

Abstract
The goal of this study was to optimize the transcranial magnetic stimulation (TMS) protocol for acquiring a reliable estimate of corticospinal excitability (CSE) using single-pulse TMS. Moreover, the minimal number of stimuli required to obtain a reliable estimate of CSE was investigated. In addition, the effect of two frequently used stimulation intensities [110% relative to the resting motor threshold (rMT) and 120% rMT] and gender was evaluated. Thirty-six healthy young subjects (18 males and 18 females) participated in a double-blind crossover procedure. They received 2 blocks of 40 consecutive TMS stimuli at either 110% rMT or 120% rMT in a randomized order. Based upon our data, we advise that at least 30 consecutive stimuli are required to obtain the most reliable estimate for CSE. Stimulation intensity and gender had no significant influence on CSE estimation. In addition, our results revealed that for subjects with a higher rMT, fewer consecutive stimuli were required to reach a stable estimate of CSE. The current findings can be used to optimize the design of similar TMS experiments.
Introduction
Corticospinal (CS) excitability can be estimated by measuring motor evoked potentials (MEPs) elicited by transcranial magnetic stimulation (TMS). MEPs provide information about the functionality of the human nervous system and are commonly measured in both research and clinical settings for diagnostic and therapeutic purposes. Because the MEP amplitude is highly variable within the same subject [1]–[4], consecutive measurements are required to obtain a reliable estimate of the CSE of the stimulated cortical site. Although the nature of MEP amplitude variability remains mainly unclear, randomness in processes such as the firing of neurons and motor neuron transmission might play a role [3], [5]. Additionally, trial-to-trial fluctuations in MEP amplitude might be caused by rapid fluctuating changes in cortical and spinal excitability [1], [6], [7]. It is reported that other physiological factors influence the MEP amplitude as well, such as pre-stimulus contraction [1], [8], attention [9], arousal, desynchronization of action potentials [3], and afferent feedback [10]. Besides these physiological factors, physical parameters such as coil orientation [11], optimal scalp location [12] and environmental noise might also play an important role in the variability of the MEP amplitude.
Whereas, in most studies 6 to 10 pulses were applied to determine CSE of the region of interest [13], it is still unclear how many TMS pulses are required to acquire a reliable estimate of CSE. Maximizing reliability is important for the assessment of neurophysiological effects caused by interventions that affect CSE. Although TMS has already been used for several decades, previous studies showed no consistency regarding the number of pulses used for measuring CSE.
Another relevant question is whether the number of pulses is mediated by factors such as stimulation intensity and/or gender. Previously, a sigmoidal relationship between stimulation intensity and average MEP amplitude was reported [5], showing that well-tolerated stimulation intensities, which are represented in the middle of the recruitment curve are sensitive for detecting changes in CSE as they are not subjected to either floor or ceiling effects. When applying single-pulse TMS at rest, two intensities relative to the resting motor threshold (rMT) are frequently applied to measure CSE, respectively 110% rMT [14]–[16] and 120% rMT [17]–[19]. However, it is not clear whether the number of pulses required to acquire a reliable estimate of CSE is mediated by these different stimulation intensities. Another parameter that might influence CME measurement is gender, as previous findings indicated that MEP variation was more pronounced in females [5], [20], [21]. Nonetheless, it should be mentioned that increased MEP variability was only reported using a paired-pulse TMS paradigm for measuring changes in intracortical inhibition [20], [22]. Yet, it is not clear whether similar effects will be found when single-pulse TMS is used.
The goal of the current study was to optimize the TMS protocol for acquiring a reliable estimate of CSE. Therefore, the first aim of the study was to examine the number of stimuli required to obtain a reliable estimate of CSE. The second aim was to evaluate the effect of two frequently used intensities (110% rMT and 120% rMT) on the reliability of CSE. And finally, we aimed to investigate whether gender acts as a covariate in estimating CSE.
Materials and Methods
Ethics Statement
Subjects provided written informed consent and experimental procedures were approved by the Ethics Committee of the University of Hasselt. The study conforms to the principles stated in the Declaration of Helsinki.
Subjects
Thirty-six subjects, 18 men and 18 women, aged 19 to 24 years (mean age±SD = 20.47±1.21) participated in this study. Handedness was assessed with the Edinburgh Handedness Inventory [23]. All participants were right-handed (mean LQ±SD = 91.16±13.32) and naïve for TMS. The resting motor threshold (rMT) ranged between 32% and 50% of the maximum stimulator output (mean rMT±SD = 39.81±4.70). Before inclusion all subjects were screened for TMS contra-indications [24]; and for pathologies associated with peripheral and/or central sensory dysfunction and for central nervous system-acting, psychotropic or antiepileptic medication intake.
