull et al., 2007; Guest et al., 2007). Latrophilins are conserved in animal nervous systems and are found in the pre-synaptic membrane, where they induce transmitter release (Silva et al., 2009) via an interaction with UNC-13 dependent vesicle priming (Willson et al., 2004). In C. elegans at least, the evidence seems clear that SLO-1 is a more important emodepside target than LAT-1, but it is possible that this is not the case for all the parasites against which the drug is active.
Many neurotransmitters act via GPCRs (Fig. 3), which can be present on pre-, post- and extra-synaptic membranes and whose function may be to modulate fast neurotransmission mediated by ligand-gated ion channels. A well studied group is the biogenic amines, which includes 5-hydroxytryptamine (5-HT, or serotonin), dopamine, tyramine and octopamine. All of these molecules act at GPCRs (Komuniecki et al., 2004), though, as stated earlier, nematodes also possess chloride channels gated by 5-HT, dopamine and tyramine. At least some of these receptors have also been identified in parasitic species (Huang et al., 1999, 2002; Smith et al., 2003, 2007) and many have distinct pharmacological characteristics that suggest that nematode-specific agonists and antagonists could be developed as useful anthelmintics (White et al., 2007). In C. elegans, dopamine and 5-HT have well-established roles in modulating behavior, including locomotion, feeding and egg-laying (Chase and Koelle, 2007). In H. contortus, 5-HT was detected in amphid and pharyngeal neurons, and in male-specific neurons, whereas dopamine was only found in motor neuron commissures; application of either caused inhibition of movement, in the case of dopamine this was confined to the central part of the worm (Rao et al., 1987). Exogenous 5-HT also stimulates feeding in A. suum and Nippostrongylus brasiliensis (Brownlee et al., 1997; Trim et al., 2001; Huang et al., 2010). By contrast, it had no effect on the contractile activity of Dirofilaria immitis (Bowen and Vitayavirasak, 2008).
Other classical neurotransmitters, including acetylcholine and GABA, also act at G-protein coupled receptors and regulate nematode locomotion (Dittman and Kaplan, 2008). ACh acts at muscarinic receptors, encoded in nematodes by the gar genes (Hwang et al., 1999; Lee et al., 1999, 2000). These are present in parasitic species, and have atypical muscarinic pharmacology (Segerberg and Stretton, 1993; Kimber et al., 2009), which again suggest that helminth-specific molecules could be developed.
6. Peptide signaling systems
Nematodes encode a multitude of peptide transmitters and hormones, including FLPs (FMRFamide-like peptides), NLPs (neuropeptide-like peptides) and insulin-like peptides (Li and Kim, 2010; Marks and Maule, 2010) and more continue to be discovered (Jarecki et al., 2011); these peptides play multiple modulating roles in the nervous system (McVeigh et al., 2006; Husson et al., 2007; Walker et al., 2009). Many of the peptides in nematodes, such as the FLPs, are not found in mammals and so peptidergic signaling by these molecules can be considered a promising target for new drug discovery (Martin and Robertson, 2010). Despite the example of emodepside (Harder and von Samson-Himmelstjerna, 2002), peptides themselves are generally considered unpromising molecules from which to develop anthelmintic drugs, and so attention has focused on the use of non-peptide small molecules (Geary, 2010). Many peptides act at GPCRs; though a huge number of neuropeptides has been identified by biochemical and genetic means, the identification of the cognate receptors for these peptides has lagged somewhat behind. Interestingly, when surface plasmon resonance studies were carried out using a recombinant 54 kDa aminoterminal fragment of HC110-R, the latrophilin-like molecule identified as a possible emodepside receptor (Saeger et al., 2001), AF1, AF10, and PF2, three FMRFamide-like peptides, were found to bind (Muhlfeld et al., 2009), suggesting that these peptides might act on this receptor to modulate pharyngeal pumping. Some peptides, for example, PF4, may directly gate ion channels (Purcell et al., 2002) and, if rapid and easily observable effects are desired, might be more profitable leads to follow. Signaling downstream of the peptide receptors will affect many other components of the nervous system, and effects on the conductance of potassium and calcium channels have been observed (Verma et al., 2007, 2009), showing the cross-talk that exists between peptide and other signaling pathways.
7. Other aspects of neurotransmission
7.1. Acetylcholinesterase
Acetylcholinesterase (AChE) is responsible for breaking down acetylcholine once it has bound to, and activated, its receptors. Inhibition of this enzyme therefore causes a spastic paralysis through enhancing Ach activity and this has been exploited not only in the development of pesticides and anthelmintics, but also nerve gases (Martin, 1997). Organophosphates, such as dichlorvos and halaxon, and carbamates have been used to control plant and animal parasitic nematodes (Selkirk et al., 2005) and act by covalently modifying the active site of the enzyme. Perhaps not surprisingly, these compounds have safety problems and they are now little used for treating parasitic nematode infections. However, it may still be possible to produce selective inhibitors of nematode AChEs. A very interesting variant form of AChE is that secreted by many parasitic nematodes, especially those that colonize mucosal surfaces within the gastro-intestinal tract (Ogilvie et al., 1973; Selkirk et al., 2005). N. brasiliensis secretes three such enzymes (Hussein et al., 1999, 2002a,b), Dictyocaulus viviparous secretes at least two (Lazari et al., 2003), Trichostrongylus colubriformis and Necator americanus at least one (Griffiths and Pritchard, 1994; Pritchard et al., 1994).
7.2. Other potential targets
Neurotransmission involves a large number of proteins and cellular process, many of which are unique to excitable cells. Practically all of these are conserved in nematodes, and could, potentially, be exploited as anthelmintic targets. Many of these proteins and processes have been targeted for therapeutic purposes in mammals, which demonstrate at least the theoretical possibility that helminth specific compounds could be developed. Potential targets, in addition to those already discussed, could include the transporters that carry transmitters across plasma and vesicle membranes, selective serotonin reuptake inhibitors are an example of a drug class that acts here (Briley and Moret, 1993), or the enzymes involved in the biosynthesis, processing and degradation of transmitters, especially of neuropeptides (Londershausen, 1996). There are also many downstream signaling proteins and chaperones that are essential for accurate neurotransmission and disruption of which, by genetic means, have been shown to be lethal or deleterious to C. elegans, but whose potential as antiparasitic targets remains unexplored.
8. Concluding remarks
The nematode nervous system has long been a profitable source of anthelmintic drugs, and this looks set to continue. The advantages of interfering with neurotransmission are obvious if your goal is to kill a multi-cellular organism, and this has been exploited for the control of worms, insects, lice, mollusks and, unfortunately, mammals. Not only is the effect rapid, it is often very easy to observe and many of the molecules involved are amenable to high-throughput screening strategies (Woods and Knauer, 2010; Woods et al., 2011). Fig. 3 shows some of the proteins and processes targeted by our existing anti-nematode drugs, put into the context of a theoretical and highly-stylized synapse. The figure also serves to highlight some of the other potential targets present at the same structure. These other targets may be worth considering, as the limitations of our current anthelmintics, especially for human medicine (Geary et al., 2010), the problems of resistance in the veterinary arena (Kaplan, 2004) and the almost complete lack of good methods for controlling plant parasitic nematodes (Holden-Dye and Walker, 2011; Molinari, 2011), make the investment in continued anthelmintic discovery essential. Regardless of whether any individual component of the nematode synapse turns out to be useful in drug development, it seems certain that the nervous system as a whole will continue to be studied and exploited for many years to come.
Acknowledgments
Research in the author’s laboratory is supported by awards 1R21AI092185 and 2R56AI047194 from the National Institute of Allergy and Infectious Diseases. The content is solely the responsibility of the author and does not necessarily represent the official views of the National Institute of Allergy and Infectious Diseases or the National Institutes of Health.
