

Abstract
Respiratory syncytial virus (RSV)-induced bronchiolitis in infants is not responsive to glucocorticoids. We have shown that RSV infection impairs glucocorticoid receptor (GR) function. In this study, we have investigated the mechanism by which RSV impairs GR function. We have shown that RSV repression of GR-induced transactivation is not mediated through a soluble autocrine factor. Knock-down of mitochondrial antiviral signaling protein (MAVS), but not retinoic acid-inducible gene 1 (RIG-I) or myeloid differentiation primary response gene 88 (MyD88), impairs GR-mediated gene activation even in mock-infected cells. Over-expression of the RSV nonstructural protein NS1, but not NS2, impairs glucocorticoid-induced transactivation and viruses deleted in NS1 and/or NS2 are unable to repress glucocorticoid-induction of the known GR regulated gene glucocorticoid-inducible leucine zipper (GILZ). These data suggest that the RSV nonstructural proteins mediate RSV repression of GR-induced transactivation and that inhibition of the nonstructural proteins may be a viable target for therapy against RSV-related disease.
INTRODUCTION
Respiratory syncytial virus (RSV) is the major cause of severe lower respiratory tract infections in children, resulting in 132,000–172,000 infant hospitalizations/year in the USA (Stockman et al., 2012). Although most children survive in the developed world, there is a significant economic burden associated with RSV disease. Treatment of severe RSV symptoms in children is estimated to cost $600 billion annually in the USA (Paramore et al., 2004). In addition to children, immunocompromised adults and the elderly are also at risk from severe RSV disease (Falsey and Walsh, 2005; Raboni et al., 2003).
RSV is a single-strand negative-sense RNA pneumovirus of the Paramyxoviridae family that causes bronchiolitis, an inflammatory disease of the bronchioles. Glucocorticoids, one of the most powerful anti-inflammatory agents available, have no beneficial effect for infants with RSV-induced bronchiolitis (Buckingham et al., 2002; Bulow et al., 1999; Cade et al., 2000; Ermers et al., 2009; Loppow et al., 2001; Panickar et al., 2009; Richter and Seddon, 1998; Roosevelt et al., 1996; Somers et al., 2009). In addition, glucocorticoids show impaired suppression of RSV-induced cytokines in vitro (Bonville et al., 2001; Carpenter et al., 2002; Hinzey et al., 2011). These data suggest that RSV may have a deleterious effect on glucocorticoid signaling. In fact, we have recently shown that RSV infection represses glucocorticoid receptor (GR)-mediated gene activation (Hinzey et al., 2011).
Viral infection could interfere with host GR signaling by three potential pathways: production of autocrine factors such as cytokines; activation of other host signaling pathways; or through a direct effect of the viral proteins or RNA. RSV infection of lung epithelial cells results in the production and release of a number of cytokines and in the activation of several intracellular signaling pathways (Garofalo et al., 1996; Lindemans et al., 2006; Mastronarde et al., 1996; Singh et al., 2007; Thomas et al., 2002). In this study we investigated which of these mechanisms RSV utilizes to impair GR function. We show that the RSV nonstructural proteins mediate these repressive actions of RSV infection on GR function through inhibition of mitochrondrial antiviral signaling protein (MAVS).
MATERIALS AND METHODS
Materials
Dexamethasone was purchased from Sigma-Aldrich (St. Louis, MO) and was dissolved in 99.5% ethanol.
Cell Culture
A549 cells, an alveolar cell carcinoma derived cell line which retains features of the type II alveolar epithelial cells, are routinely used as a model for RSV infection of epithelial cells (Huang et al., 2008). A549 cells (American Type Culture Collection (ATCC), Manassas, VA) were grown in DMEM/F12 (50/50) media with 10% fetal bovine serum (FBS) and Cos7 cells (ATCC, Manassas, VA), were grown in DMEM media with 10% FBS at 37 °C and 5% CO2.
Production of NS deletion recombinant RSV expressing GFP
Recombinant RSV (rRSV) expressing enhanced green fluorescent protein (eGFP) were constructed by amplifying eGFP from pEGFP-C3 (Promega Corp., Madison, WI) by PCR using 5′ primers containing either an NheI (NS1) or Acc65I (NS2) and 3′ primers containing either a SpeI (NS1) or BsiWI (NS2) site. These eGFP fragments were digested with the appropriate enzymes and inserted into similarly digested pGEM-NSsites (Ling et al., 2008). Full-length mutant D53 plasmids were produced and recombinant RSV expressing eGFP in place of NS1 (ΔNS1e), NS2 (ΔNS2e), or both NS1 and NS2 (ΔNS1/2e) were recovered as described (Tran et al., 2007).
Virus Preparation
Recombinant green fluorescent protein (GFP)-expressing RSV (rgRSV) was grown in HeLa cells, separated from debris by low speed centrifugation and further purified by pelleting in a high speed centrifuge (Hallak et al., 2000). Specifically, monolayer HeLa cells were inoculated with rgRSV for 2 h. Media was removed and fresh media added. Two days later cells were harvested, virus detached by vortexing and cells removed by low speed centrifugation at 1,200 rpm for 5 min. Virus was further purified by high speed centrifugation at 20,000 x g for 1.5 h. Viral pellets were resuspended in HBSS and snap frozen Mock-infected HeLa-conditioned media was produced by treating the cells in the same manner but no RSV was used to initially infect the cells. Viral titer was determined by a fluorescent titration assay as previously described (Hinzey et al., 2011). Virus stocks of recombinant wild-type RSV (rA2) and NS-deletion viruses were produced in Vero cells and titered by plaque assay as described (Tran et al., 2007).
UV-irradiation of RSV
Viral replication was inhibited by UV irradiation. The virus, rgRSV was diluted in media without FBS, distributed in 800 μl aliquots in an open 12-well cell culture plate and UV-irradiated at 184 μJ/cm2 for 20 min. Success of UV-irradiation was confirmed by the failure to detect GFP in cells inoculated with the UV-rgRSV.
Viral infection of A549 cells
For viral adsorption, media was removed from A549 cells and fresh media without FBS containing RSV at the required multiplicity of infection (MOI) was added to the cells for 1 h. The media was then replaced with fresh media containing 10% serum and treated as indicated.
Production of RSV conditioned media
A549 cells were inoculated with rgRSV (MOI=1) or mock-infected with HeLa-conditioned media for 1 h. The media was aspirated and fresh media supplied. After 2 days supernatants were collected from mock-infected cells or RSV-infected cells, centrifuged at 12,000 x g for 5 min and then either applied directly or after UV-irradiation to uninfected A549 cells.
