

Abstract
The causes of multiple myeloma (MM) remain obscure and there are few known risk factors; however, natural killer T (NKT) cell abnormalities have been reported in patients with MM, and therapeutic targeting of NKT cells is promoted as a potential treatment. We characterized NKT cell defects in treated and untreated patients with MM and determined the impact of lenalidomide therapy on the NKT cell pool. Lenalidomide is an immunomodulatory drug with co-stimulatory effects on NKT cells in vitro and is an approved treatment for MM, although its mode of action in that context is not well defined. We find that patients with relapsed/progressive MM had a marked deficiency in NKT cell numbers. In contrast, newly diagnosed patients had relatively normal NKT cell frequency and function prior to treatment, although a specific NKT cell deficiency emerged after high-dose melphalan and autologous stem cell transplantation (ASCT) regimen. This also impacted NK cells and conventional T cells, but the recovery of NKT cells was considerably delayed, resulting in a prolonged, treatment-induced NKT cell deficit. Longitudinal analysis of individual patients revealed that lenalidomide therapy had no in-vivo impact on NKT cell numbers or cytokine production, either as induction therapy, or as maintenance therapy following ASCT, indicating that its clinical benefits in this setting are independent of NKT cell modulation.
Keywords: lenalidomide, multiple myeloma, NKT cells
Introduction
Natural killer T (NKT) cells are CD1d-restricted T cells that recognize lipid-based antigens. They are capable of rapidly releasing cytokines that affect a wide range of innate and adaptive immune responses, including against cancer 1,2. Several subpopulations of NKT cells have been identified, but the most widely studied are type 1 NKT cells, which express an evolutionarily conserved semi-invariant T cell receptor (TCR) 1–4. Defects in type 1 NKT cells have been associated with multiple myeloma (MM) and are the focus of this study, so for simplicity we will refer to them as NKT cells from this point onwards.
Defects in the NKT cell pool are associated with autoimmune diseases 5–6, allergies 7 and many forms of cancer 8–14, but there is an emerging focus on NKT cells in the context of multiple myeloma (MM), where immune dysfunction has been identified as potentially important in disease predisposition and progression 8–16. Defective interferon (IFN)-γ production by NKT cells and a trend towards lower NKT cell frequencies reportedly affect patient groups with MM and are also observed in animal models of MM 8,12. As a result, many studies have nominated NKT cells as a potential new target for treatment of MM in humans 12–20.
The mechanism by which NKT cells may protect against MM development is not defined, although several studies link these cells to protective anti-myeloma responses. For example, NKT cell lines isolated from patients with MM respond to primary MM cells pulsed with α-galactosylceramide (α-GalCer) (a potent NKT cell agonist), resulting in cytokine release and lysis of the myeloma cells 19. The CD1d antigen-presenting molecule recognized by NKT cells is highly expressed in premalignant and early myeloma plasma cells, and progression of MM is associated with reduced CD1d expression by myeloma cells 21. Engagement of CD1d (NKT ligand) also kills tumour cells in vitro 21, and NKT cells specific for an inflammation-associated lipid antigen, lysophosphatidylcholine, have been detected in patients with advanced myeloma 22. Collectively, these findings indicate why a defective NKT cell pool may be associated with MM onset and relapse and support the concept that therapeutic targeting of NKT cells may be a useful strategy in treatment of MM in humans 12–23.
The immunomodulatory drug lenalidomide (Revlimid®, Celgene Corp., Summit, NJ, USA) has been approved as a treatment for MM 24–25, although its exact mode of action is not well understood. Lenalidomide could potentially impact on MM through several mechanisms, including inhibition of angiogenesis, modulation of plasma cell proliferation and promoting immune cell adhesion and activation 25–26. Lenalidomide also promotes T cell and NK cell activation 27, and it has specific stimulatory effects on NKT cells that are independent of conventional T cells 12–28. These include enhancing NKT cell proliferation and cytokine production, and promoting a T helper type 1 (Th1) cytokine profile 18–31.
The well-documented in-vitro impact of lenalidomide treatment on NKT cells is consistent with a report showing increased frequency and cytokine responsiveness of NKT cells from patients treated with lenalidomide 32. Only two patients with MM were examined in that study, but a more recent analysis of patients with asymptomatic myeloma treated with lenalidomide in combination with αGalCer loaded monocyte-derived dendritic cells (DCs) also identified activated NKT cells and reduced serum paraprotein 20. Although it was difficult to isolate the impact of the lenalidomide from the transferred DCs in that study, the study speculated that combination therapies targeting NKT cells could help to prevent disease progression in humans.
We and others have argued that although these data are promising, more knowledge is required about whether NKT cell defects contribute to MM in humans, and whether NKT cell agonists (possibly including lenalidomide) are viable approaches to anti-MM treatment. In this study, we present findings from a longitudinal analysis of NKT cells from a clinical trial exploring the efficacy of lenalidomide therapy in newly diagnosed patients with untreated MM. These results were compared to patients enrolled in a lenalidomide clinical trial for MM, which had relapsed, or was refractory to prior anti-MM therapy; and to a control group of healthy donors. We characterized the frequency and functional defects of NKT cells patients with MM, and determined their NKT cell response to lenalidomide therapy.