Experimental Design
Prior to the start of the experiment, subjects were asked to report their level of attention, fatigue and arousal using a visual analogue scale (VAS) score. Next, the hotspot of the first dorsal interosseous (FDI) muscle and the rMT was determined using TMS. Then, subjects moved on to a double-blind crossover procedure. Both, subjects and the experimenter applying TMS were blinded for the stimulation intensity. Subjects received two blocks of 40 consecutive TMS pulses, one block at 110% rMT and another at 120% rMT. Neither the experimenter nor the subjects received feedback (visualization of the MEPs). Subjects were instructed to keep their eyes open during the experiment. After the experiment, VAS scores were assessed again to monitor changes in attention, fatigue and arousal during the experiment, which lasted approximately 30 min.
Transcranial Magnetic Stimulation (TMS)
Magnetic stimuli (Magstim BiStim2, Whitland, South West Wales, UK) were delivered using a 70-mm loop-diameter figure-of-eight coil. For each subject, an orthogonal 1×1 cm coordinate system was marked on a swimming cap with references to anatomical landmarks (left and right external auditory meatus, occiput and vertex). Then, the hotspot (scalp location resulting the highest mean MEP after five consecutive magnetic stimuli) of the relaxed FDI muscle was determined. The coil was positioned on the left hemisphere with the coil handle pointing backward and rotated 45° away from the midsagital line [11]. Next, the rMT was defined as the lowest stimulation intensity evoking MEPs with an amplitude larger than 50 µV peak-to-peak in at least five of ten consecutive trials. Finally, two blocks of 40 consecutive TMS pulses (40 pulses at 110% rMT and 40 pulses at 120% rMT) were administered in a randomized order. A two-minute break was provided between blocks. The interval between TMS stimuli was randomized (5–8 s.).
Electromyographic Recordings (EMG)
Electromyographic signals from the FDI muscle were continuously monitored and measured using EMG (Bagnoli-16, Delsys Inc, Boston, USA). After amplification (gain = 1000), band pass filtering (4–1500 Hz) and 50/60 Hz noise elimination (Humbug, Quest Scientific, North Vancouver, Canada) the recorded EMG signals were digitized at 5000 Hz (CED Signal Version 3.03, Cambridge Electronic Design, Cambridge, UK) and were stored on a laboratory computer for offline analysis.
Data Analysis
Visual Analogue Scale (VAS)
Because the Shapiro-Wilk test indicated that the data was not normally distributed, the Wilcoxon signed-rank test was used for statistical analysis of the VAS scores. The level of significance was set at p<0.05.
MEP data analysis
Before analysis, individual MEPs were screened for artefacts and voluntary contraction; and excluded (<1%) if the root mean square EMG exceeded 5 µV during the 50-ms period immediately preceding the onset of the TMS pulse.
In the main analysis, data of all 36 subjects were analysed. For each subject, the average MEP was calculated for subsets of consecutive stimuli: 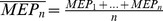 , where n: 2…40. In this experiment
, where n: 2…40. In this experiment 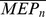 can be assumed as most accurate estimate of the true underlying MEP value. The main goal of this study was to define the number of consecutive stimuli needed for to approach
can be assumed as most accurate estimate of the true underlying MEP value. The main goal of this study was to define the number of consecutive stimuli needed for to approach 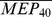 . In order to evaluate the accuracy of the estimates, a 95% confidence interval (CI) was calculated using all 40 stimuli for each subject (see figure 1). Based on both the
. In order to evaluate the accuracy of the estimates, a 95% confidence interval (CI) was calculated using all 40 stimuli for each subject (see figure 1). Based on both the 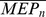 values and the CI, it is possible to determine whether
values and the CI, it is possible to determine whether 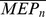 is included in CI, yielding a binary variable (0 = not included in the CI, 1 = included in the CI). The procedure described above was applied to all subjects for both stimulation intensities. Additionally, we analysed if either gender and/or stimulation intensity had an effect on the total number of stimuli required to obtain a reliable estimate of CSE. The change in attention (post TMS – pre TMS), arousal, and fatigue; and rMT were included as covariates in our model. The level of significance was set at p<0.05. In order to investigate these effects a generalized estimating equation (GEE) analysis was used (SAS v9.2). More specifically, this technique estimates the parameters of a general linear model taking into account the correlation between two measurements of the same subject with different stimulation intensities. In the current analysis a binary response variable indicates whether an estimate of the true underlying MEP value (
is included in CI, yielding a binary variable (0 = not included in the CI, 1 = included in the CI). The procedure described above was applied to all subjects for both stimulation intensities. Additionally, we analysed if either gender and/or stimulation intensity had an effect on the total number of stimuli required to obtain a reliable estimate of CSE. The change in attention (post TMS – pre TMS), arousal, and fatigue; and rMT were included as covariates in our model. The level of significance was set at p<0.05. In order to investigate these effects a generalized estimating equation (GEE) analysis was used (SAS v9.2). More specifically, this technique estimates the parameters of a general linear model taking into account the correlation between two measurements of the same subject with different stimulation intensities. In the current analysis a binary response variable indicates whether an estimate of the true underlying MEP value (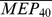 ), based on a certain number of repetitions, falls within the confidence interval (response = 1) or not (response = 0). This yields a vector consisting of the values 0 and 1 for each subject. Since there exists no convenient specification of the joint distribution for such a vector of binary responses, Generalized Estimating Equations (GEE) was introduced as a valuable alternative for maximum likelihood estimation [25], [26]. In more detail, within the GEE approach a marginal model is considered with a mean function of the predictor variables similar to the well-known linear models in regression and ANOVA. Furthermore, within the GEE approach this is combined with a known variance function and a working correlation matrix. The main idea is to generalize the univariate likelihood equations by means of a covariance matrix of the response vector.