References
- Amazigo U. The African programme for onchocerciasis control (APOC) Ann. Trop. Med. Parasitol. 2008;102:19–22. doi: 10.1179/136485908X337436. [DOI] [PubMed] [Google Scholar]
- Aronoff R., Mellem J.E., Maricq A.V., Sprengel R., Seeburg P.H. Neuronal toxicity in Caenorhabditis elegans from an editing site mutant in glutamate receptor channels. J. Neurosci. 2004;24:8135–8140. doi: 10.1523/JNEUROSCI.2587-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atkinson N.S., Robertson G.A., Ganetzky B. A component of calcium-activated potassium channels encoded by the Drosophila slo locus. Science. 1991;253:551–555. doi: 10.1126/science.1857984. [DOI] [PubMed] [Google Scholar]
- Awadzi K., Dadzie K.Y., Shulzkey H., Haddock D.R.W., Gilles H.M., Aziz M.A. The chemotherapy of onchocerciasis. 10. An assessment of 4 single dose treatment regimes of mk-933 (ivermectin) in human onchocerciasis. Ann. Trop. Med. Parasitol. 1985;79:63–78. [PubMed] [Google Scholar]
- Bargmann C.I. Neurobiology of the Caenorhabditis elegans genome. Science. 1998;282:2028–2033. doi: 10.1126/science.282.5396.2028. [DOI] [PubMed] [Google Scholar]
- Bockarie M.J., Deb R.M. Elimination of lymphatic filariasis: do we have the drugs to complete the job. Curr. Opin. Infect. Dis. 2010;23:617–620. doi: 10.1097/QCO.0b013e32833fdee5. [DOI] [PubMed] [Google Scholar]
- Boulin T., Gielen M., Richmond J.E., Williams D.C., Paoletti P., Bessereau J.-L. Eight genes are required for functional reconstitution of the Caenorhabditis elegans levamisole-sensitive acetylcholine receptor. Proc. Natl. Acad. Sci. USA. 2008;105:18590–18595. doi: 10.1073/pnas.0806933105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boulin T., Fauvin A., Charvet C.L., Cortet J., Cabaret J., Bessereau J.-L., Neveu C. Functional reconstitution of Haemonchus contortus acetylcholine receptors in Xenopus oocytes provides mechanistic insights into levamisole resistance. Br. J. Pharmacol. 2011;164:1421–1432. doi: 10.1111/j.1476-5381.2011.01420.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowen J.M., Vitayavirasak B. Pharmacological evaluation of contractile activity of the dog heartworm Dirofilaria immitis. Vet. Res. Commun. 2008;32:231–241. doi: 10.1007/s11259-007-9022-x. [DOI] [PubMed] [Google Scholar]
- Briley M., Moret C. Neurobiological mechanisms involved in antidepressant therapies. Clin. Neuropharmacol. 1993;16:387–400. doi: 10.1097/00002826-199310000-00002. [DOI] [PubMed] [Google Scholar]
- Brooker S., Hotez P.J., Bundy D.A.P. The global atlas of helminth infection: mapping the way forward in neglected tropical disease control. PLoS Negl. Trop. Dis. 2010;4:e779. doi: 10.1371/journal.pntd.0000779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brownlee D.J.A., Holden-Dye L., Walker R.J. Actions of the anthelmintic ivermectin on the pharyngeal muscle of the parasitic nematode, Ascaris suum. Parasitology. 1997;115:553–561. doi: 10.1017/s0031182097001601. [DOI] [PubMed] [Google Scholar]
- Bull K., Cook A., Hopper N.A., Harder A., Holden-Dye L., Walker R.J. Effects of the novel anthelmintic emodepside on the locomotion, egg-laying behaviour and development of Caenorhabditis elegans. Int. J. Parasitol. 2007;37:627–636. doi: 10.1016/j.ijpara.2006.10.013. [DOI] [PubMed] [Google Scholar]
- Buxton S.K., Neveu C., Charvet C.L., Robertson A.P., Martin R.J. On the mode of action of emodepside: slow effects on membrane potential and voltage-activated currents in Ascaris suum. Br. J. Pharmacol. 2011;164:453–470. doi: 10.1111/j.1476-5381.2011.01428.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campbell W.C., Fisher M.H., Stapley E.O., Albers-Schönberg G., Jacob T.A. Ivermectin: a potent new antiparasitic agent. Science. 1983;221:823–828. doi: 10.1126/science.6308762. [DOI] [PubMed] [Google Scholar]
- Carr J.A., Parashar A., Gibson R., Robertson A.P., Martin R.J., Pandey S. A microfluidic platform for high-sensitivity, real-time drug screening on C. elegans and parasitic nematodes. Lab Chip. 2011;11:2385–2396. doi: 10.1039/c1lc20170k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chalfie M. Neurosensory mechanotransduction. Nat. Rev. Mol. Cell Biol. 2009;10:44–52. doi: 10.1038/nrm2595. [DOI] [PubMed] [Google Scholar]
- Chase, D.L., Koelle, M.R., 2007. Biogenic amine neurotransmitters. In: The C. elegans Research Community (Ed.), Wormbook. [DOI] [PMC free article] [PubMed]
- Cheeseman C.L., Delany N.S., Woods D.J., Wolstenholme A.J. High-affinity ivermectin binding to recombinant subunits of the Haemonchus contortus glutamate-gated chloride channel. Mol. Biochem. Parasitol. 2001;114:161–168. doi: 10.1016/s0166-6851(01)00258-4. [DOI] [PubMed] [Google Scholar]
- Cleland T.A. Inhibitory glutamate receptor channels. Mol. Neurobiol. 1996;13:97–136. doi: 10.1007/BF02740637. [DOI] [PubMed] [Google Scholar]
- Crisford A., Murray C., O’Connor V., Edwards R.J., Kruger N., Welz C., von Samson-Himmelstjerna G., Harder A., Walker R.J., Holden-Dye L. Selective toxicity of the anthelmintic emodepside revealed by heterologous expression of human KCNMA1 in Caenorhabditis elegans. Mol. Pharmacol. 2011;79:1031–1043. doi: 10.1124/mol.111.071043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cully D.F., Vassilatis D.K., Liu K.K., Paress P., van der Ploeg L.H.T., Schaeffer J.M., Arena J.P. Cloning of an avermectin-sensitive glutamate-gated chloride channel from Caenorhabditis elegans. Nature. 1994;371:707–711. doi: 10.1038/371707a0. [DOI] [PubMed] [Google Scholar]
- Davis E.L., Mitchum M.G. Nematodes. Sophisticated parasites of legumes. Plant Physiol. 2005;137:1182–1188. doi: 10.1104/pp.104.054973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dent J.A. Evidence for a diverse Cys-loop ligand-gated ion channel superfamily in early bilateria. J. Mol. Evol. 2006;62:523–535. doi: 10.1007/s00239-005-0018-2. [DOI] [PubMed] [Google Scholar]
- Dent J.A., Davis M.W., Avery L. avr-15 encodes a chloride channel subunit that mediates inhibitory glutamatergic neurotransmission and ivermectin sensitivity in Caenorhabditis elegans. EMBO J. 1997;16:5867–5879. doi: 10.1093/emboj/16.19.5867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dent J.A., Smith M.M., Vassilatis D.K., Avery L. The genetics of ivermectin resistance in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA. 2000;97:2674–2679. doi: 10.1073/pnas.97.6.2674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dittman J.S., Kaplan J.M. Behavioral impact of neurotransmitter-activated G-protein-coupled receptors: muscarinic and GABA(B) receptors regulate Caenorhabditis elegans locomotion. J. Neurosci. 2008;28:7104–7112. doi: 10.1523/JNEUROSCI.0378-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Driscoll M. Molecular genetics of cell death in the nematode Caenorhabditis elegans. J. Neurobiol. 1992;23:1327–1351. doi: 10.1002/neu.480230919. [DOI] [PubMed] [Google Scholar]
- El-Abdellati A., De Graef J., Van Zeveren A., Donnan A., Skuce P., Walsh T., Wolstenholme A., Tait A., Vercruysse J., Claerebout E., Geldhof P. Altered avr-14B gene transcription patterns in ivermectin-resistant isolates of the cattle parasites, Cooperia oncophora and Ostertagia ostertagi. Int. J. Parasitol. 2011;41:951–957. doi: 10.1016/j.ijpara.2011.04.003. [DOI] [PubMed] [Google Scholar]
- Evans, C.C., Moorhead, A., Storey, B., Dzimianski, M., Wolstenholme, A., Blagburn, B., Carmichael, J., Schenker, R., Kaplan, R., 2011. In vitro bioassay for detection of anthelmintic resistance in Dirofilaria immitis. In: Proceedings of 23rd International Conference of the World Association for the Advancement of Veterinary Parasitology pp. 272.
