

Abstract
Notch signalling acts in virtually every tissue during the lifetime of metazoans. Recent studies have pointed to multiple roles for Notch in stem cells during quiescence, proliferation, temporal specification, and maintenance of the niche architecture. Skeletal muscle has served as an excellent paradigm to examine these diverse roles as embryonic, foetal, and adult skeletal muscle stem cells have different molecular signatures and functional properties, reflecting their developmental specification during ontology. Notably, Notch signalling has emerged as a major regulator of all muscle stem cells. This review will provide an overview of Notch signalling during myogenic development and postnatally, and underscore the seemingly opposing contextual activities of Notch that have lead to a reassessment of its role in myogenesis.
Keywords: Notch, Skeletal muscle, Quiescence, Regeneration
Background
Notch signalling is one the major regulatory pathways that define multicellular development, principally by dictating the fate of one cell in relation to that of its neighbouring cell [1]. This pathway relies on cell-to-cell communication, by virtue of the fact that both the Notch receptors (Notch1-4 in mammals) and their ligands (Delta-like1, Delta-like4, Jagged1, and Jagged2 in mammals) are transmembrane proteins (note: Dll3 is not considered, as it does not act as a Notch ligand, [2,3]). The cleaved, intracellular domain of Notch, NICD, is also a transcriptional coactivator [4-6] that interacts with RBPJ [7-10] and induces its binding to DNA to regulate gene expression [11,12].
Although the fundamental role of Notch signalling during development has been well studied, its function in the adult organism was overshadowed until the stem cells of fully-grown and aged organisms became a subject of systematic research. Today, we know that Notch activity is critical for the maintenance of diverse types of adult somatic stem cells, both quiescent and proliferating [13-17]. In the skeletal muscle, adult stem (satellite) cells that are quiescent during homeostasis, have robust regenerative capacity and are regulated by Notch signalling activities.
The origin of satellite cells in the body can be traced back to mesodermal cells of the dermomyotome [18], a transient epithelial structure of the somites formed in the mouse around embryonic day (E)9. These skeletal muscle founder stem cells proliferate and express the paired-box/homeodomain transcription factors Pax3 and Pax7 [19-21]. They undergo myogenic specification following the sequential expression of the bHLH Myogenic Regulatory Factors (MRFs), Myf5, Mrf4, Myod, and subsequently the differentiation gene Myogenin [22-24]. Juvenile satellite cells that are Pax7+ are observed under the myofibre basement membrane from E16.5 [20] and they act as reservoirs of adult satellite cells that emerge between 2–3 weeks postnatally [22,25,26].
The main focus of this review is to examine the role of the Notch pathway during the establishment, maintenance, and activation of satellite cells in the mouse. We speculate on the cellular contexts, during mouse development and postnatally, that ensure proper activation of Notch signalling in skeletal muscle stem cells. Moreover, we underscore the opposing contextual outcomes of Notch activity and propose alternative views for its role during quiescence and regeneration.
Notch receptors and their ligands in myogenic stem cells during development
Notch signalling plays a central role in maintaining muscle stem cells during prenatal and postnatal skeletal muscle development. Therefore, there is a requirement for continuous Notch signalling during the ontogeny of muscle, as well as during adult muscle homeostasis. So, what is the identity and the source of the ligand required to maintain appropriate Notch signalling in the right cells at the right time?
In the embryo, the major player appears to be Delta-like1 (Dll1), provided mainly by the committed myoblasts [13,27-29]. These cells are Myod+ that, in analogy to neuronal bHLH factors [30], induces the expression of Dll1 [31] to signal to upstream cells and sustain the stem/progenitor cell population by direct cell-cell interaction. This feedback mechanism of receptor/ligand regulation, utilized in many other cell systems [32,33], is coherent with the transcriptional signature of upstream (high levels of Pax7, “Pax7Hi”) and differentiating (Pax7Low) myogenic cells that show high levels of Notch receptors (Notch-1, -2,-3) and activity in the former, and high Dll1 ligand expression in the latter [13]. Notably, the Pax7 population in the mouse does not express detectable levels of Dll4, and the transcripts of the Notch ligand and target Jagged-1 are detectable only in the Pax7Hi cells that receive Notch signalling (PM, ST, unpublished observations).
The prevalence of Dll1 in committed myogenic cells is consistent with the muscle phenotypes in mouse embryos with reduced levels of this ligand (hypomorphic over null Dll1 allele), where there is severe muscle hypotrophy due to precocious differentiation of the muscle stem cell population [29]. With the caveat that this was a germline mutation and not cell type specific, this study strongly suggests that Dll1 is a necessary and sufficient Notch ligand for the maintenance of myogenic stem cells during embryogenesis. Similarly, loss of myogenic stem cells was accompanied by increased differentiation in mouse embryos upon muscle-specific, conditional deletion of Rbpj[34]. In a complementary approach, it was shown that Myf5Cre driven expression of constitutively active Notch1 (Myf5Cre:R26stop-NICD) maintained upstream myogenic cells undifferentiated and Pax7+, even in the absence of differentiating progeny [35]. By extrapolation, we can conclude that Dll1-triggered canonical Notch signalling fulfils the requirements as the principal pathway, sufficient to autonomously maintain embryonic myogenic cells throughout development.
A distinct source of Dll1 ligand was described for a subpopulation of muscle progenitors during early myogenesis in the chick embryo [36]. In that study, migrating neural crest cells expressing Dll1 trigger the transient activation of Notch signalling in muscle progenitors specifically within the dorsal dermomyotome of the somite, as these delaminate to commit to myogenesis. This attractive regulatory mechanism, which involves the coordination of different cell types, remains to be tested in diverse model organisms and in other muscle progenitors outside the dorsomedial lip of the dermomyotome.