Materials and methods
Trial and study design
Anti-coagulated whole blood from healthy donors was obtained from the Australian Red Cross Blood Bank Service (Southbank, Melbourne, Australia). Patient samples were obtained from two clinical trials: (1) the Revlite trial (RL NCT00482261) in patients with either relapsed/refractory MM evaluating the effects of low-dose lenalidomide (Revlimid; Celgene) (15 mg D1-21) with high-dose dexamethasone (20 mg d1-4, 9–11 and 17–21) [these patients had been treated previously with chemotherapy and ASCT (for most)]; and (2) the LitVacc trial (ACTRN12613000344796) in patients with newly diagnosed MM undergoing four cycles of induction with low-dose lenalidomide (15 mg D1-21), low-dose dexamethasone (20 mg weekly) followed by high-dose cyclophosphamide and granulocyte colony-stimulating factor (G-CSF) stem cell mobilization and high-dose melphalan (200 mg/m2) AuSCT followed by lenalidomide maintenance commencing on D21-35 post-transplant (25 mg d1-21/28-day cycle) and DC vaccination with autologous DC loaded with primary MM cell lysate. All enrolled patients in either trial had active MM requiring treatment. Patients with monoclonal gammopathy of undetermined significance (MGUS) or smouldering MM were not included. The trials were approved by the Peter MacCallum Centre Human Research Ethics committee and are registered on http://ClinicalTrials.gov.
Sample processing and storage
Serial blood samples were obtained as per the study protocol. Samples from patients in the Revlite trial were taken at enrolment only and samples from patients in the LitVacc trial were taken at enrolment and on day 1 of lenalidomide induction cycles (C) 2 and 3, at the end of the induction (EOI) immediately prior to ASCT, 21 days post-ASCT and on day 1 of maintenance (M) cycles 2, 4 and 6 (Supporting information, Figure S1).
Peripheral blood mononuclear cells (PBMCs) were isolated from whole anti-coagulated blood by gradient centrifugation by Histopaque (density 1·077 g/ml; Sigma-Aldrich, St Louis, MO, USA) and then cryopreserved in 10% dimethyl sulphoxide and 90% fetal bovine serum at −80°C prior to storage in liquid nitrogen for later batch analysis.
Antibodies and flow cytometry
Fluorochrome-labelled antibodies used for flow cytometry [fluorescein isothiocyanate (FITC)-conjugated anti-CD25, anti-immunoglobulin (Ig)G1 and anti-IFN-γ (4S.B3), phycoerythrin-cyanin 7 (PE-Cy7)-conjugated anti-CD3 (SK7), allophycocyanin (APC)-H7-conjugated anti-CD8 (SK1), APC-conjugated anti-IL-4 (MP4-25D2) and anti-IgG1, AlexaFluor 700-conjugated anti-CD56 and anti-tumour necrosis factor (TNF) (MAb11) and Pacific Blue-conjugated anti-CD4 (RPA-T4)] were purchased from BD Biosciences (San Diego, CA, USA). APC-conjugated anti-CD161 (191B8) was purchased from Miltenyi Biotech (Bergisch Gladbach, Germany). PE-conjugated αGalCer-loaded mouse CD1d tetramer was produced in-house by Konstantinos Kyparissoudis (University of Melbourne) using a construct provided originally by Mitchell Kronenberg, La Jolla Institute for Allergy and Immunology. Flow cytometry data were acquired on a LSRII (BD Biosciences) and analysed using FlowJo software (Treestar Inc., Ashland, OR, USA). Autofluorescent cells, doublets, non-specifically stained cells and dead cells were excluded using 7-aminoactinomycin D (Invitrogen Life Technologies, Carlsbad, CA, USA) and vehicle-loaded CD1d tetramer.
PBMC stimulation
Cells were cultured in 12-well plates in RPMI-1640 medium (Invitrogen Life Technologies) supplemented with 10% heat-inactivated fetal bovine serum (JRH Biosciences, St Lenexa, KS, USA), 100 U penicillin (Invitrogen Life Technologies), 100 μg/ml streptomycin (Invitrogen Life Technologies), 2 mM glutamax (Invitrogen Life Technologies), 1 mM sodium pyruvate (Invitrogen Life Technologies), 15 mM HEPES (Invitrogen Life Technologies), 0·1 mM non-essential amino acids (Invitrogen Life Technologies) and 50 μM 2-mercaptoethanol (Sigma-Aldrich) at 37°C and 5% CO2. For in-vitro stimulation, 10 ng/ml phorbol 12-myristate 13-acetate (PMA; Sigma-Aldrich) and 1 μg/ml ionomycin (Sigma-Aldrich) for 4 h with 2 μM monensin (GolgiStop; BD Biosciences). After stimulation, cells were prepared for intracellular analysis of IFN-γ, TNF and IL-4 using the Cytofix/Cytoperm staining kit (BD Biosciences).
Statistical analyses
Statistical analyses were performed with GraphPad Prism version 5.0 software (GraphPad Software Inc., San Diego, CA, USA) using the Mann–Whitney test or Kruskal–Wallis test with Dunn's multiple comparisons post-test as appropriate.