- Fauvin A., Charvet C., Issouf M., Cortet J., Cabaret J., Neveu C. CDNA–AFLP analysis in levamisole-resistant Haemonchus contortus reveals alternative splicing in a nicotinic acetylcholine receptor subunit. Mol. Biochem. Parasitol. 2010;170:105–107. doi: 10.1016/j.molbiopara.2009.11.007. [DOI] [PubMed] [Google Scholar]
- Forrester S.G., Prichard R.K., Beech R.N. A glutamate-gated chloride channel subunit from Haemonchus contortus: expression in a mammalian cell line, ligand binding, and modulation of anthelmintic binding by glutamate. Biochem. Pharmacol. 2002;63:1061–1068. doi: 10.1016/s0006-2952(02)00852-3. [DOI] [PubMed] [Google Scholar]
- Forrester S.G., Prichard R.K., Dent J.A., Beech R.N. Haemonchus contortus: HcGluCla expressed in Xenopus oocytes forms a glutamate-gated ion channel that is activated by ibotenate and the antiparasitic drug ivermectin. Mol. Biochem. Parasitol. 2003;129:115–121. doi: 10.1016/s0166-6851(03)00102-6. [DOI] [PubMed] [Google Scholar]
- Forrester S.G., Beech R.N., Prichard R.K. Agonist enhancement of macrocyclic lactone activity at a glutamate-gated chloride channel subunit from Haemonchus contortus. Biochem. Pharmacol. 2004;67:1019–1024. doi: 10.1016/j.bcp.2003.08.047. [DOI] [PubMed] [Google Scholar]
- Freeman A.S., Nghiem C., Li J., Ashton F.T., Guerrero J., Shoop W.L., Schad G.A. Amphidial structure of ivermectin-resistant and susceptible laboratory and field strains of Haemonchus contortus. Vet. Parasitol. 2003;110:217–226. doi: 10.1016/s0304-4017(02)00321-7. [DOI] [PubMed] [Google Scholar]
- Gao S.B., Zhen M. Action potentials drive body wall muscle contractions in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA. 2011;108:2557–2562. doi: 10.1073/pnas.1012346108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garrity P.A., Goodman M.B., Samuel A.D., Sengupta P. Running hot and cold: behavioral strategies, neural circuits, and the molecular machinery for thermotaxis in C. elegans and Drosophila. Genes Dev. 2010;24:2365–2382. doi: 10.1101/gad.1953710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geary T.G. Nonpeptide ligands for peptidergic G protein-coupled receptors. In: Geary T.G., Maule A.G., editors. vol. 692. Springer-Verlag; Berlin: 2010. pp. 10–26. (Neuropeptide Systems as Targets for Parasite and Pest Control, Advances in Experimental Medicine and Biology). [Google Scholar]
- Geary T.G., Sims S.M., Thomas E.M. Haemonchus contortus: ivermectin-induced paralysis of the pharynx. Exp. Parasitol. 1993;77:88–96. doi: 10.1006/expr.1993.1064. [DOI] [PubMed] [Google Scholar]
- Geary T.G., Woo K., McCarthy J.S., Mackenzie C.D., Horton J., Prichard R.K., de Silva N.R., Olliaro P.L., Lazdins-Helds J.K., Engels D.A., Bundy D.A. Unresolved issues in anthelmintic pharmacology for helminthiases of humans. Int. J. Parasitol. 2010;40:1–13. doi: 10.1016/j.ijpara.2009.11.001. [DOI] [PubMed] [Google Scholar]
- Gill J.H., Lacey E. Avermectin/milbemycin resistance in trichostrongyloid nematodes. Int. J. Parasitol. 1998;28:863–877. doi: 10.1016/s0020-7519(98)00068-x. [DOI] [PubMed] [Google Scholar]
- Glendinning S.K., Buckingham S.D., Sattelle D.B., Wonnacott S., Wolstenholme A.J. Glutamate-gated chloride channels of Haemonchus contortus restore drug sensitivity to ivermectin resistant Caenorhabditis elegans. PLoS One. 2011;6:e22390. doi: 10.1371/journal.pone.0022390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldschmidt R. Das nervensystem von Ascaris lumbricoides und Megalocephala. Ein Versuch, in den Aufbau eines einfachen Nervensystems einzudringen, Zweiter Teil. Z. Wiss. Zool. 1909;92:306–357. [Google Scholar]
- Greenberg R.M. Are Ca2+ channels targets of praziquantel action? Int. J. Parasitol. 2005;35:1–9. doi: 10.1016/j.ijpara.2004.09.004. [DOI] [PubMed] [Google Scholar]
- Griffiths G., Pritchard D.I. Purification and biochemical-characterization of acetylcholinesterase (AChE) from the excretory/secretory products of Trichostrongylus colubriformis. Parasitology. 1994;108:579–586. doi: 10.1017/s0031182000077441. [DOI] [PubMed] [Google Scholar]
- Guest M., Bull K., Walker R.J., Amliwala K., O’Connor V., Harder A., Holden-Dye L., Hopper N.A. The calcium-activated potassium channel, SLO-1, is required for the action of the novel cyclo-octadepsipeptide anthelmintic, emodepside, in Caenorhabditis elegans. Int. J. Parasitol. 2007;37:1577–1588. doi: 10.1016/j.ijpara.2007.05.006. [DOI] [PubMed] [Google Scholar]
- Harder A., von Samson-Himmelstjerna G. Cyclooctadepsipeptides – a new class of anthelmintically active compounds. Parasitol. Res. 2002;88:481–488. doi: 10.1007/s00436-002-0619-2. [DOI] [PubMed] [Google Scholar]
- Harder A., Schmitt-Wrede H.P., Krucken J., Marinovski P., Wunderlich F., Willson J., Amliwala K., Holden-Dye L., Walker R. Cyclooctadepsipeptides – an anthelmintically active class of compounds exhibiting a novel mode of action. Int. J. Antimicrob. Agents. 2003;22:318–331. doi: 10.1016/s0924-8579(03)00219-x. [DOI] [PubMed] [Google Scholar]
- Hibbs R.E., Gouaux E. Principles of activation and permeation in an anion-selective Cys-loop receptor. Nature. 2011;474:54–60. doi: 10.1038/nature10139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoerauf A., Buttner D.W., Adjei O., Pearlman E. Science, medicine, and the future – onchocerciasis. Br. Med. J. 2003;326:207B–210B. doi: 10.1136/bmj.326.7382.207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holden-Dye L., Walker R.J. Actions of glutamate and ivermectin on the pharyngeal muscle of Ascaridia galli: a comparative study with Caenorhabditis elegans. Int. J. Parasitol. 2006;36:395–402. doi: 10.1016/j.ijpara.2005.11.006. [DOI] [PubMed] [Google Scholar]
- Holden-Dye L., Walker R.J. Neurobiology of plant parasitic nematodes. Invert. Neurosci. 2011;11:9–19. doi: 10.1007/s10158-011-0117-2. [DOI] [PubMed] [Google Scholar]
- Holdendye L., Krogsgaardlarsen P., Nielsen L., Walker R.J. GABA receptors on the somatic muscle-cells of the parasitic nematode, Ascaris suum – stereoselectivity indicates similarity to a GABAA-type agonist recognition site. Br. J. Pharmacol. 1989;98:841–850. doi: 10.1111/j.1476-5381.1989.tb14613.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hong S.T., Driscoll M. A transmembrane domain of the putative chanel subunit MEC-4 influences mechanostransduction and neurodegeneration in C. elegans. Nature. 1994;367:470–473. doi: 10.1038/367470a0. [DOI] [PubMed] [Google Scholar]
- Horoszok L., Raymond V., Sattelle D.B., Wolstenholme A.J. GLC-3: a novel fipronil and BIDN-sensitive, but picrotoxinin-insensitive, l-glutamate-gated chloride channel subunit from Caenorhabditis elegans. Br. J. Pharmacol. 2001;132:1247–1254. doi: 10.1038/sj.bjp.0703937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu Y., Xiao S.H., Aroian R.V. The new anthelmintic tribendimidine is an L-type (levamisole and pyrantel) nicotinic acetylcholine receptor agonist. PLoS Negl. Trop. Dis. 2009;3:e499. doi: 10.1371/journal.pntd.0000499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang X.Y., Duran E., Diaz F., Xiao H., Messer W.S., Komuniecki R. Alternative-splicing of serotonin receptor isoforms in the pharynx and muscle of the parasitic nematode, Ascaris suum. Mol. Biochem. Parasitol. 1999;101:95–106. doi: 10.1016/s0166-6851(99)00059-6. [DOI] [PubMed] [Google Scholar]