Transient Transfection
Cos7 cells were plated in 24-well cell culture plates in DMEM containing 10% charcoal stripped serum (CSS) at a concentration of 5 × 104 cells/well. The next day cells were transfected with 20 ng hGR, 100 ng of the GR-responsive promoter, MMTVLuc, 20 ng of the constitutively active renilla internal control, pRL-TK (Promega Corp., Madison, WI), HA- or FLAG-tagged NS1 and/or NS2 or the empty plasmid SB897 and pSG5 (empty plasmid) using TransIT-LT1 (Mirus Bio, Madison, WI) according to the manufacturer’s instructions. The following day the cells were treated with vehicle or 100 nM dexamethasone for 5 h, the cells were lysed and luciferase measured using the Dual Luciferase Assay (Promega Corp., Madison, WI) on a Packard LumiCount luminometer (PerkinElmer, Waltham, MA).
Real-time PCR
Total RNA was extracted using TRIzol (Life technologies, Grand Island, NY) according to the manufacturer’s instructions. 500 ng RNA was reverse transcribed using the High Capacity cDNA Reverse Transcription Kit with RNase inhibitor (Life Technologies, Grand Island, NY) in a 20 μl reaction. cDNA was then diluted by addition of 100 μl DEPC-treated H2O. Real-time PCR was performed on 8 μl diluted cDNA in a 20 μl reaction containing 1X Power SYBR® Green PCR Master Mix (Life technologies, Grand Island, NY) and 0.125 μM of each primer ((Hinzey et al., 2011) and Table 1) using the following protocol on an ABI 7300 Real-Time PCR system. Plates were heated at 50 °C for 2 min, denatured for 10 min at 95°C and subjected to 40 cycles of 95 °C for 15 sec and 60 °C for 1 min. Cycle threshold (CT) values of duplicate samples were analyzed using the comparative CT (ΔΔCT) method (Life technologies, Grand Island, NY). The fold induction (2−ΔΔCt) by dexamethasone was obtained by normalizing to the average of two endogenous genes, GAPDH (glyceraldehyde-3-phosphate dehydrogenase) and CAP-1 (Cyclic AMP-accessory protein) (Gavrilin et al., 2006), and expressed relative to the amount in non-treated cells. Relative copy numbers (RCN) were calculated as 2−ΔCt × 100 (Gavrilin et al., 2006).
TABLE 1.
Real-time PCR primers.
| Gene name | Accession number | Direction | Primer sequence (5′–3′) | Reference |
|---|---|---|---|---|
| CAP-1 | NM_006367.3 | F R |
ATTCCCTGGATTGTGAAATAGTC ATTAAAGTCACCGCCTTCTGTAG |
(26) |
| GAPDH | NM_002046.3 | F R |
ACTTTGGTATCGTGGAAGGACT GTAGAGGCAGGGATGATGTTCT |
(26) |
| GILZ | NM_004089 | F R |
GCACAATTTCTCCATCTCCTTCTT TCAGCATGATTCTTCACCAGATCCA |
(16) |
| IL-8 | NM_000584.2 | F R |
AGTTTTTGAAGAGGGCTGAGAAT CAACAGACCCACACAATACATGA |
(16) |
| MAVS | NM_020746.4 | F R |
AGACCAGGATCGACTGCGGGC AGAGGCCACTTCGTCCGCGA |
|
| MKP-1 | NM_004417 | F R |
CTGCCTTGATCAACGTCTCA ACCCTTCCTCCAGCATTCTT |
(16) |
| MyD88 | NM_001172567.1 | F R |
GAGCTGGCGGGCATCAC TCGAAACGCTCAGGCATATG |
|
| RIG-I | NM_014314.3 | F R |
ACCAGAGCACTTGTGGACGCT TGCCGGGAGGGTCATTCCTGT |
Western blotting
Total cellular protein was isolated using M-PER in the presence of protease inhibitors (Thermo Fisher Scientific Inc., Pittsburgh, PA) according to the manufacturer’s instructions. 10 μg protein was subjected to SDS-PAGE on NuPAGE Novex 4–12% bis-tris pre-cast gels (Life Technologies, Grand Island, NY). Proteins were then transferred to nitrocellulose membranes by semi-dry blotting and blocked in 5% non-fat dry milk in Tris-buffered saline – 0.05% Tween 20 (TBST). They were then incubated with primary antibody raised against the HA-tag (Cell Signaling Technology Inc., Danver, MA; 3742; 1:1000) or the FLAG-tag (Cell Signaling Technology Inc., Danver, MA; 2368; 1:1000) in 5% milk in TBST overnight at 4°C. Membranes were then washed three times with TBST and incubated with goat anti-rabbit IgG-HRP (Santa Cruz Biotechnology, Santa Cruz, CA; sc-2004; 1:2000) in TBST for 1 h at room temperature. Membranes were again washed with TBST and chemiluminescence detected using SuperSignal West Pico Chemiluminscence Substrate (Thermo Fisher Scientific Inc., Pittsburgh, PA) according to the manufacturer’s instructions and exposed to autoradiographic film. Membranes were stripped for 15 min at room temperature in Restore Stripping buffer (Thermo Fisher Scientific Inc., Pittsburgh, PA) and re-probed using a b-actin primary antibody (Santa Cruz Biotechnology, Santa Cruz, CA; sc-47778; 1:2000) and goat anti-mouse IgG-HRP secondary antibody (Santa Cruz Biotechnology, Santa Cruz, CA; sc-2005; 1:5000).
siRNA-mediated gene silencing
50 nM siRNA to myeloid differentiation primary response gene 88 (MyD88), retinoic acid-inducible gene 1 (RIG-I), MAVS or control siRNA-A (Santa Cruz Biotechnology, Santa Cruz, CA; sc-35986, sc-61480, sc-75755 and sc-37007 respectively) was transfected into A549 cells using DharmaFECT 2 (Thermo Fisher Scientific Inc., Pittsburgh, PA, USA) according to the manufacturer’s instructions. 48 h post transfection the cells were inoculated with rgRSV at an MOI of 3 as described above. After 1 h the inoculum was removed and cells treated with vehicle or 100 nM dexamethasone and the expression of GR-regulated gene, glucocorticoid-inducible leucine zipper (GILZ), determined by real-time PCR. The efficiency of siRNA silencing was determined by real-time PCR using primers in Table 1.
Statistical Analysis
Transfection experiments were performed in triplicate and real-time experiments in duplicate and the average was treated as one value. The average values of n independent experiments were analyzed for statistical significance using the two-tailed unpaired Student’s t-test or an ANOVA and appropriate post hoc test using Prism 5 software (Graphpad Software Inc., La Jolla, CA).