Results
Relapsed/refractory MM patients (RMM) have lower NKT cell frequencies than healthy donors
We analysed T and NKT cells from peripheral blood of 24 patients with relapsed/refractory MM (RMM) and compared them to samples from 47 healthy blood donors, some of whom were also part of an earlier study 29. RMM patients had a normal distribution of CD4+, CD4− and CD8+ subsets, but a statistically significant NKT cell deficiency (Fig. 1). The conventional T cell pool of RMM patients was also moderately deficient, but NKT cells remained selectively deficient even when expressed relative to T cells (rather than as a percentage of PBMCs), which indicates a specific NKT cell deficiency in RMM patients.
Figure 1.
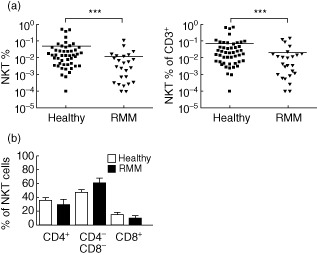
Natural killer (NK) T cell deficiency in pretreated multiple myeloma patients. Peripheral blood mononuclear cells (PBMCs) isolated from blood collected from relapsed or refractory multiple myeloma (MM) patients (RMM) (n = 24) and healthy donors (n = 47) were stained with CD3 and αGalCer-loaded CD1d tetramer to detect NKT cells. (a) RMM patients had a significantly lower NKT cell frequency compared to healthy controls, irrespective of whether the frequency was expressed as a proportion of overall lymphocytes or of CD3+ T cells. (b) The distribution of CD4+, DN and CD8+ NKT cell subsets are shown for RMM patients and healthy donors.
NKT cell frequency in patients with newly diagnosed MM prior to treatment
Given the significant NKT cell deficit in RMM patients, it was important to investigate whether or not this was related directly to MM. We therefore compared the RMM group to newly diagnosed MM patients who had not commenced treatment and to healthy donors (Fig. 2). Analysis of the T cell and NKT cell pools showed that newly diagnosed patients had a normal NKT cell frequency. The overall frequency of conventional T cell levels in untreated patients with MM and the ratio between CD4+ and CD8+ T cells was comparable to that of healthy subjects (Fig. 2b,c). Interestingly, there was a trend towards an abnormal ratio between CD4+ and CD4− NKT cell subsets in untreated patients with MM, reflecting a comparative deficit of the CD4− NKT cell subset that is regarded as the most important for anti-cancer activities 4–36; however, this was variable and not statistically significant.
Figure 2.
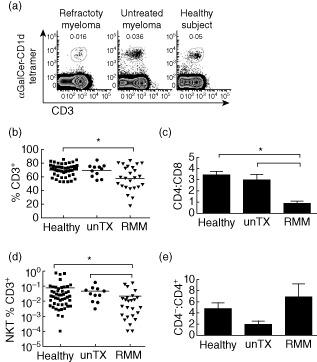
T and natural killer (NK) T cell frequency in patients with multiple myeloma (MM). (a) Representative flow cytometry plots of NKT cells from refractory MM (RMM) (n = 24), untreated patients with MM (n = 11) and healthy controls (n = 47) are shown. (b) Individual peripheral blood T cell frequencies for RMM, untreated MM and healthy donors are shown. (c) The CD4+ T cell: CD8+ T cell ratio was assessed for each group. (d) Individual peripheral blood NKT cell frequencies for RMM, untreated MM and healthy donors are shown. The dotted line represents a NKT % below the threshold of detection. (e) The ratio of CD4− NKT: CD4+ NKT cells were also assessed. Means of each group were compared using the Kruskal–Wallis statistical test with Dunn's multiple comparison post-test.
NKT cell frequency after lenalidomide and ASCT
Our data indicated that NKT cell numbers were relatively normal in newly diagnosed patients with MM, suggesting that the deficit we observed in the group of relapsed or progressive patients with MM may have been due to the treatment of the disease rather than MM itself. To examine this directly and to measure the impact of standard therapy with lenalidomide, we conducted longitudinal analysis of T cells, NK cells and NKT cells in 11 MM patients throughout treatment as part of the LitVacc trial (Fig. 3). Blood samples were taken prior to treatment and throughout a treatment course of lenalidomide and dexamethasone induction therapy, ASCT and subsequent lenalidomide–DC vaccination maintenance therapy (DC vaccination was given to 10 of 11 patients).
Figure 3.