- Huang X.Y., Xiao H., Rex E.B., Hobson R.J., Messer W.S., Komuniecki P.R., Komuniecki R.W. Functional characterization of alternatively spliced 5-HT2 receptor isoforms from the pharynx and muscle of the parasitic nematode, Ascaris suum. J. Neurochem. 2002;83:249–258. doi: 10.1046/j.1471-4159.2002.01067.x. [DOI] [PubMed] [Google Scholar]
- Huang S.C.C., Chan D.T.Y., Smyth D.J., Ball G., Gounaris K., Selkirk M.E. Activation of Nippostrongylus brasiliensis infective larvae is regulated by a pathway distinct from the hookworm Ancylostoma caninum. Int. J. Parasitol. 2010;40:1619–1628. doi: 10.1016/j.ijpara.2010.06.004. [DOI] [PubMed] [Google Scholar]
- Hussein A.S., Chacon M.R., Smith A.M., Tosado-Acevedo R., Selkirk M.E. Cloning, expression, and properties of a nonneuronal secreted acetylcholinesterase from the parasitic nematode Nippostrongylus brasiliensis. J. Biol. Chem. 1999;274:9312–9319. doi: 10.1074/jbc.274.14.9312. [DOI] [PubMed] [Google Scholar]
- Hussein A.S., Harel M., Selkirk M.E. A distinct family of acetylcholinesterases is secreted by Nippostrongylus brasiliensis. Mol. Biochem. Parasitol. 2002;123:125–134. [PubMed] [Google Scholar]
- Hussein A.S., Kichenin K., Selkirk M.E. Suppression of secreted acetylcholinesterase expression in Nippostrongylus brasiliensis by RNA interference. Mol. Biochem. Parasitol. 2002;122:91–94. doi: 10.1016/s0166-6851(02)00068-3. [DOI] [PubMed] [Google Scholar]
- Husson S.J., Mertens I., Janssen T., Lindemans M., Schoofs L. Neuropeptidergic signaling in the nematode Caenorhabditis elegans. Prog. Neurobiol. 2007;82:33–55. doi: 10.1016/j.pneurobio.2007.01.006. [DOI] [PubMed] [Google Scholar]
- Hwang J.M., Chang D.J., Kim U.S., Lee Y.S., Park Y.S., Kaang B.K., Cho N.J. Cloning and functional characterization of a Caenorhabditis elegans muscarinic acetylcholine receptor. Recept. Channels. 1999;6:415–424. [PubMed] [Google Scholar]
- Jagannathan S., Laughton D.L., Critten C.L., Skinner T.M., Horoszok L., Wolstenholme A.J. Ligand-gated chloride channel subunits encoded by the Haemonchus contortus and Ascaris suum orthologues of the Caenorhabditis elegans gbr-2 (avr-14) gene. Mol. Biochem. Parasitol. 1999;103:129–140. doi: 10.1016/s0166-6851(99)00120-6. [DOI] [PubMed] [Google Scholar]
- Jarecki J.L., Frey B.L., Smith L.M., Stretton A.O. Discovery of neuropeptides in the nematode Ascaris suum by database mining and tandem mass spectrometry. J. Proteome Res. 2011;10:3098–3106. doi: 10.1021/pr2001176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones A.K., Sattelle D.B. Functional genomics of the nicotinic acetylcholine receptor gene family of the nematode, Caenorhabditis elegans. Bioessays. 2004;26:39–49. doi: 10.1002/bies.10377. [DOI] [PubMed] [Google Scholar]
- Jospin M., Qi Y.B., Stawicki T.M., Boulin T., Schuske K.R., Horvitz H.R., Bessereau J.-L., Jorgensen E., Jin Y. A neuronal acetylcholine receptor regulates the balance of muscle excitation and inhibition in Caenorhabditis elegans. PLoS Biol. 2009;7:e1000265. doi: 10.1371/journal.pbio.1000265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaczorowski G.J., Knaus H.G., Leonard R.J., McManus O.B., Garcia M.L. High-conductance calcium-activated potassium channels; structure, pharmacology, and function. J. Bioenerg. Biomembr. 1996;28:255–267. doi: 10.1007/BF02110699. [DOI] [PubMed] [Google Scholar]
- Kaminsky R., Ducray P., Jung M., Clover R., Rufener L., Bouvier J., Schorderet Weber S., Wenger A., Wieland-Berghausen S., Goebel T., Gauvry N., Pautrat F., Skripsky T., Froelich O., Komoin-Oka C., Westlund B., Sluder A., Maser P. A new class of anthelmintics effective against drug-resistant nematodes. Nature. 2008;452:176–180. doi: 10.1038/nature06722. [DOI] [PubMed] [Google Scholar]
- Kaminsky R., Gauvry N., Weber S.S., Skripsky T., Bouvier J., Wenger A., Schroeder F., Desaules Y., Hotz R., Goebel T., Hosking B.C., Pautrat F., Wieland-Berghausen S., Ducray P. Identification of the amino-acetonitrile derivative monepantel (AAD 1566) as a new anthelmintic drug development candidate. Parasitol. Res. 2008;103:931–939. doi: 10.1007/s00436-008-1080-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kano T., Brockie P.J., Sassa T., Fujimoto H., Kawahara Y., Lino Y., Mellem J.E., Madsen D.M., Hosono R., Maricq A.V. Memory in Caenorhabditis elegans is mediated by NMDA-type ionotropic glutamate receptors. Curr. Biol. 2008;18:1010–1015. doi: 10.1016/j.cub.2008.05.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaplan R.M. Drug resistance in nematodes of veterinary importance: a status report. Trends Parasitol. 2004;20:477–481. doi: 10.1016/j.pt.2004.08.001. [DOI] [PubMed] [Google Scholar]
- Kaplan R.M., Nielsen M.K. An evidence-based approach to equine parasite control: it ain’t the 60s anymore. Equine Vet. Educ. 2010;22:306–316. [Google Scholar]
- Kass I.S., Wang C.C., Walrond J.P., Stretton A.O.W. Avermectin B1A, a paralysing anthelmintic that affects interneurones and inhibitory motor neurones in Ascaris. Proc. Natl. Acad. Sci. USA. 1980;77:6211–6215. doi: 10.1073/pnas.77.10.6211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keiser J., Utzinger J. Efficacy of current drugs against soil-transmitted helminth infections – systematic review and meta-analysis. J. Am. Med. Assoc. 2008;299:1937–1948. doi: 10.1001/jama.299.16.1937. [DOI] [PubMed] [Google Scholar]
- Kimber M.J., Sayegh L., El-Shehabi F., Song C.Z., Zamanian M., Woods D.J., Day T.A., Ribeiro P. Identification of an Ascaris G protein-coupled acetylcholine receptor with atypical muscarinic pharmacology. Int. J. Parasitol. 2009;39:1215–1222. doi: 10.1016/j.ijpara.2009.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kohler P. The biochemical basis of anthelmintic action and resistance. Int. J. Parasitol. 2001;31:336–345. doi: 10.1016/s0020-7519(01)00131-x. [DOI] [PubMed] [Google Scholar]
- Kohn A.B., Anderson P.A., Roberts-Misterly J.M., Greenberg R.M. Schistosome calcium channel beta subunits. Unusual modulatory effects and potential role in the action of the antischistosomal drug praziquantel. J. Biol. Chem. 2001;276:36873–36876. doi: 10.1074/jbc.C100273200. [DOI] [PubMed] [Google Scholar]
- Komuniecki R.W., Hobson R.J., Rex E.B., Hapiak V.M., Komuniecki P.R. Biogenic amine receptors in parasitic nematodes: what can be learned from Caenorhabditis elegans? Mol. Biochem. Parasitol. 2004;137:1–11. doi: 10.1016/j.molbiopara.2004.05.010. [DOI] [PubMed] [Google Scholar]
