

Abstract
Abstract The brain is programmed to drive behaviour by precisely wiring the appropriate neuronal circuits. Wiring and rewiring of neuronal circuits largely depends on the orchestrated changes in the strengths of synaptic contacts. Here, we review how the rules of synaptic plasticity change during development of the brain, from birth to independence. We focus on the changes that occur at the postsynaptic side of excitatory glutamatergic synapses in the rodent hippocampus and neocortex. First we summarize the current data on the structure of synapses and the developmental expression patterns of the key molecular players of synaptic plasticity, N-methyl-d-aspartate (NMDA) and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors, as well as pivotal kinases (Ca2+/calmodulin-dependent protein kinase II, protein kinase A, protein kinase C) and phosphatases (PP1, PP2A, PP2B). In the second part we relate these findings to important characteristics of the emerging network. We argue that the concerted and gradual shifts in the usage of plasticity molecules comply with the changing need for (re)wiring neuronal circuits.
 [ Christian Lohmann is Senior Group Leader and Head of the Synapse and Network Development Laboratory at the Netherlands Institute for Neuroscience in Amsterdam. His group studies mechanisms of synapse development and, in particular, the signaling cascades that lead to the establishment of precisely connected networks using imaging and electrophysiology in brain slices and in vivo. After studying Biology in Mainz and Tbingen he did his PhD at the University of Frankfurt with Dr. Eckhard Friauf. Next, he pursued his research as a post-doctoral fellow with Dr. Rachel Wong at Washington University Medical School in St. Louis. In 2002 he started working as a research associate at the Max-Planck Institute of Neurobiology with Dr. Tobias Bonhoeffer and in 2008 he established his group in Amsterdam.
[ Christian Lohmann is Senior Group Leader and Head of the Synapse and Network Development Laboratory at the Netherlands Institute for Neuroscience in Amsterdam. His group studies mechanisms of synapse development and, in particular, the signaling cascades that lead to the establishment of precisely connected networks using imaging and electrophysiology in brain slices and in vivo. After studying Biology in Mainz and Tbingen he did his PhD at the University of Frankfurt with Dr. Eckhard Friauf. Next, he pursued his research as a post-doctoral fellow with Dr. Rachel Wong at Washington University Medical School in St. Louis. In 2002 he started working as a research associate at the Max-Planck Institute of Neurobiology with Dr. Tobias Bonhoeffer and in 2008 he established his group in Amsterdam.
Helmut W. Kessels is a Group Leader at the Netherlands Institute for Neuroscience (NIN) of the Royal Dutch Academy of Sciences, Amsterdam, the Netherlands. He completed his Masters in Molecular Sciences in 1996 from Wageningen University, Wageningen, the Netherlands and received his PhD in Immunology from the Netherlands Cancer Institute, Amsterdam, the Netherlands in 2003. He was a Postdoctoral fellow at the Department of Neuroscience, Erasmus Medical Center, Rotterdam, the Netherlands (Advisor: Prof. Dr. Y. Elgersma) in 2003–2004 and at the Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, USA (Advisor: Prof. Dr. R. Malinow) in 2004–2008.]
Introduction
Our brains enable us to perceive and interact with the outer world. Synaptic connections between neurons can be adjusted to respond flexibly to changes in the environment, for example when we learn. In addition, plasticity mechanisms play important roles during early development, even before we are born, to prepare the neuronal circuitry for processing sensory information when we open our eyes. The demands for synaptic plasticity keep changing during the course of a lifetime. At the first developmental stage of life, neuronal networks are being built in the brain to prepare itself for dealing with the outside world after birth (enabling perceptions and the programming of innate behaviour). To accomplish this, synaptic contacts are being shaped in the absence of sensory input. In the next phase of life, the infant has to absorb and process a great deal of new information in a short time (parents, family, language, cultural behaviour), which demands high levels of synaptic plasticity. At the mature stage the need for synaptic plasticity becomes gradually less urgent: a picture of the outside world is made that only incidentally requires adaptation.
Here, we review how the rules of synaptic plasticity change during development of the brain, from birth to independence. The different phases of synaptic plasticity are extensively studied in the rodent brain. Mice and rats open their eyes approximately 2 weeks after birth, and are considered sexually mature at 2 months of age, suggesting that the different phases of synaptic plasticity take place within that time frame. As an attempt to visualize how the demands for plasticity change in the rodent brain, the expression profiles of a number of key synaptic components during the first weeks of the rodent life were gathered from literature and portrayed in Figs 1 and 2. In the first part of this review we will discuss the plasticity changes during the first weeks of the rodent life from the perspective of each individual synaptic component (Fig. 1). In the second part we will discuss the key synaptic plasticity mechanisms from the perspective of each developmental phase (Fig. 2). We focus on the postsynaptic changes that occur at excitatory glutamatergic synapses in the hippocampus and neocortex in rats and mice. Other important aspects of the development of synaptic physiology and plasticity have been reviewed excellently elsewhere (Huang & Scheiffele, 2008; Kerchner & Nicoll, 2008; Feldmeyer & Radnikow, 2009; Groc et al. 2009; Owald & Sigrist, 2009; Sanes & Kotak, 2011; Clarke et al. 2012).
Figure 1. Time course of various synaptic components.
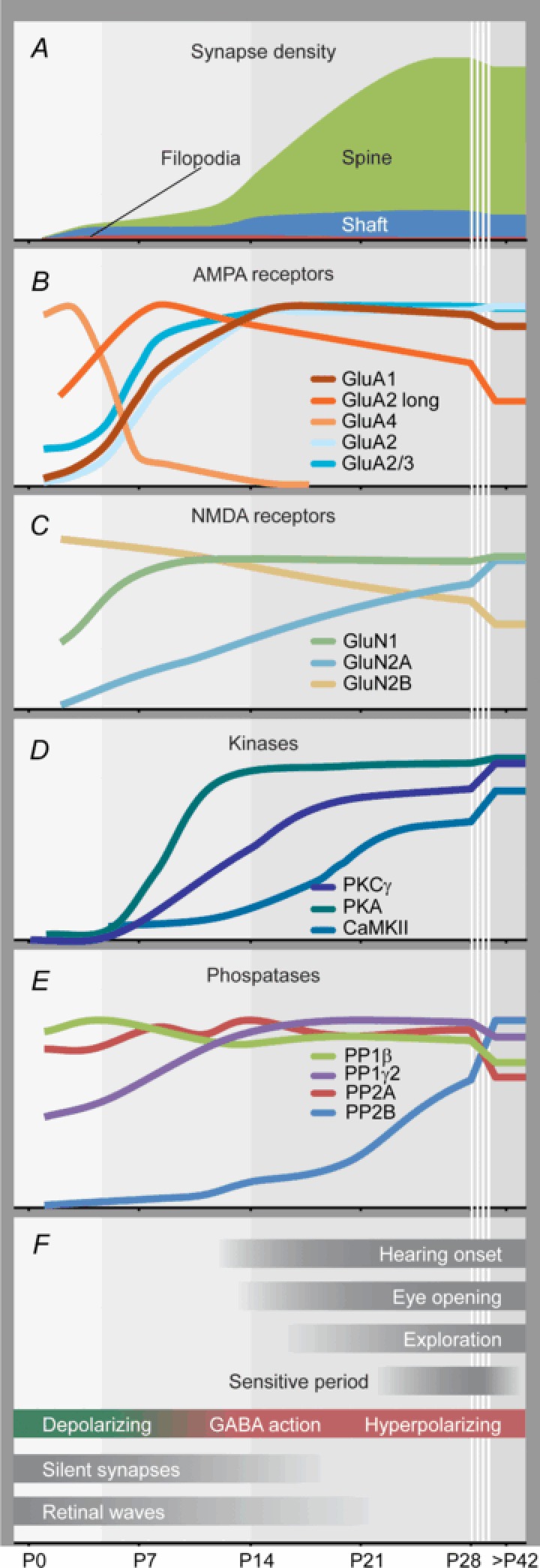
Time course of the postnatal development of synapses (A) and postsynaptic plasticity molecules (B–E) in the hippocampus, forebrain or whole brain in comparison with cellular, network and behavioural development (F). Curves in A–E are expressed as a percentage of the maximal density or expression level. Original data were smoothed (running average). Original data: A, synapse density (Fiala et al. 1998; Steward & Falk, 1991). Proportion of filopodia, shaft and spine synapses (P1–P12: Fiala et al. 1998; P21: Boyer et al. 1998; adult: Harris et al. 1992). B, GluA1, 2, 2/3, 4 (Zhu et al. 2000); GluA2long (Kolleker et al. 2003). C, GluN1, 2A, 2B (Sans et al. 2000). D, PKCγ (Roisin & Barbin, 1997); PKA (Kelly, 1982); CaMKII (Kelly & Vernon, 1985). E, PP1β, PP1γ2 (Strack et al. 1999); PP2A, 2B (Takahashi et al. 2000).
Figure 2. Schematic of the structure and plasticity molecules of glutamatergic synapses at four fundamental stages of development.
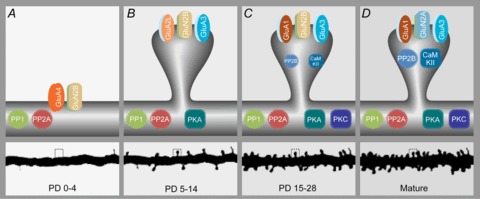
A, between postnatal day 0 and 5 most synapses are located on the dendritic shaft, AMPARs only consist of GluA4 and NMDARs have all GluN2B. Phosphatases PP1 and PP2A are present, but the prime kinases are still absent. B, between postnatal days 5 and 14 synapses are still scarce but start occurring on spines, GluA2long is temporarily the dominant long-tailed AMPAR subunit, while short-tailed AMPAR subunits GluA2/3 also start being expressed. PKA is becoming the dominant kinase. C, in the third and fourth weeks after birth spine numbers increase exponentially. Two types of AMPARs dominate from this time onward: GluA1/2s and GluA2/3s. The kinase PKC is reaching maximal levels. CaMKII and calcineurin (PP2B) expression levels are rising. D, when the rodent is mature, spine numbers maximize, the developmental switch of GluN2B to GluN2A has occurred and CaMKII and calcineurin (PP2B) have reached maximal levels.
Structural and molecular determinants of synaptic plasticity
The ability of synapses to undergo plastic changes of postsynaptic responses is determined by many factors. First, synaptic structure plays an important role, for example the presence or absence of synaptic spines. Spines represent specialized dendritic compartments that isolate synapses biochemically and electrically from each other and may thus help to confine plasticity events to individual synapses. Besides synaptic structure, also the molecular machinery within a synapse defines the malleability of synaptic transmission. The basic molecular components that underlie synaptic transmission are the neurotransmitter receptors that transform chemical signals into changes of membrane polarization. These receptors are also an important substrate of synaptic plasticity, because their numbers and functionality can increase or decrease to potentiate or depress synaptic function. Furthermore, intracellular signalling cascades are required to translate specific activity patterns into changes of synaptic strength. Central to many plasticity mechanisms are kinases and phosphatases that mediate or influence synaptic plasticity. Importantly, these molecular components of the synaptic plasticity machinery change in parallel with the changing demands for neuronal adaptation during development.
Synaptic structure (Fig. 1A)
During the first weeks of development neurons undergo substantial structural alterations: dendrites and axons grow at a fast rate and establish new synaptic contacts. Furthermore, the fine structure of synapses changes dramatically. At birth the number of synapses is low and most are located on dendritic shafts or small protrusions called filopodia (Fiala et al. 1998). In the third week after birth the number of synapses increases sharply (De Felipe et al. 1997; Steward & Falk, 1991) and the great majority of newly generated synapses are located on dendritic spines (Boyer et al. 1998). During the fourth week the generation of synapses and spines slows down and reaches mature levels in the cortex and hippocampus. The absolute number of shaft synapses in the hippocampus does not change much during development after the first postnatal week and therefore only a small minority (<10%) of excitatory synapses are located on shafts in mature hippocampal neurons (Harris et al. 1992; Boyer et al. 1998). The relative number of filopodial synapses also decreases during maturation and is very low in the adult. Although in the mature brain synaptic structures can still be dynamic, for example during learning, overall synaptic efficacies remain stationary, and average spine density and spine size are constant or become slightly reduced (Steward & Falk, 1991; De Felipe et al. 1997; Holtmaat et al. 2005; Loewenstein et al. 2011).
That the developmental increase in spine synapses coincides with a relative decrease in shaft synapses led to the hypothesis that shaft synapses are precursors to spine synapses (Fiala et al. 1998; Ethell & Pasquale, 2005). However, long-term live imaging showed that newly emerging dendritic protrusions occur adjacent to shaft synapses and that new spines turn into functional postsynaptic compartments only after their emergence (Nagerl et al. 2007; Zito et al. 2009; Kwon & Sabatini, 2011). A shaft-to-spiny synapse transition was never observed. In addition, spine growth was shown to precede synapse formation (Knott et al. 2006). Several lines of evidence suggest that shaft synapses and spine synapses represent different types of synapses: they are of different size (the postsynaptic densities of shaft synapses are on average 2-fold larger; Rusakov et al. 1998), they are differentially regulated (Geinisman et al. 1996; Helmeke et al. 2001; Zha et al. 2005; Aoto et al. 2007; Nedelescu et al. 2010), and can receive input from different sources (e.g. in the amygdala; Dong et al. 2010).
There are several advantages for having a synapse located on a spine, and these can be of chemical (Muller & Connor, 1991; Koch et al. 1992; Koch & Zador, 1993; Yuste & Denk, 1995; Sabatini et al. 2002), electrical (Araya et al. 2006a2006a,b2006b; Grunditz et al. 2008; Bloodgood et al. 2009; Yuste, 2011; Gulledge et al. 2012; Harnett et al. 2012) and spatial (Chklovskii, 2004) nature. The influx of cations, and in particular calcium, during synaptic activation is more restricted within the head compartment of a spine. The signal transduction pathways that are activated by the increase in calcium are therefore more local, too, because the likelihood that calcium or second messengers diffuse to neighbouring synapses is low. Secondly, the spine necks form electrical filters that amplify voltage changes in the spine head. Thirdly, the presence of dendritic spines maximizes connectivity within the available volume of tissue.
If there are all these advantages for having synapses located on dendritic spines, why are dendrites in early development lacking them? The requirement of spatial optimization may be less stringent during early development where interstitial space is abundant. Whether there is less need for chemical or electrical isolation of synapses is unknown. Perhaps with shaft synapses there is a higher degree of flexibility: it may be less costly to rearrange shaft synapses than those on spines. It is also possible that chemical or electrical isolation of synapses is less desirable during early development. We speculate that there is a need for neighbouring synapses to be influenced by each other chemically or electrically to facilitate clustering of synchronized inputs (Kleindienst et al. 2011; Takahashi et al. 2012).
AMPA receptor composition (Fig. 1B)
Most synapses use α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA)-type glutamate receptors (AMPARs) for fast transmission of signals across the synapse. The efficacy by which signals are transmitted across the synapse depends on the number of AMPARs at the postsynaptic site. Long-term potentiation (LTP) and long-term depression (LTD) of synaptic strength are expressed by the addition and removal of synaptic AMPARs (Shepherd & Huganir, 2007; Malinow & Malenka, 2002; Bassani et al. 2013), processes that play a role in the rewiring of neuronal circuits and the adaptation of behaviour (Kessels & Malinow, 2009). AMPARs are tetrameric molecules, consisting of four AMPAR subunits. There are four different AMPAR genes, Gria1, Gria2, Gria3 and Gria4, which encode for AMPAR subunits GluA1, GluA2, GluA3 and GluA4 respectively. These AMPAR subunits are most distinctively different in the structure of their cytoplasmic regions. GluA1, GluA4 and a splice variant of GluA2 named GluA2long have long cytoplasmic tails, while the c-tails of GluA2 and GluA3 are short and similarly structured.