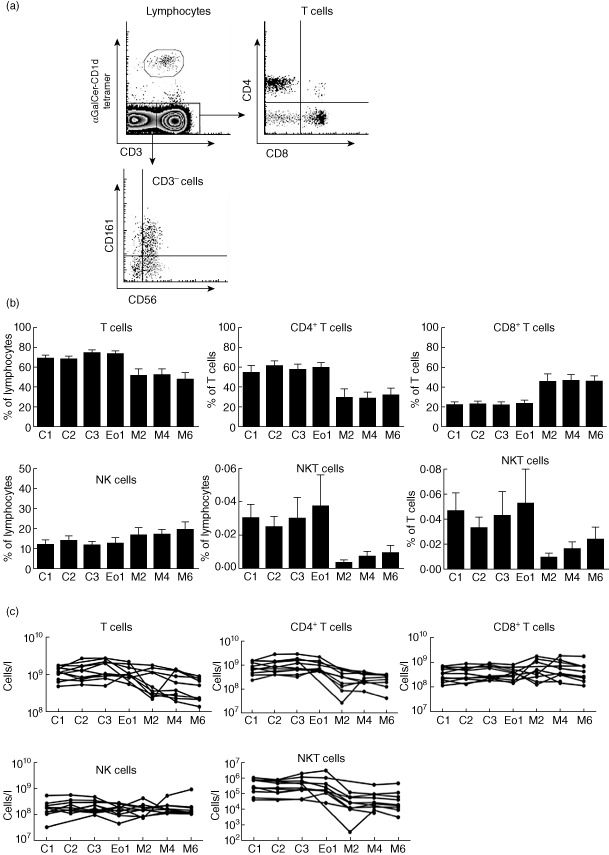
Poor recovery of natural killer (NK) T cells in untreated multiple myeloma (MM) patients after autologous stem cell transplant. Peripheral blood mononuclear cells (PBMCs) were isolated from blood collected from untreated MM patients prior to treatment (C1), after two cycles of lenalidomide induction (LEN) (C2), after three cycles of LEN induction (C3), at the end of induction (EoI), two maintenance cycles post-AuSCT (M2), four maintenance cycles post-autologous stem cell transplantation (ASCT) (M4) and six maintenance cycles post-ASCT (M6). (a) Representative fluorescence activated cell sorter (FACS) plots demonstrate how NKT cell subsets are analysed. (b) The frequency of T cells, NK cells and NKT cells were assessed at each time-point. CD4+ T cells and CD8+ T cells are expressed as a proportion of T cells. T cells, NK cells and NKT cells are expressed as a proportion of lymphocytes and of T cells as indicated. (c) T cells, NK cells and NKT cell for individual untreated MM patients are shown.
These data showed that lenalidomide induction therapy did not result in a significant increase in the frequency or absolute number of NKT cells (Fig. 3b,c). At the end of induction (EoI) patients underwent ASCT, which caused an expected decline in T cell frequency of most patients (Fig. 3b,c). A selective reduction in frequencies and absolute numbers of circulating CD4+ T cells was also observed after ASCT (Fig. 3b,c), coinciding with a proportional increase in CD8 T cells (Fig. 3b,c). Proportions and absolute numbers of NK cells did not change significantly post-ASCT (Fig. 3b,c), in keeping with the typically rapid recovery of this lymphocyte population post-ASCT.
In contrast to conventional T cells, the NKT cell pool underwent striking post-ASCT changes (Fig. 3b,c). The proportions and absolute numbers of NKT cells were greatly decreased, did not recover to pre-ASCT during lenalidomide maintenance therapy and were decreased more severely than for conventional CD4+ and CD8+ T cells and NK cells (Fig. 3b,c).
Longitudinal analysis of NKT cell subsets
Having identified a severe and prolonged NKT cell deficiency in patients with MM after ASCT therapy, we examined separately the impact on functionally distinct NKT cell subsets. We analysed NKT cell subsets defined by expression of CD4 and CD8, NK markers such as CD161 and CD56 and the CD25 activation marker in patients with MM during induction therapy, after ASCT and during maintenance therapy (Figs 3c and 4a) 4–37. Induction therapy did not alter significantly the composition of the NKT cell compartment, and the proportions and numbers of different NKT cell subsets remained relatively consistent (Fig. 4b,c). Analysis of five patients post-ASCT revealed a decline in numbers across all NKT cell subsets and none recovered fully during maintenance therapy (Fig. 4c). CD25 expression was uniformly low on all groups and at all time-points, indicating that NKT cells were not activated in these patients (Fig. 4a and data not shown).
Figure 4.
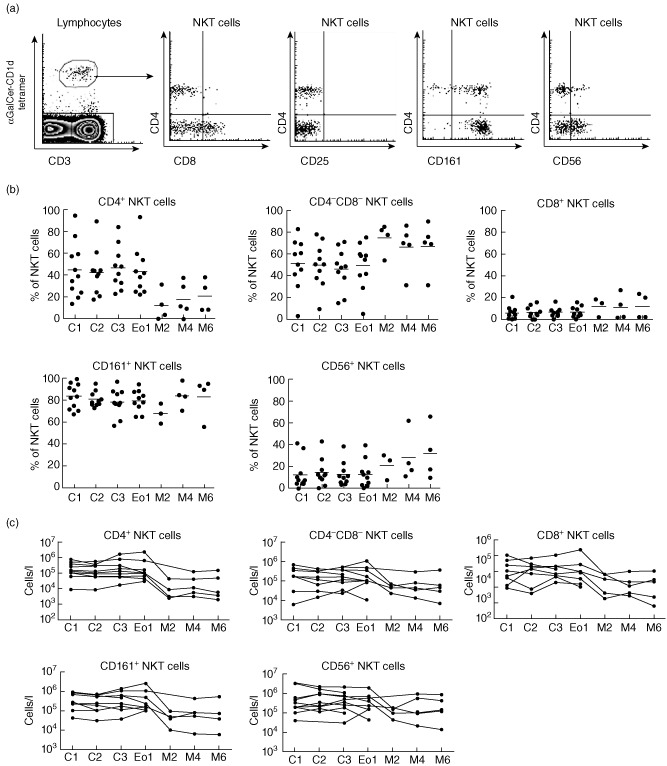
The natural killer (NK) T cell compartment is deficient following autologous stem cell transplantation (ASCT). (a) Representative fluorescence activated cell sorter (FACS) plots demonstrate how NKT cell subsets are analysed. (b) CD4+, CD4−CD8−, CD4−CD8+, CD161+ and CD56+ NKT cell subsets were monitored during a longitudinal study of previously untreated multiple myeloma (MM) patients receiving lenalidomide, ASCT and dendritic cell (DC) vaccination in MM. (c) Cell numbers for CD4+, CD4−CD8−, CD4−CD8+, CD161+ and CD56+ NKT cell subsets are shown for individual patients.
