

Abstract
The green micro-alga Chlamydomonas reinhardtii is an elegant model organism to study all aspects of oxygenic photosynthesis. Chlorophyll (Chl) and heme are major tetrapyrroles that play an essential role in energy metabolism in photosynthetic organisms and are synthesized via a common branched tetrapyrrole biosynthetic pathway. One of the enzymes in the pathway is Mg chelatase (MgChel) which inserts Mg 2+ into protoporphyrin IX (PPIX, proto) to form magnesium-protoporphyrin IX (MgPPIX, Mgproto), the first biosynthetic intermediate in the Chl branch. MgChel is a multimeric enzyme that consists of three subunits designated CHLD, CHLI and CHLH. Plants have two isozymes of CHLI (CHLI1 and CHLI2) which are 70%-81% identical in protein sequences. Although the functional role of CHLI1 is well characterized, that of CHLI2 is not. We have isolated a non-photosynthetic light sensitive mutant 5A7 by random DNA insertional mutagenesis that is devoid of any detectable Chl. PCR based analyses show that 5A7 is missing the CHLI1 gene and at least eight additional functionally uncharacterized genes. 5A7 has an intact CHLI2 gene. Complementation with a functional copy of the CHLI1 gene restored Chl biosynthesis, photo-autotrophic growth and light tolerance in 5A7. We have identified the first chli1 mutant of Chlamydomonas reinhardtii and in green algae. Our results show that in the wild type Chlamydomonas CHLI2 protein amount is lower than that of CHLI1 and the chli1 mutant has a drastic reduction in CHLI2 protein levels although it possesses the CHLI2 gene. Our chli1 mutant opens up new avenues to explore the functional roles of CHLI1 and CHLI2 in Chl biosynthesis and chloroplast to nucleus retrograde signaling in Chlamydomonas, which has never been studied before.
Introduction
The green micro-alga Chlamydomonas reinhardtii possesses a photosynthetic apparatus very similar to that of higher plants, can grow photo-autotrophically and heterotrophically (it can metabolize exogenous acetate as a carbon source) and possesses a completely sequenced genome 1. These attributes make it an elegant model organism to study oxygenic photosynthesis and chloroplast biogenesis 2, 3. In photosynthetic organisms, tetrapyrroles like Chl and heme are essential for energy metabolism (i.e. photosynthesis and respiration). Biosynthesis of Chl and heme occur via a common branched pathway that involves both nuclear- and chloroplast-encoded enzymes in most photosynthetic organisms 4. In photosynthetic eukaryotes, 5-aminolevulinic acid (ALA) is synthesized from glutamine through glutamyl-tRNA 5. Conversion of ALA through several steps yields protoporphyrin IX (PPIX), the last common precursor for both heme and Chl biosynthesis 5. Ferrochelatase inserts iron in the center of PPIX thus committing it to the heme branch of the pathway. Insertion of Mg 2+ in PPIX by MgChel leads to Mgproto, the first biosynthetic intermediate in the Chl branch 6. Magnesium chelatase has three subunits, which are CHLD, CHLH and CHLI 7. The ATP-dependent catalytic mechanism of the heterotrimeric MgChel complex includes at least two steps 7– 9: an activation step, followed by the Mg 2+ insertion 8. Activation of MgChel with ATP involves CHLD and CHLI while CHLH is required for the chelation step 10. CHLI belongs to the AAA+ family of ATPases. Plants have two isozymes of CHLI1 (CHLI1 and CHLI2) which are 70%–81% identical in protein sequences 10. Although the functional role of CHLI1 is well characterized, that of CHLI2 is not. Most of the data on CHLI comes from studies on Arabidopsis thaliana chli mutants and the functional significance of CHLI1 and CHLI2 has not been studied in green algae 10– 16. In Arabidopsis CHLI2 plays a limited role in Chl biosynthesis because of its lower expression level compared to that of CHLI1 12– 15. In Arabidopsis the CHLI2 protein amount is lower than that of CHLI1. When overexpressed, CHLI2 can fully rescue an Arabidopsis chli1chli2 double mutant 12.
We have isolated the first chli1 mutant of Chlamydomonas reinhardtii ( 5A7) which possesses an intact CHLI2 gene. Transformation of 5A7 with a functional copy of the CHLI1 gene restored Chl biosynthesis. Western analyses show that the CHLI2 protein level is lower than that of CHLI1 in the wild type strain and CHLI2 protein is barely detectable in the mutant strain. In this study, we present our molecular data on the identification of the mutation locus in 5A7 and its complementation.
Materials and methods
Algal media and cultures
Chlamydomonas strains 4A+ (a gift from Dr. Krishna Niyogi (UC, Berkeley), 5A7/chli1 (generated by our laboratory) and chli1 complements (generated by our laboratory) were grown either in Tris-Acetate Phosphate (TAP) heterotrophic media or in Sueoka’s High Salt (HS) photo-autotrophic media. TAP and HS liquid media and agar plates were prepared in the lab using reagents from Fisher Scientific (Pittsburg, PA) according to the protocol given in Gorman and Levine (1965) 17 and Sueoka (1960) 18, respectively. The 4A+ strain and chli1 complements were maintained on TAP agar plates and TAP+zeocin (Sigma, St. Louis, MO) plates, respectively under dim light intensities (10–15 µmol photons m -2s -1) at 25°C. The final zeocin concentration was 15 µg/ml). The chli1 mutant ( 5A7) was maintained in the dark on TAP 1.5% agar plates containing 10 µg/ml of paromomycin (Sigma, St. Louis, MO). Liquid algal cultures used for RNA and genomic DNA extractions and protein analyses were grown in 100 ml flasks on the New Brunswick Scientific Excella E5 platform shaker (Enfield, CT) at 150 rpm in the dark or in the dim light.