- Kopp S.R., Kotze A.C., McCarthy J.S., Traub R.J., Coleman G.T. Pyrantel in small animal medicine: 30 years on. Vet. J. 2008;178:177–184. doi: 10.1016/j.tvjl.2007.06.021. [DOI] [PubMed] [Google Scholar]
- Kotze A.C., Clifford S., O’Grady J., Behnke J.M., McCarthy J.S. An in vitro larval motility assay to determine anthelmintic sensitivity for human hookworm and Strongyloides species. Am. J. Trop. Med. Hyg. 2004;71:608–616. [PubMed] [Google Scholar]
- Kotze A.C., Le Jambre L.F., O’Grady J. A modified larval migration assay for detection of resistance to macrocyclic lactones in Haemonchus contortus, and drug screening with trichostrongylidae parasites. Vet. Parasitol. 2006;137:294–305. doi: 10.1016/j.vetpar.2006.01.017. [DOI] [PubMed] [Google Scholar]
- Kruger N., Harder A., von Samson-Himmelstjerna G. The putative cyclooctadepsipeptide receptor depsiphilin of the canine hookworm Ancylostoma caninum. Parasitol. Res. 2009;105:S91–S100. doi: 10.1007/s00436-009-1500-3. [DOI] [PubMed] [Google Scholar]
- Kwok T.C.Y., Ricker N., Fraser R., Chan A.W., Burns A., Stanley E.F., McCourt P., Cutler S.R., Roy P.J. A small-molecule screen in C. elegans yields a new calcium channel antagonist. Nature. 2006;441:91–95. doi: 10.1038/nature04657. [DOI] [PubMed] [Google Scholar]
- Laughton D.L., Lunt G.G., Wolstenholme A.J. Reporter gene constructs suggest the Caenorhabditis elegans avermectin receptor β-subunit is expressed solely in the pharynx. J. Exp. Biol. 1997;200:1509–1514. doi: 10.1242/jeb.200.10.1509. [DOI] [PubMed] [Google Scholar]
- Laughton D.L., Lunt G.G., Wolstenholme A.J. Alternative splicing of a Caenorhabditis elegans gene produces two novel inhibitory amino acid receptor subunits with identical ligand-binding domains but different ion channels. Gene. 1997;201:119–125. doi: 10.1016/s0378-1119(97)00436-8. [DOI] [PubMed] [Google Scholar]
- Lazari O., Hussein A.S., Selkirk M.E., Davidson A.J., Thompson F.J., Matthews J.B. Cloning and expression of two secretory acetylcholinesterases from the bovine lungworm, Dictyocaulus viviparus. Mol. Biochem. Parasitol. 2003;132:83–92. doi: 10.1016/j.molbiopara.2003.09.001. [DOI] [PubMed] [Google Scholar]
- Lee Y.S., Park Y.S., Chang D.J., Hwang J.M., Min C.K., Kaang B.K., Cho N.J. Cloning and expression of a G protein-linked acetylcholine receptor from Caenorhabditis elegans. J. Neurochem. 1999;72:58–65. doi: 10.1046/j.1471-4159.1999.0720058.x. [DOI] [PubMed] [Google Scholar]
- Lee Y.S., Park Y.S., Nam Y., Suh S.J., Lee F., Kaang B.K., Cho N.J. Characterization of GAR-2, a novel G protein-linked acetylcholine receptor from Caenorhabditis elegans. J. Neurochem. 2000;75:1800–1809. doi: 10.1046/j.1471-4159.2000.0751800.x. [DOI] [PubMed] [Google Scholar]
- Li C., Kim K. Neuropeptide gene families in Caenorhabditis elegans. In: Geary T.G., Maule A.G., editors. vol. 692. Springer-Verlag; Berlin: 2010. pp. 98–137. (Neuropeptide Systems as Targets for Parasite and Pest Control, Advances in Experimental Medicine and Biology). [DOI] [PubMed] [Google Scholar]
- Little P.R., Hodge A., Maeder S.J., Wirtherle N.C., Nicholas D.R., Cox G.G., Conder G.A. Efficacy of a combined oral formulation of derquantel–abamectin against the adult and larval stages of nematodes in sheep, including anthelmintic-resistant strains. Vet. Parasitol. 2011;181:180–193. doi: 10.1016/j.vetpar.2011.05.008. [DOI] [PubMed] [Google Scholar]
- Londershausen M. Approaches to new parasiticides. Pestic. Sci. 1996;48:269–292. [Google Scholar]
- Lynagh T., Lynch J.W. An improved ivermectin-activated Cl channel receptor for inhibiting electrical activity in defined neuronal populations. J. Biol. Chem. 2010;285:14890–14897. doi: 10.1074/jbc.M110.107789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lynagh T., Lynch J.W. A glycine residue essential for high ivermectin sensitivity in Cys-loop ion channel receptors. Int. J. Parasitol. 2010;40:1477–1481. doi: 10.1016/j.ijpara.2010.07.010. [DOI] [PubMed] [Google Scholar]
- Marks N.J., Maule A.G. Neuropeptides in helminths: occurrence and distribution. In: Geary T.G., Maule A.G., editors. vol. 692. Springer-Verlag; Berlin: 2010. pp. 49–77. (Neuropeptide Systems as Targets for Parasite and Pest Control, Advances in Experimental Medicine and Biology). [Google Scholar]
- Martin R.J. γ-Aminobutyric acid- and piperazine-activated single-channel currents from Ascaris suum body muscle. Br. J. Pharmacol. 1985;84:445–461. doi: 10.1111/j.1476-5381.1985.tb12929.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin R.J. Modes of action of anthelmintic drugs. Vet. J. 1997;154:11–34. doi: 10.1016/s1090-0233(05)80005-x. [DOI] [PubMed] [Google Scholar]
- Martin R.J., Robertson A.P. Mode of action of levamisole and pyrantel, anthelmintic resistance, E153 and Q57. Parasitology. 2007;134:1093–1104. doi: 10.1017/S0031182007000029. [DOI] [PubMed] [Google Scholar]
- Martin R.J., Robertson A.P. Control of nematode parasites with agents acting on neuro-musculature systems: lessons for neuropeptide ligand discovery. In: Geary T.G., Maule A.G., editors. vol. 692. Springer-Verlag; Berlin: 2010. pp. 138–154. (Neuropeptide Systems as Targets for Parasite and Pest Control, Advances in Experimental Medicine and Biology). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin R.J., Pennington A.J., Duittoz A.H., Robertson S., Kusel J.R. The physiology and pharmacology of neuromuscular transmission in the nematode parasite, Ascaris suum. Parasitology. 1991;102:S41–S58. doi: 10.1017/s0031182000073285. [DOI] [PubMed] [Google Scholar]
- Martin R.J., Bai G.X., Clark C.L., Robertson A.P. Methyridine (2-[2-methoxyethyl]-pyridine]) and levamisole activate different ACh receptor subtypes in nematode parasites: a new lead for levamisole resistance. Br. J. Pharmacol. 2004;140:1068–1076. doi: 10.1038/sj.bjp.0705528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin R.J., Clark C.L., Trailovic S.M., Robertson A.P. Oxantel is an N-type (methyridine and nicotine) agonist not an L-type (levamisole and pyrantel) agonist: classification of cholinergic anthelmintics in Ascaris. Int. J. Parasitol. 2004;34:1083–1090. doi: 10.1016/j.ijpara.2004.04.014. [DOI] [PubMed] [Google Scholar]
- Maryon E.B., Coronado R., Anderson P. Unc-68 encodes a ryanodine receptor involved in regulating C. elegans body-wall contraction. J. Cell Biol. 1996;134:885–893. doi: 10.1083/jcb.134.4.885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayer M.L. Structure and mechanism of glutamate receptor ion channel assembly, activation and modulation. Curr. Opin. Neurobiol. 2011;21:283–290. doi: 10.1016/j.conb.2011.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCavera S., Rogers A.T., Yates D.M., Woods D.J., Wolstenholme A.J. An ivermectin-sensitive glutamate-gated chloride channel from the parasitic nematode, Haemonchus contortus. Mol. Pharmacol. 2009;75:1347–1355. doi: 10.1124/mol.108.053363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKay J.P., Raizen D.M., Gottschalk A., Schafer W.R., Avery L. eat-2 and eat-18 are required for nicotinic neurotransmission in the Caenorhabditis elegans pharynx. Genetics. 2004;166:161–169. doi: 10.1534/genetics.166.1.161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKellar Q.A., Jackson F. Veterinary anthelmintics: old and new. Trends Parasitol. 2004;20:456–461. doi: 10.1016/j.pt.2004.08.002. [DOI] [PubMed] [Google Scholar]