Longitudinal analysis of NKT cell function
A previous study reported that NKT cells from patients with progressive MM lacked the ability to produce IFN-γ after stimulation 8. The regulatory functions of NKT cells rely upon rapid release of cytokines in response to stimulation; lenalidomide has been proposed as a means of boosting NKT cell function in patients with MM because several studies have demonstrated improved cytokine production by NKT cells in the presence of lenalidomide 18,20.
We have reported previously that lenalidomide treatment of patients with MDS did not impact NKT cell frequency or function significantly 29. To establish a more complete understanding of the nature and kinetics of the NKT cell cytokine defect in patients with MM, and the impact of lenalidomide treatment on this important function, we assessed NKT cell cytokine responses from four MM patients throughout a 6–8-month course of lenalidomide and dexamethasone induction therapy, ASCT and lenalidomide and DC vaccination maintenance therapy (Fig. 5a,b). We stimulated PBMCs in vitro with PMA and ionomycin and assayed the T cells and NKT cells from these cultures for intracellular IFN-γ, TNF and IL-4 using flow cytometry (Fig. 5a). NKT cell numbers of two patients did not recover sufficiently for cytokine analysis post-ASCT, but in the others assessed there was NKT cell production of IFN-γ, TNF and IL-4 for 6 months after ASCT (Fig. 5b). The small blood volumes available from these patients, coupled with the low frequency of NKT cells, meant that we were unable to conduct functional assays for all patients. Despite this, our data show that NKT cells from patients with MM remained capable of significant cytokine production of IFN-γ, IL-4 and TNF at each time-point prior to and after treatment.
Figure 5.
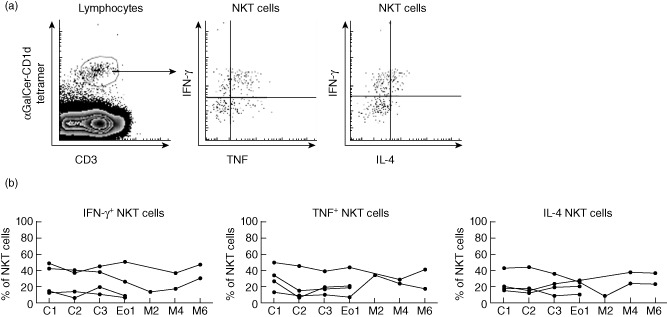
Natural killer (NK) T cells retain functional capacity after autologous stem cell transplantation (ASCT). Peripheral blood mononuclear cells (PBMCs) were stimulated ex vivo with phorbol myristate acetate (PMA)/ionomycin for 4 h. (a) Representative fluorescence activated cell sorter (FACS) plots from previously untreated patients are shown. (b) The percentage of NKT cells capable of interferon (IFN)-γ, tumour necrosis factor (TNF) and interleukin (IL)-4 production was assessed for four patients with sufficient NKT cells for cytokine analysis.
Discussion
We report on a longitudinal study of NKT cells in patients with MM undergoing treatment with lenalidomide. Earlier reports had indicated that NKT cell defects may be important in the development of MM 8–18 and that NKT cells could be a new target for immune-based treatment of MM patients 20–23. To investigate the therapeutic significance of NKT cells in MM, we analysed NKT cell frequency and function in longitudinal studies of newly diagnosed patients with MM throughout several stages of therapy, with an emphasis on identifying and defining any NKT cell defects and determining the impact of lenalidomide treatment on NKT cell frequency and function. The latter objective followed reports that lenalidomide, which is already an approved treatment of MM, could act to increase NKT cell numbers and cytokine expression 18–20.
Our analysis of relapsed patients with MM identified a significant NKT cell deficiency, but the frequency and cytokine production of NKT cells was normal in newly diagnosed patients. Longitudinal analysis of newly diagnosed MM patients demonstrated that a profound and extended deficiency of NKT cells developed after high-dose chemotherapy with ASCT. We observed that NKT cells were capable of producing IFN-γ, TNF and IL-4 at all time-points in our study, including prior to treatment, which contrasts with earlier reports of severely impaired IFN-γ production by NKT cells in patients with MM.
Our data suggest that NKT cell defects are not present at the time of MM diagnosis and therefore would appear not to predispose to the onset of MM. Rather, NKT cells are markedly depleted by ASCT and/or associated prior chemotherapy and recover far more slowly than conventional T cells or NK cells. The impact of NKT cell deficiency in patients with MM after intensive prior therapy (regardless of cause) is not known, but is worthy of further investigation given the significance of NKT cell deficiency in other patient groups. For example, we do not exclude the possibility that such a defect could still potentially impact upon the duration of remission from MM and/or their response to subsequent treatments that affect, or rely upon, NKT cells.