RESULTS
Effect of soluble secreted factors on GR-mediated transactivation
RSV induces the expression of soluble cytokines, and cytokines have been shown to play a role in glucocorticoid insensitivity (Hu et al., 2009; Irusen et al., 2002; Ishiguro, 1999; Leung et al., 1995; Matthews et al., 2004; Pace et al., 2011; Sher et al., 1994; Tliba et al., 2008). Therefore, we tested the effect of conditioned media produced from RSV- or mock-infected A549 cells on the ability of dexamethasone to induce a known GR-regulated gene. Analysis of the conditioned media from RSV-infected cells showed increased levels of 27 cytokines, including RANTES, interleukin 6 (IL-6), IL-8 and IL-1α that were not present in conditioned media from mock-infected cells (data not shown). Conditioned media from RSV-infected cells, which also contains live virus, was either applied directly to uninfected cells or UV-irradiated, to inactivate live virus, prior to addition to the uninfected cells. Both of these treatments increased the expression of IL-8 mRNA whereas conditioned media from mock-infected cells did not (Fig. 1A). Dexamethasone induction of GILZ (Fig. 1B) or mitogen-activated protein (MAP) kinase phosphatase 1 (MKP-1) (Fig. 1C) was similar in cells treated with conditioned media from mock-infected cells or from RSV-infected cells, either untreated or UV-irradiated.
FIG. 1.
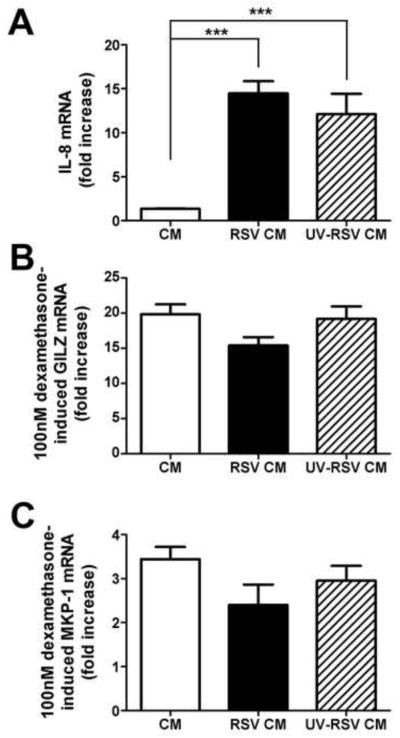
RSV repression of GR-induced genes is not mediated by a soluble secreted factor. Conditioned media from mock-infected cells (CM) (open bar), RSV-infected cells (RSV CM) (solid bar) or UV-irradiated conditioned media from RSV-infected cells (UV-RSV CM) (hatched bar) was added to uninfected A549 cells. After 5 hours vehicle or dexamethasone was added such that the final concentration was 100 nM and cells were incubated for a further 5 h. A. IL-8 mRNA was detected in vehicle only treated cells by real-time PCR, normalized to GAPDH and CAP-1 and compared to untreated cells. Dexamethasone-induced GILZ (B) and MKP-1 (C) mRNA was determined by real-time PCR and normalized to GAPDH and CAP-1. Means and SD are shown (n= 5). Significance was tested using a 1-way ANOVA and Dunnett post hoc test (*** depicts p<0.001).
Role of intracellular signaling pathways in RSV suppression of GR-mediated transactivation
RSV is known to activate a number of intracellular signaling pathways (Garofalo et al., 1996; Lindemans et al., 2006; Mastronarde et al., 1996; Singh et al., 2007; Thomas et al., 2002), therefore, the role of these pathways in RSV-mediated suppression of glucocorticoid induction of a GR-regulated gene was investigated. Specifically, RSV is known to stimulate the Toll-like receptor (TLR) and RIG-like helicase (RLR) pathways (Haeberle et al., 2002; Kurt-Jones et al., 2000; Liu et al., 2007; Liu et al., 2008; Lukacs et al., 2010; Murawski et al., 2009; Segovia et al., 2012). MyD88, the common adaptor molecule of the Toll-like receptors (TLRs), and RIG-I were knocked down by siRNA. RSV infection of A549 cells transfected with siRNAs against MyD88, RIG-I or a control siRNA repressed dexamethasone induction of GILZ to similar levels compared to mock-infected cells (Fig. 2A). Dexamethasone induction of GILZ was slightly higher in mock-infected cells transfected with RIG-I siRNA compared to control siRNA. Knock-down of individual gene expression was confirmed by real-time PCR (Fig. 2B & C).
FIG 2.

Knock-down of MyD88 or RIG-I does not reverse RSV repression of dexamethasone-induced GILZ mRNA. A549 cells were transfected with control siRNA or siRNA against MyD88 or RIG-I. 48 h later they were inoculated with rgRSV (MOI=3) (solid bar) or mock-infected with HeLa-conditioned media (open bar) for 1 h. The media was aspirated and cells treated with vehicle or 100 nM dexamethasone for 5 h. Dexamethasone-induced GILZ mRNA was determined by real-time PCR and normalized to GAPDH and CAP-1 (A). siRNA knockdown was confirmed by real-time PCR (B & C). Means and SD are shown (n= 3). Significance was tested using a 1-way ANOVA and Tukey post hoc test (** depicts p = 0.01-0.001; *** depicts p<0.001).
Knock-down of MAVS, the common adaptor of the RIG-I-like receptors (RLR), reduced dexamethasone induction of GILZ in mock-infected cells compared to control siRNA (Fig. 3A). Glucocorticoid induction of GILZ in cells transfected with MAVS siRNA was further reduced by RSV infection, though this slight effect was not statistically significant. As above, efficiency of siRNA knock-down was confirmed by real-time PCR (Fig. 3B).
FIG 3.

Knock-down of MAVS reduces glucocorticoid induction of GILZ and eliminates RSV repression of glucocorticoid-induced GILZ. A549 cells were transfected with control siRNA or siRNA against MAVS. 48 h later they were inoculated with rgRSV (MOI=3) (solid bar) or mock-infected with HeLa-conditioned media (open bar) for 1 h. The media was aspirated and cells treated with vehicle or 100 nM dexamethasone for 5 h. Dexamethasone-induced GILZ mRNA was determined by real-time PCR and normalized to GAPDH and CAP-1 (A). siRNA knockdown was confirmed by real-time PCR (B). Means and SD are shown (n= 3). Significance was tested using a 1-way ANOVA and Tukey post hoc test (* depicts p = 0.05-0.01; ** depicts p = 0.01-0.001).