Generation of the 5A7 mutant
The purified pBC1 plasmid from the DH5α Escherichia coli clone harboring the pBC1 plasmid (obtained from Dr. Krishna Niyogi’s laboratory at UC, Berkeley) was used for random DNA insertional mutagenesis. This plasmid contains two antibiotic resistance genes: APHVIII and Amp R ( Figure 1). APHVIII confers resistance against the antibiotic paromomycin (Sigma, St. Louis MO) and was used as a selection marker for screening of Chlamydomonas transformants. Amp R was used as a selection marker for screening of E. coli clones harboring the pBC1 plasmid. E.coli was grown in 1 l of Luria Bertani (LB) broth containing 1% tryptone, 0.5% of yeast extract, 1% NaCl and ampicillin [final concentration of ampicillin:100 µg/ml]. LB reagent was prepared in the laboratory using reagents purchased from Fisher (Pittsburgh, PA). Ampicillin was purchased from Fisher (Pittsburgh, PA). The culture was incubated at 37°C overnight. Plasmid purification from E. coli cells was facilitated by Qiagen plasmid mega kit according to the protocol given in the technical manual (Qiagen, Valencia, CA). Once purified from E. coli, the circular pBC1 vector was linearized with the restriction enzyme Kpn1 (NEB, Beverly, MA) according to the protocol given in the technical manual. The linearized DNA was purified using a QIAEX II gel extraction kit (Qiagen, Valencia, CA) according to the protocol given in the technical manual. All agarose DNA gel electrophoresis was visualized by BioRad Molecular Imager Gel Doc XR+ (BioRad, Hercules, CA). Transformation of parental strain 4A+ by the linearized pBC1 vector was performed utilizing the glass bead transformation technique described by Kindle et al. (1989) 19 and Dent et al. (2005) 2. Transformants were plated onto fresh TAP agar plates containing 10 µg/ml paromomycin (TAP+P) in the dark. Single colonies of mutants were picked and transferred onto fresh TAP+P plates using a numbered grid layout. Screening of photosynthetic and pigment deficient mutants was done by visual inspection and monitoring of growth under different light intensities in heterotrophic, mixotrophic and photo-autotrophic conditions 2.
Figure 1. Linearized pBC1 plasmid used for random insertional mutagenesis.

The cleavage site of the Kpn1 restriction enzyme, used for linearization of the vector is shown. APHVIII is under the control of combo promoters consisting of the promoter of the gene encoding the small subunit of Rubisco (RbcS2) and the promoter of the gene encoding the heat shock protein 70A (Hsp70A). pBC1 is a phagemid and its F1 origin (F1 ori) and pUC origin (pUC ori) are shown. The size of the plasmid is 4763 bp.
Genomic DNA and RNA extraction
4A+, chli1 complements and 5A7/chli1 were grown in TAP liquid media in the dark to a cell density of about 5 × 10 6 cells/ml of the culture. Genomic DNA was purified using a phenol-chloroform extraction method 19. RNA extraction was facilitated by TRIzol reagent from Invitrogen (Carlsbad, CA) following the protocol in the technical manual. DNA and RNA concentrations were measured using a Nanodrop 1000 spectrophotometer from Thermo Fisher Scientific (Wilmington, DE). DNase treatment was performed using Ambion’s TURBO DNA-free kit from Invitrogen (Carlsbad, CA) following the protocol in the technical manual to remove genomic DNA from the RNA preparation. Generation of cDNA was performed using Life Technologies Superscript III First-Strand Synthesis System from Invitrogen (Carlsbad, CA) following the protocol in the technical manual.
Thermal asymmetric interLaced PCR
TAIL (Thermal Asymmetric InterLaced) PCR was implemented, following the protocol of Dent et al. (2005) 2. This protocol was implemented with one modification of utilizing a non-degenerate primer (AD2) derived from the original degenerate primer (AD) for TAIL PCR as this non-degenerate primer was giving us optimum yield without generating excess nonspecific products. HotStar Taq Plus DNA polymerase kit reagents (Qiagen, Valencia, CA) were used for PCR. The PCR reaction mixture consisted of 1× PCR buffer, 200 µM of each dNTP, 1× Q-solution, 2.5 units of HotStar Taq Plus DNA polymerase, 60 pmoles of the non-degenerate primer AD2 and 5 pmol of the APHVIII specific primer. Primers were ordered from IDT (Skokie, IL; Table 1). The non-degenerate primer AD2 has a T m of 46°C while the three APHVIII specific primers used had T m ranging from 58°C to 64°C. PCR cycling programs were created using the program given in Dent et al. (2005) 2. TAIL1 PCR product was diluted 10 and 25-fold and 2 μl of the diluted TAIL1 PCR product was used for TAIL2 PCR reactions. The TAIL2 PCR product was gel purified using QIAEX II gel extraction kit (Qiagen, Valencia, CA) according to the protocol given in the technical manual. The purified TAIL2 PCR product was sequenced at the UC, Berkeley DNA Sequencing Facility (Berkeley, CA).
Table 1. List of primers used for Thermal Asymmetric InterLaced (TAIL) PCR, verification of TAIL-PCR product and DNA sequencing.
| Primer name | Sequence of primer | Location |
|---|---|---|
| AD | 5´-NTC GWG WTS CNA GC-3´ | Random degenerate primer |
| AD2 | 5´-ATCGTGTTCCCAGC-3´ | Non-degenerate primer derived from the primer AD |
| 1F | 5´-AAA GAC TGA TCA GCA CGA AAC GGG-3´ | APHVIII 3´UTR |
| 2F | 5´-TAA GCT ACC GCT TCA GCA CTT GAG-3´ | APHVIII 3´UTR |
| 2R | 5´-CTC AAG TGC TGA AGC GGT AGC TTA-3´ | APHVIII 3´UTR |
| 3R | 5´-TCT TCT GAG GGA CCT GAT GGT GTT-3´ | APHVIII 3´UTR |
| 4R | 5´-GGG CGG TAT CGG AGG AAA AGC TG-3´ | APHVIII 3´UTR |
Figure 3. Locating the APHVIII flanking genomic sequence in 5A7.

( A) A diagram showing a truncated pBC1 illustrating the APHVIII end of the linearized pBC1 vector. Primers used for PCR are shown by numbered black arrows. Thermal Asymmetric InterLaced 1 (TAIL1) PCR was performed using primer 4R and AD2. ( B) TAIL2 PCR was performed using primer 3R and AD2. Lanes 1 and 4 are zero DNA lanes; in lane 2, a 10-fold diluted TAIL1 PCR product was used for TAIL2 PCR; in lane 5, a 25-fold diluted TAIL1 PCR product was used for TAIL2 PCR; lanes 3 and 6 are blank lanes. The 850 bp product used for DNA sequencing is highlighted. ( C) Gel purified DNA product (850 bp) from the TAIL2 PCR was used to verify if the product was specific to the APHVIII gene. F and R stand for forward and reverse primers, respectively. AD2 is a non-degenerate primer. PCR primer names are labeled on the top of the gel. In lanes A and B, where triple primers were used for PCR, PCR products are labeled by the corresponding primer combinations that gave rise to the specific product. PCR product sizes are shown beside the primer combinations. All primer sequences are shown in Table 1. ST stands for 1 kb plus ladder (Invitrogen, Carlsbad, CA). DNA samples were run on a 1% agarose gel.