Notch receptors and their ligands in satellite cells
In adult homeostatic muscle, quiescent satellite cells express Notch -1, -2, and -3 as well as high levels of the Notch/Rbpj targets Hey1, HeyL, and Hes1, thereby reflecting high Notch activity. Abrogation of Notch signalling by targeted deletion of Rbpj results in the spontaneous differentiation of this cell population. Therefore, satellite cells are sustained in a quiescent state by canonical Notch activity [13,37]. Interestingly, Notch3 germline knock-out mice have a seemingly opposite phenotype, with an abnormally high number of satellite cells and hypertrophic regenerated muscle even after seven rounds of injury, indicating an antagonistic function with the other Notch receptors [38]. Although canonical Notch signalling is transduced by Rbpj, how this transcription factor relays signalling from each of the Notch receptors is a critical question that could unveil further surprises. Conditional deletion of Notch3, as well as that of Notch-1 and -2, would provide useful information for the functional relationship of the Notch paralogues in satellite cell homeostasis.
Based on the anatomical position of adult satellite cells between the myofibre and the basement membrane, the muscle fibre is the most likely source of ligand. However, the lack of reliable mouse Dll1 antibodies has hindered the direct visualization of the protein, especially relative to the position of the satellite cells. Genetic, inducible depletion of Dll1 and/or Dll4 specifically in the myofibres should be performed to validate the main source of the ligand. The basal lamina of the basement membrane, a cell-free extracellular matrix protein rich structure, is located in apposition to the myofibre. Although proteoglycans of the basal lamina bind secreted cytokines and other signalling molecules [39], they are not expected to bind Notch ligands as these are transmembrane proteins and their soluble form is not active [40,41].
Alternatively, several cell types that reside outside the basement membrane, including pericytes, endothelial cells, PICs (Pw1+ interstitial cells), fibro-adipogenic and mesenchymal cells, could potentially act as source of ligand [42]. In addition, satellite cells have been shown to be closely associated to capillaries of human and mouse muscle [43]. Though no apparent physical contact has been demonstrated yet between satellite and endothelial or pericyte cells, the latter cell types might contribute to Notch activation in the satellite cells. Indeed, both in vertebrates and invertebrates Dll-bearing cellular protrusions (filopodia) capable of activating Notch signalling at a long-range have been described [44-46], providing a possible mode of cell interactions crossing the basement membrane. Moreover, even in the absence of cell contact, soluble factors secreted by interstitial cells might enhance stimulation of the Notch pathway in satellite cells by a paracrine mechanism.
The muscle fibre, its ensheathing basement membrane, as well as the various cell types indicated above provide a complex microenvironment that maintains satellite cells in a G0 reversible cell cycle state, whilst retaining their extraordinary regenerative potential. Disruption of this satellite cell niche invariably leads to exit from quiescence and entry into a phase of active proliferation. The status of Notch signalling in satellite cells during the transition from G0-exit to the proliferation of myogenic progeny cells has been difficult to decipher. Recent studies point to a more complex role than previously anticipated.
Proliferating and Quiescent myogenic stem cells: two distinct cell states regulated by Notch
In the mouse embryo, Notch signalling is essential for the maintenance of proliferating Pax7+ cells, although it appears not to be required for their emergence in the dermomyotome [29,34]. In the adult, active Notch signalling is critical for sustaining quiescent satellite cells during homeostasis [13,37]. In addition to maintaining quiescence, Notch activity is likely to facilitate the transition into that cell state, as forced expression of NICD promotes precocious cell cycle exit prenatally, characterised by the quiescence marker signature: Pax7+, CalR+ (CalR: Calcitonin receptor, marker of adult satellite cells), Myod— [35]. To date, it is unclear how Notch maintains stem cells in a proliferative state during development and in a quiescent state in the adult. One common feature is by preventing differentiation thereby raising the question whether the quiescent state reflects an absence of proliferation and differentiation or a bona fide defined cell state.
Notch signalling collapses during satellite cell activation: a prerequisite or consequence?
In the context of skeletal muscle repair, previous reports suggested that satellite cell activation and subsequent proliferation is driven by Notch signalling [47-49]. This notion was in agreement with the role of this signalling pathway during development. More recent studies, however, have fundamentally changed this view by examining the activity of the Notch pathway directly in myogenic cells in vivo. Here we highlight the recent reports that point to a more nuanced role for Notch during adult myogenesis. We will first describe the key steps that characterise satellite cell activation, then discuss the role of Notch signalling during these critical stages.
One feature of G0-exit is the prolonged G1 phase that follows the breaking of quiescence. This is indeed the case for satellite cells: in injured muscles, the first S-phase of activated satellite cells in vivo initiates between 14 h and 18 h post-injury (Notexin-injury), [50]. Accordingly, the first mitoses of quiescent satellite cells isolated by FACS are observed between 18-27 h after plating in low oxygen [13,50]. To date, the earliest molecular marker of activated satellite cells is phosphorylated p38 (pp38α/β), followed by Myod [51-53]. Also, Myf5 is highly upregulated upon activation [54,55], but unlike Myod, Myf5 protein is detectable in the majority of quiescent satellite cells [55]. Interestingly, during the G1-phase following quiescence, Myod does not promote differentiation, but instead it directly regulates the expression of Cdc6, a gene involved in rendering chromatin accessible for DNA replication [53,56]. Once activated, myoblasts enter the cell cycle and continue to divide every 7-8 h (both in vivo and in culture in low O2; [50]) until the majority progress to terminal differentiation by downregulating Pax7 and upregulating Myogenin. A fraction of the proliferating population, however, is able to self-renew and return to quiescence (Pax7+/Myod-).