Effect of the RSV nonstructural proteins on GR-mediated transactivation
The RSV nonstructural proteins have been shown to inhibit interferon induction by direct interaction with components of the RLR pathway (Boyapalle et al., 2012; Ling et al., 2009), Since siRNA knockdown of MAVS abrogated RSV-mediated inhibition of GR-induced GILZ expression, we tested whether the RSV NS proteins were responsible for the effect of RSV on GR-mediated transactivation. Over-expression of either HA- (Fig. 4A) or FLAG-tagged (Fig. 4B) NS1 repressed dexamethasone induction of the glucocorticoid-responsive promoter MMTVLuc when expressed by transient transfection in Cos7 cells. Over-expression of NS2 or the empty vector had no effect of dexamethasone-induced MMTVLuc activity. Co-expression of NS1 and NS2 repressed dexamethasone-induced MMTVLuc activity similarly to NS1 alone. Expression was confirmed by western blotting using antibodies directed against the HA- or FLAG-tags (Fig. 4C and D). The HA-tagged NS1 was not detected by the HA-tag antibody although there is clearly an effect of transfection of this construct.
FIG 4.

Over-expression of NS1 represses glucocorticoid-induced MMTVLuc activity. Cos7 cells were transfected with hGR, the GR-responsive MMTVLuc promoter; a constitutively active control renilla vector, and either a HA-(A and C) or FLAG-tagged (B and D) NS1 or NS2 expression vector or the empty vector SB897. Cells were treated with vehicle or 100 nM dexamethasone for 5 h. Firefly luciferase was normalized to renilla luciferase and fold increase following dexamethasone treatment was calculated. Means and SD are shown (n= 3). Significance was tested using a 1-way ANOVA and Dunnett post hoc test. a; p = 0.05-0.01 when compared to 5 ng SB897: b; p = 0.05-0.01 when compared to 10 ng SB897; c; p < 0.001 when compared to 5 ng SB897; d; p < 0.001 when compared to 10 ng SB897. Cell lysates were taken for western blotting using antibodies directed against the HA- (C) or FLAG-tags (D). The blots were stripped and re-probed for β-actin. A representative figure of two repeats is shown.
To confirm this effect in the context of viral infection, we infected A549 cells with either wild-type recombinant RSV (rA2) or recombinant RSV that lack NS1 (ΔNS1e), NS2 (ΔNS2e), or both NS1 and NS2 (ΔNS1/2e). These recombinant viruses contain GFP inserted in place of the deleted genes. Infection of A549 cells with wild-type rA2 virus repressed dexamethasone-induced GILZ whereas infection with RSV lacking NS1 (ΔNS1e and ΔNS1/2e) had no effect of glucocorticoid-induced GILZ (Fig. 5). Unexpectedly, infection with ΔNS2e also did not inhibit GILZ expression after dexamethasone treatment. Real-time PCR confirmed the expected expression patterns of the NS genes in cells infected with rA2 or the NS-deletion recombinant RSVs. In addition, viral genome levels were not affected by 5 h dexamethasone treatment and were relatively similar among the cells infected by the NS-deletion and wild type rRSV, indicating equal input infection and a lack of effect of dexamethasone on RSV infection (data not shown). To examine transcription, we determined F mRNA levels, since F is downstream of the NS genes and should be less affected by changes in the transcription gradient due to NS gene alteration than genes that are more promoter-proximal. We found that F RNA levels were unaffected by dexamethasone treatment, similar to the viral genome levels, and expressed to comparable levels in rA2-, ΔNS1, and ΔNS1/2-infected cells (data not shown). However, F RNA was present at higher levels in ΔNS2-infected cells, though this expression did not appear to affect GR activation.
FIG 5.

RSV deleted in the NS1 and/or NS2 genes does not repress glucocorticoid-induced GILZ. A549 cells were inoculated with rA2, ΔNS1e, ΔNS2e, ΔNS12e (MOI=3) or mock-infected with HeLa-conditioned media for 1 h. The media was changed and cells were treated with 100 nM dexamethasone for 5 h. Dexamethasone-induced GILZ mRNA was determined by real-time PCR and normalized to GAPDH and CAP-1 (A). NS1 (open bar) and NS2 (solid bar) expression was confirmed by real-time PCR (B). Means and SD are shown (n= 3). Significance was tested using a 1-way ANOVA and Dunnett post hoc test (* depicts p = 0.05-0.01; ** depicts p = 0.01-0.001).
DISCUSSION
We previously showed that RSV infection represses glucocorticoid-induced gene expression (Hinzey et al., 2011). In this study, we examined the mechanisms by which RSV accomplishes this effect. RSV infection could impair GR function by three potential mechanisms: by production of secreted autocrine factors; by activation of intracellular signaling pathways; or by a direct protein/RNA effect. RSV-infected lung epithelial cells produce a number of cytokines. IL-1α has been previously shown to function in an autocrine fashion to mediate RSV-induction of IL-6, IL-8 and intracellular adhesion molecule 1 (ICAM-1) (Jiang et al., 1998; Patel et al., 1998; Patel et al., 1995). IL-10 acts in an autocrine manner to reduce inflammation by inhibiting T-cell responses (Loebbermann et al., 2012; Sun et al., 2011). The role of cytokines in glucocorticoid insensitivity/resistance has been previously described. Cytokines, such as IL-2/IL-4, IL-13 tumor necrosis factor α (TNFα), IL-6, IL-8, interferon γ (IFNγ) and IFNα have been shown to impair GR function and are proposed to be involved in glucocorticoid resistance/insensitivity in diseases such as asthma, ulcerative colitis, and depression (Hu et al., 2009; Irusen et al., 2002; Ishiguro, 1999; Leung et al., 1995; Matthews et al., 2004; Pace et al., 2011; Sher et al., 1994; Tliba et al., 2008). Thus, the hypothesis that RSV-induced cytokines could impair GR function was tested. We have shown that RSV-conditioned media-induced IL-8 transcription as previously described (Patel et al., 1998). However, this RSV-conditioned media did not reduce glucocorticoid induction of the known GR-regulated genes GILZ and MKP-1 (Fig. 1), suggesting that the repression of GR function by RSV infection is not mediated by a soluble autocrine factor.