- McVeigh P., Geary T.G., Marks N.J., Maule A.G. The FLP-side of nematodes. Trends Parasitol. 2006;22:385–396. doi: 10.1016/j.pt.2006.06.010. [DOI] [PubMed] [Google Scholar]
- Mellem J.E., Brockie P.J., Madsen D.M., Maricq A.V. Action potentials contribute to neuronal signaling in C. elegans. Nat. Neurosci. 2008;11:865–867. doi: 10.1038/nn.2131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molinari S. Natural genetic and induced plant resistance, as a control strategy to plant–parasitic nematodes alternative to pesticides. Plant Cell Rep. 2011;30:311–323. doi: 10.1007/s00299-010-0972-z. [DOI] [PubMed] [Google Scholar]
- Moreno Y., Nabhan J.F., Solomon J., Mackenzie C.D., Geary T.G. Ivermectin disrupts the function of the excretory–secretory apparatus in microfilariae of Brugia malayi. Proc. Natl. Acad. Sci. USA. 2010;107:20120–20125. doi: 10.1073/pnas.1011983107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muhlfeld S., Schmitt-Wrede H.P., Harder A., Wunderlich F. FMRFamide-like neuropeptides as putative ligands of the latrophilin-like HC110-R from Haemonchus contortus. Mol. Biochem. Parasitol. 2009;164:162–164. doi: 10.1016/j.molbiopara.2008.12.003. [DOI] [PubMed] [Google Scholar]
- Njue A.I., Hayashi J., Kinne L., Feng X.-P., Prichard R.K. Mutations in the extracellular domain of glutamate-gated chloride channel α3 and β subunits from ivermectin-resistant Cooperia oncophora affect agonist sensitivity. J. Neurochem. 2004;89:1137–1147. doi: 10.1111/j.1471-4159.2004.02379.x. [DOI] [PubMed] [Google Scholar]
- Ogilvie B.M., Rothwell T.L.W., Bremner K.C., Schitzerling H.J., Nolan J., Keith R.K. Acetylcholinesterase secretion by parasitic nematodes, 1. Evidence for secretion of the enzyme by a number of species. Int. J. Parasitol. 1973;3:589–597. doi: 10.1016/0020-7519(73)90083-0. [DOI] [PubMed] [Google Scholar]
- Osei-Atweneboana M.Y., Awadzi K., Attah S.K., Boakye D.A., Gyapong J.O., Prichard R.K. Phenotypic evidence of emerging ivermectin resistance on Onchocerca volvulus. PLoS Negl. Trop. Dis. 2011;5:e998. doi: 10.1371/journal.pntd.0000998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ottesen E.A., Hooper P.J., Bradley M., Biswas G. The global programme to eliminate lymphatic filariasis: health impact after 8 years. PLoS Negl. Trop. Dis. 2008;2:e317. doi: 10.1371/journal.pntd.0000317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pirri J.K., McPherson A.D., Donnelly J.L., Francis M.M., Akkema M.J. A tyramine-gated chloride channel coordinates distinct motor programs of a Caenorhabditis elegans escape response. Neuron. 2009;62:526–538. doi: 10.1016/j.neuron.2009.04.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Portillo V., Jagannathan S., Wolstenholme A.J. Distribution of glutamate-gated chloride channel subunits in the parasitic nematode Haemonchus contortus. J. Comp. Neurol. 2003;462:213–222. doi: 10.1002/cne.10735. [DOI] [PubMed] [Google Scholar]
- Pritchard D.I., Brown A., Toutant J.P. The molecular forms of acetylcholinesterase from Necator americanus (nematoda), a hookworm parasite of the human intestine. Eur. J. Biochem. 1994;219:317–323. doi: 10.1111/j.1432-1033.1994.tb19943.x. [DOI] [PubMed] [Google Scholar]
- Purcell J., Robertson A.P., Thompson D.P., Martin R.J. PF4, a FMRFamide-related peptide, gates low-conductance Cl− channels in Ascaris suum. Eur. J. Pharmacol. 2002;456:11–17. doi: 10.1016/s0014-2999(02)02622-5. [DOI] [PubMed] [Google Scholar]
- Putrenko I., Zakikhani M., Dent J.A. A family of acetylcholine-gated chloride channel subunits in Caenorhabditis elegans. J. Biol. Chem. 2005;280:6392–6398. doi: 10.1074/jbc.M412644200. [DOI] [PubMed] [Google Scholar]
- Qian H., Martin R.J., Robertson A.P. Pharmacology of N-, L-, and B-subtypes of nematode nAChR resolved at the single-channel level in Ascaris suum. FASEB J. 2006;20:2606. doi: 10.1096/fj.06-6264fje. [DOI] [PubMed] [Google Scholar]
- Ranganathan R., Cannon S.C., Horvitz H.R. MOD-1 is a serotonin-gated chloride channel that modulates locomotory behaviour in C. elegans. Nature. 2000;408:470–475. doi: 10.1038/35044083. [DOI] [PubMed] [Google Scholar]
- Rao U.R., Chandrashekar R., Subrahmanyam D. Effect of ivermectin on serum dependent cellular interactions to Dipetalonema viteae microfilariae. Trop. Med. Parasitol. 1987;38:123–127. [PubMed] [Google Scholar]
- Rao V.T.S., Siddiqui S.Z., Prichard R.K., Forrester S.G. A dopamine-gated ion channel (HcGGR3∗) from Haemonchus contortus is expressed in the cervical papillae and is associated with macrocyclic lactone resistance. Mol. Biochem. Parasitol. 2009;166:54–61. doi: 10.1016/j.molbiopara.2009.02.011. [DOI] [PubMed] [Google Scholar]
- Rao V.T.S., Accardi M.V., Siddiqui S.Z., Beech R.N., Prichard R.K., Forrester S.G. Characterization of a novel tyramine-gated chloride channel from Haemonchus contortus. Mol. Biochem. Parasitol. 2010;173:64–68. doi: 10.1016/j.molbiopara.2010.05.005. [DOI] [PubMed] [Google Scholar]
- Raymond V., Mongan N.P., Sattelle D.B. Anthelmintic actions on homomer forming nicotinic acetylcholine receptor subunits: chicken alpha 7 and ACR-16 from the nematode Caenorhabditis elegans. Neuroscience. 2000;101:785–791. doi: 10.1016/s0306-4522(00)00279-7. [DOI] [PubMed] [Google Scholar]
- Richmond J.E., Jorgensen E.M. One GABA and two acetylcholine receptors function at the C. elegans neuromuscular junction. Nat. Neurosci. 1999;2:791–797. doi: 10.1038/12160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ringstad N., Abe N., Horvitz H.R. Ligand-gated chloride channels are receptors for biogenic amines in C. elegans. Science. 2009;325:96–100. doi: 10.1126/science.1169243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robertson A.P., Clark C.L., Burns T.A., Thompson D.P., Geary T.G., Trailovic S.M., Martin R.J. Paraherquamide and 2-deoxy-paraherquamide distinguish cholinergic receptor subtypes in Ascaris muscle. J. Pharmacol. Exp. Ther. 2002;302:853–860. doi: 10.1124/jpet.102.034272. [DOI] [PubMed] [Google Scholar]
- Rohrer S.P., Birzin E.T., Eary C.H., Schaeffer J.M., Shoop W.L. Ivermectin binding sites in sensitive and resistant Haemonchus contortus. J. Parasitol. 1994;80:493–497. [PubMed] [Google Scholar]
- Rolfe R.N., Barrett J., Perry R.N. Electrophysiological analysis of responses of adult females of Brugia pahangi to some chemicals. Parasitology. 2001;122:347–357. doi: 10.1017/s0031182001007375. [DOI] [PubMed] [Google Scholar]