Figure 4. The APHVIII flanking genomic DNA sequence in 5A7.

Primer 2R ( Table 1), specific to the 3´UTR of the APHVIII gene was used for sequencing the 850 bp Thermal Asymmetric InterLaced 2 (TAIL2) PCR product. 3´UTR sequence of APHVIII is in bold black, extra nucleotide additions are in bold blue. The flanking Chlamydomonas UP6 genomic sequence is denoted in red. The APHVIII end of the plasmid has been inserted after the eighth nucleotide in the fourth exon of UP6 gene.
Genomic and reverse transcription PCR
Primers were designed based on genomic DNA sequences available in the Chlamydomonas genome database in Phytozome. Amplifications of genomic DNA and cDNA were executed using MJ Research PTC-200 Peltier Thermal Cycler (Watertown, MA). HotStar Taq Plus DNA polymerase kit (Qiagen, Valencia, CA) was used for PCR following the cycling conditions given in the Qiagen protocol booklet. Annealing temperature was between 55 and 60°C depending on the T m of the primers. Extension time was varied according to the size of the PCR product amplified. Final extension was set at 72°C for ten minutes. All genomic and reverse transcription PCR products were amplified for a total of thirty-five cycles. 50–150 ng of genomic DNA or cDNA templates were used for PCR reactions. For semi-quantitative RT-PCR using CHLI1 and CHLI2 gene specific primers, 3 μg of total RNA was converted into cDNA and then 150 ng of cDNA templates were used for RT-PCR. Sequences of primers used for genomic and RT-PCR are shown in Table 2, Table 3 and Table 4.
Table 2. List of primers used for amplifying CHLI1 and four neighboring genes downstream of CHLI1.
These primers were used to generate the data in Figure 6A. The gene loci in Phytozome ( http://www.phytozome.net/) are: CHLI1 (Cre06.g306300), UP1 (Cre06.g306250), UP2 (Cre06.g306200), UP3 (Cre06.g306150) and UP4 (Cre06.g306100).
| Primer name | Sequence of primer | Gene |
|---|---|---|
| CHLI1AF | 5´-ACTACGACTTCCGCGTCAAGATCA-3´ | CHLI1 |
| CHLI1BR | 5´-CATGCCGAACACCTGCTTGAAGAT-3´ | CHLI1 |
| PB138 | 5´-ATGACCGTAACGCTGCGTAC-3´ | UP1 |
| PB139 | 5´-CGTGAACGACAGTGTTTAGCG-3´ | UP1 |
| PB132 | 5´-AAGGGCATCAGCTACAAGGTC-3´ | UP2 |
| PB133 | 5´-GGCATCGAGGATGTATTGGTTG-3´ | UP2 |
| UP3F | 5´-GGC ACA CAA GCG TGA TTT TCT GG-3´ | UP3 |
| UP3R | 5´-CGG CAC GTC GAA GAC AAA CT-3´ | UP3 |
| UP4F | 5´-CTT TGA CCT GCA AAG AGA GAA AGC G-3´ | UP4 |
| UP4R | 5´-CAC CAC CTT GAT GCC CTT GAG-3´ | UP4 |
Figure 6. PCR analyses using primers specific to eight genes neighboring the CHLI1 locus.

( A) PCR using the genomic DNA of 5A7 and 4A+ with primers specific to CHLI1 and four neighboring genes ( UP1, UP2, UP3, UP4) downstream of the CHLI1 gene. The sizes of the genomic DNA PCR products for CHLI1, UP1, UP2, UP3 and UP4 are, 459, 100, 342, 550 and 672 (bp), respectively. Odd numbered lanes denote 5A7; even numbered lanes denote 4A+; ST denotes 1 kb plus DNA ladder. ( B) PCR using the genomic DNA of 5A7 and 4A+ with primers specific to CHLI1 and four neighboring genes ( FDX3, AMT, UP5 and UP6) upstream of the CHLI1 gene. The sizes of the genomic DNA PCR products for CHLI1, FDX3, AMT, UP5 and UP6 are, 459, 90, 369, 379 and 369 (bp), respectively. Odd numbered lanes denote 5A7; even numbered lanes denote 4A+; M denotes 50 bp DNA ladder (NEB, Beverly, MA). ( C) PCR and RT-PCR with UP6 gene specific primers using the 5A7 and 4A+ genomic DNA and cDNA. Actin was used as a control. Actin genomic and cDNA product sizes are 527 and 305 (bp), respectively. Odd numbered lanes denote genomic DNA PCR products; even numbered lanes denote cDNA products. All primers used spanned an intron. M denotes 50 bp DNA ladder. All DNA samples were run on a 1.8% agarose gel. Gene names are given at the bottom of the gel. Primer sequences are shown in Table 2 and Table 3.
Table 3. List of primers used for amplifying CHLI1, four neighboring genes upstream of CHLI1 and the actin gene and transcript.
These primers were used to generate the data in Figure 6B and 6C. The gene loci in Phytozome ( http://www.phytozome.net/) are: CHLI1 (Cre06.g306300), FDX3 (Cre06.g306350), AMT (g7098), UP5 (Cre06.g306450) and UP6 (Cre06.g306500) and Actin (Cre13.g603700).
| Primer name | Sequence of primer | Gene |
|---|---|---|
| CHLI1AF | 5´-ACTACGACTTCCGCGTCAAGATCA-3´ | CHLI1 |
| CHLI1BR | 5´-CATGCCGAACACCTGCTTGAAGAT-3´ | CHLI1 |
| PB134 | 5´-CTGGAGCGCACCTTTATGAAG-3´ | AMT |
| PB135 | 5´-AGTGGAACAGGTTCTCGATGAC-3´ | AMT |
| PB132 | 5´-AAGGGCATCAGCTACAAGGTC-3´ | FDX3 |
| PB133 | 5´-GGCATCGAGGATGTATTGGTTG-3´ | FDX3 |
| UP5F | 5´-GGG CAA CTG GAG CTT TGG C-3´ | UP5 |
| UP5R | 5´-CGT CTA TGT GCG CCA CGT C-3´ | UP5 |
| UP6F | 5´-GCA ACT GGA GCT TCG GCG-3´ | UP6 |
| UP6R | 5´-CGT AGG CGC CAA ACA CCG-3´ | UP6 |
| F2 | 5´-ACGACACCACCTTCAACTCCATCA-3´ | Actin |
| R2 | 5´-TTAGAAGCACTTCCGGTGCACGAT-3´ | Actin |
Table 4. List of primers for amplifying CHLI transcripts and complement testing.