RSV infection activates a number of intracellular signaling pathways, including Nuclear factor κ B (NFκB), phosphoinositide 3-kinase (PI3K), and MAP kinase pathways (Garofalo et al., 1996; Lindemans et al., 2006; Mastronarde et al., 1996; Singh et al., 2007; Thomas et al., 2002). Activation of such pathways could result in glucocorticoid insensitivity/resistance as GR is known to cross-talk with and mutually antagonize these pathways (Adcock and Lane, 2003; Ayroldi et al., 2012; Irusen et al., 2002; Marwick et al., 2009; Ray and Prefontaine, 1994). In our earlier work, we showed that RSV repression of GR function was not mediated though the NFκB pathway (Hinzey et al., 2011). In order to investigate this further we inhibited the activation of multiple signaling pathways by interfering with the initiation of these signaling cascades. RSV is sensed through pathogen recognition receptors in the TLR (Haeberle et al., 2002; Kurt-Jones et al., 2000; Lukacs et al., 2010; Murawski et al., 2009; Segovia et al., 2012) and RLR pathways (Liu et al., 2007; Liu et al., 2008), resulting in the activation of type I interferon responses. SiRNA knock-down of MyD88, the common adapter of the TLR pathways, or RIG-I did not impair RSV repression of glucocorticoid-induced GILZ mRNA (Fig. 2), suggesting that activation of these receptors does not mediate RSV repression of GR function. However, we cannot fully rule out TLR3 as some RSV effects mediated through TLR3 are MyD88 independent (Rudd et al., 2005).
Knock-down of MAVS, the downstream adapter molecule of RIG-I, repressed glucocorticoid-induction of GILZ even in mock-infected cells (Fig. 3). In the presence of RSV infection glucocorticoid-induction of GILZ was further repressed. Even though this was not significantly significant we cannot rule out biological significance due to the reduced GILZ induction in the mock-infected cells. RSV expresses two small nonstructural proteins, NS1 and NS2, that suppress type I interferon response in part by inhibiting the RIG-I-MAVS interaction (Boyapalle et al., 2012; Ling et al., 2009). Therefore, we tested if NS1 and NS2 could mediate the effect of RSV on GR function. First, we used a transient transfection system where we over-expressed HA- or FLAG-tagged NS1 and/or NS2. In this system, over-expression of NS1, but not NS2, repressed glucocorticoid induction of the glucocorticoid responsive MMTVLuc promoter. Co-expression of NS1 and NS2 repressed glucocorticoid-induced MMTVLuc activity similarly to NS1 alone (Fig. 4). To confirm these findings, we used viruses with deletions in either the NS1 or NS2 genes or both. Deletion of NS1, alone or in combination with NS2, prevented RSV suppression of glucocorticoid-induced GILZ (Fig. 5). Unexpectedly, deletion of NS2 also abrogated the RSV effect on glucocorticoid responsiveness. These data show that the nonstructural proteins mediate the RSV suppression of GR function.
It is not entirely clear why only NS1 was able to repress glucocorticoid-induced MMTVLuc activity in the transient transfection system whereas deletion of either NS1 or NS2 from the virus reversed the repression of glucocorticoid-induced GILZ. It is possible that this is an artifact of the transient transfection system. Certainly differences in subcellular localization of NS1 and NS2 following transient transfection, compared to infection, have been noted. NS2 is found in the mitochondria and cytoplasm when expressed on its own; NS1 is found to be in the nucleus and in mitochondria either when expressed alone or when co-expressed with NS2 (Boyapalle et al., 2012; Swedan et al., 2011; Tan et al., 2013).
Direct impact of a viral component (protein or RNA) on GR function is not unknown. The Vpr protein of HIV binds directly to GR, functioning as a coactivator to up-regulate GR (Kino et al., 1999; Kino et al., 2002; Sherman et al., 2000). The Poxvirus molluscum contagiosum (MC) protein MC013L binds and inhibits GR (Chen et al., 2000) and a noncoding RNA has also been shown to impair GR function (Kino et al., 2010). Our data suggest that the RSV nonstructural proteins similarly impair GR function. Further studies are ongoing to determine if the effects of the nonstructural proteins are mediated through direct protein-protein interactions or if they are due to the inhibition of the type I interferon pathway. Repression of the interferon pathway by glucocorticoids is known (Diez et al., 2012; Flammer et al., 2010; McCoy et al., 2008) but there is no literature to suggest that the interferon pathway can repress GR function. In addition, other viruses, such as pneumonia virus of mice, Sendai virus, influenza, dengue virus and Ebola virus, have proteins similar to the RSV nonstructural proteins that inhibit the interferon response (Aguirre et al., 2012; Buchholz et al., 2009; Heinze et al., 2011; Mibayashi et al., 2007; Prins et al., 2009; Strahle et al., 2007). It has yet to be determined if these proteins from other viruses also impair GR function.
To date there is no vaccine for RSV and early attempts with a formalin-inactivated vaccine proved fatal. Interestingly, viruses deleted in NS1 and/or NS2 are novel candidates for vaccine development (Wright et al., 2006). Although this is primarily due to their limited virulence it is possible that the loss of the glucocorticoid suppressive action may also help in the production of a useful vaccine. In addition, inhibition of the nonstructural proteins may be a viable target for therapy against RSV-related disease. Inhibition of these proteins would not only enhance host anti-viral interferon responses but would also enhance glucocorticoid responsiveness.
RSV repression of GR function is not mediated by a soluble autocrine factor.
Knock-down of MAVS represses GR-mediated gene activation in mock-infected cells.
Over-expression of RSV NS1 represses GR-mediated transactivation.
Loss of NS proteins eliminates the effect of RSV infection on GR-mediated gene activation.