- Rufener L., Maser P., Roditi I., Kaminsky R. Haemonchus contortus acetylcholine receptors of the DEG-3 subfamily and their role in sensitivity to monepantel. PLoS Pathog. 2009;5:e1000380. doi: 10.1371/journal.ppat.1000380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rufener L., Baur R., Kaminsky R., Maser P., Sigel E. Monepantel allosterically activates DEG-3/DES-2 channels of the gastrointestinal nematode Haemonchus contortus. Mol. Pharmacol. 2010;78:895–902. doi: 10.1124/mol.110.066498. [DOI] [PubMed] [Google Scholar]
- Rufener L., Keiser J., Kaminsky R., Maser P., Nilsson D. Phylogenomics of ligand-gated ion channels predicts monepantel effect. PLoS Pathog. 2010;6:e1001091. doi: 10.1371/journal.ppat.1001091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruiz-Lancheros E., Viau C., Walter T.N., Francis A., Geary T.G. Activity of novel nicotinic anthelmintics in cut preparations of Caenorhabditis elegans. Int. J. Parasitol. 2011;41:455–461. doi: 10.1016/j.ijpara.2010.11.009. [DOI] [PubMed] [Google Scholar]
- Saeger B., Schmitt-Wrede H.P., Dehnhardt M., Benten W.P.M., Krucken J., Harder A., von Samson-Himmelstjerna G., Wiegand H., Wunderlich F. Latrophilin-like receptor from the parasitic nematode Haemonchus contortus as target for the anthelmintic depsipeptide PF1022A. FASEB J. 2001;15:1332–1334. doi: 10.1096/fj.00-0664fje. [DOI] [PubMed] [Google Scholar]
- Sager H., Hosking B., Bapst B., Stein P., Vanhoff K., Kaminsky R. Efficacy of the amino-acetonitrile derivative, monepantel, against experimental and natural adult stage gastro-intestinal nematode infections in sheep. Vet. Parasitol. 2009;159:49–54. doi: 10.1016/j.vetpar.2008.10.006. [DOI] [PubMed] [Google Scholar]
- Salkoff L., Butler A., Ferreira G., Santi C., Wei A. High-conductance potassium channels of the SLO family. Nat. Rev. Neurosci. 2006;7:921–931. doi: 10.1038/nrn1992. [DOI] [PubMed] [Google Scholar]
- Sasser J.N. Root-knot nematodes – a global menace to crop production. Plant Dis. 1980;64:36–41. [Google Scholar]
- Sattelle D.B. Invertebrate nicotinic acetylcholine receptors – targets for chemicals and drugs important in agriculture, veterinary medicine and human health. J. Pestic. Sci. 2009;34:233–240. [Google Scholar]
- Sattelle D.B., Cordova D., Cheek T.R. Insect ryanodine receptors: molecular targets for novel pest control chemicals. Invert. Neurosci. 2008;8:107–119. doi: 10.1007/s10158-008-0076-4. [DOI] [PubMed] [Google Scholar]
- Segerberg M.A., Stretton A.O.W. Actions of cholinergic drugs in the nematode Ascaris suum – complex pharmacology of muscle and motor-neurons. J. Gen. Physiol. 1993;101:271–296. doi: 10.1085/jgp.101.2.271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selkirk M.E., Lazari O., Matthews J.B. Functional genomics of nematode acetylcholinesterases. Parasitology. 2005;131:S3–S18. doi: 10.1017/S0031182005008206. [DOI] [PubMed] [Google Scholar]
- Sheriff J.C., Kotze A.C., Sangster N.C., Martin R.J. Effects of macrocyclic lactone anthelmintics on feeding and pharyngeal pumping in Trichostrongylus colubriformisin vitro. Parasitology. 2002;125:477–484. doi: 10.1017/s0031182002002251. [DOI] [PubMed] [Google Scholar]
- Sheriff J.C., Kotze A.C., Sangster N.C., Hennessy D.R. Effect of ivermectin on feeding by Haemonchus contortusin vivo. Vet. Parasitol. 2005;128:341–346. doi: 10.1016/j.vetpar.2004.12.008. [DOI] [PubMed] [Google Scholar]
- Silva J.P., Suckling J., Ushkaryov Y. Penelope’s web: using α-latrotoxin to untangle the mysteries of exocytosis. J. Neurochem. 2009;111:275–290. doi: 10.1111/j.1471-4159.2009.06329.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith M.W., Borts T.L., Emkey R., Cook C.A., Wiggins C.J., Gutierrez J.A. Characterization of a novel G-protein coupled receptor from the parasitic nematode H. contortus with high affinity for serotonin. J. Neurochem. 2003;86:255–266. doi: 10.1046/j.1471-4159.2003.01849.x. [DOI] [PubMed] [Google Scholar]
- Smith K.A., Rex E.B., Komuniecki R.W. Are Caenorhabditis elegans receptors useful targets for drug discovery: pharmacological comparison of tyramine receptors with high identity from C. elegans (TYRA-2) and Brugia malayi (Bm4) Mol. Biochem. Parasitol. 2007;154:52–61. doi: 10.1016/j.molbiopara.2007.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smout M.J., Kotze A.C., McCarthy J.S., Loukas A. A novel high throughput assay for anthelmintic drug screening and resistance diagnosis by real-time monitoring of parasite motility. PLoS Negl. Trop. Dis. 2010;4:e885. doi: 10.1371/journal.pntd.0000885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sonnichsen B., Koski L.B., Walsh A., Marschall P., Neumann B., Brehm M., Alleaume A.M., Artelt J., Bettencourt P., Cassin E., Hewitson M., Holz C., Khan M., Lazik S., Martin C., Nitzsche B., Ruer M., Stamford J., Winzi M., Heinkel R., Roder M., Finell J., Hantsch H., Jones S.J.M., Jones M., Piano F., Gunsalus K.C., Oegema K., Gonczy P., Coulson A., Hyman A.A., Echeverri C.J. Full-genome RNAi profiling of early embryogenesis in Caenorhabditis elegans. Nature. 2005;434:462–469. doi: 10.1038/nature03353. [DOI] [PubMed] [Google Scholar]
- Steinmann P., Zhou X.N., Du Z.W., Jiang J.Y., Xiao S.H., Wu Z.X., Zhou H., Utzinger J. Tribendimidine and albendazole for treating soil-transmitted helminths, Strongyloides stercoralis and Taenia spp.: open-label randomized trial. PLoS Negl. Trop. Dis. 2008;2:e322. doi: 10.1371/journal.pntd.0000322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sutherland I.A., Leathwick D.M. Anthelmintic resistance in nematode parasites of cattle: a global issue? Trends Parasitol. 2011;27:176–181. doi: 10.1016/j.pt.2010.11.008. [DOI] [PubMed] [Google Scholar]
- Tandon R., LePage K.T., Kaplan R.M. Cloning and characterization of genes encoding alpha and beta subunits of glutamate-gated chloride channel protein in Cylicocyclus nassatus. Mol. Biochem. Parasitol. 2006;150:46–55. doi: 10.1016/j.molbiopara.2006.06.007. [DOI] [PubMed] [Google Scholar]
- Thompson A.J., Lester H.A., Lummis S.C.R. The structural basis of function in Cys-loop receptors. Quart. Rev. Biophys. 2010;43:449–499. doi: 10.1017/S0033583510000168. [DOI] [PubMed] [Google Scholar]
- Tompkins J.B., Stitt L.E., Ardelli B.F. Brugia malayi: in vitro effects of ivermectin and moxidectin on adults and microfilariae. Exp. Parasitol. 2010;124:394–402. doi: 10.1016/j.exppara.2009.12.003. [DOI] [PubMed] [Google Scholar]
- Touroutine D., Fox R.M., Von Stetina S.E., Burdina A., Miller D.M., Richmond J.E. acr-16 encodes an essential subunit of the levamisole-resistant nicotinic receptor at the Caenorhabditis elegans neuromuscular junction. J. Biol. Chem. 2005;280:27013–27021. doi: 10.1074/jbc.M502818200. [DOI] [PubMed] [Google Scholar]
- Treinin M., Chalfie M. A mutated acetylcholine receptor subunit causes neuronal degeneration in C. elegans. Neuron. 1995;14:871–877. doi: 10.1016/0896-6273(95)90231-7. [DOI] [PubMed] [Google Scholar]