Before birth and in the first postnatal week GluA4 is expressed in excitatory neurons of the hippocampus and cortex. Since the levels of the other subunits are low at this time, AMPARs at this early stage probably consist predominantly of GluA4. Entry of GluA4 homomers into synapses is activity dependent and a low level of activity (i.e. spontaneous activity) is sufficient for trafficking of GluA4 into synapses (Zhu et al. 2000). Soon after birth the expression levels of GluA4 decline as a consequence of increased neuronal activity (Akaneya, 2007). When GluA4 levels drop, another developmental AMPAR rises: GluA2long. GluA2long is an alternatively spliced isoform of GluA2 with a long cytoplasmic tail that is similar in structure to that of GluA4. The expression levels of GluA2long suggest that it is specialized in mediating synaptic strengthening during development: its expression peaks in the second week after birth, and is low in the mature hippocampus (Kolleker et al. 2003). The activity patterns that trigger GluA2long insertion into synapses are thought to lie between those of GluA4 and GluA1: both spontaneous activity and LTP-inducing activity patterns mediate synaptic insertion of GluA2long homomers (Kolleker et al. 2003).
In the second postnatal week the expression levels of GluA1, GluA2 and GluA3 start to rise in parallel and reach the high levels seen in adults by the third postnatal week. A unique feature of GluA2 is that its RNA is edited such that it has a positively charged arginine in its transmembrane region (Sommer et al. 1991). As a result, GluA2 homomers are unstable (Greger et al. 2002) and thus GluA2 predominantly forms heteromers with the other subunits (Greger et al. 2003). At the mature stage, three types of AMPARs are formed in excitatory neurons of the hippocampus and cortex: GluA1/2 heteromers, GluA2/3 heteromers and a smaller population of GluA1 homomers (Wenthold et al. 1996). The relative amounts of AMPAR subunits GluA1, GluA2 and GluA3 in hippocampal dendrites are estimated to be at a ratio of 1:2:1 (Kessels et al. 2009), suggesting that GluA1/2 and GluA2/3 are present in equivalent amounts. To determine the AMPAR subunit composition in synapses, synapse strength was measured after deletion of either GluA1 or GluA3 during the first weeks of development. GluA1-deficient neurons were 80% decreased in synaptic AMPAR currents, while currents in GluA3-deficient synapses were reduced by 15%, suggesting that the majority of synaptic AMPARs are GluA1/2s (Lu et al. 2009). However, the expression of an AMPAR subunit may not be the only factor that determines synaptic strength. GluA1-lacking neurons have very few functional AMPARs on their extrasynaptic surface (Andrasfalvy et al. 2003), and as extrasynaptic AMPARs are the main source of receptors for obtaining LTP (Makino & Malinow, 2009; Patterson et al. 2010), GluA1-deficient neurons have a major impairment in LTP (Zamanillo et al. 1999; Granger et al. 2013). While LTD is intact (Selcher et al. 2012), the lack of LTP probably contributes to the large decrease in average synapse strength compared with GluA1-expressing neurons. In GluA3-deficient neurons LTP is not impaired (Meng et al. 2003), which may explain their modest decrease in synapse strength. The notion that synaptic currents depend not only on AMPAR expression levels is supported by the observation that the acute expression of GluA1 in GluA1-deficient neurons did not result in a change in synaptic strength (Kessels et al. 2009). A more definitive estimation of AMPAR subunit usage in synapses awaits experimentation without AMPAR manipulation.
The population of GluA2-lacking AMPARs (i.e. GluA1 homomers) can vary in size, and is regulated by the level of RNA editing of GluA2 (Seeburg & Hartner, 2003; Kwak & Weiss, 2006) or by changing the relative expression levels of GluA1 and GluA2 (Ju et al. 2004; Grooms et al. 2006; Mameli et al. 2007). Neurons that are highly active tend to increase their production of GluA2, while activity blockade selectively increases GluA1 production (Sutton & Schuman, 2006; Mameli et al. 2007; Liu et al. 2010). Since the GluA2-lacking AMPARs are Ca2+ permeable, their presence at the synapse can boost Ca2+ influx upon synaptic activation and thereby lower the threshold for synaptic plasticity (Liu et al. 2006). This suggests that GluA2-lacking AMPARs are involved in a form of homeostatic plasticity (Isaac et al. 2007): an overactive neuron will gradually impede strengthening of its synapses by decreasing its levels of Ca2+-permeable AMPARs, while vice versa a quiet neuron will increase its levels of Ca2+-permeable AMPARs to promote synaptic strengthening. Endogenous GluA2-lacking AMPARs can be transiently inserted into synapses upon LTP induction (Plant et al. 2006), although this is not always detected (Adesnik & Nicoll, 2007). GluA2-lacking AMPARs can also be found enriched in synapses after in vivo experience in the cortex (Clem & Barth, 2006) or memory formation in the hippocampus (Clem & Huganir, 2010).
The role of GluA1-containing AMPARs in LTP was studied by overexpressing GluA1 in hippocampal neurons, which mostly produces GluA1 homomers (Hayashi et al. 2000). Upon overexpression of a subunit, the total AMPAR levels are less than ∼50% above endogenous levels, indicating that the GluA1 homomers largely replace the existing dendritic AMPAR pool without overloading the dendritic AMPAR delivery machinery (Kessels et al. 2009). Under basal conditions these GluA1 homomers are excluded from entering the synapse. Upon stimulation with high-frequency activity, synapses are strengthened through the synaptic insertion of GluA1 homomers (Shi et al. 1999; Hayashi et al. 2000). This LTP-dependent synaptic trafficking of GluA1 is controlled by protein interactions at its c-terminal tail. While a small mutation in the c-terminal PDZ domain of overexpressed GluA1 prevents synaptic delivery (Hayashi et al. 2000; Passafaro et al. 2001), a larger PDZ mutation in a knock-in model did not affect trafficking (Kim et al. 2005). These differences were not a consequence of differences in experimental models, but point towards a mechanism of competitive protein interactions at the GluA1 PDZ domain that regulates GluA1 synaptic delivery (Boehm et al. 2006a2006a). The activity-dependent synaptic delivery of recombinantly expressed GluA1 homomers was shown to also take place in vivo upon amygdala-dependent associative short-term memory formation (Rumpel et al. 2005), hippocampus-dependent short-term contextual memory formation (Mitsushima et al. 2011) and barrel cortex-dependent whisker experience (Takahashi et al. 2003). Notably, GluA1 homomers were shown to behave the same as GluA1/2 heteromers in the whisker experience-dependent synaptic trafficking (Makino & Malinow, 2011). Combined, these studies suggest that GluA1 c-tail-containing AMPARs traffic into synapses in an activity-dependent manner.
As a model for GluA1-lacking AMPARs, overexpression of unedited GluA2 was used. In the absence of a GluA1 c-tail, these GluA2 homomers traffic into synapses independently of activity and without a change in synaptic strength (Shi et al. 2001; Makino & Malinow, 2011). In the barrel cortex synaptic trafficking of GluA2 homomers takes place independently of whisker experience (Takahashi et al. 2003) and mirrors the trafficking of GluA2/3 heteromers (Shi et al. 2001; Makino & Malinow, 2011). GluA2/3s constitutively replace synaptic AMPARs, potentially to keep synaptic strength stable in the face of continuous protein turnover. The replacement of overexpressed GluA1 homomers inserted into hippocampal synapses upon LTP induction or cortical synapses after whisker experience has been shown to not require activity and to take approximately 24 h (Takahashi et al. 2003; McCormack et al. 2006). Whether GluA2/3s are also used for synaptic strengthening during LTP is currently a matter of debate. A recent paper showed that in principle any type of receptor (independent of the c-tail) can be driven into synapses to achieve LTP (Granger et al. 2013). However, when both long-tailed and short-tailed AMPARs are present, the long-tailed ones appear to be preferentially used for synaptic strengthening. This was shown for recombinantly expressed AMPARs (Jia et al. 1996; Hayashi et al. 2000; Shi et al. 2001; Lee et al. 2003; Kopec et al. 2006; Makino & Malinow, 2009; Makino et al. 2011; Tanaka & Hirano, 2012), as well as for endogenous AMPARs in amygdala synapses upon learning (Nedelescu et al. 2010). In addition, targeted in vivo mutations of AMPAR subunits demonstrate that the protein interactions and phosphorylations at the c-tail control their synaptic targeting (Lee et al. 2003; Steinberg et al. 2006). Thus, the long c-tail seems to give AMPARs a competitive advantage to be inserted into synapses upon synaptic strengthening (Sheng et al. 2013). The function of GluA2/3s in memory formation or information processing remains largely unclear. Mice that lack GluA2/3s (i.e. GluA3-knockout mice) have few behavioural deficits (Adamczyk et al. 2012) and create fear memories normally (Humeau et al. 2007). GluA2/3s have been implicated to play a role in the homeostatic scaling of synapse strength (Rial Verde et al. 2006; Gainey et al. 2009; Makino & Malinow, 2011). It is tempting to speculate that the increased propensity of GluA3-deficient mice to develop seizures (Steenland et al. 2008) is due to a defect in synaptic scaling.
NMDA receptor composition (Fig. 1C)
NMDARs are ionotropic receptors that – by transmitting calcium ions – mediate multiple forms of synaptic plasticity (Malenka & Nicoll, 1993). Furthermore, NMDARs serve a structural role and can stabilize synapses (Alvarez et al. 2007). Mechanistically important is their sophisticated regulation of the ion pore. Under resting conditions the channel is blocked by magnesium ions, even in the presence of the endogenous agonist glutamate. Only upon significant depolarization from the resting potential is the magnesium block removed. As a consequence, NMDARs function as coincidence detectors: they only open their channels when presynaptic glutamate release and a postsynaptic depolarization precisely coincide. In most synapses, NMDARs are the major mediators of calcium influx. Therefore, the activity of NMDARs is crucial for the activation of Ca2+-dependent signalling pathways. For example, NMDAR activation and a subsequent intracellular rise in Ca2 are required for the induction of LTP. Also, LTD is triggered by NMDARs and a long held view postulates that NMDAR activation leads to LTD when low amounts of Ca2+ enter the synapse (Cummings et al. 1996). However, a recent study demonstrated that NMDAR-dependent LTD does not depend on calcium influx (Nabavi et al. 2013). Instead, NMDARs also have metabotropic functions beyond their role as ion channels: persistent glutamate binding to the NMDAR in the absence of channel opening activates a signalling pathway that leads to synaptic depression (Kessels et al. 2013; Nabavi et al. 2013).
NMDARs are tetramers, which are composed of two GluN1 subunits and two GluN2 subunits. There are four types of GluN2 subunits, GluN2A–D, which are differentially expressed across brain regions and developmental stages (Cull-Candy et al. 2001; Paoletti et al. 2013). The different GluN2 subunits confer strikingly variable characteristics on the functional receptors, including their kinetics, magnesium sensitivity, ion conductance, molecular interaction partners and – as a consequence – role in synaptic plasticity. As GluN2A and GluN2B predominate in hippocampal and cortical neurons (GluN2C is mainly found in cerebellum and GluN2D is only sparsely expressed) we will discuss the development of these two subspecies of NMDARs (Ewald & Cline, 2009).
NMDAR subunit composition changes during development. GluN2B expression levels are high before birth and in the first 2 weeks after birth. Thereafter, the expression levels of GluN2B decrease (Kirson et al. 1999; Sans et al. 2000). In contrast, GluN2A occurs later during development and the expression of GluN2A increases steadily throughout the first month after birth (Monyer et al. 1994; Sheng et al. 1994; Sans et al. 2000). As a consequence, the ratio of GluN2B to GluN2A in synapses increases during development in an activity-dependent fashion (Carmignoto & Vicini, 1992; Quinlan et al. 1999; Bellone & Nicoll, 2007). The GluN2B and GluN2A subunits differ in several functional aspects. (1) GluN2Bs have a higher affinity for glutamate (Laurie & Seeburg, 1994) and deactivate and desensitize more slowly than GluN2As (Flint et al. 1997; Vicini et al. 1998). (2) The voltage-dependent gating is slightly slower in GluN2B-containing NMDARs (Erreger et al. 2005; Clarke et al. 2013). (3) GluN2B-containing NMDARs traffic more rapidly in the cell membrane than those containing GluN2A, suggesting that in mature neurons NMDARs are more stably anchored (Groc et al. 2006). (4) The structure of the c-terminal region, with which they recruit cytoplasmic proteins to the synapse, differs between GluN2B and GluN2A. For instance, the GluN2B c-tail has a higher affinity for Ca2+/calmodulin-dependent protein kinase II (CaMKII) than the GluN2A c-tail (Strack & Colbran, 1998; Leonard et al. 1999), a protein that is crucial for the induction of LTP (see below). As a consequence, the presence of the GluN2B c-tail at a synapse facilitates LTP induction (Barria & Malinow, 2005; Foster et al. 2010; Ryan et al. 2013). GluN2B-rich synapses are therefore more plastic than synapses that are rich in GluN2A; they have a lower threshold for both LTP (Tang et al. 1999; Clayton et al. 2002) and LTD (Cui et al. 2013). In the mature hippocampus GluN2A eventually outnumbers GluN2B in synapses. Synaptic NMDARs in mature neurons consist of GluN1/GluN2A or GluN1/GluN2A/GluN2B heteromers (Rauner & Kohr, 2011), while GluN1/GluN2B receptors can mostly be found outside synapses (Petralia et al. 2009). Therefore, mature synapses have a reduced ability to undergo plasticity compared with young synapses (Yashiro & Philpot, 2008; Lee et al. 2010b2010b).
Kinases (Fig. 1D) and phosphatases (Fig. 1E)
Synaptic plasticity events are mediated by altering protein interactions through phosphorylation. The phosphorylation targets comprise a myriad of synaptic proteins – including NMDARs and AMPARs. The majority of these phosphorylations take place on serine or threonine residues and a minority on tyrosine residues. There are many types of kinases and phosphatases that differ in substrate selectivity and regulation mechanisms and many of them have roles in synaptic plasticity. Here, we focus on those Ser/Thr kinases/phosphatases that are classically considered the most important for synaptic plasticity: the kinases protein kinase A (PKA), protein kinase C (PKC) and CaMKII, and the phosphatases PP1, PP2A and calcineurin (PP2B).
PKA expression starts after the first week of development and reaches mature levels in the second postnatal week. PKA is a tetrameric molecule with two regulating domains that inhibit the two catalytic domains (Francis & Corbin, 1994). Upon binding of cyclic AMP to the regulatory domain, the catalytic domain is released to phosphorylate its substrates. PKA has a wide range of targets, amongst which are N-methyl-d-aspartate receptors (NMDARs) and AMPARs. Early during development, the PKA-mediated phosphorylation of GluA4 is sufficient for synaptic potentiation (Esteban et al. 2003). At later stages when GluA1 is expressed, PKA phosphorylation of GluA1-containing AMPARs increases their surface levels (Man et al. 2007) and lowers the threshold for LTP (Esteban et al. 2003; Hu et al. 2007; Man et al. 2007; Seol et al. 2007; Lee et al. 2010a2010a).