), based on a certain number of repetitions, falls within the confidence interval (response = 1) or not (response = 0). This yields a vector consisting of the values 0 and 1 for each subject. Since there exists no convenient specification of the joint distribution for such a vector of binary responses, Generalized Estimating Equations (GEE) was introduced as a valuable alternative for maximum likelihood estimation [25], [26]. In more detail, within the GEE approach a marginal model is considered with a mean function of the predictor variables similar to the well-known linear models in regression and ANOVA. Furthermore, within the GEE approach this is combined with a known variance function and a working correlation matrix. The main idea is to generalize the univariate likelihood equations by means of a covariance matrix of the response vector.
Figure 1. Data for subject 36 (a) and 34 (b) is illustrated.

The Y-axis shows the MEP amplitude (mV), while the number of TMS stimuli (n) is shown on the X-axis. White dots represent the individual (raw) MEPs, whereas the black dots represent the average of consecutive MEPs (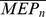 ). Dashed lines represent the 95% confidence interval (CI), which is based upon all 40 stimuli. The upper panel (a) illustrates data that was included in the statistical analysis (slope estimate: 0.007; p = 0.355). For this particular subject, 8 consecutive stimuli were sufficient to enter the CI. The lower panel (b) shows data that has been excluded due to a significant change in slope over time (slope estimate: 0.062; p<0.001).
). Dashed lines represent the 95% confidence interval (CI), which is based upon all 40 stimuli. The upper panel (a) illustrates data that was included in the statistical analysis (slope estimate: 0.007; p = 0.355). For this particular subject, 8 consecutive stimuli were sufficient to enter the CI. The lower panel (b) shows data that has been excluded due to a significant change in slope over time (slope estimate: 0.062; p<0.001).
Although the MEP amplitude is highly variable, no significant trend in MEP amplitude over time is expected during rest [27]. Therefore, we performed an identical supplementary analysis. However, if a significant change (slope) in CSE over time was detected (using a simple linear regression analysis) within one of the stimulation intensities, the data corresponding to this intensity was excluded from the analysis. After excluding subjects 2 [Male (M)], 11 (M), 12 (M), 16 [Female (F)], 17 (F) and 26 (F) for 110% rMT and subjects 8 (F), 10 (F), 14 (M) and 34 (F) for 120% rMT, respectively 30 (110% rMT) and 32 (120% rMT) subjects were maintained in each condition for statistical analysis (see figure 1 for an example of included/excluded data).
Results
Estimation of Corticospinal Excitability
Table 1 shows the probability that the average of these stimuli will be in the CI for the total sample irrespective of intensity and gender as a function of the number of consecutive stimuli. Exploratory data analysis revealed that a probability of 1 was reached only after at least 30 consecutive stimuli.
Table 1. Probability table.
| Number of consecutive stimuli | Probability of hitting the 95% CI |
| 2 | 0.39 |
| 3 | 0.30 |
| 4 | 0.32 |
| 5 | 0.38 |
| 6 | 0.38 |
| 7 | 0.44 |
| 8 | 0.50 |
| 9 | 0.55 |
| 10 | 0.58 |
| 11 | 0.58 |
| 12 | 0.65 |
| 13 | 0.66 |
| 14 | 0.65 |
| 15 | 0.71 |
| 16 | 0.78 |
| 17 | 0.79 |
| 18 | 0.82 |
| 19 | 0.83 |
| 20 | 0.86 |
| 21 | 0.86 |
| 22 | 0.86 |
| 23 | 0.89 |
| 24 | 0.90 |
| 25 | 0.92 |
| 26 | 0.99 |
| 27 | 0.99 |
| 28 | 0.99 |
| 29 | 0.99 |
| 30–40 | 1.00 |
The number of consecutive stimuli required as a function of the probability of hitting the 95% confidence interval (CI).