- Trim J.E., Holden-Dye L., Willson J., Lockyer M., Walker R.J. Characterization of 5-HT receptors in the parasitic nematode, Ascaris suum. Parasitology. 2001;122:207–217. doi: 10.1017/s003118200100703x. [DOI] [PubMed] [Google Scholar]
- Unwin N. Refined structure of the nicotinic acetylcholine receptor at 4 Å resolution. J. Mol. Biol. 2005;346:967–989. doi: 10.1016/j.jmb.2004.12.031. [DOI] [PubMed] [Google Scholar]
- Vassilatis D.K., Arena J.P., Plasterk R.H.A., Wilkinson H., Schaeffer J.M., Cully D.F., van der Ploeg L.H.T. Genetic and biochemical evidence for a novel avermectin sensitive chloride channel in C. elegans isolation and characterisation. J. Biol. Chem. 1997;272:33167–33174. doi: 10.1074/jbc.272.52.33167. [DOI] [PubMed] [Google Scholar]
- Vassilatis D.K., Elliston K.O., Paress P.S., Hamelin M., Arena J.P., Schaeffer J.M., van der Ploeg L.H.T., Cully D.F. Evolutionary relationship of the ligand-gated ion channels and the avermectin-sensitive, glutamate-gated chloride channels. J. Mol. Evol. 1997;44:501–508. doi: 10.1007/pl00006174. [DOI] [PubMed] [Google Scholar]
- Vercruysse, J., Albonico, M., Behnke, J., Kotze, A., Prichard, R., McCarthy, J.S., Montresor, A., Levecke, B., 2011. Is anthelmintic resistance a concern for the control of human soil-transmitted helminths? Int. J. Parasitol. – Drugs Drug Resist. 1, in press. doi:10.1016/j.ijpddr.2011.09.002. [DOI] [PMC free article] [PubMed]
- Verma S., Robertson A.P., Martin R.J. The nematode neuropeptide, AF2 (KHEYLRF-NH2), increases voltage-activated calcium currents in Ascaris suum muscle. Br. J. Pharmacol. 2007;151:888–899. doi: 10.1038/sj.bjp.0707296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verma S., Robertson A.P., Martin R.J. Effects of SDPNFLRF-amide (PF1) on voltage-activated currents in Ascaris suum muscle. Int. J. Parasitol. 2009;39:315–326. doi: 10.1016/j.ijpara.2008.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wagland B.M., Jones W.O., Hribar L., Bendixsen T., Emery D.L. A new simplified assay for larval migration-inhibition. Int. J. Parasitol. 1992;22:1183–1185. doi: 10.1016/0020-7519(92)90040-r. [DOI] [PubMed] [Google Scholar]
- Walker R.J., Papaioannou S., Holden-Dye L. A review of FMRFamide- and RFamide-like peptides in metazoa. Invert. Neurosci. 2009;9:111–153. doi: 10.1007/s10158-010-0097-7. [DOI] [PubMed] [Google Scholar]
- Welz C., Kruger N., Schniederjans M., Miltsch S.M., Krucken J., Guest M., Holden-Dye L., Harder A., von Samson-Himmelstjerna G. SLO-1-channels of parasitic nematodes reconstitute locomotor behaviour and emodepside sensitivity in Caenorhabditis elegansslo-1 loss of function mutants. PLoS Pathog. 2011;7:e1001330. doi: 10.1371/journal.ppat.1001330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White J.G., Southgate E., Thomson J.N., Brenner S. The structure of the nervous system of the nematode Caenorhabditis elegans. Philos. Trans. R. Soc. Lond. Ser. B. 1986;314:1–340. doi: 10.1098/rstb.1986.0056. [DOI] [PubMed] [Google Scholar]
- White W.H., Gutierrez J.A., Naylor S.A., Cook C.A., Gonzalez I.C., Wisehart M.A., Smith C.K., Thompson W.A. In vitro and in vivo characterization of p-amino-phenethyl-m-trifluoromethylphenyl piperazine (PAPP), a novel serotonergic agonist with anthelmintic activity against Haemonchus contortus, Teladorsagia circumcincta and Trichostrongylus colubriformis. Vet. Parasitol. 2007;146:58–65. doi: 10.1016/j.vetpar.2007.02.014. [DOI] [PubMed] [Google Scholar]
- Williamson S.M., Walsh T.K., Wolstenholme A.J. The Cys-loop ligand-gated ion channel gene family of Brugia malayi and Trichinella spiralis: a comparison with Caenorhabditis elegans. Invert. Neurosci. 2007;7:219–226. doi: 10.1007/s10158-007-0056-0. [DOI] [PubMed] [Google Scholar]
- Williamson S.M., Robertson A.P., Brown L., Williams T., Woods D.J., Martin R.J., Sattelle D.B., Wolstenholme A.J. The nicotinic acetylcholine receptors of the parasitic nematode Ascaris suum: formation of two distinct drug targets by varying the relative expression levels of two subunits. PLoS Pathog. 2009;5:e1000517. doi: 10.1371/journal.ppat.1000517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willson J., Amliwala K., Harder A., Holden-Dye L., Walker R.J. The effect of the anthelmintic emodepside at the neuromuscular junction of the parasitic nematode Ascaris suum. Parasitology. 2003;126:79–86. doi: 10.1017/s0031182002002639. [DOI] [PubMed] [Google Scholar]
- Willson J., Amliwala K., Davis A., Cook A., Cuttle M.F., Kriek N., Hopper N.A., O’Connor V., Harder A., Walker R.J., Holden-Dye L. Latrotoxin receptor signaling engages the UNC-13-dependent vesicle-priming pathway in C. elegans. Curr. Biol. 2004;14:1374–1379. doi: 10.1016/j.cub.2004.07.056. [DOI] [PubMed] [Google Scholar]
- Wolstenholme A.J., Rogers A.T. Glutamate-gated chloride channels and the mode of action of the avermectin/milbemycin anthelmintics. Parasitology. 2005;131:S85–S95. doi: 10.1017/S0031182005008218. [DOI] [PubMed] [Google Scholar]
- Wolstenholme A.J., Fairweather I., Prichard R.K., von Samson-Himmelstjerna G., Sangster N. Drug resistance in veterinary helminths. Trends Parasitol. 2004;20:469–476. doi: 10.1016/j.pt.2004.07.010. [DOI] [PubMed] [Google Scholar]
- Wolstenholme A.J., Williamson S.M., Reaves B.J. TRP channels in parasites. In: Islam M.S., editor. vol. 704. Springer-Verlag; Berlin: 2011. pp. 359–371. (Transient Receptor Potential Channels, Advances in Experimental Medicine and Biology). [DOI] [PubMed] [Google Scholar]
- Woods D.J., Knauer C.S. Discovery of veterinary antiparasitic agents in the 21st century: a view from industry. Int. J. Parasitol. 2010;40:1177–1181. doi: 10.1016/j.ijpara.2010.04.005. [DOI] [PubMed] [Google Scholar]
- Woods D.J., Vaillancourt V.A., Wendt J.A., Meeus P.F. Discovery and development of veterinary antiparasitic drugs: past, present and future. Future Med. Chem. 2011;3:887–896. doi: 10.4155/fmc.11.39. [DOI] [PubMed] [Google Scholar]
- Wu S.N., Wu A.Z., Lin M.W. Pharmacological roles of the large-conductance calcium-activated potassium channel. Curr. Top Med. Chem. 2006;6:1025–1030. doi: 10.2174/156802606777323764. [DOI] [PubMed] [Google Scholar]
- Yassin L., Gillo B., Kahan T., Halevi S., Eshel M., Treinin M. Characterization of the DEG-3/DES-2 receptor: a nicotinic acetylcholine receptor that mutates to cause neuronal degeneration. Mol. Cell Neurosci. 2001;17:589–599. doi: 10.1006/mcne.2000.0944. [DOI] [PubMed] [Google Scholar]
- Yates D.M., Wolstenholme A.J. An ivermectin-sensitive glutamate-gated chloride channel subunit from Dirofilaria immitis. Int. J. Parasitol. 2004;34:1075–1081. doi: 10.1016/j.ijpara.2004.04.010. [DOI] [PubMed] [Google Scholar]
- Zahner H., Schmidtchen D., Mutasa J.A. Ivermectin-induced killing of microfilariae in vitro by neutrophils mediated by NO. Exp. Parasitol. 1997;86:110–117. doi: 10.1006/expr.1997.4160. [DOI] [PubMed] [Google Scholar]