Acknowledgments
This study was supported by NIAID (R01AI081977, MNT) and the Joy McCann Culverhouse Foundation (MNT). We would like to thank Christopher Burnsides, Emaline Wise and Amanda Selhorst for technical assistance. rgRSV was kindly provided by Mark Peeples (The Research Institute at Nationwide Children’s Hospital, Department of Pediatrics, OSU, Columbus, OH) and by Peter Collins (National Institute of Allergy and Infectious Diseases, NIH, Bethesda, MD). We also thank Peter Collins for the use of the RSV reverse genetics system. Expression plasmids for hGR and the MMTVLuc reporter plasmid were provided by S. Stoney Simons Jr. (National Institute of Diabetes and Digestive and Kidney Diseases, NIH, Bethesda, MD). Expression plasmids for HA- and FLAG-tagged NS1 and NS2 were provided by Sailen Barik (Cleveland State University, Cleveland, OH).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Adcock IM, Lane SJ. Corticosteroid-insensitive asthma: Molecular mechanisms. J Endocrinol. 2003;178:347–355. doi: 10.1677/joe.0.1780347. [DOI] [PubMed] [Google Scholar]
- Aguirre S, Maestre AM, Pagni S, Patel JR, Savage T, Gutman D, Maringer K, Bernal-Rubio D, Shabman RS, Simon V, Rodriguez-Madoz JR, Mulder LC, Barber GN, Fernandez-Sesma A. DENV inhibits type I IFN production in infected cells by cleaving human STING. PLoS Pathog. 2012;8:e1002934. doi: 10.1371/journal.ppat.1002934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayroldi E, Cannarile L, Migliorati G, Nocentini G, Delfino DV, Riccardi C. Mechanisms of the anti-inflammatory effects of glucocorticoids: Genomic and nongenomic interference with MAPK signaling pathways. FASEB J. 2012;26:4805–4820. doi: 10.1096/fj.12-216382. [DOI] [PubMed] [Google Scholar]
- Bonville CA, Mehta PA, Krilov LR, Rosenberg HF, Domachowske JB. Epithelial cells infected with respiratory syncytial virus are resistant to the anti-inflammatory effects of hydrocortisone. Cell Immunol. 2001;213:134–140. doi: 10.1006/cimm.2001.1869. [DOI] [PubMed] [Google Scholar]
- Boyapalle S, Wong T, Garay J, Teng M, San Juan-Vergara H, Mohapatra S, Mohapatra S. Respiratory syncytial virus NS1 protein colocalizes with mitochondrial antiviral signaling protein MAVS following infection. PLoS One. 2012;7:e29386. doi: 10.1371/journal.pone.0029386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buchholz UJ, Ward JM, Lamirande EW, Heinze B, Krempl CD, Collins PL. Deletion of nonstructural proteins NS1 and NS2 from pneumonia virus of mice attenuates viral replication and reduces pulmonary cytokine expression and disease. J Virol. 2009;83:1969–1980. doi: 10.1128/JVI.02041-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckingham SC, Jafri HS, Bush AJ, Carubelli CM, Sheeran P, Hardy RD, Ottolini MG, Ramilo O, DeVincenzo JP. A randomized, double-blind, placebo-controlled trial of dexamethasone in severe respiratory syncytial virus (RSV) infection: Effects on RSV quantity and clinical outcome. J Infect Dis. 2002;185:1222–1228. doi: 10.1086/340024. [DOI] [PubMed] [Google Scholar]
- Bulow SM, Nir M, Levin E, Friis B, Thomsen LL, Nielsen JE, Holm JC, Moller T, Bonde-Hansen ME, Nielsen HE. Prednisolone treatment of respiratory syncytial virus infection: a randomized controlled trial of 147 infants. Pediatrics. 1999;104:e77. doi: 10.1542/peds.104.6.e77. [DOI] [PubMed] [Google Scholar]
- Cade A, Brownlee KG, Conway SP, Haigh D, Short A, Brown J, Dassu D, Mason SA, Phillips A, Eglin R, Graham M, Chetcuti A, Chatrath M, Hudson N, Thomas A, Chetcuti PA. Randomised placebo controlled trial of nebulised corticosteroids in acute respiratory syncytial viral bronchiolitis. Arch Dis Child. 2000;82:126–130. doi: 10.1136/adc.82.2.126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carpenter LR, Moy JN, Roebuck KA. Respiratory syncytial virus and TNF alpha induction of chemokine gene expression involves differential activation of Rel A and NF-kappa B1. BMC Infect Dis. 2002;2:5. doi: 10.1186/1471-2334-2-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen N, Baudino T, MacDonald PN, Green M, Kelley WL, Burnett JW, Buller RM. Selective inhibition of nuclear steroid receptor function by a protein from a human tumorigenic poxvirus. Virology. 2000;274:17–25. doi: 10.1006/viro.2000.0410. [DOI] [PubMed] [Google Scholar]
- Diez D, Goto S, Fahy JV, Erle DJ, Woodruff PG, Wheelock AM, Wheelock CE. Network analysis identifies a putative role for the PPAR and type 1 interferon pathways in glucocorticoid actions in asthmatics. BMC Med Genomics. 2012;5:27. doi: 10.1186/1755-8794-5-27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ermers MJ, Rovers MM, van Woensel JB, Kimpen JL, Bont LJ. The effect of high dose inhaled corticosteroids on wheeze in infants after respiratory syncytial virus infection: Randomised double blind placebo controlled trial. BMJ. 2009;338:b897. doi: 10.1136/bmj.b897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falsey AR, Walsh EE. Respiratory syncytial virus infection in elderly adults. Drugs Aging. 2005;22:577–587. doi: 10.2165/00002512-200522070-00004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flammer JR, Dobrovolna J, Kennedy MA, Chinenov Y, Glass CK, Ivashkiv LB, Rogatsky I. The type I interferon signaling pathway is a target for glucocorticoid inhibition. Mol Cell Biol. 2010;30:4564–4574. doi: 10.1128/MCB.00146-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garofalo R, Sabry M, Jamaluddin M, Yu RK, Casola A, Ogra PL, Brasier AR. Transcriptional activation of the interleukin-8 gene by respiratory syncytial virus infection in alveolar epithelial cells: Nuclear translocation of the RelA transcription factor as a mechanism producing airway mucosal inflammation. J Virol. 1996;70:8773–8781. doi: 10.1128/jvi.70.12.8773-8781.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gavrilin MA, Bouakl IJ, Knatz NL, Duncan MD, Hall MW, Gunn JS, Wewers MD. Internalization and phagosome escape required for Francisella to induce human monocyte IL-1beta processing and release. Proc Natl Acad Sci U S A. 2006;103:141–146. doi: 10.1073/pnas.0504271103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haeberle HA, Takizawa R, Casola A, Brasier AR, Dieterich HJ, Van Rooijen N, Gatalica Z, Garofalo RP. Respiratory syncytial virus-induced activation of nuclear factor-kappaB in the lung involves alveolar macrophages and toll-like receptor 4-dependent pathways. J Infect Dis. 2002;186:1199–1206. doi: 10.1086/344644. [DOI] [PubMed] [Google Scholar]