The PKC family of isozymes is a Ca2+-dependent phospholipid-sensitive group of Ser/Thr kinases. PKC is present in neuronal tissue in high concentration and participates in various neuronal functions including synaptic plasticity (Malenka et al. 1986; Malinow et al. 1989). Phosphorylation of long-tailed AMPARs GluA4 and GluA1 by PKC promotes their surface expression (Gomes et al. 2007; Lin et al. 2009) and LTP (Boehm et al. 2006b2006b). PKC also phosphorylates the short c-tail of GluA2, but with an opposite effect: it leads to internalization of GluA2-containing AMPARs and LTD of synaptic strength (Chung et al. 2003; Seidenman et al. 2003; Lu & Ziff, 2005; Steinberg et al. 2006). The outcome of PKC phosphorylation on synaptic strength therefore probably depends on the AMPAR subunit usage of a synapse. PKC isoforms alpha, beta, epsilon and zeta are expressed both during development and in adults (Roisin & Barbin, 1997). The most abundant isoform in hippocampus, cortex and cerebellum is the neuron-specific PKCγ (Saito & Shirai, 2002). PKCγ expression starts relatively late in comparison with PKA, in the second postnatal week, and reaches adult levels after 3 weeks (Roisin & Barbin, 1997).
CaMKII is crucial for the induction of LTP (Lisman et al. 2012). CaMKII is a circular holoenzyme that consists of 12 (alpha or beta) subunits. Upon LTP induction, calcium influx through the NMDAR channel leads to Ca2+-calmodulin binding to CaMKII and the activation of its catalytic kinase domain. A major phosphorylation target of CaMKII is CaMKII itself. LTP-induced autophosphorylation between neighbouring subunits can keep the CaMKII complex in an active state for up to 1 min after calcium levels have dropped (Miller & Kennedy, 1986; Lee et al. 2009). Upon autophosphorylation CaMKII translocates to the synapse (Shen & Meyer, 1999), where it can bind the GluN2B subunit of the NMDARs (Strack & Colbran, 1998; Leonard et al. 1999), thereby locking CaMKII in an active state (Bayer et al. 2001). Once activated and translocated to the synapse, CaMKII induces spine growth independent of its kinase activity (Pi et al. 2010), suggesting that CaMKII has a crucial structural role during LTP. The CaMKII–GluN2B interaction is vital for the induction of LTP (Barria & Malinow, 2005), and is either reversible or permanent depending on the level and duration of synaptic stimulation (Bayer et al. 2006). After LTP induction CaMKII can reside at the synapse for prolonged periods of time (Otmakhov et al. 2004), where its presence is required for the maintenance of synaptic strength (Sanhueza et al. 2011). Activation of CaMKII is sufficient to induce synaptic strengthening through the addition of GluA1-containing AMPARs to synapses (Hayashi et al. 2000). Besides its autophosphorylation required for LTP induction, CaMKII phosphorylation of GluA1 (Barria et al. 1997; Lu et al. 2010,b2010) and its auxiliary subunits (Tomita et al. 2005; Opazo et al. 2010) facilitate AMPAR insertion into synapses. It is possible that these phosphorylation events may only indirectly promote synaptic AMPAR trafficking by preventing AMPAR degradation and increasing their cell surface levels (Hayashi et al. 2000; He et al. 2009; Kessels et al. 2009). CaMKII phosphorylation events on GluA1 complexes also contribute to LTP by increasing AMPAR single-channel conductance (Derkach et al. 1999; Kristensen et al. 2011). CaMKII-dependent synaptic strengthening is common in adult neurons, but not during early development. In relation to this, CaMKII expression is relatively low during the first 2 weeks of development and increases in the third and fourth week of development (Kelly & Vernon, 1985).
In contrast to LTP, NMDAR-dependent LTD depends largely on phosphatase activity (Malenka & Bear, 2004). LTD is prevented by inhibiting the phosphatases calcineurin (PP2B; Mulkey et al. 1994), PP1 (Mulkey et al. 1993; Kirkwood & Bear, 1994) or PP2A (Nicholls et al. 2008; Mauna et al. 2011). Calcineurin is a Ca2+-calmodulin dependent Ser/Thr phosphatase, is one of the most abundant phosphatases in the nervous system and acts on multiple substrates (Baumgartel & Mansuy, 2012). Calcineurin has a high affinity for Ca2+/calmodulin, and is already active under basal conditions when free Ca2+ levels within the neuron are low, thereby constitutively inhibiting the insertion of GluA1-containing AMPARs into synapses and providing the force to generate LTD (Nabavi et al. 2013). An alternative route to achieve synaptic potentiation is lowering intracellular calcium levels or blocking calcineurin activity (Wang & Kelly, 1997; Nabavi et al. 2013). Previously identified types of synaptic potentiation that involve prolonged silencing or loss of Ca2+ channels (Turrigiano et al. 1998; Ultanir et al. 2007; Adesnik et al. 2008) could potentially be driven by reduced calcineurin activity as a consequence of a drop in intracellular Ca2+. Calcineurin expression starts late during development: expression levels are low during the first 2 weeks of development, and gradually increase during the third and fourth weeks. In contrast, PP1 and PP2A are expressed at maximal levels from birth onwards (Strack et al. 1999; Takahashi et al. 2000) and therefore may control synaptic depression during early stages. Phosphatase activity of PP1 and PP2A is not Ca2+-dependent, but it is regulated through binding of accessory proteins or through their phosphorylation (Munton et al. 2004).
Developmental phases of synaptic plasticity
The expression levels of the key molecules underlying synaptic plasticity change during the first 4 weeks of mouse and rat development. Even though these changes are gradual, they can be placed roughly into three developmental and one mature phase (Fig. 2). Interestingly, these phases are also associated with specific developmental stages beyond the structure and molecular composition of excitatory synapses. These comprise the presence of silent synapses, the switch from depolarization to hyperpolarization at GABAergic synapses, the onset of vision, hearing and active exploration when evoked activity adds to spontaneous activity in shaping circuitry, and finally the critical period after which brain function is usually considered to be mature (Fig. 1F). In the following we describe these developmental stages and analyse how the changes on the molecular and physiological levels may help shape the networks that eventually direct behaviour.
Early postnatal stages of synapse development, PD 0–4 (Fig. 2A)
The behaviour of the newborn rodent is based largely on reflexes. For instance, the suckling reflex requires olfaction and tactile sensation, and is mediated by the brainstem. The hippocampus and cortex receive only limited experiential sensory input during the first days of life (Akerman et al. 2002; Leinekugel et al. 2002; Khazipov et al. 2004; Colonnese et al. 2010). The density of synapses is still very low in hippocampus (Steward & Falk, 1991; Fiala et al. 1998) and cortex (De Felipe et al. 1997), and they are predominantly located on dendritic shafts or filopodia. However, the occurrence of dendrite–axon contacts increases during this period of development. Early steps of synaptic partner selection that are based on cellular recognition mechanisms during the formation of these contacts involve calcium signalling, but occur independently of neuronal activity (Lohmann & Bonhoeffer, 2008; Lohmann, 2009).
Despite the low numbers of functional synapses that are present at this age, spontaneous network activity is prevalent (Allene & Cossart, 2010; Blankenship & Feller, 2010). The transmission of early spontaneous network activity is mediated through gap junctions (Kandler & Katz, 1995; Khazipov & Luhmann, 2006; Niculescu & Lohmann, 2013), transmembrane channels that directly connect the cytoplasm of adjacent cells. Gap junctions mediate both electrical and metabolic coupling of neurons by allowing the passage of ions and small signalling molecules. Through this they coordinate transcriptional activities in developing neurons (Meyer, 1991; Kandler & Katz, 1995; Roerig & Feller, 2000) and contribute to synchronized spontaneous activity. The incidence of gap junction coupling decreases during postnatal weeks 1–3 of the rat hippocampus (Strata et al. 1997) and neocortex (Connors et al. 1983; Peinado et al. 1993). This developmental uncoupling coincides with the major period of synaptogenesis and increased synaptic activity (Kandler & Katz, 1998). Spontaneous activity plays an important role in the establishment of early connections (Katz & Shatz, 1996; Feller, 1999) and can trigger synaptic plasticity in developing networks. For example, pairing spontaneous bursts of network activity with Schaffer collateral stimulation potentiates CA3 to CA1 synapses in a brain-derived neurotrophic factor (BDNF) and extracellular signal-regulated kinase (ERK) pathway-dependent fashion (Kasyanov et al. 2004; Mohajerani et al. 2007). Such correlation-based plasticity mechanisms act on quite long time scales at these ages (several hundred milliseconds; Lee et al. 2002; Kasyanov et al. 2004; Butts et al. 2007; Gjorgjieva et al. 2009) matching the duration of spontaneous bursts. It is likely that the presence of the GluN2B subunit of the NMDAR at this developmental stage, which deactivates and desensitizes more slowly than GluN2As (Flint et al. 1997; Vicini et al. 1998), underlies the ability to integrate correlations across entire bursts of activity.
Furthermore, spontaneous activity is important for an even finer scale of connection specificity: both action potential firing and NMDAR activation are required for sorting synapses on dendrites of hippocampal pyramidal cells in dependence of their presynaptic activity patterns, i.e. the functional clustering of synaptic inputs (Kleindienst et al. 2011). However, how this early plasticity relates to classical LTP mechanisms is unclear, as the critical molecular players involved are either absent or present only at low levels. Furthermore, LTP-inducing protocols trigger postsynaptic depolarizations less efficiently and thus young neurons show higher thresholds for LTP induction (Liao & Malinow, 1996).
Notably, one of the long-tail glutamate receptor subunits, GluA4, is maximally expressed during this period. As GluA4-containing AMPARs can be incorporated into synapses during early steps of synapse formation (Sia et al. 2007), the question emerges as to whether GluA4 may support some form of early synaptic potentiation. Interestingly, GluA4 can be targeted to synapses through a mechanism that requires spontaneous activity in slightly older neurons (P5–7, see below; Zhu et al. 2000). Therefore, it is likely that during the first few days of rodent life the dominant mechanism for AMPAR recruitment to synapses is this spontaneous activity-driven GluA4 trafficking. The GluA4-driven synapse strengthening may play an important role in the initial establishment of the functional circuitry. Interestingly, the GluA4-knockout mouse displays behavioural deficits that mimic aspects of schizophrenia-like behaviour (Sagata et al. 2010). Altered brain development at the fetal or neonatal stage can indeed be the origin of schizophrenia. Whether schizophrenia can originate from mutations in GluA4 is unclear; an association between polymorphisms in the GluA4 gene Gria4 and susceptibility for schizophrenia was found among a Japanese population (Makino et al. 2003), but not in a Chinese or Korean population (Guo et al. 2004; Crisafulli et al. 2012).
Synapse development before onset of visual and auditory experience, PD5–14 (Fig. 2B)
The second postnatal phase of brain development leads up to the moment when mice and rats open their eyes, begin hearing and start interacting with the environment at the end of the second postnatal week. Synapse numbers increase steadily, at a sub-maximal rate in the hippocampus (Steward & Falk, 1991; Fiala et al. 1998) and almost maximally in the cortex (De Felipe et al. 1997). While some sensory inputs can be measured, for example in central somatosensory and visual pathways (Akerman et al. 2002; Khazipov et al. 2004; Colonnese et al. 2010), spontaneously generated activity still dominates the emerging networks (Leinekugel et al. 2002; Demas et al. 2003; Allene et al. 2008; Golshani et al. 2009; Rochefort et al. 2009; Colonnese et al. 2010; Ackman et al. 2012; Siegel et al. 2012). Besides glutamatergic and gap junction signalling (see below), GABAergic transmission is required for normal spontaneous network activity at these ages (Khazipov & Luhmann, 2006; Ben Ari et al. 2007; Allene & Cossart, 2010). Counterintuitively, GABA acts as a depolarizing transmitter during the first postnatal week and becomes inhibitory only during the second week. The cause of the shift from depolarizing to hyperpolarizing action of GABA is a reduction in the intracellular chloride concentration during the second postnatal week, which is mediated in turn by changes in chloride transporter expression (Rivera et al. 1999; Sipila et al. 2006). Importantly, depolarizations at GABAergic synapses can remove the magnesium block of NMDARs during this stage of development (Leinekugel et al. 1997) and may therefore contribute directly to the plasticity or unsilencing of excitatory synapses (Chancey et al. 2013).
The progression of spontaneous activity across brain regions requires, besides chemical synapses, gap junctions during the first week of postnatal development in several rodent brain structures, for example the retina and the cortex (Syed et al. 2004; Dupont et al. 2006; Siegel et al. 2012). Furthermore, recent studies suggest that early gap junction coupling is required for wiring neurons into emerging circuits (Li et al. 2012; Yu et al. 2012). During the first postnatal week, clonally related neurons in the visual cortex are highly connected with each other via gap junctions, but not with other nearby neurons. Later in life, these clones are still highly interconnected, although not through gap junctions, but rather by functional synapses. Remarkably, early electrical coupling is necessary for the formation of chemical synapses between sister neurons later on (Yu et al. 2012) and for the establishment of similar receptive fields among them after eye opening (Li et al. 2012). An intriguing possibility is that gap junctions facilitate simultaneous firing of electrically coupled neurons and thus increase the occurrence of LTP in their common inputs (Yu et al. 2012; Ko et al. 2013).
While the initiation of LTP is not fully developed at the end of the first postnatal week, 1 week later the ability to undergo potentiation is already maximal at hippocampal synapses (Harris & Teyler, 1984; Liao & Malinow, 1996). This correlates with an increase in the number of spine synapses and – on the molecular level – the expression of the key kinases PKA and PKC during the second postnatal week. CaMKII is still expressed at low levels (Kelly & Vernon, 1985) and is, in contrast to PKA, not required for LTP before P14 (Yasuda et al. 2003). The dependence of LTP on PKA during the second week of development correlates with a change in AMPAR subunit usage. Between the first and the second postnatal week, the expression of the long-tailed AMPAR subunits GluA4, GluA2long and GluA1 peaks sequentially (Zhu et al. 2000; Kolleker et al. 2003). The two early expressed subunits, GluA4 and GluA2long, are targeted to synapses by spontaneous activity (Zhu et al. 2000; Kolleker et al. 2003). Also, PKA activation is sufficient to target GluA4 to synapses through phosphorylation of its c-tail (Esteban et al. 2003). As the structures of the GluA4 and GluA2long c-tail are highly conserved around the PKA phosphorylation site, it is possible that PKA phosphorylation of the GluA2long c-tail suffices, too. In contrast, for GluA1, which is expressed later, spontaneous activity or PKA activation is insufficient; GluA1 requires more elaborate activity patterns and CaMKII activation to be inserted into synapses during LTP (Hayashi et al. 2000). The question of whether the evolving changes in AMPAR usage reflect differences in demands for LTP was addressed in a study in the barrel cortex of the rat (Miyazaki et al. 2012). Whisker experience leads to GluA4 synaptic targeting at P8–10 when CaMKII levels are still low. A few days later, at P12–14, the preference is reversed: GluA1 subunits are inserted into synapses upon whisker experience and not GluA4 subunits. Thus, the developmental change in plasticity mechanisms (e.g. PKA or CaMKII driven) coincides with the preference for AMPAR subtype to be used for synaptic insertion.
At the end of the second postnatal week, mice and rats open their eyes and start interacting with their environment. This moment seems to be critical in hippocampal and cortical development. In developmental disorders, errors in connectivity become frequently apparent at this age. For example, SynGAP mutant mice, a model of intellectual disability and autism, exhibit premature spine structure and synaptic function at P14. And even though spines and synapse function appear to be normal at later stages, the behavioural abnormalities are permanent (Clement et al. 2012).