These primers were used in the experiments that generated the data in Figure 7 and Figure 10 and also used for CHLI1 cDNA amplification for cloning.
| Primer name | Sequence of primer | Gene/purpose |
|---|---|---|
| CHLI1CR | 5´-TTGACCCTTTGACACGAACCAACC-3´ | CHLI1 |
| CHLI1BR | 5´-CATGCCGAACACCTGCTTGAAGAT-3´ | CHLI1 |
| CHLI1AF | 5´-ACTACGACTTCCGCGTCAAGATCA-3´ | CHLI1 |
| CHLI2BF | 5´-TGACGCATTTGTGGACTCGTGCAA-3´ | CHLI2 |
| CHLI2CR | 5´-CACACTTACACGTTCACGCAGCAA-3´ | CHLI2 |
| CHLI1XF | 5´-GGAATTCCATATGGCCTGAACATGCGTGTTTC-3´ | CHLI1 cDNA amplification for cloning |
| CHLI1XR | 5´-CCGGAATTCTTACTCCATGCCGAACACCTGCTT-3´ | CHLI1 cDNA amplification for cloning |
| PsaDF1 | 5´-CCACTGCTACTCACAACAAGCCCA-3´ | Complementation testing |
Figure 7. PCR analyses to check for the presence of the CHLI1 and CHLI2.

( A) Semi-quantitative RT-PCR analyses of 5A7 and 4A+ using CHLI1 and CHLI2 primers. ( B) PCR analyses using 5A7 and 4A+ genomic DNA with CHLI2 gene specific primers. Odd numbered lanes denote 5A7; even numbered lanes denote 4A+. PCR product sizes (bp) are labeled. ST denotes 1 kb plus DNA ladder. All DNA samples were run on a 1.8% agarose gel. Gene names are given at the top of the gel. Primer sequences are shown in Table 4.
Figure 10. Molecular analysis of chli1 complements.

( A) A schematic of the native CHLI1 gene. The tan bars denote UnTranslated Regions (UTRs), the white arrows represent exons and the black lines denote introns. ( B) A schematic of the CHLI1-pDBle complementation vector containing the CHLI1 cDNA. PsaD promoter, 5´UTR, 3´UTR and CHLI1 specific primers are labeled. ( C) Genomic DNA PCR using a PsaD 5´UTR specific primer and a CHLI1 specific primer. Product size: 1272 bp. Lane 1: 5A7; Lane 2: 4A+; Lane 3: chli1-7; Lane 4: chli1-8. ST represents 1 kb plus DNA ladder. ( D) Genomic DNA PCR using CHLI1 specific primers. Genomic DNA product size: 459 bp; cDNA product size: 249 bp. Lanes 3 and 4 show smaller PCR products compared to that in lane 2 as cDNA was used for complementation. Lane 1: 5A7; Lane 2: 4A+; Lane 3: chli1-7; Lane 4: chli1-8. ST represents 1 kb plus DNA ladder. All primer sequences are shown in Table 4.
Cloning of the CHLI1 cDNA in the pDBle vector
The pDBle vector (obtained from Dr. Saul Purton, University College London, UK) was double-digested with restriction enzymes EcoR1 and Nde1 (NEB, Beverly, MA) according to the protocol given in the technical manual. The CHLI1 cDNA template was amplified using primers given in Table 4. Ligation of the double digested ( NdeI and EcoR1 digested) CHLI1 cDNA and the NdeI/ EcoRI double-digested pDBle vector was done using the T4 ligase and 1 mM ATP (NEB, Beverly, MA). Chemically competent (CaCl 2 treated) E. coli cells were used for transformation. After transformation, E. coli cells were plated on LB+ampicillin (100 µg/ml) plates and incubated at 37°C overnight. Single colonies were picked the next day and plasmids were isolated from these clones. Isolated plasmids were double-digested with EcoR1 and Nde1 to verify the cloning of the CHLI1 cDNA. The CHLI1-pDBle construct from the selected clone was sequenced by the UC, Berkeley DNA Sequencing Facility (Berkeley, CA). Chromas Lite ( http://technelysium.com.au/) and BLAST were used to analyze DNA sequences.
Generation and screening of chli1 complements
Complementation of the chli1 was performed utilizing the glass bead transformation technique described by Kindle et al. 1989 20. 2 µg of the linearized CHLI1-pDBle was used to complement chli1. Transformed cells were plated onto fresh TAP plates containing 15 µg/ml zeocin (Z) and placed in the dark at 25°C. Single colonies were picked and transferred onto fresh TAP+Z plates using a numbered grid template for screening of potential chli1 complements. Screening of chli1 complements was done by visual inspection of green coloration and monitoring growth of light adapted complement strain cells either on TAP in the dark or in the dim light or HS plates under medium light (300 μmol photons m -2s -1).
Cellular protein analysis
Chlamydomonas cells from different strains grown in TAP in the dark were harvested, washed twice with fresh medium and resuspended in TEN buffer (10 mM Tris-HCl, 10 mM EDTA and 150 mM NaCl; pH 8). Protein concentrations of samples were determined by the method of Lowry et al. (1951) 21 with bovine serum albumin as standard. Gel lanes were either loaded with an equal amount of Chl (4 μg Chl) or with 40 μg of protein. Resuspended cell suspension was mixed in a 1:1 ratio with the sample solubilization buffer SDS-urea buffer (150 mM Tris-HCl, pH 6.8; 7% w/v SDS; 10% w/v glycerol; 2 M urea, bromophenol blue and 10% β-mercaptoethanol) and were incubated at room temperature for about thirty minutes, with intermittent vortexing. The sample solubilization buffer was prepared according to the protocol of Smith et al. (1990) 22 using reagents from Fisher (Pittsburgh, PA). After incubation, the solubilized protein samples were vortexed and spun at a maximum speed of 20,000 g in a microcentrifuge for five minutes at 4°C. The soluble fraction was loaded on a "any kD™ Mini-PROTEAN ® TGX™ Precast Gel" (BioRad, Hercules, CA) and SDS-PAGE analysis was performed according to Laemmli (1970) 23 using a Page Ruler prestained or unstained protein ladder (Fermentas, Glen Burnie, Maryland) at a constant current of 80 V for 2 hours. Gels were stained with colloidal Coomassie Gel code blue stain reagent (Thermo Fisher Scientific, Rockford, IL) for protein visualization.