- Hallak LK, Spillmann D, Collins PL, Peeples ME. Glycosaminoglycan sulfation requirements for respiratory syncytial virus infection. J Virol. 2000;74:10508–10513. doi: 10.1128/jvi.74.22.10508-10513.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heinze B, Frey S, Mordstein M, Schmitt-Graff A, Ehl S, Buchholz UJ, Collins PL, Staeheli P, Krempl CD. Both nonstructural proteins NS1 and NS2 of pneumonia virus of mice are inhibitors of the interferon type I and type III responses in vivo. J Virol. 2011;85:4071–4084. doi: 10.1128/JVI.01365-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hinzey A, Alexander J, Corry J, Adams KM, Claggett AM, Traylor ZP, Davis IC, Webster Marketon JI. Respiratory syncytial virus represses glucocorticoid receptor-mediated gene activation. Endocrinology. 2011;152:483–494. doi: 10.1210/en.2010-0774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu F, Pace TW, Miller AH. Interferon-alpha inhibits glucocorticoid receptor-mediated gene transcription via STAT5 activation in mouse HT22 cells. Brain Behav Immun. 2009;23:455–463. doi: 10.1016/j.bbi.2009.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang YC, Li Z, Hyseni X, Schmitt M, Devlin RB, Karoly ED, Soukup JM. Identification of gene biomarkers for respiratory syncytial virus infection in a bronchial epithelial cell line. Genomic Med. 2008;2:113–125. doi: 10.1007/s11568-009-9080-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Irusen E, Matthews JG, Takahashi A, Barnes PJ, Chung KF, Adcock IM. p38 Mitogen-activated protein kinase-induced glucocorticoid receptor phosphorylation reduces its activity: Role in steroid-insensitive asthma. J Allergy Clin Immunol. 2002;109:649–657. doi: 10.1067/mai.2002.122465. [DOI] [PubMed] [Google Scholar]
- Ishiguro Y. Mucosal proinflammatory cytokine production correlates with endoscopic activity of ulcerative colitis. J Gastroenterol. 1999;34:66–74. doi: 10.1007/s005350050218. [DOI] [PubMed] [Google Scholar]
- Jiang Z, Kunimoto M, Patel JA. Autocrine regulation and experimental modulation of interleukin-6 expression by human pulmonary epithelial cells infected with respiratory syncytial virus. J Virol. 1998;72:2496–2499. doi: 10.1128/jvi.72.3.2496-2499.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kino T, Gragerov A, Kopp JB, Stauber RH, Pavlakis GN, Chrousos GP. The HIV-1 virion-associated protein vpr is a coactivator of the human glucocorticoid receptor. J Exp Med. 1999;189:51–62. doi: 10.1084/jem.189.1.51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kino T, Gragerov A, Slobodskaya O, Tsopanomichalou M, Chrousos GP, Pavlakis GN. Human immunodeficiency virus type 1 (HIV-1) accessory protein Vpr induces transcription of the HIV-1 and glucocorticoid-responsive promoters by binding directly to p300/CBP coactivators. J Virol. 2002;76:9724–9734. doi: 10.1128/JVI.76.19.9724-9734.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kino T, Hurt DE, Ichijo T, Nader N, Chrousos GP. Noncoding RNA gas5 is a growth arrest- and starvation-associated repressor of the glucocorticoid receptor. Sci Signal. 2010;3:ra8. doi: 10.1126/scisignal.2000568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurt-Jones EA, Popova L, Kwinn L, Haynes LM, Jones LP, Tripp RA, Walsh EE, Freeman MW, Golenbock DT, Anderson LJ, Finberg RW. Pattern recognition receptors TLR4 and CD14 mediate response to respiratory syncytial virus. Nat Immunol. 2000;1:398–401. doi: 10.1038/80833. [DOI] [PubMed] [Google Scholar]
- Leung DY, Martin RJ, Szefler SJ, Sher ER, Ying S, Kay AB, Hamid Q. Dysregulation of interleukin 4, interleukin 5, and interferon gamma gene expression in steroid-resistant asthma. J Exp Med. 1995;181:33–40. doi: 10.1084/jem.181.1.33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindemans CA, Coffer PJ, Schellens IM, de Graaff PM, Kimpen JL, Koenderman L. Respiratory syncytial virus inhibits granulocyte apoptosis through a phosphatidylinositol 3-kinase and NF-kappaB-dependent mechanism. J Immunol. 2006;176:5529–5537. doi: 10.4049/jimmunol.176.9.5529. [DOI] [PubMed] [Google Scholar]
- Ling Z, Tran KC, Arnold JJ, Teng MN. Purification and characterization of recombinant human respiratory syncytial virus nonstructural protein NS1. Protein Expr Purif. 2008;57:261–270. doi: 10.1016/j.pep.2007.09.017. [DOI] [PubMed] [Google Scholar]
- Ling Z, Tran KC, Teng MN. Human respiratory syncytial virus nonstructural protein NS2 antagonizes the activation of beta interferon transcription by interacting with RIG-I. J Virol. 2009;83:3734–3742. doi: 10.1128/JVI.02434-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu P, Jamaluddin M, Li K, Garofalo RP, Casola A, Brasier AR. Retinoic acid-inducible gene I mediates early antiviral response and Toll-like receptor 3 expression in respiratory syncytial virus-infected airway epithelial cells. J Virol. 2007;81:1401–1411. doi: 10.1128/JVI.01740-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu P, Li K, Garofalo RP, Brasier AR. Respiratory syncytial virus induces RelA release from cytoplasmic 100-kDa NF-kappa B2 complexes via a novel retinoic acid-inducible gene-I NF- kappa B-inducing kinase signaling pathway. J Biol Chem. 2008;283:23169–23178. doi: 10.1074/jbc.M802729200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loebbermann J, Schnoeller C, Thornton H, Durant L, Sweeney NP, Schuijs M, O’Garra A, Johansson C, Openshaw PJ. IL-10 regulates viral lung immunopathology during acute respiratory syncytial virus infection in mice. PLoS One. 2012;7:e32371. doi: 10.1371/journal.pone.0032371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loppow D, Schleiss MB, Kanniess F, Taube C, Jorres RA, Magnussen H. In patients with chronic bronchitis a four week trial with inhaled steroids does not attenuate airway inflammation. Respir Med. 2001;95:115–121. doi: 10.1053/rmed.2000.0960. [DOI] [PubMed] [Google Scholar]
- Lukacs NW, Smit JJ, Mukherjee S, Morris SB, Nunez G, Lindell DM. Respiratory virus-induced TLR7 activation controls IL-17-associated increased mucus via IL-23 regulation. J Immunol. 2010;185:2231–2239. doi: 10.4049/jimmunol.1000733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marwick JA, Caramori G, Stevenson CS, Casolari P, Jazrawi E, Barnes PJ, Ito K, Adcock IM, Kirkham PA, Papi A. Inhibition of PI3Kdelta restores glucocorticoid function in smoking-induced airway inflammation in mice. Am J Respir Crit Care Med. 2009;179:542–548. doi: 10.1164/rccm.200810-1570OC. [DOI] [PubMed] [Google Scholar]