The GEE analysis (Table 2) showed only a significant effect for number of consecutive stimuli, indicating that the probability that the estimate will be in the CI increased when the number of consecutive stimuli increased (p = 0.033). No significant effects were reported for stimulation intensity or gender (all, p>0.05; see Figure 2). Explorative data analysis showed that, at least 26 consecutive stimuli were required to reach a probability of 1, when stimulating at an intensity of 110% rMT and at least 30 consecutive stimuli at 120% rMT. For females 30 consecutive stimuli were required, while for males 26 consecutive stimuli were needed to reach a probability of 1.
Table 2. Generalized estimating equation (GEE) analysis.
| Parameter | Estimate | Z-value | p-value |
| Intercept | −4.7675 | −3.24 | 0.001 |
| Number of stimuli | 0.2333 | 2.13 | 0.033 |
| Resting motor threshold | 0.0715 | 2.35 | 0.019 |
| Resting motor threshold× Number of stimuli | −0.0019 | −0.71 | 0.481 |
Estimates and p-values are shown for the number of consecutive stimuli, arousal, fatigue, resting motor threshold and the interaction between arousal and fatigue. P-values in bold highlight a significant effect.
Figure 2. Results for gender (a) and intensity (b) based upon the raw data are illustrated.

The Y-axis shows probability of inclusion in the 95% CI, while the number of TMS stimuli (n) is shown on the X-axis. The upper panel (a) illustrates the probability of inclusion in the 95% CI for females (white dots) and males rMT (black dots). The lower panel (b) illustrates the probability of inclusion in the 95% CI for the stimulation intensity of 110% rMT (white dots) and 120% rMT (black dots).
Resting Motor Threshold
The GEE analysis showed a significant effect of rMT on CSE estimates (p = 0.019). Moreover, as shown in Table 3, for subjects with a higher rMT, fewer consecutive stimuli were required to reach a stable estimate of CSE.
Table 3. The number of TMS stimuli required to reach a probability of 1.0 for hitting the 95% CI was estimated using the GEE analysis for different levels of resting motor threshold (rMT).
| rMT(% max. stim.output) | TMS stimuli requiredfor probability = 1.0 |
| 32–34 | 28 |
| 35–42 | 27 |
| 43–47 | 26 |
| 47–50 | 25 |
Visual Analogue Scale Scores
Table 4 shows the data for attention, fatigue and arousal obtained prior and after the experiment. For the entire group a significant decrease in attention (p<0.001), arousal (p<0.001) and a significant increase in fatigue (p = 0.014) were reported during the course of the experiment. Similar results were reported for males (all, p<0.05). However, for females, there was only a significant decrease in arousal (p = 0.026). Attention and fatigue did not change significantly over time in this subgroup (all, p>0.05).
Table 4. Visual Analogue Scales (VAS).
| Parameter | PRE | POST | Z-value | p-value | |||
| Mean | SD | Mean | SD | ||||
| Allsubjects | 6.30 | (1.70) | 5.31 | (1.80) | −3.567 | <0.001 | |
| Attention | Male | 6.67 | (1.52) | 5.47 | (2.03) | −3.435 | <0.001 |
| Female | 5.93 | (1.83) | 5.16 | (1.58) | −1.801 | 0.074 | |
| Allsubjects | 5.71 | (1.99) | 5.24 | (2.22) | −2.436 | 0.014 | |
| Fatigue | Male | 6.16 | (1.70) | 5.23 | (2.03) | −2.746 | 0.004 |
| Female | 5.27 | (2.19) | 5.26 | (2.46) | −0.632 | 0.543 | |
| Allsubjects | 3.32 | (2.37) | 2.23 | (2.11) | −3.441 | <0.001 | |
| Arousal | Male | 2.53 | (1.96) | 1.76 | (2.01) | −2.842 | 0.003 |
| Female | 4.10 | (2.53) | 2.69 | (2.15) | −2.200 | 0.026 | |
The mean VAS score ± standard deviation (SD) is shown for attention (0 = no attention; 10 = maximal attention), fatigue (0 = no fatigue; 10 = maximal fatigue) and arousal (0 = no arousal; 10 = maximal arousal). Measurements were obtained prior (PRE) and after (POST) the experiment. Data is shown for all subjects and for males and females separately. P-values in bold highlight a significant effect between PRE and POST measurement.