Western analysis
Electrophoretic transfer of the SDS-PAGE resolved proteins onto an Immobilon P–PVDF membrane (Millipore, Billerica, MA) was carried out for 2 hours at a constant current of 400 mA in the transfer buffer (25 mM Tris, 192 mM glycine and 20% methanol). The CHLI1 polyclonal antibody was raised in rabbit against the full length Arabidopsis thaliana CHLI1 mature protein that lacks the predicted transit peptide 24. This antibody is a gift from Dr. Robert Larkin (Michigan State University). CHLI1 primary antibodies were diluted to a ratio of 1:2,000 before being used as a primary probe. The secondary antibodies used for Western blotting were conjugated to horseradish peroxidase (Pierce protein research product, Thermo Fisher Scientific, Rockford, IL) and diluted to a ratio of 1:20,000 with the antibody buffer. Western blots were developed by using the Supersignal West Pico chemiluminescent substrate kit (Pierce protein research product, Thermo Fisher Scientific, Rockford, IL).
Cell counts and chlorophyll extraction
Cell density (number of cells per ml of the culture) was calculated by counting the cells using a Neubauer ultraplane hemacytometer (Hausser Scientific, Horsham, PA). Pigments from intact cells were extracted in 80% acetone and cell debris was removed by centrifugation at 10,000 g for 5 minutes. The absorbance of the supernatant was measured with a Beckman Coulter DU 730 Life science UV/Vis spectrophotometer (Brea, CA). Chl a and b concentrations were determined by Arnon (1949) 25 equations, with corrections as described by Melis et al. (1987) 26.
Results
Generation and identification of the mutant 5A7
Mutant 5A7 was generated by random insertional mutagenesis of the Chlamydomonas reinhardtii wild type strain 4A+ (137c genetic background). 5A7 lacks detectable chlorophyll, appears yellowish-brown in color and grows only under heterotrophic conditions in the dark or in the dim light in the presence of acetate in the growth media ( Figure 1). It is incapable of photosynthesis and is sensitive to light intensities higher than 20 μmol photons m -2s -1 ( Figure 2).
Figure 2. Growth phenotype of 5A7.

( A) This figure shows the phenotypic difference of 5A7 compared to the parental strain, 4A+ on heterotrophic/mixotrophic agar media (TAP) plates under five different light conditions: dark + paromomycin (P), dark, very dim light (VDL, 2–4 μmol photons m -2s -1), dim light A (DLA, 10–15 μmol photons m -2s -1) and dim light B (DLB, 20–25 μmol photons m -2s -1). ( B) This figure shows the growth phenotype of 5A7 in liquid photo-autotrophic media (HS) under dim light (DL = 10–15 μmol photons m -2s -1).
Molecular characterization of the mutation in 5A7
The linearized plasmid pBC1 was used to generate 5A7 ( Figure 1). To find the insertion of the APHVIII end of the plasmid in 5A7, a modified TAIL (Thermal Asymmetric InterLaced) PCR method was used. Figure 3A shows the position of the vector specific TAIL PCR primers and also shows the arbitrary position of the random non-degenerate primer. A 850 bp DNA product from TAIL2 PCR was purified from the agarose gel ( Figure 3B, Table 1). This purified DNA product was used for PCR using internal primers specific to the 3´UTR (UnTranslated Region) of the APHVIII gene. The PCR results confirmed that the 850 bp DNA product contains the 3´UTR of the APHVIII gene ( Figure 3C). Sequencing of the 850 bp TAIL2 PCR product revealed that the APHVIII end of the plasmid has been inserted in the fourth exon of a hypothetical gene which we have named as UP6 ( Figure 4). UP6 (Cre06.g306500) is located on chromosome 6.
Figure 5 shows a schematic map of the UP6 locus with its eight neighboring genes UP4 (Cre06.g306100), UP3 (Cre06.g306150), UP1 (Cre06.g306250), UP2 (Cre06.g306200) CHLI1 (Cre06.g306300), FDX3 (Cre06.g306350), AMT (g7098) and UP5 (Cre06.g306450). It is to be noted that we have named all of these genes arbitrarily for our study except for the CHLI1 and FDX3 genes, which were annotated in the Chlamydomonas genome database. Readers are requested to identify these unknown genes by the gene locus number (Cre or g number) in the Phytozome database. PCR analyses with the genomic DNA of 4A+ and 5A7 were performed using primers specific to four neighboring genes upstream of the CHLI1 (including UP6) and four neighboring genes downstream of the CHLI1 locus ( Table 2 and Table 3; Figure 5A and 5B). PCR analyses revealed that all eight genes neighboring the CHLI1 locus were deleted or displaced from their native location ( Figure 5A and 5B). UP5 primers gave nonspecific multiple products in 5A7 ( Figure 5B). The first two exons of UP6 are present in the 5A7 genome as the UP6 primers spanning the first and the second exon, gave similar genomic DNA PCR product of the expected size as in the 4A+ lane ( Figure 5B). Reverse transcription (RT-PCR) analyses using the same UP6 primers on 5A7 and 4A+ cDNA did not yield a PCR product in 5A7 unlike that in 4A+ ( Figure 5C; Table 3). This shows that the insertion of the plasmid in the fourth exon of UP6 in 5A7 has hampered the transcription of the UP6 gene.
Figure 5. A schematic map of the UP6 locus on chromosome 6.
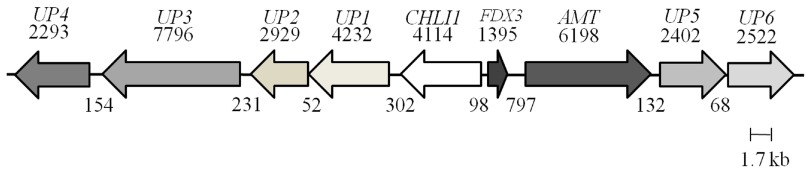
The map shows a 35.7 kb genomic DNA region that harbors the UP6 and eight genes located upstream of it. Each arrow represents a gene. The name of the gene is given on top of the arrow. The black numbers on the top of arrows denote sizes of genes (bp) while black numbers below denote distances in between genes (bp).
Taken together the data shows that at least a 35,715 bp genomic region has been deleted and or/displaced when the plasmid got inserted in the 5A7 genome. Except for the CHLI1 gene, the functions of the remaining eight genes (including UP6) are not known. We do not yet know the exact location of the pUC origin (pUC ori) end of the plasmid ( Figure 1) in the 5A7 genome.