Our longitudinal analysis of individuals also allowed us to investigate the effect of lenalidomide on NKT cells. This followed several in-vitro studies (including our own 29) 19–31 and one in-vivo study 18, which showed that lenalidomide could promote NKT cell proliferation and cytokine production, suggesting that lenalidomide's mode of action in MM might involve direct effects on NKT cells 12,20. However, NKT cell frequency and function were normal in patients with MM prior to treatment, and subsequent induction therapy with lenalidomide had no significant impact on the number or cytokine responsiveness of NKT cells. The results suggest that the clinical benefit of lenalidomide treatment of patients with MM does not rely upon counteracting a pre-existing NKT cell defect, or upon stimulating residual NKT cells.
Our finding that lenalidomide had no measurable effect on NKT cells in patients with MM, even after an extended treatment regimen, is consistent with our recent analysis of NKT cells from patients with myelodsyplastic syndromes (MDS) treated with lenalidomide 29. We cannot exclude formally that lenalidomide promoted a moderate NKT cell expansion masked by similar NKT cell losses through death or selective egress from the blood, however, we contend that this is extremely unlikely because our longitudinal analysis showed no major changes in NKT cell frequency or function at any stage after lenalidomide treatment. The more likely explanation is that NKT cells were not affected meaningfully by lenalidomide in vivo and are unlikely to have contributed significantly to clinical improvements.
The similar frequencies and cytokine production of NKT cells from MM patients and healthy donors means that we should not discount the possibility that NKT cells could be targeted for therapeutic advantage in patients with MM. A recent clinical trial reported that a combination therapy approach of α-GalCer-loaded monocyte-derived DCs and low-dose lenalidomide to treat patients with MM led to NKT cell activation and tumour regression, with the authors concluding that direct NKT cell activation (and the synergistic activation of innate cells), perhaps in combination with the co-stimulatory activities of lenalidomide, may benefit anti-cancer treatment in humans 20. The stimulation of NKT cells in that study was attributed partly to the effects of lenalidomide on NKT cells, however, we question whether it is possible to fully delineate the respective roles of NKT cells versus lenalidomide because lenalidomide and the α-GalCer-loaded DCs were administered together, and because NKT cells, NK cells and other innate cells activated in the study can stimulate one another reciprocally. Furthermore, although the potential impact of dexamethasone should be considered [38], the clinical benefits of immunotherapies that act via NKT cell stimulation are likely to be maximized in the setting of normal NKT cell populations. This would require that therapies should be employed early in the treatment schedule when NKT cells are preserved, and not be delayed to a time where NKT cells have been functionally depleted, unless additional treatments are used to restore NKT cell numbers, such as the administration of glycolipid-based vaccines 39.
An issue worthy of further investigation is the distribution of NKT cell subsets in patients with MM. Prior to treatment, the average CD4− to CD4+ ratio of NKT cell subsets was more than twofold higher among patients with MM, however, there was high variability of this parameter and the difference between groups was not significant. The question of whether the different means is indicative of a true defect in patients with MM is important to resolve, because CD4− NKT cells have potent anti-tumour activities and the CD4− subset deficiency implied by our study could conceivably contribute to tumour predisposition, even when overall NKT cell numbers are normal.
In summary, we report that patients with newly diagnosed MM were not significantly defective in NKT cell frequency or cytokine production, however, NKT cells were specifically depleted after ASCT and remained so even after lenalidomide treatment. This indicated that NKT cell defects do not predispose to MM and that the positive clinical response to lenalidomide in patients with MM does not rely upon stimulation of the NKT cell pool.
Acknowledgments
The authors gratefully acknowledge the participation of patients and blood donors in the study, and the contributions of Rhonda Holdsworth (Australian Red Cross Blood Bank) and Kon Kyparissoudis (research assistant). The authors acknowledge the generous funding of this study by the National Health and Medical Research Council (NHMRC), Project grant (no. 454363), the Fiona Elsey Cancer Research Institute (FECRI) and the Victorian Cancer Agency. The authors also acknowledge the following funding support for this research: S. P. B. is a recipient of an NHMRC Career Development Award and current Research Fellowships from University of Ballarat (Robert H. T. Smith Fellowship) and FECRI (Dorevitch Research Fellowship); D. I. G. is supported by an NHMRC Program grant (no. 251608, renewed as no. 454569) and an NHMRC Senior Principal Research Fellowship.
Disclosure
The authors declare no conflicts of interest.
Supporting Information
Additional Supporting information may be found in the online version of this article at the publisher's website
Multiple myeloma clinical trial design. Previously untreated newly diagnosed patients were enrolled onto the Litvacc study (a), where following lenalidomide + dexamethasone (LEN + DEX) induction the patients received an autologous stem cell transplantation (ASCT) followed by myeloma lysate-pulsed dendritic cells (DCs) + LEN maintenance or len maintenance alone (in our study, 10 of 11 patients received myeloma lysate-pulsed DCs + LEN maintenance). Patients with refractory relapsed disease were enrolled into the RevLite study (b) to receive LEN + DEX induction, then repeated cycles of LEN + DEX until disease progression. A comparison between the two trials is provided in table (c) including disease state, drug dose and cycles of therapy.