In addition, the GEE analysis revealed no effect for none of these covariates on the CSE estimates (all, p>0.05), indicating that neither the change in attention, nor the change in arousal or fatigue influenced the CSE estimates.
Supplemental Data Analysis
Similar results were obtained after exclusion of subjects who showed a significant trend in MEP amplitude over time (for details, see Supporting Information). Moreover, the GEE analysis (Table S2) revealed only significant effects for the number of pulses (p<0.001) and rMT (p = 0.005, Table S3). There were no effects for gender, attention, arousal, fatigue, or their interactions (all, p>0.05). The number of consecutive pulses required to obtain the most reliable estimate for CSE was at least 26 (Table S1).
Discussion
The primary aim of the present study was to optimize the TMS protocol for acquiring a reliable estimate of CSE. During our measurements, we attempted to control for the factors affecting MEP variability, such as pre-stimulus contraction [1], [8], attention [9], arousal and coil orientation [11]. Our results revealed that at least 30 consecutive stimuli were required to obtain the most accurate estimate of CSE. In addition, stimulation intensity and/or gender had no significant effect on the estimation of CSE.
As the MEP amplitude is highly variable within subjects [1]–[4] several consecutive TMS stimuli were applied in most studies. Until now, no clear advice was available with respect to the amount of stimuli required to obtain reliable estimates of CSE. This basic methodological information might play a crucial role in the development of reliable TMS protocols.
Interestingly, our results revealed no significant differences between two frequently used intensities (110% and 120% rMT). As illustrated in studies measuring TMS recruitment curves [28]–[30], these intensities are well situated in the rising part of the (sigmoidal) curve, making them sensitive candidates for evaluating shifts in CSE.
In the current study no significant gender effect was reported. This finding was in line with Pitcher et al. (2003), who did not found a main-effect of gender on MEP variation when exploring TMS recruitment curve characteristics. However, they did report a significant interaction between stimulus intensity, age, rMT and gender. Moreover, females tended to have increased MEP variability than males, but age and rMT were much stronger modulators of MEP variation than gender [5]. In contrast with our results, previous findings [20], [21] indicated that MEP variation was greater in females (due to changes in ovarian steroid levels during the various stages of the menstrual cycle). Nevertheless, we should be careful with comparing our results with the gender-related differences reported in earlier studies because different TMS protocols were used focussing primarily on inhibitory networks [20], [22]. More specifically, it is thought that variability in inhibition is due to allosteric action of progesterone-derived neurosteroids on GABAA receptor transmission [31], [32], indicating that ovarian hormones exert an effect on brain function. However, it might be argued that gender-related variability in specific intracortical γ-aminobutyric acid A (GABAA)-mediated inhibitory networks within M1 may not necessary translate into variability in the activation of the cortical network targeted by single-pulse TMS.
Our results revealed that rMT contributed significantly to the estimation of CSE. More specifically, for subjects with a higher rMT, fewer stimuli were required to reach a stable estimate of CSE. This is in line with Smith et al. (2011), indicating that subjects with high rMTs showed less MEP variability at a given stimulus intensity, as compared to subjects with low rMTs [30]. Unfortunately, the current literature offers no obvious neurophysiological explanation for this finding.
Though the current results are clear, we need to be cautious with the interpretation of our results. Firstly, we have to be careful with extrapolating our findings to other populations. Since our data was obtained in healthy young subjects, a different number of consecutive stimuli might be required when estimating CSE in elderly or neurodegenerative populations. As compared to young subjects, trial-to-trial variability was shown to be increased in elderly, specifically at low, near threshold intensities [5]. In patients suffering from neurodegenerative disease, it is reported that MEP amplitudes are often reduced or even absent [33]–[35]. Secondly, estimates of CSE can be influenced by the experimental set-up. For example, the use of navigated TMS, different coil types and shapes, EMG hardware configuration and noise elimination can affect variability and reliability of the measurements. With respect to TMS navigation, a recent study comparing non-navigated and MRI-guided navigated TMS [36] reported that the stability of MEPs increased significantly (lower MEP variability) when MRI-guided navigated TMS was used. In contrast, findings from Jung et al. (2010) revealed no significant difference in MEP variability and reproducibility between non-navigated and optically tracked TMS navigation [37]. Although we did not use MRI-guided navigated TMS in the current study, there is evidence that experimenters using non-navigated TMS can reach a performance level, which is comparable with optically tracked navigated TMS measurements as indicated by Jung et al. (2010).