Checking for the absence/presence of the CHLI1 transcript and the CHLI2 gene and transcript
As CHLI plays a role in Chl biosynthesis, we checked for the presence/absence of the CHLI1 and CHLI2 in 5A7. RT-PCR results show that CHLI1 transcript is absent and CHLI2 transcript is present in 5A7 ( Figure 7A, Table 4). Figure 7B shows the presence of the CHLI2 gene in 5A7.
Complementation of 5A7
We will be referring to strain 5A7 as chli1 from here onward. As our chli1 lacks Chl and CHLI1 is involved in Chl biosynthesis, we cloned the CHLI1 cDNA in the pDBle vector to transform 5A7 ( Figure 8, Table 4). CHLI1 expression is driven by the constitutive PsaD promoter in the CHLI1-pDBle construct ( Figure 8). pDBle has two Ble genes that confer resistance to the antibiotic zeocin. Figure 9 shows growth phenotypes of two chli1 complements ( chli1-7 and chli1-8); 5A7 and 4A+. chli1 complements are able to synthesize Chl, are not light sensitive and are capable of photosynthesis ( Figure 9). As the chli1 complements harbor the Ble gene (from the pDBle vector) and APHVIII gene (derived from the parental strain 5A7), they can grow both on zeocin and paromomycin media plates unlike chli1 and 4A+ ( Figure 9).
Figure 8. A schematic figure of the pDBle vector used for complementation of chli1.
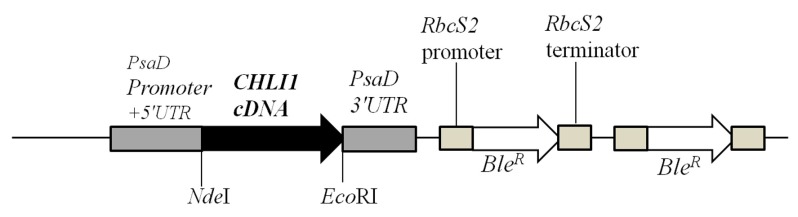
NdeI/ EcoR1 double digested CHLI1 cDNA (1260 bp) was cloned into the NdeI/ EcoRI double digested pDBle plasmid. Primers used for amplification of CHLI1 cDNA are shown in Table 4. CHLI1 expression is driven by the constitutive PsaD promoter. NdeI and EcoRI restriction sites are labeled. pDBle contains two copies of Ble R genes driven by the Rubisco ( RbcS2) promoter. The size of the CHLI1- pDBle construct is 7957 bp. Black arrow and white arrow denotes CHLI1 cDNA and Ble R gene, respectively. Grey boxes denote UnTranslated regions (UTR).
Figure 9. Growth phenotype analysis of chli1 complements.

chli1-7 and chli1-8 complements were grown with 5A7 and 4A+ under five growth conditions: TAP+Z (zeocin) in the dark, TAP in dim light (DL) (15 µmol photons m -2s -1), TAP+P (paromomycin) in DL, TAP in medium light ML (300 µmol photons m -2s -1) and HS in ML.
Chl analyses show that both chli1 complements are about 33–46% Chl deficient. chli1complements have a similar Chl a/ b ratio as that of the wild type ( Table 5, Data File below). Figure 10A and 10B show a schematic figure of the native Chlamydomonas CHLI1 gene and the trans CHLI1 gene used for complementation, respectively. PCR analyses using the genomic DNA show that the chli1 complements have the trans CHLI1 gene ( Figure 10C and 10D). In Figure 10D the genomic DNA PCR product sizes in the two complement lanes are smaller than that in the 4A+ lane as we have cloned the CHLI1 cDNA for complementation. The Chlamydomonas CHLI1 protein has about 71% sequence identity to the Arabidopsis CHLI1 protein. Figure 11A shows a stained protein gel. The two chli1 complements and the 4A+ were loaded on an equal Chl basis in each lane in the protein gel ( Figure 11A). As 5A7 lacks Chl, the maximum amount of protein (40 µg) that can be loaded in a mini protein gel, was used ( Figure 11A). Light harvesting complex proteins (LHCs) can barely be detected in the chli1 mutant ( Figure 11A).
Figure 11. SDS-PAGE and Western analyses.

( A) A stained protein gel. Lanes 1, 2, 3 and 4 represent chli1-8, chli1-7, 4A+ and 5A7, respectively. Light harvesting complex (LHC) protein bands are labeled. PS and US denote pre stained and unstained molecular weight protein ladders, respectively. Total cell extract of different strains were loaded on equal Chl basis (4 µg of Chl) in lanes 1, 2 and 3. In lane 4, 40 µg of protein (the maximum amount of protein that can be loaded on a mini protein gel) was loaded as 5A7 lacks Chl. ( B) Western analyses using a CHLI1 antibody generated against the Arabidopsis CHLI1 protein. Lanes 1, 2, 3 and 4 represent chli1-8, chli1-7, 4A+ and 5A7, respectively. CHLI1 (40 kDa) and CHLI2 (42 kDa) proteins detected by the antibody are labeled.
Table 5. Spectrophotometric analyses of chlorophyll in 4A+, chli1, chli1-7 and chli1-8 strains.
Chlorophyll analyses were done on three biological replicates for each strain. Strains were grown mixotrophically in TAP under 15–20 µmol photons m -2s -1. Mean values are shown in the table. Statistical error (± SD) was ≤10% of the values shown. ND: not detected.
| Parameter | 4A+ | Strains chli1 | chli1-7 | chli1-8 |
|---|---|---|---|---|
| Chl/cell (nmoles/cell) | 3.6 × 10 -6 | ND | 2.0 × 10 -6 | 2.5 × 10 -6 |
| Chl a/b ratio | 2.8 | – | 2.7 | 2.8 |
Chlorophyll analyses were done on three biological replicates for each strain. Strains were grown mixotrophically in TAP medium under 15-20 µmol photons m-2s-1. Chlorophyl measurements are in nmol/ml; column I is the uncorrected value, column J is the corrected value. Measurements were taken on 07/02/2013. Chl - chlorophyll
Western analyses of the two chli1 complements with a CHLI1 antibody show that the CHLI1 protein is absent in the chli1 mutant but present in the chli1 complements ( Figure 11B). Western analyses also show that the Arabidopsis CHLI1 antibody detects both the CHLI1 (40 kDa) and CHLI2 (42 kDa) protein in Chlamydomonas as the Chlamydomonas CHLI2 has about 62% sequence identity to the Arabidopsis CHLI1 ( Figure 11B). In the wild type the CHLI2 protein amount is much lower than that of CHLI1. As the chli1 complements are Chl deficient compared to the wild type, the two complement lanes show higher amount of protein loadings ( Figure 11A). Although more protein was loaded in the 5A7 lane in the protein gel compared to that in the 4A+ and the chli1 complement lanes, the CHLI2 protein was barely detectable in 5A7 ( Figure 11B).