It was previously proposed that Notch signalling is the driving force for the proliferation of activated satellite cells, and this notion was consistent with the general pro-mitotic action of NICD in other systems [57,58]. However, recent experimental evidence has revised our view of the role of Notch signalling during activation. These are based on: 1) the drastic decrease of endogenous Notch activity in vivo following muscle injury; 2) the continued proliferation and efficient regeneration of Rbpj null satellite cells; 3) the hindered proliferation of activated satellite cells that express constitutively active NICD.
Notably, analysis of activated myoblasts isolated from regenerating muscles showed that endogenous Notch signalling is dramatically reduced upon activation, relative to the non-activated satellite cells [13,37]. In both studies, activated satellite cells from hindlimb muscles injured either by Notexin venom [13] or BaCl2 salt [37] were analysed by quantitative RT-PCR. Regardless of the injury model, the expression of the established Notch/Rbpj targets Hey1, Hey2, HeyL Hes5, and Hes1 was reduced by up to 80-90% compared to quiescent satellite cells. Of note, only one Hes family member, Hes6, was found to be upregulated. However, Hes6 is not a direct Notch/Rbpj target, and it promotes myogenic differentiation [59]. Importantly, microarray data comparing quiescent with activated satellite cells (isolated by FACS) in vivo, confirmed that Notch signalling was dramatically reduced in proliferating myoblasts [60]. Consistently, analysis of GFP+ cells from injured transgenicTg:Pax7-nGFP mice showed that the reduction in Notch activity takes place almost immediately (within 20 hours) after injury [13], that is, before the first cell division occurs in vivo [50]. Subsequently, Notch signals weakly in the proliferating myoblasts and it starts to increase again by day 4–5 post-injury, a time point that correlates with the appearance of differentiated, Myogenin+ cells in the population and a decline in overall proliferation of the myogenic cells. We propose that the upregulation of Notch during regeneration, reflects the Pax7Hi, self-renewing myogenic cells. Maximal levels of Notch activity are then restored by days 20–30 post-injury, corresponding to tissue homeostasis and re-establishment of self-renewed satellite cells. Therefore, the majority of myogenic cells exhibit little to no Notch activity immediately following satellite cell activation at a time when these cells exit quiescence to undergo exponential expansion (Figure 1).
Figure 1.

Notch signalling activity during muscle regeneration. Upper panel: In wild type muscle, quiescent, G0-arrested satellite cells have high Notch activity (purple line), which maintains Pax7 and inhibits Myod (indirectly: dotted line) and Myogenin (directly via Hey1) expression. Immediately after activation, satellite cells downregulate Notch activity and express Myod that is required for appropriate Cdc6 expression and S-phase entry. During the amplification phase high Notch activity is restricted to upstream, Pax7Hi cells that remain undifferentiated and self-renew to replenish the satellite cell pool. Notch activation is triggered by Dll1-bearing differentiating myoblasts. Non-muscle cells, like infiltrating inflammatory cells and fibro/adipogenic progenitors could also trigger or influence Notch activation. The expression of Dll ligands by the mature myofibres is likely, but remains to be demonstrated. Lower panel: Rbpj null satellite cells (no Notch activity: dotted purple line) enter the cell cycle normally and start proliferating. Mutant satellite cells differentiate faster (yellow cells) and fail to self-renew.
It is clear also that during regeneration, activated myoblasts interact with other cell types, including infiltrating inflammatory cells [61,62] and fibro/adipogenic progenitors [63]. These interactions might be productive in nature, at least in part, via Notch signalling. On this point, manipulations of the Notch pathway by injection of inhibitory or activating reagents in regenerating muscle can potentially affect myogenic and non-myogenic cell populations thereby resulting in indirect unrelated phenotypes. Moreover, discrepancies between compounds, or mutations targeting the Notch or Rbpj genes, could expose putative non-canonical functions, both of the receptors and Rbpj.
In accordance with the expression data indicated above, genetic analysis of mice with activated or blocked Notch signalling support the view that elevated Notch signalling is not a requirement and likely it is incompatible with transit amplification of myoblasts. The dispensability of Notch activity during myoblast proliferation is illustrated by the phenotypes of Rbpj null mice in the context of regeneration. When muscles with Rbpj null satellite cells were injured (injury performed 16 days after tamoxifen-induced Rbpj gene deletion to ensure complete clearance of protein), tissue repair was overtly normal with only slightly smaller muscle fibres ([13], and data not shown; Figure 2). Moreover, the first mitosis of cultured Rbpj null satellite cells was similar in timing to control Rbpj+/- cells, as scored by videomicroscopy [13]. Furthermore, Rbpj null cells can amplify in culture for the first 3–4 days, that is, before significant differentiation takes place. In a complementary study, it was shown that quiescent satellite cells expressing constitutively activated Notch1 (Pax7 CreERT2 : R26 stop-NICD ), failed to exit from quiescence, as subsequent proliferation was significantly hindered [64]. Taken together, these data strongly suggest that a temporary decrease in Notch/Rbpj signalling at the time of satellite cell activation is a prerequisite for transit amplification of the myogenic population.
Figure 2.