Synapse development after activation of sensory inputs, P15–28 (Fig. 2C)
During the next 2 weeks when young animals start exploring their environment synapse density increases fast in the cortex and hippocampus (Steward & Falk, 1991; De Felipe et al. 1997). This increase is almost exclusively driven by the emergence of spine synapses, and is critically dependent on NMDAR function (Ultanir et al. 2007). During the third week of development, when spines are very dynamic and synapse numbers increase rapidly, synaptic NMDARs are dominantly composed of GluN2B. In neurons in which GluN2B is selectively knocked-out the number of synapses is increased, suggesting that GluN2B mediates the loss of synapses that lack correlated activity (Gray et al. 2011). The expression levels of GluN2A gradually increase at this stage and the GluN2B to GluN2A ratio in synapses decreases. As a consequence of this GluN2B to GluN2A switch, functional and structural changes in spines become more difficult and synapse numbers stabilize (Gambrill & Barria, 2011; Gray et al. 2011). Synapse density reaches a maximum at the end of the fourth week (Steward & Falk, 1991; De Felipe et al. 1997).
On the molecular level, the expression of several late occurring, but central, components of the synaptic plasticity machinery increases steadily until P28. Besides the NMDAR subunit GluN2A, the most important molecular factors are the kinase CaMKII and the phosphatase calcineurin (PP2B). In the third week of development, LTP is only partly dependent on PKA and becomes increasingly dependent on CaMKII, which starts to be expressed in hippocampal neurons at this time. At the end of the fourth postnatal week LTP is fully CaMKII dependent (Yasuda et al. 2003). The increasing expression of calcineurin during the third and fourth weeks of development impedes long-tailed AMPAR entry into synapses (Nabavi et al. 2013), which may raise the threshold for LTP induction. Indeed, whereas in young rats (P12–14) whisker experience is sufficient to drive GluA1-containing AMPARs into synapses, at the third week (P21–23) when functional reorganization of the whisker-barrel map is complete, whisking is insufficient (Jitsuki et al. 2011).
Plasticity in the mature brain, P28 and onwards (Fig. 2D)
Between P28 and P32, synaptic plasticity is temporally enhanced in the mouse visual cortex, and is known to be the critical period for ocular dominance plasticity (Espinosa & Stryker, 2012; Levelt & Hübener, 2012). During this period, a frequently employed plasticity paradigm, monocular deprivation, leads to decreased responses from the deprived eye and increased responses from the open eye and these functional changes are associated with significant rearrangements of synaptic connections. As absolute synapse numbers have reached a maximum by now and are in fact slightly reduced after this period (Steward & Falk, 1991; De Felipe et al. 1997), synapse elimination is probably contributing to refining network connections. The late plasticity molecules, CaMKII, calcineurin and GluN2A, approach adult levels during the critical period, at the end of the fourth week of life. While GluN2A is not required for ocular dominance plasticity in general (Fagiolini et al. 2003; Cho et al. 2009), the relative contribution of LTP is increased whereas LTD is decreased in GluN2A-knockout mice (Cho et al. 2009), suggesting very specific contributions of GluN2A- and GluN2B-mediated synaptic plasticity in adaptations of the visual system during the critical period. In addition, altered plasticity at inhibitory synapses (van Versendaal et al. 2012; Chen et al. 2012) plays a prominent role in the observed differences in visual plasticity during and after the critical period. After the critical period the threshold for plasticity in the visual system is reduced, but it is still possible and previous experience-induced plasticity (during the critical period) reduces the threshold for plasticity in adults (Hofer et al. 2006, 2009,). A reduction in structural plasticity might prevent large-scale reorganization of connections after the critical period (Levelt & Hübener, 2012).
When mature, several types of hippocampal learning and cortical information processing depend on the NMDAR-dependent, CaMKII-mediated synaptic strengthening through the insertion of GluA1-containing AMPARs (Kessels & Malinow, 2009; Mayford et al. 2012). The ability to have synaptic plasticity is reduced in the mature nervous system, partly caused by an increased GluN2A to GluN2B ratio at the level of individual synapses. The adult rodent is capable of having synaptic plasticity, but this is largely absent under basal conditions and requires special circumstances. For instance, in mature mice whisking does not lead to synaptic strengthening at layer 2/3 pyramidal synapses under normal conditions. However, when surrounding whiskers are removed, stimulation of the remaining whisker does lead to synaptic strengthening through deprivation-mediated dis-inhibition (Gambino & Holtmaat, 2012). The most important factor that facilitates synaptic plasticity in the mature brain is emotion. An emotionally charged experience induces the release of modulatory neurotransmitters such as dopamine, serotonin, noradrenaline and glucocorticoids, which can facilitate plasticity through multiple signalling pathways, including synaptic AMPAR trafficking (Hu et al. 2007; Krugers et al. 2010; Jitsuki et al. 2011). For instance, both dopamine and noradrenaline signalling lead to PKA phosphorylation of AMPAR subunit GluA1 (Vanhoose 2003; Winder, 2003; Mangiavacchi & Wolf, 2004), and neurons from mature mice depend more on this phosphorylation event to obtain LTP compared with neurons from young animals (Lee et al. 2003; Lu et al. 2007). The noradrenaline-induced phosphorylation of GluA1 indeed promotes memory formation of a fearful event (Hu et al. 2007).
Conclusion
We have discussed here how the synapses in the brain adapt to the changing requirements for synaptic plasticity. While the emergence of synapse structure and function is gradual during development, we show that it can be divided roughly into four phases based on the expression levels of key plasticity molecules as illustrated in Figs 1 and 2. Early in rodent development, synaptic plasticity mechanisms are employed to create the synaptic network. In the second developmental phase synaptic connections are being fine-tuned to optimally prepare the brain for interpreting and responding to the outside world. During the third phase, synaptic plasticity is used to process, interpret and encode all this new information in the brain. In phase four, the mature hippocampus and cortex use synaptic plasticity to learn, adapt behaviour and create memories. We hope that this overview may serve as a ‘seed crystal’ to establish a framework of the molecular and physiological stages of synapse development. It will be important to investigate whether the expression levels of other developmentally regulated plasticity molecules, in particular those that are involved in presynaptic, dendritic or axonal plasticity mechanisms, correlate with the developmental phases as defined in Fig. 1.
In the rodent brain the developmental phases of synaptic plasticity are completed within 6 weeks after birth. The human brain develops in a much different time frame compared with rodents. First, while rodents are blind until P4–5 and open their eyes at P14, human babies have a functional visual system already at gestational week 20 and open their eyes immediately after birth. This may mean that the first two phases after rodent birth are actually accomplished in the human brain before birth. After birth, however, the human brain takes a much longer and slower path to maturity. It will be interesting to establish how the expression levels of the key synaptic plasticity proteins as shown in Fig. 1 develop in humans.
It is clear that synaptic plasticity mechanisms are important for establishment of the neuronal circuitry. Therefore, disruptions of developmental plasticity mechanisms have long-lasting consequences for cognition and behaviour (Zoghbi, 2003; Sudhof, 2008; Bhakar et al. 2012; Castren et al. 2012). Developmental disorders and other neuropsychiatric illnesses such as autism, schizophrenia, attention deficit hyperactivity disorder (ADHD), anxiety disorders, depression or addiction can be caused genetically (Vorstman & Ophoff, 2013), through physical, hormonal or pharmacological impact during gestation or birth, and through stressful events during birth or early childhood (Insel & Fernald, 2004). Therefore, the mapping of active plasticity mechanisms during the phases of development may provide insights into the specific causes of developmental disorders. Further research integrating molecular, physiological and behavioural aspects will provide a more complete map of synaptic plasticity mechanisms during the different phases of brain development and may inform us at what stage the brain is particularly sensitive to errors in mechanisms of synaptic plasticity and how interventions may prevent cognitive deficits.
Acknowledgments
The authors would like to thank members of their labs for insightful discussions and Drs Maarten Kole and Takuya Takahashi for critical reading of the manuscript.
Additional information
Competing interests
None declared.
Author contributions
None declared.
Funding
Research in the Lohmann and the Kessels labs is supported by the Netherlands Organization for Scientific Research.
References
- Ackman JB, Burbridge TJ, Crair MC. Retinal waves coordinate patterned activity throughout the developing visual system. Nature. 2012;490:219–225. doi: 10.1038/nature11529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adamczyk A, Mejias R, Takamiya K, Yocum J, Krasnova IN, Calderon J, Cadet JL, Huganir RL, Pletnikov MV, Wang T. GluA3-deficiency in mice is associated with increased social and aggressive behavior and elevated dopamine in striatum. Behav Brain Res. 2012;229:265–272. doi: 10.1016/j.bbr.2012.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adesnik H, Li G, During MJ, Pleasure SJ, Nicoll RA. NMDA receptors inhibit synapse unsilencing during brain development. Proc Natl Acad Sci U S A. 2008;105:5597–5602. doi: 10.1073/pnas.0800946105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adesnik H, Nicoll RA. Conservation of glutamate receptor 2-containing AMPA receptors during long-term potentiation. J Neurosci. 2007;27:4598–4602. doi: 10.1523/JNEUROSCI.0325-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akaneya Y( Activity regulates the expression of AMPA receptor subunit GluR4 in developing visual cortex. Eur J Neurosci. 2007;25:1641–1646. doi: 10.1111/j.1460-9568.2007.05388.x. [DOI] [PubMed] [Google Scholar]
- Akerman CJ, Smyth D, Thompson ID. Visual experience before eye-opening and the development of the retinogeniculate pathway. Neuron. 2002;36:869–879. doi: 10.1016/s0896-6273(02)01010-3. [DOI] [PubMed] [Google Scholar]
- Allene C, Cossart R. Early NMDA receptor-driven waves of activity in the developing neocortex: physiological or pathological network oscillations. J Physiol. 2010;588:83–91. doi: 10.1113/jphysiol.2009.178798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allene C, Cattani A, Ackman JB, Bonifazi P, Aniksztejn L, Ben-Ari Y, Cossart R. Sequential generation of two distinct synapse-driven network patterns in developing neocortex. J Neurosci. 2008;28:12851–12863. doi: 10.1523/JNEUROSCI.3733-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alvarez VA, Ridenour DA, Sabatini BL. Distinct structural and ionotropic roles of NMDA receptors in controlling spine and synapse stability. J Neurosci. 2007;27:7365–7376. doi: 10.1523/JNEUROSCI.0956-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andrasfalvy BK, Smith MA, Borchardt T, Sprengel R, Magee JC. Impaired regulation of synaptic strength in hippocampal neurons from GluR1-deficient mice. J Physiol. 2003;552:35–45. doi: 10.1113/jphysiol.2003.045575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aoto J, Ting P, Maghsoodi B, Xu N, Henkemeyer M, Chen L. Postsynaptic EphrinB3 promotes shaft glutamatergic synapse formation. J Neurosci. 2007;27:7508–7519. doi: 10.1523/JNEUROSCI.0705-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Araya R, Eisenthal KB, Yuste R. Dendritic spines linearize the summation of excitatory potentials. Proc Natl Acad Sci U S A. 2006a;103:18799–18804. doi: 10.1073/pnas.0609225103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Araya R, Jiang J, Eisenthal KB, Yuste R. The spine neck filters membrane potentials. Proc Natl Acad Sci U S A. 2006b;103:17961–17966. doi: 10.1073/pnas.0608755103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barria A, Malinow R. NMDA receptor subunit composition controls synaptic plasticity by regulating binding to CaMKII. Neuron. 2005;48:289–301. doi: 10.1016/j.neuron.2005.08.034. [DOI] [PubMed] [Google Scholar]
- Barria A, Muller D, Derkach V, Griffith LC, Soderling TR. Regulatory phosphorylation of AMPA-type glutamate receptors by CaM-KII during long-term potentiation. Science. 1997;276:2042–2045. doi: 10.1126/science.276.5321.2042. [DOI] [PubMed] [Google Scholar]
- Bassani S, Folci A, Zapata J, Passafaro M. AMPAR trafficking in synapse maturation and plasticity. Cell Mol Life Sci. 2013 doi: 10.1007/s00018-013-1309-1. doi: 10.1007/s00018-013-1309-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baumgartel K, Mansuy IM. Neural functions of calcineurin in synaptic plasticity and memory. Learn Mem. 2012;19:375–384. doi: 10.1101/lm.027201.112. [DOI] [PubMed] [Google Scholar]
- Bayer KU, De KP, Leonard AS, Hell JW, Schulman H. Interaction with the NMDA receptor locks CaMKII in an active conformation. Nature. 2001;411:801–805. doi: 10.1038/35081080. [DOI] [PubMed] [Google Scholar]
- Bayer KU, Lebel E, McDonald GL, O’Leary H, Schulman H, De KP. Transition from reversible to persistent binding of CaMKII to postsynaptic sites and NR2B. J Neurosci. 2006;26:1164–1174. doi: 10.1523/JNEUROSCI.3116-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bellone C, Nicoll RA. Rapid bidirectional switching of synaptic NMDA receptors. Neuron. 2007;55:779–785. doi: 10.1016/j.neuron.2007.07.035. [DOI] [PubMed] [Google Scholar]