References
- 1.Godfrey DI, MacDonald HR, Kronenberg M, Smyth MJ, Van Kaer L. NKT cells: what's in a name? Nat Rev Immunol. 2004;4:231–237. doi: 10.1038/nri1309. [DOI] [PubMed] [Google Scholar]
- 2.Bendelac A, Savage PB, Teyton L. The biology of NKT cells. Annu Rev Immunol. 2007;25:297–336. doi: 10.1146/annurev.immunol.25.022106.141711. [DOI] [PubMed] [Google Scholar]
- 3.Godfrey DI, Kronenberg M. Going both ways: immune regulation via CD1d-dependent NKT cells. J Clin Invest. 2004;114:1379–1388. doi: 10.1172/JCI23594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Berzins SP, Smyth MJ, Baxter AG. Presumed guilty: natural killer T cell defects and human disease. Nat Rev Immunol. 2011;11:131–142. doi: 10.1038/nri2904. [DOI] [PubMed] [Google Scholar]
- 5.Wu L, Van Kaer L. Natural killer T cells and autoimmune disease. Curr Mol Med. 2009;9:4–14. doi: 10.2174/156652409787314534. [DOI] [PubMed] [Google Scholar]
- 6.van der Vliet HJJ, von Blomberg BME, Nishi N, et al. Circulating V alpha 24(+) V beta 11(+) NKT cell numbers are decreased in a wide variety of diseases that are characterized by autoreactive tissue damage. Clin Immunol. 2001;100:144–148. doi: 10.1006/clim.2001.5060. [DOI] [PubMed] [Google Scholar]
- 7.Iwamura C, Nakayama T. Role of NKT cells in allergic asthma. Curr Opin Immunol. 2010;22:807–813. doi: 10.1016/j.coi.2010.10.008. [DOI] [PubMed] [Google Scholar]
- 8.Dhodapkar MV, Geller MD, Chang DH, et al. A reversible defect in natural killer T cell function characterizes the progression of premalignant to malignant multiple myeloma. J Exp Med. 2003;197:1667–1676. doi: 10.1084/jem.20021650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Swann JB, Coquet JM, Smyth MJ, Godfrey DI. CD1-restricted T cells and tumor immunity. Curr Top Microbiol Immunol. 2007;314:293–323. doi: 10.1007/978-3-540-69511-0_12. [DOI] [PubMed] [Google Scholar]
- 10.Terabe M, Berzofsky JA. The role of NKT cells in tumor immunity. Adv Cancer Res. 2008;101:277–348. doi: 10.1016/S0065-230X(08)00408-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Balato A, Unutmaz D, Gaspari AA. Natural killer T cells: an unconventional T-cell subset with diverse effector and regulatory functions. J Invest Dermatol. 2009;129:1628–1642. doi: 10.1038/jid.2009.30. [DOI] [PubMed] [Google Scholar]
- 12.Neparidze N, Dhodapkar MV. Harnessing CD1d-restricted T cells toward antitumor immunity in humans. Ann NY Acad Sci. 2009;1174:61–67. doi: 10.1111/j.1749-6632.2009.04931.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dhodapkar MV, Richter J. Harnessing natural killer T (NKT) cells in human myeloma: progress and challenges. Clin Immunol. 2011;140:160–166. doi: 10.1016/j.clim.2010.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Vivier E, Ugolini S, Blaise D, Chabannon C, Brossay L. Targeting natural killer cells and natural killer T cells in cancer. Nat Rev Immunol. 2012;12:239–252. doi: 10.1038/nri3174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rajkumar SV, Buadi F. Multiple myeloma: new staging systems for diagnosis, prognosis and response evaluation. Best Pract Res Clin Haematol. 2007;20:665–680. doi: 10.1016/j.beha.2007.10.002. [DOI] [PubMed] [Google Scholar]
- 16.Ghalamfarsa G, Hadinia A, Yousefi M, Jadidi-Niaragh F. The role of natural killer T cells in B cell malignancies. Tumour Biol. 2013;34:1349–1360. doi: 10.1007/s13277-013-0743-x. [DOI] [PubMed] [Google Scholar]
- 17.Nur H, Fostier K, Aspeslagh S, et al. Preclinical evaluation of invariant natural killer T cells in the 5T33 multiple myeloma model. PLOS One. 2013;8:e65075. doi: 10.1371/journal.pone.0065075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chang DH, Liu N, Klimek V, et al. Enhancement of ligand-dependent activation of human natural killer T cells by lenalidomide: therapeutic implications. Blood. 2006;108:618–621. doi: 10.1182/blood-2005-10-4184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Song W, van der Vliet HJ, Tai YT, et al. Generation of antitumor invariant natural killer T cell lines in multiple myeloma and promotion of their functions via lenalidomide: a strategy for immunotherapy. Clin Cancer Res. 2008;14:6955–6962. doi: 10.1158/1078-0432.CCR-07-5290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Richter J, Neparidze N, Zhang L, et al. Clinical regressions and broad immune activation following combination therapy targeting human NKT cells in myeloma. Blood. 2013;121:423–430. doi: 10.1182/blood-2012-06-435503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Spanoudakis E, Hu M, Naresh K, et al. Regulation of multiple myeloma survival and progression by CD1d. Blood. 2009;113:2498–2507. doi: 10.1182/blood-2008-06-161281. [DOI] [PubMed] [Google Scholar]