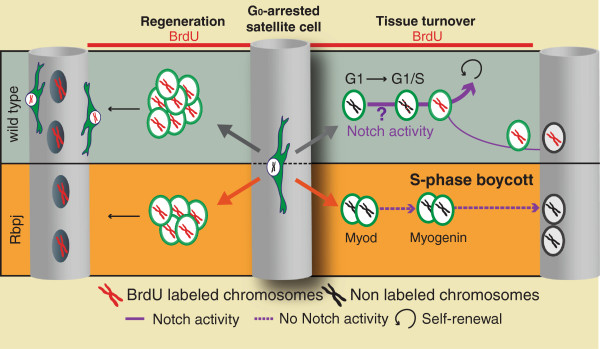
Differential outcomes of loss of Notch activity during regeneration and tissue turnover. During regeneration (left) both wild type and Rbpj null myoblasts proliferate and form new muscle fibres. In the absence of Notch signalling, the size of the amplified population is smaller since the cells fail to self-renew. During tissue homeostasis (right) a small fraction of satellites cells exit quiescence and divide (red chromosomes, BrdU+) to give rise to new satellite cells and fusion-competent myoblasts. Satellite cell specific deletion of Rbpj results in an increase in the number of cells that exit quiescence; these cells differentiate spontaneously. In sharp contrast to wild type cells the majority of Rbpj null satellite cells become Myogenin-positive without going through S-phase (black chromosomes).
A seemingly contradictory observation is that freshly isolated canine satellite cells plated on Dll1 ligand, expanded more than non-treated cells [65]. One major difference between this and the Pax7CreERT2: R26stop-NICD experiment cited above, is that cells were exposed to the ligand after their isolation, hence they were already activated. If Notch downregulation is indeed critical for the Myod-driven G1/S transition, it is possible that some of the dissociated cells were already engaged in the Myod program. Alternatively, chronic Notch signalling in culture might result in the eventual amplification of the cell population after an initial delay. These seemingly disparate observations might expose qualitative differences between the first G1/S transition, and those taking place during myogenic cell amplification.
A model for Notch signalling during early satellite cell activation and homeostasis
In an attempt to rationalize the contextual roles of Notch signalling, we propose that the immediate collapse in activity following injury reflects the physical dissociation of satellite cells from the ligand source (likely the myofibre). The observation that pp38α/β and Myod are early markers of activated satellite cells is also coherent with a downregulation of Notch signalling, as both have been shown to be inhibited by activated Notch [35,51]. Indeed, it is reasonable to expect that Notch signalling should be transiently shut-off in order to allow the expression of Myod, whose activity via Cdc6 is important for exiting quiescence [53] (Figure 1). The increase in Notch activity at day 4–5 post-injury could be the outcome of signal emitting (Dll1+/Myog+) and signal receiving (Pax7Hi) cells, either implicating asymmetric or symmetric cell divisions. Analysis at the single-cell level would be required to investigate if the residual Notch activity scored in proliferating myogenic cells is restricted to a specific subpopulation, for example the self-renewing Pax7Hi cells.
Notably, during homeostasis, loss of Rbpj activity results in depletion of the satellite cell pool after a delayed period of 5–7 weeks [13,37]. In stark contrast to activated satellite cells, the majority of the Rbpj null cells differentiate directly in the absence of muscle injury, without entering S-phase, then they fuse with the myofibres (Figure 2). This observation strongly suggests that in the context of normal satellite cell turnover, Notch activity is required for executing the G1/S transition. Analysis of Notch null clones in the Drosophila eye imaginal disc clearly demonstrates that Notch is required to overcome the G1-S check-point [66], consistent with the observations cited above. The finding that during regeneration Rbpj null satellite cells undergo, seemingly normally, the first G1/S phase following quiescence and they can transiently amplify, points to a secondary pathway that intervenes during injury, but that is absent during homeostasis. Once again, these contrasting behaviours underscore the distinct contextual outcomes of Notch activity in homeostasis vs. injury. Interestingly, in a model where constitutive expression of NICD was maintained in the upstream myogenic population (Myf5Cre-NICD), proliferating myogenic cells proceeded during development to express embryonic and foetal specific markers, then they prematurely exited the cell cycle to adopt a quiescent state late during foetal development. Here too, there are seemingly opposing behaviours for NICD: compatible with proliferating myogenic cells during early embryogenesis, and with the quiescent state in the late foetal phase [35]. These findings lead us to propose that Notch plays a role as facilitator, rather than an instigator, of cell fate decisions in this scenario, leaving the door open to explore alternative pathways that could promote stem cell quiescence.
Towards understanding what lies downstream of Notch signalling
An important initial step towards understanding the developmental outputs of Notch signalling activation is to identify its downstream targets and the degree of conservation under diverse cellular contexts. In an attempt to address this question, a whole-genome ChIP-seq screen was performed for direct Rbpj/NICD targets in cultured myogenic cells, under activated or inhibited Notch signalling [11]. From this study, two points are pertinent to consider. First, in mouse muscle cells, binding of Rbpj to DNA is regulated dynamically by Notch signalling, thereby extending across-species, and at genome-wide scale, the binding behaviour of Drosophila Rbpj to the E(spl) gene cluster [12]. In other words, Rbpj is poorly bound, or not at all, to DNA in the absence of Notch signalling. By contrast, activation of Notch signalling results in recruitment of Rbpj together with NICD to enhancers/promoters, and subsequent activation of transcription. The dynamic association of Rbpj to DNA contradicted the prevalent view that this transcription factor statically occupies its binding sites, while exchanging repressors for activators in response to NICD. Unexpectedly, a second category of binding-sites was identified and these sequences were not co-occupied by NICD, but Rbpj was bound constitutively. Interestingly, this screen uncovered a number of Rbpj/NICD enhancers linked to genes encoding extracellular matrix associated proteins. This raises the intriguing possibility that skeletal muscle stem cells can partially contribute to the composition of their niche via cell-autonomous, Notch-regulated mechanisms. Such a mechanism is in agreement with a recent report that Notch signals control the homing of satellite cells by influencing the proper assembly of components in the basal lamina [67]. Moreover, myoblasts expressing constitutive activated Notch1 (Myf5Cre:R26stop-NICD), survive throughout development in the absence of myofibres, and in this scenario, cells leave the cell cycle in late foetal stages and are surrounded by laminin, possibly mimicking the niche [35].