- Ben Ari Y, Gaiarsa JL, Tyzio R, Khazipov R. GABA: a pioneer transmitter that excites immature neurons and generates primitive oscillations. Physiol Rev. 2007;87:1215–1284. doi: 10.1152/physrev.00017.2006. [DOI] [PubMed] [Google Scholar]
- Bhakar AL, Dölen G, Bear MF. The pathophysiology of fragile x (and what it teaches us about synapses) Annu Rev Neurosci. 2012;35:417–443. doi: 10.1146/annurev-neuro-060909-153138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blankenship AG, Feller MB. Mechanisms underlying spontaneous patterned activity in developing neural circuits. Nat Rev Neurosci. 2010;11:18–29. doi: 10.1038/nrn2759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bloodgood BL, Giessel AJ, Sabatini BL. Biphasic synaptic Ca influx arising from compartmentalized electrical signals in dendritic spines. PLoS Biol. 2009;7:e1000190. doi: 10.1371/journal.pbio.1000190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boehm J, Ehrlich I, Hsieh H, Malinow R. Two mutations preventing PDZ-protein interactions of GluR1 have opposite effects on synaptic plasticity. Learn Mem. 2006a;13:562–565. doi: 10.1101/lm.253506. [DOI] [PubMed] [Google Scholar]
- Boehm J, Kang MG, Johnson RC, Esteban J, Huganir RL, Malinow R. Synaptic incorporation of AMPA receptors during LTP is controlled by a PKC phosphorylation site on GluR1. Neuron. 2006b;51:213–225. doi: 10.1016/j.neuron.2006.06.013. [DOI] [PubMed] [Google Scholar]
- Boyer C, Schikorski T, Stevens CF. Comparison of hippocampal dendritic spines in culture and in brain. J Neurosci. 1998;18:5294–5300. doi: 10.1523/JNEUROSCI.18-14-05294.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Butts DA, Kanold PO, Shatz CJ. A burst-based “Hebbian” learning rule at retinogeniculate synapses links retinal waves to activity-dependent refinement. PLoS Biol. 2007;5:e61. doi: 10.1371/journal.pbio.0050061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carmignoto G, Vicini S. Activity-dependent decrease in NMDA receptor responses during development of the visual cortex. Science. 1992;258:1007–1011. doi: 10.1126/science.1279803. [DOI] [PubMed] [Google Scholar]
- Castren E, Elgersma Y, Maffei L, Hagerman R. Treatment of neurodevelopmental disorders in adulthood. J Neurosci. 2012;32:14074–14079. doi: 10.1523/JNEUROSCI.3287-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chancey JH, Adlaf EW, Sapp MC, Pugh PC, Wadiche JI, Overstreet-Wadiche LS. GABA depolarization is required for experience-dependent synapse unsilencing in adult-born neurons. J Neurosci. 2013;33:6614–6622. doi: 10.1523/JNEUROSCI.0781-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen JL, Villa KL, Cha JW, So PT, Kubota Y, Nedivi E. Clustered dynamics of inhibitory synapses and dendritic spines in the adult neocortex. Neuron. 2012;74:361–373. doi: 10.1016/j.neuron.2012.02.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chklovskii DB( Synaptic connectivity and neuronal morphology: two sides of the same coin. Neuron. 2004;43:609–617. doi: 10.1016/j.neuron.2004.08.012. [DOI] [PubMed] [Google Scholar]
- Cho KK, Khibnik L, Philpot BD, Bear MF. The ratio of NR2A/B NMDA receptor subunits determines the qualities of ocular dominance plasticity in visual cortex. Proc Natl Acad Sci U S A. 2009;106:5377–5382. doi: 10.1073/pnas.0808104106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung HJ, Steinberg JP, Huganir RL, Linden DJ. Requirement of AMPA receptor GluR2 phosphorylation for cerebellar long-term depression. Science. 2003;300:1751. doi: 10.1126/science.1082915. [DOI] [PubMed] [Google Scholar]
- Clarke GL, Chen J, Nishimune H. Presynaptic active zone density during development and synaptic plasticity. Front Mol Neurosci. 2012;5:12. doi: 10.3389/fnmol.2012.00012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke RJ, Glasgow NG, Johnson JW. Mechanistic and structural determinants of NMDA receptor voltage-dependent gating and slow Mg2+ unblock. J Neurosci. 2013;33:4140–4150. doi: 10.1523/JNEUROSCI.3712-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clayton DA, Mesches MH, Alvarez E, Bickford PC, Browning MD. A hippocampal NR2B deficit can mimic age-related changes in long-term potentiation and spatial learning in the Fischer 344 rat. J Neurosci. 2002;22:3628–3637. doi: 10.1523/JNEUROSCI.22-09-03628.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clem RL, Huganir RL. Calcium-permeable AMPA receptor dynamics mediate fear memory erasure. Science. 2010;330:1108–1112. doi: 10.1126/science.1195298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clem RL, Barth A. Pathway-specific trafficking of native AMPARs by in vivo experience. Neuron. 2006;49:663–670. doi: 10.1016/j.neuron.2006.01.019. [DOI] [PubMed] [Google Scholar]
- Clement JP, Aceti M, Creson TK, Ozkan ED, Shi Y, Reish NJ, Almonte AG, Miller BH, Wiltgen BJ, Miller CA, Xu X, Rumbaugh G. Pathogenic SYNGAP1 mutations impair cognitive development by disrupting maturation of dendritic spine synapses. Cell. 2012;151:709–723. doi: 10.1016/j.cell.2012.08.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colonnese MT, Kaminska A, Minlebaev M, Milh M, Bloem B, Lescure S, Moriette G, Chiron C, Ben-Ari Y, Khazipov R. A conserved switch in sensory processing prepares developing neocortex for vision. Neuron. 2010;67:480–498. doi: 10.1016/j.neuron.2010.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Connors BW, Benardo LS, Prince DA. Coupling between neurons of the developing rat neocortex. J Neurosci. 1983;3:773–782. doi: 10.1523/JNEUROSCI.03-04-00773.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crisafulli C, Chiesa A, De RD, Han C, Lee SJ, Park MH, Patkar AA, Pae CU, Serretti A. Influence of GRIA1, GRIA2 and GRIA4 polymorphisms on diagnosis and response to antipsychotic treatment in patients with schizophrenia. Neurosci Lett. 2012;506:170–174. doi: 10.1016/j.neulet.2011.10.074. [DOI] [PubMed] [Google Scholar]
- Cui Z, Feng R, Jacobs S, Duan Y, Wang H, Cao X, Tsien JZ. Increased NR2A:NR2B ratio compresses long-term depression range and constrains long-term memory. Sci Rep. 2013;3:1036. doi: 10.1038/srep01036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cull-Candy S, Brickley S, Farrant M. NMDA receptor subunits: diversity, development and disease. Curr Opin Neurobiol. 2001;11:327–335. doi: 10.1016/s0959-4388(00)00215-4. [DOI] [PubMed] [Google Scholar]
- Cummings JA, Mulkey RM, Nicoll RA, Malenka RC. Ca2+ signaling requirements for long-term depression in the hippocampus. Neuron. 1996;16:825–833. doi: 10.1016/s0896-6273(00)80102-6. [DOI] [PubMed] [Google Scholar]
- De Felipe J, Marco P, Fairen A, Jones EG. Inhibitory synaptogenesis in mouse somatosensory cortex. Cereb Cortex. 1997;7:619–634. doi: 10.1093/cercor/7.7.619. [DOI] [PubMed] [Google Scholar]
- Demas J, Eglen SJ, Wong ROL. Developmental loss of synchronous spontaneous activity in the mouse retina is independent of visual experience. J Neurosci. 2003;23:2851–2860. doi: 10.1523/JNEUROSCI.23-07-02851.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Derkach V, Barria A, Soderling TR. Ca2+/calmodulin-kinase II enhances channel conductance of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionate type glutamate receptors. Proc Natl Acad Sci U S A. 1999;96:3269–3274. doi: 10.1073/pnas.96.6.3269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong YL, Fukazawa Y, Wang W, Kamasawa N, Shigemoto R. Differential postsynaptic compartments in the laterocapsular division of the central nucleus of amygdala for afferents from the parabrachial nucleus and the basolateral nucleus in the rat. J Comp Neurol. 2010;518:4771–4791. doi: 10.1002/cne.22487. [DOI] [PubMed] [Google Scholar]
- Dupont E, Hanganu IL, Kilb W, Hirsch S, Luhmann HJ. Rapid developmental switch in the mechanisms driving early cortical columnar networks. Nature. 2006;439:79–83. doi: 10.1038/nature04264. [DOI] [PubMed] [Google Scholar]
- Erreger K, Dravid SM, Banke TG, Wyllie DJ, Traynelis SF. Subunit-specific gating controls rat NR1/NR2A and NR1/NR2B NMDA channel kinetics and synaptic signalling profiles. J Physiol. 2005;563:345–358. doi: 10.1113/jphysiol.2004.080028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Espinosa JS, Stryker MP. Development and plasticity of the primary visual cortex. Neuron. 2012;75:230–249. doi: 10.1016/j.neuron.2012.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esteban JA, Shi SH, Wilson C, Nuriya M, Huganir RL, Malinow R. PKA phosphorylation of AMPA receptor subunits controls synaptic trafficking underlying plasticity. Nat Neurosci. 2003;6:136–143. doi: 10.1038/nn997. [DOI] [PubMed] [Google Scholar]
- Ethell IM, Pasquale EB. Molecular mechanisms of dendritic spine development and remodeling. Prog Neurobiol. 2005;75:161–205. doi: 10.1016/j.pneurobio.2005.02.003. [DOI] [PubMed] [Google Scholar]
- Ewald RC, Cline HT. NMDA receptors and brain development. In: Van Dongen AM, editor. Biology of the NMDA receptor. CRC Press; 2009. Boca Raton. Chapter 1. [PubMed] [Google Scholar]
- Fagiolini M, Katagiri H, Miyamoto H, Mori H, Grant SG, Mishina M, Hensch TK. Separable features of visual cortical plasticity revealed by N-methyl-D-aspartate receptor 2A signaling. Proc Natl Acad Sci U S A. 2003;100:2854–2859. doi: 10.1073/pnas.0536089100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feldmeyer D, Radnikow G. Developmental alterations in the functional properties of excitatory neocortical synapses. J Physiol. 2009;587:1889–1896. doi: 10.1113/jphysiol.2009.169458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feller MB( Spontaneous correlated activity in developing neural circuits. Neuron. 1999;22:653–656. doi: 10.1016/s0896-6273(00)80724-2. [DOI] [PubMed] [Google Scholar]
- Fiala JC, Feinberg M, Popov V, Harris KM. Synaptogenesis via dendritic filopodia in developing hippocampal area CA1. J Neurosci. 1998;18:8900–8911. doi: 10.1523/JNEUROSCI.18-21-08900.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flint AC, Maisch US, Weishaupt JH, Kriegstein AR, Monyer H. NR2A subunit expression shortens NMDA receptor synaptic currents in developing neocortex. J Neurosci. 1997;17:2469–2476. doi: 10.1523/JNEUROSCI.17-07-02469.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foster KA, McLaughlin N, Edbauer D, Phillips M, Bolton A, Constantine-Paton M, Sheng M. Distinct roles of NR2A and NR2B cytoplasmic tails in long-term potentiation. J Neurosci. 2010;30:2676–2685. doi: 10.1523/JNEUROSCI.4022-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis SH, Corbin JD. Structure and function of cyclic nucleotide-dependent protein kinases. Annu Rev Physiol. 1994;56:237–272. doi: 10.1146/annurev.ph.56.030194.001321. [DOI] [PubMed] [Google Scholar]
- Gainey MA, Hurvitz-Wolff JR, Lambo ME, Turrigiano GG. Synaptic scaling requires the GluR2 subunit of the AMPA receptor. J Neurosci. 2009;29:6479–6489. doi: 10.1523/JNEUROSCI.3753-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gambino F, Holtmaat A. Spike-timing-dependent potentiation of sensory surround in the somatosensory cortex is facilitated by deprivation-mediated disinhibition. Neuron. 2012;75:490–502. doi: 10.1016/j.neuron.2012.05.020. [DOI] [PubMed] [Google Scholar]
- Gambrill AC, Barria A. NMDA receptor subunit composition controls synaptogenesis and synapse stabilization. Proc Natl Acad Sci U S A. 2011;108:5855–5860. doi: 10.1073/pnas.1012676108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geinisman Y, Detoledo-Morrell L, Morrell F, Persina IS, Beatty MA. Synapse restructuring associated with the maintenance phase of hippocampal long-term potentiation. J Comp Neurol. 1996;368:413–423. doi: 10.1002/(SICI)1096-9861(19960506)368:3<413::AID-CNE7>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- Gjorgjieva J, Toyoizumi T, Eglen SJ. Burst-time-dependent plasticity robustly guides ON/OFF segregation in the lateral geniculate nucleus. PLoS Comput Biol. 2009;5:e1000618. doi: 10.1371/journal.pcbi.1000618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golshani P, Goncalves JT, Khoshkhoo S, Mostany R, Smirnakis S, Portera-Cailliau C. Internally mediated developmental desynchronization of neocortical network activity. J Neurosci. 2009;29:10890–10899. doi: 10.1523/JNEUROSCI.2012-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomes AR, Correia SS, Esteban JA, Duarte CB, Carvalho AL. PKC anchoring to GluR4 AMPA receptor subunit modulates PKC-driven receptor phosphorylation and surface expression. Traffic. 2007;8:259–269. doi: 10.1111/j.1600-0854.2006.00521.x. [DOI] [PubMed] [Google Scholar]
- Granger AJ, Shi Y, Lu W, Cerpas M, Nicoll RA. LTP requires a reserve pool of glutamate receptors independent of subunit type. Nature. 2013;493:495–500. doi: 10.1038/nature11775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray JA, Shi Y, Usui H, During MJ, Sakimura K, Nicoll RA. Distinct modes of AMPA receptor suppression at developing synapses by GluN2A and GluN2B: single-cell NMDA receptor subunit deletion in vivo. Neuron. 2011;71:1085–1101. doi: 10.1016/j.neuron.2011.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greger IH, Khatri L, Kong X, Ziff EB. AMPA receptor tetramerization is mediated by Q/R editing. Neuron. 2003;40:763–774. doi: 10.1016/s0896-6273(03)00668-8. [DOI] [PubMed] [Google Scholar]
- Greger IH, Khatri L, Ziff EB. RNA editing at arg607 controls AMPA receptor exit from the endoplasmic reticulum. Neuron. 2002;34:759–772. doi: 10.1016/s0896-6273(02)00693-1. [DOI] [PubMed] [Google Scholar]
- Groc L, Gustafsson B, Hanse E. Developmental alterations in the functional properties of excitatory neocortical synapses. Trends Neurosci. 2009;587:1889–1896. [Google Scholar]