- 22.Chang DH, Deng H, Matthews P, et al. Inflammation-associated lysophospholipids as ligands for CD1d-restricted T cells in human cancer. Blood. 2008;112:1308–1316. doi: 10.1182/blood-2008-04-149831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Benson DM., Jr Can NKT cells extinguish smoldering myeloma? Blood. 2013;121:418–420. doi: 10.1182/blood-2012-11-465849. [DOI] [PubMed] [Google Scholar]
- 24.Gorgun G, Calabrese E, Soydan E, et al. Immunomodulatory effects of lenalidomide and pomalidomide on interaction of tumor and bone marrow accessory cells in multiple myeloma. Blood. 2010;116:3227–3237. doi: 10.1182/blood-2010-04-279893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Shah SR, Tran TM. Lenalidomide in myelodysplastic syndrome and multiple myeloma. Drugs. 2007;67:1869–1881. doi: 10.2165/00003495-200767130-00005. [DOI] [PubMed] [Google Scholar]
- 26.Mariz JM, Esteves GV. Review of therapy for relapsed/refractory multiple myeloma: focus on lenalidomide. Curr Opin Oncol. 2012;24(Suppl. 2):S3–11. doi: 10.1097/01.cco.0000410243.84074.dc. [DOI] [PubMed] [Google Scholar]
- 27.Davies FE, Raje N, Hideshima T, et al. Thalidomide and immunomodulatory derivatives augment natural killer cell cytotoxicity in multiple myeloma. Blood. 2001;98:210–216. doi: 10.1182/blood.v98.1.210. [DOI] [PubMed] [Google Scholar]
- 28.Quach H, Ritchie D, Stewart AK, et al. Mechanism of action of immunomodulatory drugs (IMiDS) in multiple myeloma. Leukemia. 2010;24:22–32. doi: 10.1038/leu.2009.236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chan AC, Neeson P, Leeansyah E, et al. Testing the NKT cell hypothesis in lenalidomide-treated myelodysplastic syndrome patients. Leukemia. 2010;24:592–600. doi: 10.1038/leu.2009.279. [DOI] [PubMed] [Google Scholar]
- 30.De Sanctis JB, Mijares M, Suarez A, et al. Pharmacological properties of thalidomide and its analogues. Recent Pat Inflamm Allergy Drug Discov. 2010;4:144–148. doi: 10.2174/187221310791163026. [DOI] [PubMed] [Google Scholar]
- 31.Zhu D, Corral LG, Fleming YW, Stein B. Immunomodulatory drugs Revlimid (lenalidomide) and CC-4047 induce apoptosis of both hematological and solid tumor cells through NK cell activation. Cancer Immunol Immunother. 2008;57:1849–1859. doi: 10.1007/s00262-008-0512-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Chang DH, Osman K, Connolly J, et al. Sustained expansion of NKT cells and antigen-specific T cells after injection of alpha-galactosyl-ceramide loaded mature dendritic cells in cancer patients. J Exp Med. 2005;201:1503–1517. doi: 10.1084/jem.20042592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lee PT, Benlagha K, Teyton L, Bendelac A. Distinct functional lineages of human V(alpha)24 natural killer T cells. J Exp Med. 2002;195:637–641. doi: 10.1084/jem.20011908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Gumperz JE, Miyake S, Yamamura T, Brenner MB. Functionally distinct subsets of CD1d-restricted natural killer T cells revealed by CD1d tetramer staining. J Exp Med. 2002;195:625–636. doi: 10.1084/jem.20011786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Terabe M, Berzofsky JA. NKT cells in immunoregulation of tumor immunity: a new immunoregulatory axis. Trends Immunol. 2007;28:491–496. doi: 10.1016/j.it.2007.05.008. [DOI] [PubMed] [Google Scholar]
- 36.Crowe NY, Coquet JM, Berzins SP, et al. Differential antitumor immunity mediated by NKT cell subsets in vivo. J Exp Med. 2005;202:1279–1288. doi: 10.1084/jem.20050953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Chan AC, Leeansyah E, Cochrane A, et al. Ex-vivo analysis of human natural killer T cells demonstrates heterogeneity between tissues and within established CD4(+) and CD4(–) subsets. Clin Exp Immunol. 2013;172:129–137. doi: 10.1111/cei.12045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hsu AK, Quach H, Tai T, et al. The immunostimulatory effect of lenalidomide on NK-cell function is profoundly inhibited by concurrent dexamethasone therapy. Blood. 2011;117:1605–1613. doi: 10.1182/blood-2010-04-278432. [DOI] [PubMed] [Google Scholar]
- 39.Cerundolo V, Silk JD, Masri SH, Salio M. Harnessing invariant NKT cells in vaccination strategies. Nat Rev Immunol. 2009;9:28–38. doi: 10.1038/nri2451. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Multiple myeloma clinical trial design. Previously untreated newly diagnosed patients were enrolled onto the Litvacc study (a), where following lenalidomide + dexamethasone (LEN + DEX) induction the patients received an autologous stem cell transplantation (ASCT) followed by myeloma lysate-pulsed dendritic cells (DCs) + LEN maintenance or len maintenance alone (in our study, 10 of 11 patients received myeloma lysate-pulsed DCs + LEN maintenance). Patients with refractory relapsed disease were enrolled into the RevLite study (b) to receive LEN + DEX induction, then repeated cycles of LEN + DEX until disease progression. A comparison between the two trials is provided in table (c) including disease state, drug dose and cycles of therapy.