Review and conclusion
As often reported in the past, the outcome of Notch signals is profoundly dependent on the cellular context [68,69]. Muscle stem cells are no exception; pathway activity is critical for maintaining proliferating stem cells during growth, but also adult satellite cells, which are G0-arrested. Generally, Notch action is pro-myogenic when it is linked to inhibition of differentiation, but it can also suppress growth in tissues where its physiological role is to induce cell differentiation, for example in the skin [70,71]. The quiescent state, however, constitutes an alternative, more complex paradigm, where block of differentiation and reversible cell cycle arrest simultaneously co-exist. Once again, distinct extrinsic and intrinsic factors seem to determine the phenotypic consequences of loss of Rbpj function in satellite and adult neural stem cells, where the former differentiate without entering S-phase during homeostasis [13] and the latter collectively enter the cell cycle and undergo transit amplification [14,15]. A major challenge in the future is to identify the partners and the tissue specific downstream targets of Notch signalling that influence the developmental outcome of this fundamental signalling pathway, and to elucidate the unique and common outcomes of signalling through distinct Notch receptors in the context of Rbpj function.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
PM and ST conceived the study and drafted the manuscript. Both authors read and approved the final manuscript.
Contributor Information
Philippos Mourikis, Email: philipmour@gmail.com.
Shahragim Tajbakhsh, Email: shahragim.tajbakhsh@pasteur.fr.
Acknowledgements
We thank members of the laboratory for critical comments and acknowledge support from the Institut Pasteur, Association Française contre le Myopathies, Agence Nationale de la Recherche (Laboratoire d’Excellence Revive, Investissement d’Avenir; ANR-10-LABX-73), EU Framework 7 projects EuroSystem, Optistem, NotchIT, the Association pour la Recherche sur le Cancer, and the Fondation pour la Recherche Medicale.
References
- Louvi A, Artavanis-Tsakonas S. Notch and disease: a growing field. Semin Cell Dev Biol. 2012;23(4):473–480. doi: 10.1016/j.semcdb.2012.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geffers I, Serth K, Chapman G, Jaekel R, Schuster-Gossler K, Cordes R, Sparrow DB, Kremmer E, Dunwoodie SL, Klein T. et al. Divergent functions and distinct localization of the Notch ligands DLL1 and DLL3 in vivo. J Cell Biol. 2007;178(3):465–476. doi: 10.1083/jcb.200702009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ladi E, Nichols JT, Ge W, Miyamoto A, Yao C, Yang LT, Boulter J, Sun YE, Kintner C, Weinmaster G. The divergent DSL ligand Dll3 does not activate Notch signaling but cell autonomously attenuates signaling induced by other DSL ligands. J Cell Biol. 2005;170(6):983–992. doi: 10.1083/jcb.200503113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wharton KA, Johansen KM, Xu T, Artavanis-Tsakonas S. Nucleotide sequence from the neurogenic locus notch implies a gene product that shares homology with proteins containing EGF-like repeats. Cell. 1985;43(3 Pt 2):567–581. doi: 10.1016/0092-8674(85)90229-6. [DOI] [PubMed] [Google Scholar]
- Schroeter EH, Kisslinger JA, Kopan R. Notch-1 signalling requires ligand-induced proteolytic release of intracellular domain. Nature. 1998;393(6683):382–386. doi: 10.1038/30756. [DOI] [PubMed] [Google Scholar]
- Greenwald I. lin-12, a nematode homeotic gene, is homologous to a set of mammalian proteins that includes epidermal growth factor. Cell. 1985;43(3 Pt 2):583–590. doi: 10.1016/0092-8674(85)90230-2. [DOI] [PubMed] [Google Scholar]
- Jarriault S, Brou C, Logeat F, Schroeter EH, Kopan R, Israel A. Signalling downstream of activated mammalian Notch. Nature. 1995;377(6547):355–358. doi: 10.1038/377355a0. [DOI] [PubMed] [Google Scholar]
- Kopan R, Ilagan MX. The canonical Notch signaling pathway: unfolding the activation mechanism. Cell. 2009;137(2):216–233. doi: 10.1016/j.cell.2009.03.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fortini ME, Artavanis-Tsakonas S. The suppressor of hairless protein participates in notch receptor signaling. Cell. 1994;79(2):273–282. doi: 10.1016/0092-8674(94)90196-1. [DOI] [PubMed] [Google Scholar]
- Schweisguth F, Posakony JW. Suppressor of Hairless, the Drosophila homolog of the mouse recombination signal-binding protein gene, controls sensory organ cell fates. Cell. 1992;69(7):1199–1212. doi: 10.1016/0092-8674(92)90641-O. [DOI] [PubMed] [Google Scholar]
- Castel D, Mourikis P, Bartels SJ, Brinkman AB, Tajbakhsh S, Stunnenberg HG. Dynamic binding of RBPJ is determined by Notch signaling status. Genes Dev. 2013;27(9):1059–1071. doi: 10.1101/gad.211912.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krejci A, Bray S. Notch activation stimulates transient and selective binding of Su(H)/CSL to target enhancers. Genes Dev. 2007;21(11):1322–1327. doi: 10.1101/gad.424607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mourikis P, Sambasivan R, Castel D, Rocheteau P, Bizzarro V, Tajbakhsh S. A critical requirement for notch signaling in maintenance of the quiescent skeletal muscle stem cell state. Stem Cells. 2012;30(2):243–252. doi: 10.1002/stem.775. [DOI] [PubMed] [Google Scholar]