- Groc L, Heine M, Cousins SL, Stephenson FA, Lounis B, Cognet L, Choquet D. NMDA receptor surface mobility depends on NR2A-2B subunits. Proc Natl Acad Sci U S A. 2006;103:18769–18774. doi: 10.1073/pnas.0605238103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grooms SY, Noh KM, Regis R, Bassell GJ, Bryan MK, Carroll RC, Zukin RS. Activity bidirectionally regulates AMPA receptor mRNA abundance in dendrites of hippocampal neurons. J Neurosci. 2006;26:8339–8351. doi: 10.1523/JNEUROSCI.0472-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grunditz A, Holbro N, Tian L, Zuo Y, Oertner TG. Spine neck plasticity controls postsynaptic calcium signals through electrical compartmentalization. J Neurosci. 2008;28:13457–13466. doi: 10.1523/JNEUROSCI.2702-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gulledge AT, Carnevale NT, Stuart GJ. Electrical advantages of dendritic spines. PLoS ONE. 2012;7:e36007. doi: 10.1371/journal.pone.0036007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo S, Shi Y, Zhao X, Duan S, Zhou J, Meng J, Yang Y, Gu N, Feng G, Liu H, Zhu S, He L. No genetic association between polymorphisms in the AMPA receptor subunit GluR4 gene (GRIA4) and schizophrenia in the Chinese population. Neurosci Lett. 2004;369:168–172. doi: 10.1016/j.neulet.2004.07.079. [DOI] [PubMed] [Google Scholar]
- Harnett MT, Makara JK, Spruston N, Kath WL, Magee JC. Synaptic amplification by dendritic spines enhances input cooperativity. Nature. 2012;491:599–602. doi: 10.1038/nature11554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris KM, Jensen FE, Tsao B. Three-dimensional structure of dendritic spines and synapses in rat hippocampus (CA1) at postnatal day 15 and adult ages: implications for the maturation of synaptic physiology and long-term potentiation. J Neurosci. 1992;12:2685–2705. doi: 10.1523/JNEUROSCI.12-07-02685.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris KM, Teyler TJ. Developmental onset of long-term potentiation in area CA1 of the rat hippocampus. J Physiol. 1984;346:27–48. doi: 10.1113/jphysiol.1984.sp015005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi Y, Shi SH, Esteban JA, Piccini A, Poncer JC, Malinow R. Driving AMPA receptors into synapses by LTP and CaMKII: requirement for GluR1 and PDZ domain interaction. Science. 2000;287:2262–2267. doi: 10.1126/science.287.5461.2262. [DOI] [PubMed] [Google Scholar]
- He K, Song L, Cummings LW, Goldman J, Huganir RL, Lee HK. Stabilization of Ca2+-permeable AMPA receptors at perisynaptic sites by GluR1-S845 phosphorylation. Proc Natl Acad Sci U S A. 2009;106:20033–20038. doi: 10.1073/pnas.0910338106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helmeke C, Ovtscharoff W, Jr, Poeggel G, Braun K. Juvenile emotional experience alters synaptic inputs on pyramidal neurons in the anterior cingulate cortex. Cereb Cortex. 2001;11:717–727. doi: 10.1093/cercor/11.8.717. [DOI] [PubMed] [Google Scholar]
- Hofer SB, Mrsic-Flogel TD, Bonhoeffer T, Hubener M. Prior experience enhances plasticity in adult visual cortex. Nat Neurosci. 2006;9:127–132. doi: 10.1038/nn1610. [DOI] [PubMed] [Google Scholar]
- Hofer SB, Mrsic-Flogel TD, Bonhoeffer T, Hubener M. Experience leaves a lasting structural trace in cortical circuits. Nature. 2009;457:313–317. doi: 10.1038/nature07487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holtmaat AJ, Trachtenberg JT, Wilbrecht L, Shepherd GM, Zhang X, Knott GW, Svoboda K. Transient and persistent dendritic spines in the neocortex in vivo. Neuron. 2005;45:279–291. doi: 10.1016/j.neuron.2005.01.003. [DOI] [PubMed] [Google Scholar]
- Hu H, Real E, Takamiya K, Kang MG, Ledoux J, Huganir RL, Malinow R. Emotion enhances learning via norepinephrine regulation of AMPA-receptor trafficking. Cell. 2007;131:160–173. doi: 10.1016/j.cell.2007.09.017. [DOI] [PubMed] [Google Scholar]
- Huang ZJ, Scheiffele P. GABA and neuroligin signaling: linking synaptic activity and adhesion in inhibitory synapse development. Curr Opin Neurobiol. 2008;18:77–83. doi: 10.1016/j.conb.2008.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humeau Y, Reisel D, Johnson AW, Borchardt T, Jensen V, Gebhardt C, Bosch V, Gass P, Bannerman DM, Good MA, Hvalby O, Sprengel R, Luthi A. A pathway-specific function for different AMPA receptor subunits in amygdala long-term potentiation and fear conditioning. J Neurosci. 2007;27:10947–10956. doi: 10.1523/JNEUROSCI.2603-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Insel TR, Fernald RD. How the brain processes social information: searching for the social brain. Annu Rev Neurosci. 2004;27:697–722. doi: 10.1146/annurev.neuro.27.070203.144148. [DOI] [PubMed] [Google Scholar]
- Isaac JT, Ashby M, McBain CJ. The role of the GluR2 subunit in AMPA receptor function and synaptic plasticity. Neuron. 2007;54:859–871. doi: 10.1016/j.neuron.2007.06.001. [DOI] [PubMed] [Google Scholar]
- Jia ZP, Agopyan N, Miu P, Xiong ZG, Henderson J, Gerlai R, Taverna FA, Velumian A, Macdonald J, Carlen P, Abramownewerly W, Roder J. Enhanced LTP in mice deficient in the AMPA receptor GluR2. Neuron. 1996;17:945–956. doi: 10.1016/s0896-6273(00)80225-1. [DOI] [PubMed] [Google Scholar]
- Jitsuki S, Takemoto K, Kawasaki T, Tada H, Takahashi A, Becamel C, Sano A, Yuzaki M, Zukin RS, Ziff EB, Kessels HW, Takahashi T. Serotonin mediates cross-modal reorganization of cortical circuits. Neuron. 2011;69:780–792. doi: 10.1016/j.neuron.2011.01.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ju W, Morishita W, Tsui J, Gaietta G, Deerinck TJ, Adams SR, Garner CC, Tsien RY, Ellisman MH, Malenka RC. Activity-dependent regulation of dendritic synthesis and trafficking of AMPA receptors. Nat Neurosci. 2004;7:244–253. doi: 10.1038/nn1189. [DOI] [PubMed] [Google Scholar]
- Kandler K, Katz LC. Neuronal coupling and uncoupling in the developing nervous system. Curr Opin Neurobiol. 1995;5:98–105. doi: 10.1016/0959-4388(95)80093-x. [DOI] [PubMed] [Google Scholar]
- Kandler K, Katz LC. Relationship between dye coupling and spontaneous activity in developing ferret visual cortex. Dev Neurosci. 1998;20:59–64. doi: 10.1159/000017299. [DOI] [PubMed] [Google Scholar]
- Kasyanov AM, Safiulina VF, Voronin LL, Cherubini E. GABA-mediated giant depolarizing potentials as coincidence detectors for enhancing synaptic efficacy in the developing hippocampus. Proc Natl Acad Sci U S A. 2004;101:3967–3972. doi: 10.1073/pnas.0305974101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katz LC, Shatz CJ. Synaptic activity and the construction of cortical circuits. Science. 1996;274:1133–1138. doi: 10.1126/science.274.5290.1133. [DOI] [PubMed] [Google Scholar]
- Kelly PT. Protein phosphorylation in isolated synaptic junctional structures: changes during development. Brain Res. 1982;247:85–96. doi: 10.1016/0006-8993(82)91030-7. [DOI] [PubMed] [Google Scholar]
- Kelly PT, Vernon P. Changes in the subcellular distribution of calmodulin-kinase II during brain development. Brain Res. 1985;350:211–224. doi: 10.1016/0165-3806(85)90265-2. [DOI] [PubMed] [Google Scholar]
- Kerchner GA, Nicoll RA. Silent synapses and the emergence of a postsynaptic mechanism for LTP. Nat Rev Neurosci. 2008;9:813–825. doi: 10.1038/nrn2501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kessels HW, Kopec CD, Klein ME, Malinow R. Roles of stargazin and phosphorylation in the control of AMPA receptor subcellular distribution. Nat Neurosci. 2009;12:888–896. doi: 10.1038/nn.2340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kessels HW, Malinow R. Synaptic AMPA receptor plasticity and behavior. Neuron. 2009;61:340–350. doi: 10.1016/j.neuron.2009.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kessels HW, Nabavi S, Malinow R. Metabotropic NMDA receptor function is required for beta-amyloid-induced synaptic depression. Proc Natl Acad Sci U S A. 2013;110:4033–4038. doi: 10.1073/pnas.1219605110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khazipov R, Sirota A, Leinekugel X, Holmes GL, Ben-Ari Y, Buzsaki G. Early motor activity drives spindle bursts in the developing somatosensory cortex. Nature. 2004;432:758–761. doi: 10.1038/nature03132. [DOI] [PubMed] [Google Scholar]
- Khazipov R, Luhmann HJ. Early patterns of electrical activity in the developing cerebral cortex of humans and rodents. Trends Neurosci. 2006;29:414–418. doi: 10.1016/j.tins.2006.05.007. [DOI] [PubMed] [Google Scholar]
- Kim CH, Takamiya K, Petralia RS, Sattler R, Yu S, Zhou W, Kalb R, Wenthold R, Huganir R. Persistent hippocampal CA1 LTP in mice lacking the C-terminal PDZ ligand of GluR1. Nat Neurosci. 2005;8:985–987. doi: 10.1038/nn1432. [DOI] [PubMed] [Google Scholar]
- Kirkwood A, Bear MF. Homosynaptic long-term depression in the visual cortex. J Neurosci. 1994;14:3404–3412. doi: 10.1523/JNEUROSCI.14-05-03404.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirson ED, Schirra C, Konnerth A, Yaari Y. Early postnatal switch in magnesium sensitivity of NMDA receptors in rat CA1 pyramidal cells. J Physiol. 1999;521:99–111. doi: 10.1111/j.1469-7793.1999.00099.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kleindienst T, Winnubst J, Roth-Alpermann C, Bonhoeffer T, Lohmann C. Activity-dependent clustering of functional synaptic inputs on developing hippocampal dendrites. Neuron. 2011;72:1012–1024. doi: 10.1016/j.neuron.2011.10.015. [DOI] [PubMed] [Google Scholar]
- Knott GW, Holtmaat A, Wilbrecht L, Welker E, Svoboda K. Spine growth precedes synapse formation in the adult neocortex in vivo. Nat Neurosci. 2006;9:1117–1124. doi: 10.1038/nn1747. [DOI] [PubMed] [Google Scholar]
- Ko H, Cossell L, Baragli C, Antolik J, Clopath C, Hofer SB, Mrsic-Flogel TD. The emergence of functional microcircuits in visual cortex. Nature. 2013;496:96–100. doi: 10.1038/nature12015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch C, Zador A. The function of dendritic spines: devices subserving biochemical rather than electrical compartmentalization. J Neurosci. 1993;13:413–422. doi: 10.1523/JNEUROSCI.13-02-00413.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch C, Zador A, Brown TH. Dendritic spines – convergence of theory and experiment. Science. 1992;256:973–974. doi: 10.1126/science.1589781. [DOI] [PubMed] [Google Scholar]
- Kolleker A, Zhu JJ, Schupp BJ, Qin Y, Mack V, Borchardt T, Kohr G, Malinow R, Seeburg PH, Osten P. Glutamatergic plasticity by synaptic delivery of GluR-Blong-containing AMPA receptors. Neuron. 2003;40:1199–1212. doi: 10.1016/s0896-6273(03)00722-0. [DOI] [PubMed] [Google Scholar]
- Kopec CD, Li B, Wei W, Boehm J, Malinow R. Glutamate receptor exocytosis and spine enlargement during chemically induced long-term potentiation. J Neurosci. 2006;26:2000–2009. doi: 10.1523/JNEUROSCI.3918-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kristensen AS, Jenkins MA, Banke TG, Schousboe A, Makino Y, Johnson RC, Huganir R, Traynelis SF. Mechanism of Ca2+/calmodulin-dependent kinase II regulation of AMPA receptor gating. Nat Neurosci. 2011;14:727–735. doi: 10.1038/nn.2804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krugers HJ, Hoogenraad CC, Groc L. Stress Hormones and AMPA receptor trafficking in synaptic plasticity and memory. Nat Rev Neurosci. 2010;11:675–681. doi: 10.1038/nrn2913. [DOI] [PubMed] [Google Scholar]
- Kwak S, Weiss JH. Calcium-permeable AMPA channels in neurodegenerative disease and ischemia. Curr Opin Neurobiol. 2006;16:281–287. doi: 10.1016/j.conb.2006.05.004. [DOI] [PubMed] [Google Scholar]
- Kwon HB, Sabatini BL. Glutamate induces de novo growth of functional spines in developing cortex. Nature. 2011;474:100–104. doi: 10.1038/nature09986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laurie DJ, Seeburg PH. Ligand affinities at recombinant N-methyl-D-aspartate receptors depend on subunit composition. Eur J Pharmacol. 1994;268:335–345. doi: 10.1016/0922-4106(94)90058-2. [DOI] [PubMed] [Google Scholar]
- Lee CW, Eglen SJ, Wong RO. Segregation of ON and OFF retinogeniculate connectivity directed by patterned spontaneous activity. J Neurophysiol. 2002;88:2311–2321. doi: 10.1152/jn.00372.2002. [DOI] [PubMed] [Google Scholar]
- Lee HK, Takamiya K, Han JS, Man H, Kim CH, Rumbaugh G, Yu S, Ding L, He C, Petralia RS, Wenthold RJ, Gallagher M, Huganir RL. Phosphorylation of the AMPA receptor GluR1 subunit is required for synaptic plasticity and retention of spatial memory. Cell. 2003;112:631–643. doi: 10.1016/s0092-8674(03)00122-3. [DOI] [PubMed] [Google Scholar]
- Lee HK, Takamiya K, He K, Song L, Huganir RL. Specific roles of AMPA receptor subunit GluR1 (GluA1) phosphorylation sites in regulating synaptic plasticity in the CA1 region of hippocampus. J Neurophysiol. 2010a;103:479–489. doi: 10.1152/jn.00835.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee MC, Yasuda R, Ehlers MD. Metaplasticity at single glutamatergic synapses. Neuron. 2010b;66:859–870. doi: 10.1016/j.neuron.2010.05.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee SJ, Escobedo-Lozoya Y, Szatmari EM, Yasuda R. Activation of CaMKII in single dendritic spines during long-term potentiation. Nature. 2009;458:299–304. doi: 10.1038/nature07842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leinekugel X, Khazipov R, Cannon R, Hirase H, Ben-Ari Y, Buzsaki G. Correlated bursts of activity in the neonatal hippocampus in vivo. Science. 2002;296:2049–2052. doi: 10.1126/science.1071111. [DOI] [PubMed] [Google Scholar]
- Leinekugel X, Medina I, Khalilov I, Ben-Ari Y, Khazipov R. Ca2+ oscillations mediated by the synergistic excitatory actions of GABAA and NMDA receptors in the neonatal hippocampus. Neuron. 1997;18:243–255. doi: 10.1016/s0896-6273(00)80265-2. [DOI] [PubMed] [Google Scholar]
- Leonard AS, Lim IA, Hemsworth DE, Horne MC, Hell JW. Calcium/calmodulin-dependent protein kinase II is associated with the N-methyl-D-aspartate receptor. Proc Natl Acad Sci U S A. 1999;96:3239–3244. doi: 10.1073/pnas.96.6.3239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levelt CN, Hübener M. Critical-period plasticity in the visual cortex. Annu Rev Neurosci. 2012;35:309–330. doi: 10.1146/annurev-neuro-061010-113813. [DOI] [PubMed] [Google Scholar]
- Li Y, Lu H, Cheng Pl, Ge S, Xu H, Shi SH, Dan Y. Clonally related visual cortical neurons show similar stimulus feature selectivity. Nature. 2012;486:118–121. doi: 10.1038/nature11110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liao D, Malinow R. Deficiency in induction but not expression of LTP in hippocampal slices from young rats. Learn Mem. 1996;3:138–149. doi: 10.1101/lm.3.2-3.138. [DOI] [PubMed] [Google Scholar]