Discussion
5A7 is the first chli1 mutant to be identified in C. reinhardtii and in green algae. CHLI1 deletion has affected Chl biosynthesis and photosynthetic growth in the chli1 mutant ( Figure 2). Over-accumulation of photo-excitable PPIX leads to photo-oxidative damage to the cells in presence of light and oxygen 4– 6. The light sensitivity of the chli1 is most probably due to an over-accumulation of PPIX which occurs due to the inactivity of MgChel enzyme which converts PPIX to MgPPIX. Future HPLC (High Performance Liquid Chromatography) analyses of steady state tetrapyrrole intermediates will confirm this hypothesis.
Based on the current molecular analyses, our chli1 mutant has a deletion of at least nine genes (including the CHLI1 gene). Currently we are investigating the exact insertion point of the pUC ori end of the plasmid in the chli1 genome ( Figure 1). This will provide us with a precise estimate of the number of gene deletions in chli1. Although complementation of chli1 with the CHLI1 gene restored Chl biosynthesis, tolerance to high light levels and photo-autotrophic growth, chli1 complements are still Chl deficient to some extent ( Table 5). This is probably due to a lower expression of the CHLI1 protein in these chli1 complements ( Figure 11). Semi-quantitative RT-PCR shows that the CHLI2 transcript level in chli1 is much lower than that in the wild type strain ( Figure 7). Western analyses show that the CHLI2 protein level is severely reduced in the chli1 mutant ( Figure 11). Real Time PCR analyses can be used to confirm whether the reduction in the CHLI2 protein level is due to a low abundance of the CHLI2 transcript. Additionally, the roles of any of the other missing eight genes in Chl biosynthesis cannot be ruled out as currently the functions of these genes are unknown.
In Arabidopsis, it has been shown that CHLI2 does play a limited role in Chl biosynthesis in the absence of CHLI1 12– 15. 5A7 possesses an intact CHLI2 gene but the CHLI2 protein is barely detectable in the mutant. This raises two questions:
1) Is the low abundance of the CHLI2 protein a general effect or is it a specific effect of the CHLI1 mutation?
2) Is the total absence of Chl in strain 5A7 due to the specific absence of CHLI1 or due to the absence of both CHLI1 and the near absence of CHLI2 protein?
The first question can be addressed by performing Western analyses of 5A7 with antibodies raised against any non-photosynthetic and/or photosynthetic protein. If the low abundance of the CHLI2 protein is due to a general effect of the mutation, there will be an overall reduction of different cellular proteins. The second question can be addressed by overexpressing CHLI2 in 5A7 to see if Chl biosynthesis can occur in the absence of CHLI1 or by silencing CHLI2 in the wild type strain using RNA interference or micro RNA based techniques.
Norflurazon (NF) causes photo-oxidative damage to the chloroplast by inhibiting carotenoid biosynthesis. 24, 27– 31. In Arabidopsis MgPPIX is hypothesized to be a retrograde signal from the chloroplast to the nucleus on the basis of data obtained with mutants that are defective in the NF, induced down-regulation of the transcription of the light harvesting complex protein B(LHCB) expression [gun (genomes uncoupled) phenotype] 27, 28. In Arabidopsis, there are controversies regarding whether chli1 mutants are gun mutants 12, 29– 31. Our chli1 lacks detectable LHC proteins on the stained gel ( Figure 11A). LHC transcript and protein analyses in NF treated and untreated cells and HPLC based analyses of steady state tetrapyrroles in chli1 will help to confirm if it is a " gun" mutant. In summary, in the future our chli1 mutant can be used to clarify the functional role of CHLI1 and CHLI2 in Chl biosynthesis as well as their roles in retrograde signaling in C. reinhardtii.
Acknowledgements
We would like to thank Dr. Krishna K. Niyogi (UC, Berkeley) for providing the 4A+ strain and the pBC1 plasmid that were used for mutagenesis, Dr. Saul Purton (University College London, UK) for providing the complementation vector pDBle and Dr. Robert Larkin (Michigan State University) for giving us the Arabidopsis CHLI antibodies. We are grateful to Dr. Bernhard Grimm and Dr. Pawel Brzezowski (Humboldt University, Berlin, Germany) for providing us the FDX3, UP1, UP2, and AMT gene specific primers for PCR analyses. We would also like to thank Dr. Leos Kral (University of West Georgia) for allowing us to use his nano spectrophotometer and Dr. Anastasios Melis (UC, Berkeley) for allowing us to perform the protein and Western analyses at his laboratory.