With respect to reliability and accuracy of CSE, the triple-stimulation technique [38] has shown to be superior as compared to the current (conventional) technique. Nonetheless this advanced technique has also limitations, as it is more complex, only suitable for distal muscles and less comfortable for the subject. Furthermore, as the majority of the TMS studies are performed with the conventional technique, our results yield important information for designing TMS experiments using the conventional technique.
Although, we used a standardized procedure and attempted to control for attention, fatigue and arousal, these parameters can still change during the course of the experiment, as illustrated by our results. Nonetheless all subjects were comprehensively briefed with respect to the experimental procedures, the perceived changes in attention, arousal and fatigue throughout the experiment might be explained by an increased level of arousal and attention and a decrease level of fatigue prior to the experiment due to uncertainty about their first TMS experience [39]. Therefore, a familiarization session prior to the actual experiment might be recommended to minimize these effects. With respect to attention and arousal of the subjects, our results are in line with Hess et al. (1987). They reported that the threshold for excitation of the relaxed muscle showed some variation over time, but that is was not related to attention or alertness of the subject [40]. Furthermore, during TMS measurements background EMG was monitored to make sure the level of muscle relaxation was constant. With respect to fatigue in particular, previous studies reported that MEP amplitude decreased due to fatigue or to increased relaxation of the stimulated muscle [41], [42]. This might explain why we found a trend in the change of MEP amplitude over time in some subjects. However, excluding these subjects did not dramatically change our results. Furthermore, when subjects who showed a trend in MEP amplitude over time were included in the analysis, the current results showed that, overall, the application of only 4 extra pulses was sufficient to obtain a reliable estimation of CSE.
Importantly, although VAS scales indicated that subject reported changes in attention, fatigue and arousal, these perceived changes did not contribute significantly to the estimation of CSE as revealed by the GEE analysis.
In summary we can conclude that with the current TMS procedure, at least 30 consecutive stimuli are required to obtain the most reliable estimate for CSE. In addition, no significant differences were reported for gender or stimulation intensity. rMT, however, did contribute significantly to the estimation of CSE. More specifically, for subjects with a higher rMT, fewer stimuli were required. The current findings can be used to optimize the design of TMS experiments.
Supporting Information
Probability table. The number of consecutive stimuli required as a function of the probability of hitting the 95% confidence interval (CI).
(DOCX)
Generalized estimating equation (GEE) analysis. Estimates and p-values are shown for the number of consecutive stimuli, arousal, fatigue, resting motor threshold and the interaction between arousal and fatigue. P-values in bold highlight a significant effect.
(DOCX)
The number of TMS stimuli required to reach a probability of 1.0 for hitting the 95% CI was estimated using the GEE analysis for different levels of resting motor threshold (rMT).
(DOCX)
Funding Statement
This work is supported by the Flanders Fund for Scientific Research (G075810). Koen Cuypers is supported by the Special Research Fund UHasselt. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Kiers L, Cros D, Chiappa KH, Fang J (1993) Variability of motor potentials evoked by transcranial magnetic stimulation. Electroencephalogr Clin Neurophysiol 89: 415–423. [DOI] [PubMed] [Google Scholar]
- 2. Roy Choudhury K, Boyle L, Burke M, Lombard W, Ryan S, et al. (2011) Intra subject variation and correlation of motor potentials evoked by transcranial magnetic stimulation. Ir J Med Sci 180: 873–880. [DOI] [PubMed] [Google Scholar]
- 3. Magistris MR, Rosler KM, Truffert A, Myers JP (1998) Transcranial stimulation excites virtually all motor neurons supplying the target muscle. A demonstration and a method improving the study of motor evoked potentials. Brain 121 (Pt 3): 437–450. [DOI] [PubMed] [Google Scholar]