- Lin DT, Makino Y, Sharma K, Hayashi T, Neve R, Takamiya K, Huganir RL. Regulation of AMPA receptor extrasynaptic insertion by 4.1N, phosphorylation and palmitoylation. Nat Neurosci. 2009;12:879–887. doi: 10.1038/nn.2351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lisman J, Yasuda R, Raghavachari S. Mechanisms of CaMKII action in long-term potentiation. Nat Rev Neurosci. 2012;13:169–182. doi: 10.1038/nrn3192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu B, Liao M, Mielke JG, Ning K, Chen Y, Li L, El-Hayek YH, Gomez E, Zukin RS, Fehlings MG, Wan Q. Ischemic insults direct glutamate receptor subunit 2-lacking AMPA receptors to synaptic sites. J Neurosci. 2006;26:5309–5319. doi: 10.1523/JNEUROSCI.0567-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Y, Formisano L, Savtchouk I, Takayasu Y, Szabo G, Zukin RS, Liu SJ. A single fear-inducing stimulus induces a transcription-dependent switch in synaptic AMPAR phenotype. Nat Neurosci. 2010;13:223–231. doi: 10.1038/nn.2474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loewenstein Y, Kuras A, Rumpel S. Multiplicative dynamics underlie the emergence of the log-normal distribution of spine sizes in the neocortex in vivo. J Neurosci. 2011;31:9481–9488. doi: 10.1523/JNEUROSCI.6130-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lohmann C( Calcium signaling and the development of specific neuronal connections. Prog Brain Res. 2009;175:443–452. doi: 10.1016/S0079-6123(09)17529-5. [DOI] [PubMed] [Google Scholar]
- Lohmann C, Bonhoeffer T. A role for local calcium signaling in rapid synaptic partner selection by dendritic filopodia. Neuron. 2008;59:253–260. doi: 10.1016/j.neuron.2008.05.025. [DOI] [PubMed] [Google Scholar]
- Lu W, Isozaki K, Roche KW, Nicoll RA. Synaptic targeting of AMPA receptors is regulated by a CaMKII site in the first intracellular loop of GluA1. Proc Natl Acad Sci U S A. 2010;107:22266–22271. doi: 10.1073/pnas.1016289107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu W, Shi Y, Jackson AC, Bjorgan K, During MJ, Sprengel R, Seeburg PH, Nicoll RA. Subunit composition of synaptic AMPA receptors revealed by a single-cell genetic approach. Neuron. 2009;62:254–268. doi: 10.1016/j.neuron.2009.02.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu W, Ziff EB. PICK1 interacts with ABP/GRIP to regulate AMPA receptor trafficking. Neuron. 2005;47:407–421. doi: 10.1016/j.neuron.2005.07.006. [DOI] [PubMed] [Google Scholar]
- Lu Y, Allen M, Halt AR, Weisenhaus M, Dallapiazza RF, Hall DD, Usachev YM, McKnight GS, Hell JW. Age-dependent requirement of AKAP150-anchored PKA and GluR2-lacking AMPA receptors in LTP. EMBO J. 2007;26:4879–4890. doi: 10.1038/sj.emboj.7601884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mangiavacchi S, Wolf ME. D1 dopamine receptor stimulation increases the rate of AMPA receptor insertion onto the surface of cultured nucleus accumbens neurons through a pathway dependent on protein kinase A. J Neurochem. 2004;88:1261–1271. doi: 10.1046/j.1471-4159.2003.02248.x. [DOI] [PubMed] [Google Scholar]
- Makino C, Fujii Y, Kikuta R, Hirata N, Tani A, Shibata A, Ninomiya H, Tashiro N, Shibata H, Fukumaki Y. Positive association of the AMPA receptor subunit GluR4 gene (GRIA4) haplotype with schizophrenia: linkage disequilibrium mapping using SNPs evenly distributed across the gene region. Am J Med Genet B Neuropsychiatr Genet. 2003;116B:17–22. doi: 10.1002/ajmg.b.10041. [DOI] [PubMed] [Google Scholar]
- Makino H, Malinow R. AMPA receptor incorporation into synapses during LTP: the role of lateral movement and exocytosis. Neuron. 2009;64:381–390. doi: 10.1016/j.neuron.2009.08.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makino H, Malinow R. Compartmentalized versus global synaptic plasticity on dendrites controlled by experience. Neuron. 2011;72:1001–1011. doi: 10.1016/j.neuron.2011.09.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makino Y, Johnson RC, Yu Y, Takamiya K, Huganir RL. Enhanced synaptic plasticity in mice with phosphomimetic mutation of the GluA1 AMPA receptor. Proc Natl Acad Sci U S A. 2011;108:8450–8455. doi: 10.1073/pnas.1105261108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malenka RC, Bear MF. LTP and LTD: an embarrassment of riches. Neuron. 2004;44:5–21. doi: 10.1016/j.neuron.2004.09.012. [DOI] [PubMed] [Google Scholar]
- Malenka RC, Madison DV, Nicoll RA. Potentiation of synaptic transmission in the hippocampus by phorbol esters. Nature. 1986;321:175–177. doi: 10.1038/321175a0. [DOI] [PubMed] [Google Scholar]
- Malenka RC, Nicoll RA. NMDA-receptor-dependent synaptic plasticity: multiple forms and mechanisms. Trends Neurosci. 1993;16:521–527. doi: 10.1016/0166-2236(93)90197-t. [DOI] [PubMed] [Google Scholar]
- Malinow R, Malenka RC. AMPA receptor trafficking and synaptic plasticity. Annu Rev Neurosci. 2002;25:103–126. doi: 10.1146/annurev.neuro.25.112701.142758. [DOI] [PubMed] [Google Scholar]
- Malinow R, Schulman H, Tsien RW. Inhibition of postsynaptic PKC or CaMKII blocks induction but not expression of LTP. Science. 1989;245:862–866. doi: 10.1126/science.2549638. [DOI] [PubMed] [Google Scholar]
- Mameli M, Balland B, Lujan R, Luscher C. Rapid synthesis and synaptic insertion of GluR2 for mGluR-LTD in the ventral tegmental area. Science. 2007;317:530–533. doi: 10.1126/science.1142365. [DOI] [PubMed] [Google Scholar]
- Man HY, Sekine-Aizawa Y, Huganir RL. Regulation of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor trafficking through PKA phosphorylation of the Glu receptor 1 subunit. Proc Natl Acad Sci U S A. 2007;104:3579–3584. doi: 10.1073/pnas.0611698104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mauna JC, Miyamae T, Pulli B, Thiels E. Protein phosphatases 1 and 2A are both required for long-term depression and associated dephosphorylation of cAMP response element binding protein in hippocampal area CA1 in vivo. Hippocampus. 2011;21:1093–1104. doi: 10.1002/hipo.20823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayford M, Siegelbaum SA, Kandel ER. Synapses and memory storage. Cold Spring Harb Perspect Biol. 2012;4 doi: 10.1101/cshperspect.a005751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCormack SG, Stornetta RL, Zhu JJ. Synaptic AMPA receptor exchange maintains bidirectional plasticity. Neuron. 2006;50:75–88. doi: 10.1016/j.neuron.2006.02.027. [DOI] [PubMed] [Google Scholar]
- Meng Y, Zhang Y, Jia Z. Synaptic transmission and plasticity in the absence of AMPA glutamate receptor GluR2 and GluR3. Neuron. 2003;39:163–176. doi: 10.1016/s0896-6273(03)00368-4. [DOI] [PubMed] [Google Scholar]
- Meyer T( Cell signaling by second messenger waves. Cell. 1991;64:675–678. doi: 10.1016/0092-8674(91)90496-l. [DOI] [PubMed] [Google Scholar]
- Miller SG, Kennedy MB. Regulation of brain type II Ca2+/calmodulin-dependent protein kinase by autophosphorylation: a Ca2+-triggered molecular switch. Cell. 1986;44:861–870. doi: 10.1016/0092-8674(86)90008-5. [DOI] [PubMed] [Google Scholar]
- Mitsushima D, Ishihara K, Sano A, Kessels HW, Takahashi T. Contextual learning requires synaptic AMPA receptor delivery in the hippocampus. Proc Natl Acad Sci USA. 2011;108:12503–12508. doi: 10.1073/pnas.1104558108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyazaki T, Kunii M, Tada H, Sano A, Kuroiwa Y, Goto T, Malinow R, Takahashi T. Developmental AMPA receptor subunit specificity during experience-driven synaptic plasticity in the rat barrel cortex. Brain Res. 2012;1435:1–7. doi: 10.1016/j.brainres.2011.11.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohajerani MH, Sivakumaran S, Zacchi P, Aguilera P, Cherubini E. Correlated network activity enhances synaptic efficacy via BDNF and the ERK pathway at immature CA3 CA1 connections in the hippocampus. Proc Natl Acad Sci U S A. 2007;104:13176–13181. doi: 10.1073/pnas.0704533104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monyer H, Burnashev N, Laurie DJ, Sakmann B, Seeburg PH. Developmental and regional expression in the rat brain and functional properties of four NMDA receptors. Neuron. 1994;12:529–540. doi: 10.1016/0896-6273(94)90210-0. [DOI] [PubMed] [Google Scholar]
- Mulkey RM, Endo S, Shenolikar S, Malenka RC. Involvement of a calcineurin/inhibitor-1 phosphatase cascade in hippocampal long-term depression. Nature. 1994;369:486–488. doi: 10.1038/369486a0. [DOI] [PubMed] [Google Scholar]
- Mulkey RM, Herron CE, Malenka RC. An essential role for protein phosphatases in hippocampal long- term depression. Science. 1993;261:1051–1055. doi: 10.1126/science.8394601. [DOI] [PubMed] [Google Scholar]
- Muller W, Connor JA. Dendritic spines as individual neuronal compartments for synaptic Ca2+ responses. Nature. 1991;354:73–76. doi: 10.1038/354073a0. [DOI] [PubMed] [Google Scholar]
- Munton RP, Vizi S, Mansuy IM. The role of protein phosphatase-1 in the modulation of synaptic and structural plasticity. FEBS Lett. 2004;567:121–128. doi: 10.1016/j.febslet.2004.03.121. [DOI] [PubMed] [Google Scholar]
- Nabavi S, Kessels HW, Alfonso S, Aow J, Fox R, Malinow R. Metabotropic NMDA receptor function is required for NMDA receptor-dependent long-term depression. Proc Natl Acad Sci U S A. 2013;110:4027–4032. doi: 10.1073/pnas.1219454110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagerl UV, Kostinger G, Anderson JC, Martin KA, Bonhoeffer T. Protracted synaptogenesis after activity-dependent spinogenesis in hippocampal neurons. J Neurosci. 2007;27:8149–8156. doi: 10.1523/JNEUROSCI.0511-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nedelescu H, Kelso CM, Lazaro-Munoz G, Purpura M, Cain CK, Ledoux JE, Aoki C. Endogenous GluR1-containing AMPA receptors translocate to asymmetric synapses in the lateral amygdala during the early phase of fear memory formation: an electron microscopic immunocytochemical study. J Comp Neurol. 2010;518:4723–4739. doi: 10.1002/cne.22472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicholls RE, Alarcon JM, Malleret G, Carroll RC, Grody M, Vronskaya S, Kandel ER. Transgenic mice lacking NMDAR-dependent LTD exhibit deficits in behavioral flexibility. Neuron. 2008;58:104–117. doi: 10.1016/j.neuron.2008.01.039. [DOI] [PubMed] [Google Scholar]
- Niculescu D, Lohmann C. Gap junctions in developing thalamic and neocortical neuronal networks. Cereb Cortex. 2013 doi: 10.1093/cercor/bht175. doi: 10.1093/cercor/bht175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Opazo P, Labrecque S, Tigaret CM, Frouin A, Wiseman PW, De KP, Choquet D. CaMKII triggers the diffusional trapping of surface AMPARs through phosphorylation of stargazin. Neuron. 2010;67:239–252. doi: 10.1016/j.neuron.2010.06.007. [DOI] [PubMed] [Google Scholar]
- Otmakhov N, Tao-Cheng JH, Carpenter S, Asrican B, Dosemeci A, Reese TS, Lisman J. Persistent accumulation of calcium/calmodulin-dependent protein kinase II in dendritic spines after induction of NMDA receptor-dependent chemical long-term potentiation. J Neurosci. 2004;24:9324–9331. doi: 10.1523/JNEUROSCI.2350-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owald D, Sigrist SJ. Assembling the presynaptic active zone. Curr Opin Neurobiol. 2009;19:311–318. doi: 10.1016/j.conb.2009.03.003. [DOI] [PubMed] [Google Scholar]
- Paoletti P, Bellone C, Zhou Q. NMDA receptor subunit diversity: impact on receptor properties, synaptic plasticity and disease. Nat Rev Neurosci. 2013;14:383–400. doi: 10.1038/nrn3504. [DOI] [PubMed] [Google Scholar]
- Passafaro M, Piech V, Sheng M. Subunit-specific temporal and spatial patterns of AMPA receptor exocytosis in hippocampal neurons. Nat Neurosci. 2001;4:917–926. doi: 10.1038/nn0901-917. [DOI] [PubMed] [Google Scholar]
- Patterson MA, Szatmari EM, Yasuda R. AMPA receptors are exocytosed in stimulated spines and adjacent dendrites in a Ras-ERK-dependent manner during long-term potentiation. Proc Natl Acad Sci U S A. 2010;107:15951–15956. doi: 10.1073/pnas.0913875107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peinado A, Yuste R, Katz LC. Extensive dye coupling between rat neocortical neurons during the period of circuit formation. Neuron. 1993;10:103–114. doi: 10.1016/0896-6273(93)90246-n. [DOI] [PubMed] [Google Scholar]
- Petralia RS, Al-Hallaq RA, Wenthold RJ, Van Dongen AM. Biology of the NMDA receptor. CRC Press; 2009. Trafficking and targeting of NMDA receptors. , Boca Raton. Chapter 8, ed. In. [PubMed] [Google Scholar]
- Pi HJ, Otmakhov N, El Gaamouch F, Lemelin D, De Koninck P, Lisman J. CaMKII control of spine size and synaptic strength: role of phosphorylation states and nonenzymatic action. Proc Natl Acad Sci U S A. 2010;107:14437–14442. doi: 10.1073/pnas.1009268107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plant K, Pelkey KA, Bortolotto ZA, Morita D, Terashima A, Mcbain CJ, Collingridge GL, Isaac JT. Transient incorporation of native GluR2-lacking AMPA receptors during hippocampal long-term potentiation. Nat Neurosci. 2006;9:602–604. doi: 10.1038/nn1678. [DOI] [PubMed] [Google Scholar]
- Quinlan EM, Olstein DH, Bear MF. Bidirectional, experience-dependent regulation of N-methyl-D-aspartate receptor subunit composition in the rat visual cortex during postnatal development. Proc Natl Acad Sci U S A. 1999;96:12876–12880. doi: 10.1073/pnas.96.22.12876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rauner C, Kohr G. Triheteromeric NR1/NR2A/NR2B receptors constitute the major N-methyl-D-aspartate receptor population in adult hippocampal synapses. J Biol Chem. 2011;286:7558–7566. doi: 10.1074/jbc.M110.182600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rial Verde EM, Lee-Osbourne J, Worley PF, Malinow R, Cline HT. Increased expression of the immediate-early gene ArcArg3.1 reduces AMPA receptor-mediated synaptic transmission. Neuron. 2006;52:461–474. doi: 10.1016/j.neuron.2006.09.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rivera C, Voipio J, Payne JA, Ruusuvuori E, Lahtinen H, Lamsa K, Pirvola U, Saarma M, Kaila K. The K+/Cl– co-transporter KCC2 renders GABA hyperpolarizing during neuronal maturation. Nature. 1999;397:251–255. doi: 10.1038/16697. [DOI] [PubMed] [Google Scholar]
- Rochefort NL, Garaschuk O, Milos RI, Narushima M, Marandi N, Pichler B, Kovalchuk Y, Konnerth A. Sparsification of neuronal activity in the visual cortex at eye-opening. Proc Natl Acad Sci U S A. 2009;106:15049–15054. doi: 10.1073/pnas.0907660106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roerig B, Feller MB. Neurotransmitters and gap junctions in developing neural circuits. Brain Res Brain Res Rev. 2000;32:86–114. doi: 10.1016/s0165-0173(99)00069-7. [DOI] [PubMed] [Google Scholar]