- Chapouton P, Skupien P, Hesl B, Coolen M, Moore JC, Madelaine R, Kremmer E, Faus-Kessler T, Blader P, Lawson ND. et al. Notch activity levels control the balance between quiescence and recruitment of adult neural stem cells. J Neurosci. 2010;30(23):7961–7974. doi: 10.1523/JNEUROSCI.6170-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imayoshi I, Sakamoto M, Yamaguchi M, Mori K, Kageyama R. Essential roles of Notch signaling in maintenance of neural stem cells in developing and adult brains. J Neurosci. 2010;30(9):3489–3498. doi: 10.1523/JNEUROSCI.4987-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch U, Lehal R, Radtke F. Stem cells living with a Notch. Development. 2013;140(4):689–704. doi: 10.1242/dev.080614. [DOI] [PubMed] [Google Scholar]
- Pellegrinet L, Rodilla V, Liu Z, Chen S, Koch U, Espinosa L, Kaestner KH, Kopan R, Lewis J, Radtke F. Dll1- and dll4-mediated notch signaling are required for homeostasis of intestinal stem cells. Gastroenterology. 2011;140(4):1230–1240. doi: 10.1053/j.gastro.2011.01.005. e1231-1237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gros J, Manceau M, Thome V, Marcelle C. A common somitic origin for embryonic muscle progenitors and satellite cells. Nature. 2005;435(7044):954–958. doi: 10.1038/nature03572. [DOI] [PubMed] [Google Scholar]
- Relaix F, Rocancourt D, Mansouri A, Buckingham M. A Pax3/Pax7-dependent population of skeletal muscle progenitor cells. Nature. 2005;435(7044):948–953. doi: 10.1038/nature03594. [DOI] [PubMed] [Google Scholar]
- Kassar-Duchossoy L, Giacone E, Gayraud-Morel B, Jory A, Gomes D, Tajbakhsh S. Pax3/Pax7 mark a novel population of primitive myogenic cells during development. Genes Dev. 2005;19(12):1426–1431. doi: 10.1101/gad.345505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben-Yair R, Kalcheim C. Lineage analysis of the avian dermomyotome sheet reveals the existence of single cells with both dermal and muscle progenitor fates. Development. 2005;132(4):689–701. doi: 10.1242/dev.01617. [DOI] [PubMed] [Google Scholar]
- Tajbakhsh S. Skeletal muscle stem cells in developmental versus regenerative myogenesis. J Intern Med. 2009;266(4):372–389. doi: 10.1111/j.1365-2796.2009.02158.x. [DOI] [PubMed] [Google Scholar]
- Rudnicki MA, Schnegelsberg PN, Stead RH, Braun T, Arnold HH, Jaenisch R. MyoD or Myf-5 is required for the formation of skeletal muscle. Cell. 1993;75(7):1351–1359. doi: 10.1016/0092-8674(93)90621-V. [DOI] [PubMed] [Google Scholar]
- Kassar-Duchossoy L, Gayraud-Morel B, Gomes D, Rocancourt D, Buckingham M, Shinin V, Tajbakhsh S. Mrf4 determines skeletal muscle identity in Myf5:Myod double-mutant mice. Nature. 2004;431(7007):466–471. doi: 10.1038/nature02876. [DOI] [PubMed] [Google Scholar]
- White RB, Bierinx AS, Gnocchi VF, Zammit PS. Dynamics of muscle fibre growth during postnatal mouse development. BMC Dev Biol. 2010;10:21. doi: 10.1186/1471-213X-10-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lepper C, Conway SJ, Fan CM. Adult satellite cells and embryonic muscle progenitors have distinct genetic requirements. Nature. 2009;460(7255):627–631. doi: 10.1038/nature08209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delfini MC, Hirsinger E, Pourquie O, Duprez D. Delta 1-activated notch inhibits muscle differentiation without affecting Myf5 and Pax3 expression in chick limb myogenesis. Development. 2000;127(23):5213–5224. doi: 10.1242/dev.127.23.5213. [DOI] [PubMed] [Google Scholar]
- Hirsinger E, Malapert P, Dubrulle J, Delfini MC, Duprez D, Henrique D, Ish-Horowicz D, Pourquie O. Notch signalling acts in postmitotic avian myogenic cells to control MyoD activation. Development. 2001;128(1):107–116. doi: 10.1242/dev.128.1.107. [DOI] [PubMed] [Google Scholar]
- Schuster-Gossler K, Cordes R, Gossler A. Premature myogenic differentiation and depletion of progenitor cells cause severe muscle hypotrophy in Delta1 mutants. Proc Natl Acad Sci U S A. 2007;104(2):537–542. doi: 10.1073/pnas.0608281104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castro DS, Skowronska-Krawczyk D, Armant O, Donaldson IJ, Parras C, Hunt C, Critchley JA, Nguyen L, Gossler A, Gottgens B. et al. Proneural bHLH and Brn proteins coregulate a neurogenic program through cooperative binding to a conserved DNA motif. Dev Cell. 2006;11(6):831–844. doi: 10.1016/j.devcel.2006.10.006. [DOI] [PubMed] [Google Scholar]
- Wittenberger T, Steinbach OC, Authaler A, Kopan R, Rupp RA. MyoD stimulates delta-1 transcription and triggers notch signaling in the Xenopus gastrula. EMBO J. 1999;18(7):1915–1922. doi: 10.1093/emboj/18.7.1915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henrique D, Hirsinger E, Adam J, Le Roux I, Pourquie O, Ish-Horowicz D, Lewis J. Maintenance of neuroepithelial progenitor cells by Delta-Notch signalling in the embryonic chick retina. Curr Biol. 1997;7(9):661–670. doi: 10.1016/S0960-9822(06)00293-4. [DOI] [PubMed] [Google Scholar]