Funding Statement
This project was supported by several grants awarded to Dr. Mautusi Mitra. These are: the start-up grant of the University of West Georgia (UWG), the Faculty Research Grant by the UWG College of Science and Mathematics, the Internal Development Grant by the UWG office of Research and Sponsored Project, the Research Incentive grant by the UWG College of Science and Mathematics, the UWG Student Research Assistance Program (SRAP) grant and the UWise-BOR-STEM II grant from UWG.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
v1; ref status: indexed
References
- 1.Merchant SS, Prochnik SE, Vallon O, et al. : The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science. 2007;318(5848):245–250 10.1126/science.1143609 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Dent RM, Haglund CM, Chin BL, et al. : Functional genomics of eukaryotic photosynthesis using insertional mutagenesis of Chlamydomonas reinhardtii. Plant Physiol. 2005;137(2):545–556 10.1104/pp.104.055244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Gutman BL, Niyogi KK: Chlamydomonas and Arabidopsis. A dynamic duo. Plant Physiol. 2004;135(2):607–610 10.1104/pp.104.041491 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Beale SI: Chloroplast Signaling: Retrograde Regulation Revelations. Curr Biol. 2011;21(10):R391–R393 10.1016/j.cub.2011.03.072 [DOI] [PubMed] [Google Scholar]
- 5.Moulin M, Smith AG: Regulation of tetrapyrrole biosynthesis in higher plants. Biochem Soc Trans. 2005;33(Pt 4):737–742 10.1042/BST0330737 [DOI] [PubMed] [Google Scholar]
- 6.Tanaka R, Tanaka A: Tetrapyrrole biosynthesis in higher plants. Anu Rev Plant Biol. 2007;58:321–346 10.1146/annurev.arplant.57.032905.105448 [DOI] [PubMed] [Google Scholar]
- 7.Walker CJ, Willows RD: Mechanism and regulation of Mg-chelatase. Biochem J. 1997;327(Pt 2):321–333 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Walker CJ, Weinstein JD: The Magnesium-Insertion Step Of Chlorophyll Biosynthesis Is A two-Stage Reaction. Biochem J. 1994;299(Pt 1):277–284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Masuda T: Recent overview of the Mg branch of the tetrapyrrole biosynthesis leading to chlorophylls. Photosynth Res. 2008;96(2):121–143 10.1007/s11120-008-9291-4 [DOI] [PubMed] [Google Scholar]
- 10.Apchelimov AA, Soldatova OP, Ezhova TA, et al. : The analysis of the ChlI 1 and ChlI 2 genes using acifluorfen-resistant mutant of Arabidopsis thaliana. Planta. 2007;225(4):935–943 10.1007/s00425-006-0390-1 [DOI] [PubMed] [Google Scholar]
- 11.Ikegami A, Yoshimura N, Motohashi K, et al. : The CHLI1 subunit of Arabidopsis thaliana. magnesium chelatase is a target protein of the chloroplast thioredoxin. J Biol Chem. 2007;282(27):19282–19291 10.1074/jbc.M703324200 [DOI] [PubMed] [Google Scholar]
- 12.Huang YS, Li HM: Arabidopsis CHLI2 Can Substitute for CHLI1. Plant Physiol. 2009;150(2):636–645 10.1104/pp.109.135368 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kobayashi K, Mochizuki N, Yoshimura N, et al. : Functional analysis of Arabidopsis thaliana isoforms of the Mg-chelatase CHLI subunit. Photochem Photobiol Sci. 2008;7(10):1188–1195 10.1039/b802604c [DOI] [PubMed] [Google Scholar]
- 14.Koncz C, Mayerhofer R, Koncz-Kalman Z, et al. : Isolation of a gene encoding a novel chloroplast protein by T-DNA tagging in Arabidopsis thaliana. EMBO J. 1990;9(5):1337–1346 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rissler HM, Collakova E, DellaPenna D, et al. : Chlorophyll biosynthesis. Expression of a second Chl I gene of magnesium chelatase in Arabidopsis supports only limited chlorophyll synthesis. Plant Physiol. 2002;128(2):770–779 10.1104/pp.010625 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Soldatova O, Apchelimov A, Radukina N, et al. : An Arabidopsis mutant that is resistant to the protoporphyrinogen oxidase inhibitor acifluorfen shows regulatory changes in tetrapyrrole biosynthesis. Mol Genet Genomics. 2005;273(4):311–318 10.1007/s00438-005-1129-6 [DOI] [PubMed] [Google Scholar]
- 17.Gorman DS, Levine RP: Cytochrome f and plastocyanin: their sequence in the photosynthetic electron transport chain of Chlamydomonas reinhardi. Proc Natl Acad Sci U S A. 1965;54(6):1665–1669 10.1073/pnas.54.6.1665 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sueoka N: Mitotic Replication of Deoxyribonucleic Acid in Chlamydomonas reinhardi. Proc Natl Acad Sci U S A. 1960;46(1):83–91 10.1073/pnas.46.1.83 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Davies JP, Weeks DP, Grossman AR: Expression of the arylsulfatase gene from the beta 2-tubulin promoter in Chlamydomonas reinhardi. Nucleic Acids Res. 1992;20(12):2959–2965 10.1093/nar/20.12.2959 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kindle KL, Schnell RA, Fernández E, et al. : Stable nuclear transformation of Chlamydomonas using the Chlamydomonas gene for nitrate reductase. J Cell Biol. 1989;109(6 Pt 1):2589–2601 10.1083/jcb.109.6.2589 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lowry OH, Rosebrough NJ, Farr AL, et al. : Protein measurement with the Folin phenol reagent. J Biol Chem. 1951;193(1):265–275 [PubMed] [Google Scholar]
- 22.Smith BM, Morrissey PJ, Guenther JE, et al. : Response of the Photosynthetic Apparatus in Dunaliella salina (Green Algae) to Irradiance Stress. Plant Physiol. 1990;93(4):1433–1440 10.1104/pp.93.4.1433 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Laemmli UK: Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227(5259):680–685 10.1038/227680a0 [DOI] [PubMed] [Google Scholar]
- 24.Adhikari ND, Orler R, Chory J, et al. : Porphyrins promote the association of GENOMES UNCOUPLED 4 and a Mg-chelatase subunit with chloroplast membranes. J Biol Chem. 2009;284(37):24783–24796 10.1074/jbc.M109.025205 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Arnon DI: Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949;24(1):1–15 10.1104/pp.24.1.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Melis A, Spangfort M, Andersson B: Light-Absorption And Electron-Transport Balance Between Photosystem II And Photoystem I In Spinach Chloroplasts. Photochem and Photobiol. 1987;45(1):129–136 10.1111/j.1751-1097.1987.tb08413.x [DOI] [Google Scholar]
- 27.Nott A, Jung HS, Koussevitzky S, et al. : Plastid-to-nucleus retrograde signaling. Ann Rev Plant Biol. 2006;57:739–759 10.1146/annurev.arplant.57.032905.105310 [DOI] [PubMed] [Google Scholar]
- 28.Susek RE, Ausubel FM, Chory J: Signal transduction mutants of Arabidopsisuncouple nuclear CAB and RBCS gene expression from chloroplast development. Cell. 1993;74(5):787–799 10.1016/0092-8674(93)90459-4 [DOI] [PubMed] [Google Scholar]
- 29.Gadjieva R, Axelsson E, Olsson U, et al. : Analysis of gunphenotype in barley magnesium chelatase and Mg-protoporphyrin IX monomethyl ester cyclase mutants. Plant Physiol Biochem. 2005;43(10–11):901–908 10.1016/j.plaphy.2005.08.003 [DOI] [PubMed] [Google Scholar]
- 30.Mochizuki N, Brusslan JA, Larkin R, et al. : Arabidopsisgenomes uncoupled 5 (GUN5) mutant reveals the involvement of Mg-chelatase H subunit in plastid-to-nucleus signal transduction. Proc Natl Acad Sci U S A. 2001;98(4):2053–2058 10.1073/pnas.98.4.2053 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Strand Å, Asami T, Alonso J, et al. : Chloroplast to nucleus communication triggered by accumulation of Mg-protoporphyrinIX. Nature. 2003;421(6918):79–83 10.1038/nature01204 [DOI] [PubMed] [Google Scholar]