- 4. Rosler KM, Petrow E, Mathis J, Aranyi Z, Hess CW, et al. (2002) Effect of discharge desynchronization on the size of motor evoked potentials: an analysis. Clin Neurophysiol 113: 1680–1687. [DOI] [PubMed] [Google Scholar]
- 5. Pitcher JB, Ogston KM, Miles TS (2003) Age and sex differences in human motor cortex input-output characteristics. J Physiol 546: 605–613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Ellaway PH, Davey NJ, Maskill DW, Rawlinson SR, Lewis HS, et al. (1998) Variability in the amplitude of skeletal muscle responses to magnetic stimulation of the motor cortex in man. Electroencephalogr Clin Neurophysiol 109: 104–113. [DOI] [PubMed] [Google Scholar]
- 7. Funase K, Miles TS, Gooden BR (1999) Trial-to-trial fluctuations in H-reflexes and motor evoked potentials in human wrist flexor. Neurosci Lett 271: 25–28. [DOI] [PubMed] [Google Scholar]
- 8. Darling WG, Wolf SL, Butler AJ (2006) Variability of motor potentials evoked by transcranial magnetic stimulation depends on muscle activation. Exp Brain Res 174: 376–385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Mars RB, Bestmann S, Rothwell JC, Haggard P (2007) Effects of motor preparation and spatial attention on corticospinal excitability in a delayed-response paradigm. Exp Brain Res 182: 125–129. [DOI] [PubMed] [Google Scholar]
- 10. Nielsen JF (1996) Logarithmic distribution of amplitudes of compound muscle action potentials evoked by transcranial magnetic stimulation. J Clin Neurophysiol 13: 423–434. [DOI] [PubMed] [Google Scholar]
- 11. Brasil-Neto JP, Cohen LG, Panizza M, Nilsson J, Roth BJ, et al. (1992) Optimal focal transcranial magnetic activation of the human motor cortex: effects of coil orientation, shape of the induced current pulse, and stimulus intensity. J Clin Neurophysiol 9: 132–136. [PubMed] [Google Scholar]
- 12. Meyer BU, Britton TC, Kloten H, Steinmetz H, Benecke R (1991) Coil placement in magnetic brain stimulation related to skull and brain anatomy. Electroencephalogr Clin Neurophysiol 81: 38–46. [DOI] [PubMed] [Google Scholar]
- 13.Lewis GN, Signal N, Taylor D (2013) Reliability of lower limb motor evoked potentials in stroke and healthy populations: How many responses are needed? Clin Neurophysiol. [DOI] [PubMed]
- 14. Levin O, Cuypers K, Netz Y, Thijs H, Nuttin B, et al. (2011) Age-related differences in human corticospinal excitability during simple reaction time. Neurosci Lett 487: 53–57. [DOI] [PubMed] [Google Scholar]
- 15.Mantovani A, Rossi S, Bassi BD, Simpson HB, Fallon BA, et al.. (2013) Modulation of motor cortex excitability in obsessive-compulsive disorder: An exploratory study on the relations of neurophysiology measures with clinical outcome. Psychiatry Res. [DOI] [PMC free article] [PubMed]
- 16. Veniero D, Bortoletto M, Miniussi C (2012) Cortical modulation of short-latency TMS-evoked potentials. Front Hum Neurosci 6: 352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Fujiyama H, Hinder MR, Schmidt MW, Tandonnet C, Garry MI, et al. (2012) Age-related differences in corticomotor excitability and inhibitory processes during a visuomotor RT task. J Cogn Neurosci 24: 1253–1263. [DOI] [PubMed] [Google Scholar]
- 18. Meesen RL, Cuypers K, Rothwell JC, Swinnen SP, Levin O (2011) The effect of long-term TENS on persistent neuroplastic changes in the human cerebral cortex. Hum Brain Mapp 32: 872–882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Vaalto S, Saisanen L, Kononen M, Julkunen P, Hukkanen T, et al. (2011) Corticospinal output and cortical excitation-inhibition balance in distal hand muscle representations in nonprimary motor area. Hum Brain Mapp 32: 1692–1703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Smith MJ, Adams LF, Schmidt PJ, Rubinow DR, Wassermann EM (2002) Effects of ovarian hormones on human cortical excitability. Ann Neurol 51: 599–603. [DOI] [PubMed] [Google Scholar]
- 21. Wassermann EM (2002) Variation in the response to transcranial magnetic brain stimulation in the general population. Clin Neurophysiol 113: 1165–1171. [DOI] [PubMed] [Google Scholar]
- 22. Smith MJ, Keel JC, Greenberg BD, Adams LF, Schmidt PJ, et al. (1999) Menstrual cycle effects on cortical excitability. Neurology 53: 2069–2072. [DOI] [PubMed] [Google Scholar]
- 23. Oldfield RC (1971) The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9: 97–113. [DOI] [PubMed] [Google Scholar]