- Dong Z, Yang N, Yeo SY, Chitnis A, Guo S. Intralineage directional notch signaling regulates self-renewal and differentiation of asymmetrically dividing radial glia. Neuron. 2012;74(1):65–78. doi: 10.1016/j.neuron.2012.01.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vasyutina E, Lenhard DC, Wende H, Erdmann B, Epstein JA, Birchmeier C. RBP-J (Rbpsuh) is essential to maintain muscle progenitor cells and to generate satellite cells. Proc Natl Acad Sci U S A. 2007;104(11):4443–4448. doi: 10.1073/pnas.0610647104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mourikis P, Gopalakrishnan S, Sambasivan R, Tajbakhsh S. Cell-autonomous Notch activity maintains the temporal specification potential of skeletal muscle stem cells. Development. 2012;139(24):4536–4548. doi: 10.1242/dev.084756. [DOI] [PubMed] [Google Scholar]
- Rios AC, Serralbo O, Salgado D, Marcelle C. Neural crest regulates myogenesis through the transient activation of NOTCH. Nature. 2011;473(7348):532–535. doi: 10.1038/nature09970. [DOI] [PubMed] [Google Scholar]
- Bjornson CR, Cheung TH, Liu L, Tripathi PV, Steeper KM, Rando TA. Notch signaling is necessary to maintain quiescence in adult muscle stem cells. Stem Cells. 2012;30(2):232–242. doi: 10.1002/stem.773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kitamoto T, Hanaoka K. Notch3 null mutation in mice causes muscle hyperplasia by repetitive muscle regeneration. Stem Cells. 2010;28(12):2205–2216. doi: 10.1002/stem.547. [DOI] [PubMed] [Google Scholar]
- Schonherr E, Hausser HJ. Extracellular matrix and cytokines: a functional unit. Dev Immunol. 2000;7(2–4):89–101. doi: 10.1155/2000/31748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varnum-Finney B, Wu L, Yu M, Brashem-Stein C, Staats S, Flowers D, Griffin JD, Bernstein ID. Immobilization of Notch ligand, Delta-1, is required for induction of notch signaling. J Cell Sci. 2000;113(Pt 23):4313–4318. doi: 10.1242/jcs.113.23.4313. [DOI] [PubMed] [Google Scholar]
- Mishra-Gorur K, Rand MD, Perez-Villamil B, Artavanis-Tsakonas S. Down-regulation of Delta by proteolytic processing. J Cell Biol. 2002;159(2):313–324. doi: 10.1083/jcb.200203117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pannerec A, Marazzi G, Sassoon D. Stem cells in the hood: the skeletal muscle niche. Trends Mol Med. 2012;18(10):599–606. doi: 10.1016/j.molmed.2012.07.004. [DOI] [PubMed] [Google Scholar]
- Christov C, Chretien F, Abou-Khalil R, Bassez G, Vallet G, Authier FJ, Bassaglia Y, Shinin V, Tajbakhsh S, Chazaud B. et al. Muscle satellite cells and endothelial cells: close neighbors and privileged partners. Mol Biol Cell. 2007;18(4):1397–1409. doi: 10.1091/mbc.E06-08-0693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Joussineau C, Soule J, Martin M, Anguille C, Montcourrier P, Alexandre D. Delta-promoted filopodia mediate long-range lateral inhibition in Drosophila. Nature. 2003;426(6966):555–559. doi: 10.1038/nature02157. [DOI] [PubMed] [Google Scholar]
- Cohen M, Georgiou M, Stevenson NL, Miodownik M, Baum B. Dynamic filopodia transmit intermittent Delta-Notch signaling to drive pattern refinement during lateral inhibition. Dev Cell. 2010;19(1):78–89. doi: 10.1016/j.devcel.2010.06.006. [DOI] [PubMed] [Google Scholar]
- Nelson BR, Hodge RD, Bedogni F, Hevner RF. Dynamic interactions between intermediate neurogenic progenitors and radial glia in embryonic mouse neocortex: potential role in Dll1-Notch signaling. J Neurosci. 2013;33(21):9122–9139. doi: 10.1523/JNEUROSCI.0791-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brack AS, Conboy IM, Conboy MJ, Shen J, Rando TA. A temporal switch from notch to Wnt signaling in muscle stem cells is necessary for normal adult myogenesis. Cell stem cell. 2008;2(1):50–59. doi: 10.1016/j.stem.2007.10.006. [DOI] [PubMed] [Google Scholar]
- Conboy IM, Rando TA. The regulation of Notch signaling controls satellite cell activation and cell fate determination in postnatal myogenesis. Dev Cell. 2002;3(3):397–409. doi: 10.1016/S1534-5807(02)00254-X. [DOI] [PubMed] [Google Scholar]
- Sun H, Li L, Vercherat C, Gulbagci NT, Acharjee S, Li J, Chung TK, Thin TH, Taneja R. Stra13 regulates satellite cell activation by antagonizing Notch signaling. J Cell Biol. 2007;177(4):647–657. doi: 10.1083/jcb.200609007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rocheteau P, Gayraud-Morel B, Siegl-Cachedenier I, Blasco MA, Tajbakhsh S. A subpopulation of adult skeletal muscle stem cells retains all template DNA strands after cell division. Cell. 2012;148(1–2):112–125. doi: 10.1016/j.cell.2011.11.049. [DOI] [PubMed] [Google Scholar]
- Kondoh K, Sunadome K, Nishida E. Notch signaling suppresses p38 MAPK activity via induction of MKP-1 in myogenesis. J Biol Chem. 2007;282(5):3058–3065. doi: 10.1074/jbc.M607630200. [DOI] [PubMed] [Google Scholar]