- Roisin MP, Barbin G. Differential expression of PKC isoforms in hippocampal neuronal cultures: modifications after basic FGF treatment. Neurochem Int. 1997;30:261–270. doi: 10.1016/s0197-0186(96)00095-2. [DOI] [PubMed] [Google Scholar]
- Rumpel S, LeDoux J, Zador A, Malinow R. Postsynaptic receptor trafficking underlying a form of associative learning. Science. 2005;308:83–88. doi: 10.1126/science.1103944. [DOI] [PubMed] [Google Scholar]
- Rusakov DA, Harrison E, Stewart MG. Synapses in hippocampus occupy only 1–2% of cell membranes and are spaced less than half-micron apart: a quantitative ultrastructural analysis with discussion of physiological implications. Neuropharmacology. 1998;37:513–521. doi: 10.1016/s0028-3908(98)00023-9. [DOI] [PubMed] [Google Scholar]
- Ryan TJ, Kopanitsa MV, Indersmitten T, Nithianantharajah J, Afinowi NO, Pettit C, Stanford LE, Sprengel R, Saksida LM, Bussey TJ, O’Dell TJ, Grant SG, Komiyama NH. Evolution of GluN2A/B cytoplasmic domains diversified vertebrate synaptic plasticity and behavior. Nat Neurosci. 2013;16:25–32. doi: 10.1038/nn.3277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sabatini BL, Oertner TG, Svoboda K. The life cycle of Ca2+ ions in dendritic spines. Neuron. 2002;33:439–452. doi: 10.1016/s0896-6273(02)00573-1. [DOI] [PubMed] [Google Scholar]
- Sagata N, Iwaki A, Aramaki T, Takao K, Kura S, Tsuzuki T, Kawakami R, Ito I, Kitamura T, Sugiyama H, Miyakawa T, Fukumaki Y. Comprehensive behavioural study of GluR4 knockout mice: implication in cognitive function. Genes Brain Behav. 2010;9:899–909. doi: 10.1111/j.1601-183X.2010.00629.x. [DOI] [PubMed] [Google Scholar]
- Saito N, Shirai Y. Protein kinase Cγ (PKCγ): function of neuron specific isotype. J Biochem. 2002;132:683–687. doi: 10.1093/oxfordjournals.jbchem.a003274. [DOI] [PubMed] [Google Scholar]
- Sanes DH, Kotak VC. Developmental plasticity of auditory cortical inhibitory synapses. Hear Res. 2011;279:140–148. doi: 10.1016/j.heares.2011.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanhueza M, Fernandez-Villalobos G, Stein IS, Kasumova G, Zhang P, Bayer KU, Otmakhov N, Hell JW, Lisman J. Role of the CaMKII/NMDA receptor complex in the maintenance of synaptic strength. J Neurosci. 2011;31:9170–9178. doi: 10.1523/JNEUROSCI.1250-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sans N, Petralia RS, Wang YX, Blahos J, Hell JW, Wenthold RJ. A developmental change in NMDA receptor-associated proteins at hippocampal synapses. J Neurosci. 2000;20:1260–1271. doi: 10.1523/JNEUROSCI.20-03-01260.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seeburg PH, Hartner J. Regulation of ion channel/neurotransmitter receptor function by RNA editing. Curr Opin Neurobiol. 2003;13:279–283. doi: 10.1016/s0959-4388(03)00062-x. [DOI] [PubMed] [Google Scholar]
- Seidenman KJ, Steinberg JP, Huganir R, Malinow R. Glutamate receptor subunit 2 Serine 880 phosphorylation modulates synaptic transmission and mediates plasticity in CA1 pyramidal cells. J Neurosci. 2003;23:9220–9228. doi: 10.1523/JNEUROSCI.23-27-09220.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selcher JC, Xu W, Hanson JE, Malenka RC, Madison DV. Glutamate receptor subunit GluA1 is necessary for long-term potentiation and synapse unsilencing, but not long-term depression in mouse hippocampus. Brain Res. 2012;1435:8–14. doi: 10.1016/j.brainres.2011.11.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seol GH, Ziburkus J, Huang S, Song L, Kim IT, Takamiya K, Huganir RL, Lee HK, Kirkwood A. Neuromodulators control the polarity of spike-timing-dependent synaptic plasticity. Neuron. 2007;55:919–929. doi: 10.1016/j.neuron.2007.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen K, Meyer T. Dynamic control of CaMKII translocation and localization in hippocampal neurons by NMDA receptor stimulation. Science. 1999;284:162–166. doi: 10.1126/science.284.5411.162. [DOI] [PubMed] [Google Scholar]
- Sheng M, Cummings J, Roldan LA, Jan YN, Jan LY. Changing subunit composition of heteromeric NMDA receptors during development of rat cortex. Nature. 1994;368:144–147. doi: 10.1038/368144a0. [DOI] [PubMed] [Google Scholar]
- Sheng M, Malinow R, Huganir R. Neuroscience: strength in numbers. Nature. 2013;493:482–483. doi: 10.1038/493482a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shepherd JD, Huganir RL. The cell biology of synaptic plasticity: AMPA receptor trafficking. Annu Rev Cell Dev Biol. 2007;23:613–643. doi: 10.1146/annurev.cellbio.23.090506.123516. [DOI] [PubMed] [Google Scholar]
- Shi S, Hayashi Y, Esteban JA, Malinow R. Subunit-specific rules governing AMPA receptor trafficking to synapses in hippocampal pyramidal neurons. Cell. 2001;105:331–343. doi: 10.1016/s0092-8674(01)00321-x. [DOI] [PubMed] [Google Scholar]
- Shi SH, Hayashi Y, Petralia RS, Zaman SH, Wenthold RJ, Svoboda K, Malinow R. Rapid spine delivery and redistribution of AMPA receptors after synaptic NMDA receptor activation. Science. 1999;284:1811–1816. doi: 10.1126/science.284.5421.1811. [DOI] [PubMed] [Google Scholar]
- Sia GM, Beique JC, Rumbaugh G, Cho R, Worley PF, Huganir RL. Interaction of the N-terminal domain of the AMPA receptor GluR4 subunit with the neuronal pentraxin NP1 mediates GluR4 synaptic recruitment. Neuron. 2007;55:87–102. doi: 10.1016/j.neuron.2007.06.020. [DOI] [PubMed] [Google Scholar]
- Siegel F, Heimel JA, Peters J, Lohmann C. Peripheral and central inputs shape network dynamics in the developing visual cortex in vivo. Curr Biol. 2012;22:253–258. doi: 10.1016/j.cub.2011.12.026. [DOI] [PubMed] [Google Scholar]
- Sipila ST, Schuchmann S, Voipio J, Yamada J, Kaila K. The Na–K–Cl cotransporter (NKCC1) promotes sharp waves in the neonatal rat hippocampus. J Physiol. 2006;573:765–773. doi: 10.1113/jphysiol.2006.107086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sommer B, Kohler M, Sprengel R, Seeburg PH. RNA editing in brain controls a determinant of ion flow in glutamate-gated channels. Cell. 1991;67:11–19. doi: 10.1016/0092-8674(91)90568-j. [DOI] [PubMed] [Google Scholar]
- Steenland HW, Kim SS, Zhuo M. GluR3 subunit regulates sleep, breathing and seizure generation. Eur J Neurosci. 2008;27:1166–1173. doi: 10.1111/j.1460-9568.2008.06078.x. [DOI] [PubMed] [Google Scholar]
- Steinberg JP, Takamiya K, Shen Y, Xia J, Rubio ME, Yu S, Jin W, Thomas GM, Linden DJ, Huganir RL. Targeted in vivo mutations of the AMPA receptor subunit GluR2 and its interacting protein PICK1 eliminate cerebellar long-term depression. Neuron. 2006;49:845–860. doi: 10.1016/j.neuron.2006.02.025. [DOI] [PubMed] [Google Scholar]
- Steward O, Falk PM. Selective localization of polyribosomes beneath developing synapses: a quantitative analysis of the relationships between polyribosomes and developing synapses in the hippocampus and dentate gyrus. J Comp Neurol. 1991;314:545–557. doi: 10.1002/cne.903140311. [DOI] [PubMed] [Google Scholar]
- Strack S, Colbran RJ. Autophosphorylation-dependent targeting of calcium/calmodulin-dependent protein kinase II by the NR2B subunit of the N-methyl-D-aspartate receptor. J Biol Chem. 1998;273:20689–20692. doi: 10.1074/jbc.273.33.20689. [DOI] [PubMed] [Google Scholar]
- Strack S, Kini S, Ebner FF, Wadzinski BE, Colbran RJ. Differential cellular and subcellular localization of protein phosphatase 1 isoforms in brain. J Comp Neurol. 1999;413:373–384. [PubMed] [Google Scholar]
- Strata F, Atzori M, Molnar M, Ugolini G, Tempia F, Cherubini E. A pacemaker current in dye-coupled hilar interneurons contributes to the generation of giant GABAergic potentials in developing hippocampus. J Neurosci. 1997;17:1435–1446. doi: 10.1523/JNEUROSCI.17-04-01435.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sudhof TC( Neuroligins and neurexins link synaptic function to cognitive disease. Nature. 2008;455:903–911. doi: 10.1038/nature07456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sutton MA, Schuman EM. Dendritic protein synthesis, synaptic plasticity, and memory. Cell. 2006;127:49–58. doi: 10.1016/j.cell.2006.09.014. [DOI] [PubMed] [Google Scholar]
- Syed MM, Lee S, Zheng J, Zhou ZJ. Stage-dependent dynamics and modulation of spontaneous waves in the developing rabbit retina. J Physiol. 2004;560:533–549. doi: 10.1113/jphysiol.2004.066597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi M, Tomizawa K, Ishiguro K. Distribution of tau protein kinase I/glycogen synthase kinase-3β, phosphatases 2A and 2B, and phosphorylated tau in the developing rat brain. Brain Res. 2000;857:193–206. doi: 10.1016/s0006-8993(99)02424-5. [DOI] [PubMed] [Google Scholar]
- Takahashi N, Kitamura K, Matsuo N, Mayford M, Kano M, Matsuki N, Ikegaya Y. Locally synchronized synaptic inputs. Science. 2012;335:353–356. doi: 10.1126/science.1210362. [DOI] [PubMed] [Google Scholar]
- Takahashi T, Svoboda K, Malinow R. Experience strengthening transmission by driving AMPA receptors into synapses. Science. 2003;299:1585–1588. doi: 10.1126/science.1079886. [DOI] [PubMed] [Google Scholar]
- Tanaka H, Hirano T. Visualization of subunit-specific delivery of glutamate receptors to postsynaptic membrane during hippocampal long-term potentiation. Cell Rep. 2012;1:291–298. doi: 10.1016/j.celrep.2012.02.004. [DOI] [PubMed] [Google Scholar]
- Tang YP, Shimizu E, Dube GR, Rampon C, Kerchner GA, Zhuo M, Liu G, Tsien JZ. Genetic enhancement of learning and memory in mice. Nature. 1999;401:63–69. doi: 10.1038/43432. [DOI] [PubMed] [Google Scholar]
- Tomita S, Stein V, Stocker TJ, Nicoll RA, Bredt DS. Bidirectional synaptic plasticity regulated by phosphorylation of stargazin-like TARPs. Neuron. 2005;45:269–277. doi: 10.1016/j.neuron.2005.01.009. [DOI] [PubMed] [Google Scholar]
- Turrigiano GG, Leslie KR, Desai NS, Rutherford LC, Nelson SB. Activity-dependent scaling of quantal amplitude in neocortical neurons. Nature. 1998;391:892–896. doi: 10.1038/36103. [DOI] [PubMed] [Google Scholar]
- Ultanir SK, Kim JE, Hall BJ, Deerinck T, Ellisman M, Ghosh A. Regulation of spine morphology and spine density by NMDA receptor signaling in vivo. Proc Natl Acad Sci U S A. 2007;104:19553–19558. doi: 10.1073/pnas.0704031104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanhoose AM, Winder DG. NMDA and beta1-adrenergic receptors differentially signal phosphorylation of glutamate receptor type 1 in area CA1 of hippocampus. J Neurosci. 2004;23:5827–5834. doi: 10.1523/JNEUROSCI.23-13-05827.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Versendaal D, Rajendran R, Saiepour MH, Klooster J, Smit-Rigter L, Sommeijer JP, De Zeeuw CI, Hofer SB, Heimel JA, Levelt CN. Elimination of inhibitory synapses is a major component of adult ocular dominance plasticity. Neuron. 2012;74:374–383. doi: 10.1016/j.neuron.2012.03.015. [DOI] [PubMed] [Google Scholar]
- Vicini S, Wang JF, Li JH, Zhu WJ, Wang YH, Luo JH, Wolfe BB, Grayson DR. Functional and pharmacological differences between recombinant N-methyl-D-aspartate receptors. J Neurophysiol. 1998;79:555–566. doi: 10.1152/jn.1998.79.2.555. [DOI] [PubMed] [Google Scholar]
- Vorstman JA, Ophoff RA. Genetic causes of developmental disorders. Curr Opin Neurol. 2013;26:128–136. doi: 10.1097/WCO.0b013e32835f1a30. [DOI] [PubMed] [Google Scholar]
- Wang J-H, Kelly PT. Postsynaptic calcineurin activity downregulates synaptic transmission by weakening intracellular Ca2+ signaling mechanisms in hippocampal CA1 neurons. J Neurosci. 1997;17:4600–4611. doi: 10.1523/JNEUROSCI.17-12-04600.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wenthold RJ, Petralia RS, Blahos J, II, Niedzielski AS. Evidence for multiple AMPA receptor complexes in hippocampal CA1/CA2 neurons. J Neurosci. 1996;16:1982–1989. doi: 10.1523/JNEUROSCI.16-06-01982.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yashiro K, Philpot BD. Regulation of NMDA receptor subunit expression and its implications for LTD, LTP, and metaplasticity. Neuropharmacology. 2008;55:1081–1094. doi: 10.1016/j.neuropharm.2008.07.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasuda H, Barth AL, Stellwagen D, Malenka RC. A developmental switch in the signaling cascades for LTP induction. Nat Neurosci. 2003;6:15–16. doi: 10.1038/nn985. [DOI] [PubMed] [Google Scholar]
- Yu YC, He S, Chen S, Fu Y, Brown KN, Yao XH, Ma J, Gao KP, Sosinsky GE, Huang K, Shi SH. Preferential electrical coupling regulates neocortical lineage-dependent microcircuit assembly. Nature. 2012;486:113–117. doi: 10.1038/nature10958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuste R( Dendritic spines and distributed circuits. Neuron. 2011;71:772–781. doi: 10.1016/j.neuron.2011.07.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuste R, Denk W. Dendritic spines as basic functional units of neuronal integration. Nature. 1995;375:682–684. doi: 10.1038/375682a0. [DOI] [PubMed] [Google Scholar]
- Zamanillo D, Sprengel R, Hvalby O, Jensen V, Burnashev N, Rozov A, Kaiser KM, Koster HJ, Borchardt T, Worley P, Lubke J, Frotscher M, Kelly PH, Sommer B, Andersen P, Seeburg PH, Sakmann B. Importance of AMPA receptors for hippocampal synaptic plasticity but not for spatial learning. Science. 1999;284:1805–1811. doi: 10.1126/science.284.5421.1805. [DOI] [PubMed] [Google Scholar]
- Zha XM, Green SH, Dailey ME. Regulation of hippocampal synapse remodeling by epileptiform activity. Mol Cell Neurosci. 2005;29:494–506. doi: 10.1016/j.mcn.2005.04.007. [DOI] [PubMed] [Google Scholar]
- Zhu JJ, Esteban JA, Hayashi Y, Malinow R. Postnatal synaptic potentiation: delivery of GluR4-containing AMPA receptors by spontaneous activity. Nat Neurosci. 2000;3:1098–1106. doi: 10.1038/80614. [DOI] [PubMed] [Google Scholar]
- Zito K, Scheuss V, Knott G, Hill T, Svoboda K. Rapid functional maturation of nascent dendritic spines. Neuron. 2009;61:247–258. doi: 10.1016/j.neuron.2008.10.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zoghbi HY( Postnatal neurodevelopmental disorders: meeting at the synapse. Science. 2003;302:826–830. doi: 10.1126/science.1089071. [DOI] [PubMed] [Google Scholar]