- 24. Wassermann EM (1998) Risk and safety of repetitive transcranial magnetic stimulation: report and suggested guidelines from the International Workshop on the Safety of Repetitive Transcranial Magnetic Stimulation, June 5–7, 1996. Electroencephalogr Clin Neurophysiol 108: 1–16. [DOI] [PubMed] [Google Scholar]
- 25. Liang KY, Zeger SL (1986) Longitudinal data analysis using generalized linear models. Biometrika 73: 13–22. [Google Scholar]
- 26. Zeger SL, Liang KY (1986) Longitudinal data analysis for discrete and continuous outcomes. Biometrics 42: 121–130. [PubMed] [Google Scholar]
- 27. Christie A, Fling B, Crews RT, Mulwitz LA, Kamen G (2007) Reliability of motor-evoked potentials in the ADM muscle of older adults. J Neurosci Methods 164: 320–324. [DOI] [PubMed] [Google Scholar]
- 28.Cuypers K, Leenus DJ, Van Wijmeersch B, Thijs H, Levin O, et al.. (2013) Anodal tDCS increases corticospinal output and projection strength in multiple sclerosis. Neurosci Lett. [DOI] [PubMed]
- 29. Nitsche MA, Seeber A, Frommann K, Klein CC, Rochford C, et al. (2005) Modulating parameters of excitability during and after transcranial direct current stimulation of the human motor cortex. J Physiol 568: 291–303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Smith AE, Sale MV, Higgins RD, Wittert GA, Pitcher JB (2011) Male human motor cortex stimulus-response characteristics are not altered by aging. J Appl Physiol 110: 206–212. [DOI] [PubMed] [Google Scholar]
- 31. Kokate TG, Svensson BE, Rogawski MA (1994) Anticonvulsant activity of neurosteroids: correlation with gamma-aminobutyric acid-evoked chloride current potentiation. J Pharmacol Exp Ther 270: 1223–1229. [PubMed] [Google Scholar]
- 32. Lan NC, Gee KW (1994) Neuroactive steroid actions at the GABAA receptor. Horm Behav 28: 537–544. [DOI] [PubMed] [Google Scholar]
- 33. Sahota P, Prabhakar S, Lal V, Khurana D, Das CP, et al. (2005) Transcranial magnetic stimulation: role in the evaluation of disability in multiple sclerosis. Neurol India 53: 197–201 discussion 201. [DOI] [PubMed] [Google Scholar]
- 34. Pennisi G, Alagona G, Rapisarda G, Nicoletti F, Costanzo E, et al. (2002) Transcranial magnetic stimulation after pure motor stroke. Clin Neurophysiol 113: 1536–1543. [DOI] [PubMed] [Google Scholar]
- 35. Ferreri F, Pauri F, Pasqualetti P, Fini R, Dal Forno G, et al. (2003) Motor cortex excitability in Alzheimer's disease: a transcranial magnetic stimulation study. Ann Neurol 53: 102–108. [DOI] [PubMed] [Google Scholar]
- 36. Julkunen P, Saisanen L, Danner N, Niskanen E, Hukkanen T, et al. (2009) Comparison of navigated and non-navigated transcranial magnetic stimulation for motor cortex mapping, motor threshold and motor evoked potentials. Neuroimage 44: 790–795. [DOI] [PubMed] [Google Scholar]
- 37. Jung NH, Delvendahl I, Kuhnke NG, Hauschke D, Stolle S, et al. (2010) Navigated transcranial magnetic stimulation does not decrease the variability of motor-evoked potentials. Brain Stimul 3: 87–94. [DOI] [PubMed] [Google Scholar]
- 38. Magistris MR, Rosler KM, Truffert A, Landis T, Hess CW (1999) A clinical study of motor evoked potentials using a triple stimulation technique. Brain 122 (Pt 2): 265–279. [DOI] [PubMed] [Google Scholar]
- 39. Grupe DW, Nitschke JB (2013) Uncertainty and anticipation in anxiety: an integrated neurobiological and psychological perspective. Nat Rev Neurosci 14: 488–501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Hess CW, Mills KR, Murray NM (1987) Responses in small hand muscles from magnetic stimulation of the human brain. J Physiol 388: 397–419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Buccolieri A, Abbruzzese G, Rothwell JC (2004) Relaxation from a voluntary contraction is preceded by increased excitability of motor cortical inhibitory circuits. J Physiol 558: 685–695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Taylor JL, Gandevia SC (2001) Transcranial magnetic stimulation and human muscle fatigue. Muscle Nerve 24: 18–29. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Probability table. The number of consecutive stimuli required as a function of the probability of hitting the 95% confidence interval (CI).
(DOCX)
Generalized estimating equation (GEE) analysis. Estimates and p-values are shown for the number of consecutive stimuli, arousal, fatigue, resting motor threshold and the interaction between arousal and fatigue. P-values in bold highlight a significant effect.
(DOCX)
The number of TMS stimuli required to reach a probability of 1.0 for hitting the 95% CI was estimated using the GEE analysis for different levels of resting motor threshold (rMT).
(DOCX)