- Jones NC, Tyner KJ, Nibarger L, Stanley HM, Cornelison DD, Fedorov YV, Olwin BB. The p38alpha/beta MAPK functions as a molecular switch to activate the quiescent satellite cell. J Cell Biol. 2005;169(1):105–116. doi: 10.1083/jcb.200408066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang K, Sha J, Harter ML. Activation of Cdc6 by MyoD is associated with the expansion of quiescent myogenic satellite cells. J Cell Biol. 2010;188(1):39–48. doi: 10.1083/jcb.200904144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crist CG, Montarras D, Buckingham M. Muscle satellite cells are primed for myogenesis but maintain quiescence with sequestration of Myf5 mRNA targeted by microRNA-31 in mRNP granules. Cell Stem Cell. 2012;11(1):118–126. doi: 10.1016/j.stem.2012.03.011. [DOI] [PubMed] [Google Scholar]
- Gayraud-Morel B, Chretien F, Jory A, Sambasivan R, Negroni E, Flamant P, Soubigou G, Coppee JY, Di Santo J, Cumano A. et al. Myf5 haploinsufficiency reveals distinct cell fate potentials for adult skeletal muscle stem cells. J Cell Sci. 2012;125(Pt 7):1738–1749. doi: 10.1242/jcs.097006. [DOI] [PubMed] [Google Scholar]
- Machida YJ, Hamlin JL, Dutta A. Right place, right time, and only once: replication initiation in metazoans. Cell. 2005;123(1):13–24. doi: 10.1016/j.cell.2005.09.019. [DOI] [PubMed] [Google Scholar]
- Fre S, Huyghe M, Mourikis P, Robine S, Louvard D, Artavanis-Tsakonas S. Notch signals control the fate of immature progenitor cells in the intestine. Nature. 2005;435(7044):964–968. doi: 10.1038/nature03589. [DOI] [PubMed] [Google Scholar]
- Radtke F, Raj K. The role of Notch in tumorigenesis: oncogene or tumour suppressor? Nat Rev Cancer. 2003;3(10):756–767. doi: 10.1038/nrc1186. [DOI] [PubMed] [Google Scholar]
- Gao X, Chandra T, Gratton MO, Quelo I, Prud'homme J, Stifani S, St-Arnaud R. HES6 acts as a transcriptional repressor in myoblasts and can induce the myogenic differentiation program. J Cell Biol. 2001;154(6):1161–1171. doi: 10.1083/jcb.200104058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu L, Cheung TH, Charville GW, Hurgo BM, Leavitt T, Shih J, Brunet A, Rando TA. Chromatin modifications as determinants of muscle stem cell quiescence and chronological aging. Cell Rep. 2013;4(1):189–204. doi: 10.1016/j.celrep.2013.05.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnold L, Henry A, Poron F, Baba-Amer Y, van Rooijen N, Plonquet A, Gherardi RK, Chazaud B. Inflammatory monocytes recruited after skeletal muscle injury switch into antiinflammatory macrophages to support myogenesis. J Exp Med. 2007;204(5):1057–1069. doi: 10.1084/jem.20070075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sonnet C, Lafuste P, Arnold L, Brigitte M, Poron F, Authier FJ, Chretien F, Gherardi RK, Chazaud B. Human macrophages rescue myoblasts and myotubes from apoptosis through a set of adhesion molecular systems. J Cell Sci. 2006;119(Pt 12):2497–2507. doi: 10.1242/jcs.02988. [DOI] [PubMed] [Google Scholar]
- Joe AW, Yi L, Natarajan A, Le Grand F, So L, Wang J, Rudnicki MA, Rossi FM. Muscle injury activates resident fibro/adipogenic progenitors that facilitate myogenesis. Nat Cell Biol. 2010;12(2):153–163. doi: 10.1038/ncb2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wen Y, Bi P, Liu W, Asakura A, Keller C, Kuang S. Constitutive Notch activation upregulates Pax7 and promotes the self-renewal of skeletal muscle satellite cells. Mol Cell Biol. 2012;32(12):2300–2311. doi: 10.1128/MCB.06753-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parker MH, Loretz C, Tyler AE, Duddy WJ, Hall JK, Olwin BB, Bernstein ID, Storb R, Tapscott SJ. Activation of Notch signaling during ex vivo expansion maintains donor muscle cell engraftment. Stem Cells. 2012;30(10):2212–2220. doi: 10.1002/stem.1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baonza A, Freeman M. Control of cell proliferation in the Drosophila eye by Notch signaling. Dev Cell. 2005;8(4):529–539. doi: 10.1016/j.devcel.2005.01.019. [DOI] [PubMed] [Google Scholar]
- Brohl D, Vasyutina E, Czajkowski MT, Griger J, Rassek C, Rahn HP, Purfurst B, Wende H, Birchmeier C. Colonization of the satellite cell niche by skeletal muscle progenitor cells depends on Notch signals. Dev Cell. 2012;23(3):469–481. doi: 10.1016/j.devcel.2012.07.014. [DOI] [PubMed] [Google Scholar]
- Hori K, Sen A, Artavanis-Tsakonas S. Notch signaling at a glance. J Cell Sci. 2013;126(Pt 10):2135–2140. doi: 10.1242/jcs.127308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Louvi A, Artavanis-Tsakonas S. Notch and disease: a growing field. Semin Cell Dev Biol. 2012. [DOI] [PMC free article] [PubMed]
- Koch U, Radtke F. Notch and cancer: a double-edged sword. Cell Mol Life Sci. 2007;64(21):2746–2762. doi: 10.1007/s00018-007-7164-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rangarajan A, Talora C, Okuyama R, Nicolas M, Mammucari C, Oh H, Aster JC, Krishna S, Metzger D, Chambon P. et al. Notch signaling is a direct determinant of keratinocyte growth arrest and entry into differentiation. EMBO J. 2001;20(13):3427–3436. doi: 10.1093/emboj/20.13.3427. [DOI] [PMC free article] [PubMed] [Google Scholar]