- Mastronarde JG, He B, Monick MM, Mukaida N, Matsushima K, Hunninghake GW. Induction of interleukin (IL)-8 gene expression by respiratory syncytial virus involves activation of nuclear factor (NF)-kappa B and NF-IL-6. J Infect Dis. 1996;174:262–267. doi: 10.1093/infdis/174.2.262. [DOI] [PubMed] [Google Scholar]
- Matthews JG, Ito K, Barnes PJ, Adcock IM. Defective glucocorticoid receptor nuclear translocation and altered histone acetylation patterns in glucocorticoid-resistant patients. J Allergy Clin Immunol. 2004;113:1100–1108. doi: 10.1016/j.jaci.2004.03.018. [DOI] [PubMed] [Google Scholar]
- McCoy CE, Carpenter S, Palsson-McDermott EM, Gearing LJ, O’Neill LA. Glucocorticoids inhibit IRF3 phosphorylation in response to Toll-like receptor-3 and -4 by targeting TBK1 activation. J Biol Chem. 2008;283:14277–14285. doi: 10.1074/jbc.M709731200. [DOI] [PubMed] [Google Scholar]
- Mibayashi M, Martinez-Sobrido L, Loo YM, Cardenas WB, Gale M, Jr, Garcia-Sastre A. Inhibition of retinoic acid-inducible gene I-mediated induction of beta interferon by the NS1 protein of influenza A virus. J Virol. 2007;81:514–524. doi: 10.1128/JVI.01265-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murawski MR, Bowen GN, Cerny AM, Anderson LJ, Haynes LM, Tripp RA, Kurt-Jones EA, Finberg RW. Respiratory syncytial virus activates innate immunity through Toll-like receptor 2. J Virol. 2009;83:1492–1500. doi: 10.1128/JVI.00671-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pace TW, Hu F, Miller AH. Activation of cAMP-protein kinase A abrogates STAT5-mediated inhibition of glucocorticoid receptor signaling by interferon-alpha. Brain Behav Immun. 2011;25:1716–1724. doi: 10.1016/j.bbi.2011.07.227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panickar J, Lakhanpaul M, Lambert PC, Kenia P, Stephenson T, Smyth A, Grigg J. Oral prednisolone for preschool children with acute virus-induced wheezing. N Engl J Med. 2009;360:329–338. doi: 10.1056/NEJMoa0804897. [DOI] [PubMed] [Google Scholar]
- Paramore LC, Ciuryla V, Ciesla G, Liu L. Economic impact of respiratory syncytial virus-related illness in the US: an analysis of national databases. Pharmacoeconomics. 2004;22:275–284. doi: 10.2165/00019053-200422050-00001. [DOI] [PubMed] [Google Scholar]
- Patel JA, Jiang Z, Nakajima N, Kunimoto M. Autocrine regulation of interleukin-8 by interleukin-1alpha in respiratory syncytial virus-infected pulmonary epithelial cells in vitro. Immunology. 1998;95:501–506. doi: 10.1046/j.1365-2567.1998.00640.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patel JA, Kunimoto M, Sim TC, Garofalo R, Eliott T, Baron S, Ruuskanen O, Chonmaitree T, Ogra PL, Schmalstieg F. Interleukin-1 alpha mediates the enhanced expression of intercellular adhesion molecule-1 in pulmonary epithelial cells infected with respiratory syncytial virus. Am J Respir Cell Mol Biol. 1995;13:602–609. doi: 10.1165/ajrcmb.13.5.7576697. [DOI] [PubMed] [Google Scholar]
- Prins KC, Cardenas WB, Basler CF. Ebola virus protein VP35 impairs the function of interferon regulatory factor-activating kinases IKKepsilon and TBK-1. J Virol. 2009;83:3069–3077. doi: 10.1128/JVI.01875-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raboni SM, Nogueira MB, Tsuchiya LR, Takahashi GA, Pereira LA, Pasquini R, Siqueira MM. Respiratory tract viral infections in bone marrow transplant patients. Transplantation. 2003;76:142–146. doi: 10.1097/01.TP.0000072012.26176.58. [DOI] [PubMed] [Google Scholar]
- Ray A, Prefontaine KE. Physical association and functional antagonism between the p65 subunit of transcription factor NF-kappa B and the glucocorticoid receptor. Proc Natl Acad Sci U S A. 1994;91:752–756. doi: 10.1073/pnas.91.2.752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richter H, Seddon P. Early nebulized budesonide in the treatment of bronchiolitis and the prevention of postbronchiolitic wheezing. J Pediatr. 1998;132:849–853. doi: 10.1016/s0022-3476(98)70316-6. [DOI] [PubMed] [Google Scholar]
- Roosevelt G, Sheehan K, Grupp-Phelan J, Tanz RR, Listernick R. Dexamethasone in bronchiolitis: a randomised controlled trial. Lancet. 1996;348:292–295. doi: 10.1016/s0140-6736(96)02285-4. [DOI] [PubMed] [Google Scholar]
- Rudd BD, Burstein E, Duckett CS, Li X, Lukacs NW. Differential role for TLR3 in respiratory syncytial virus-induced chemokine expression. J Virol. 2005;79:3350–3357. doi: 10.1128/JVI.79.6.3350-3357.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Segovia J, Sabbah A, Mgbemena V, Tsai SY, Chang TH, Berton MT, Morris IR, Allen IC, Ting JP, Bose S. TLR2/MyD88/NF-kappaB pathway, reactive oxygen species, potassium efflux activates NLRP3/ASC inflammasome during respiratory syncytial virus infection. PLoS One. 2012;7:e29695. doi: 10.1371/journal.pone.0029695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sher ER, Leung DY, Surs W, Kam JC, Zieg G, Kamada AK, Szefler SJ. Steroid-resistant asthma. Cellular mechanisms contributing to inadequate response to glucocorticoid therapy. J Clin Invest. 1994;93:33–39. doi: 10.1172/JCI116963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sherman MP, de Noronha CM, Pearce D, Greene WC. Human immunodeficiency virus type 1 Vpr contains two leucine-rich helices that mediate glucocorticoid receptor coactivation independently of its effects on G(2) cell cycle arrest. J Virol. 2000;74:8159–8165. doi: 10.1128/jvi.74.17.8159-8165.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh D, McCann KL, Imani F. MAPK and heat shock protein 27 activation are associated with respiratory syncytial virus induction of human bronchial epithelial monolayer disruption. Am J Physiol Lung Cell Mol Physiol. 2007;293:L436–445. doi: 10.1152/ajplung.00097.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Somers CC, Ahmad N, Mejias A, Buckingham SC, Carubelli C, Katz K, Leos N, Gomez AM, DeVincenzo JP, Ramilo O, Jafri HS. Effect of dexamethasone on respiratory syncytial virus-induced lung inflammation in children: Results of a randomized, placebo controlled clinical trial. Pediatr Allergy Immunol. 2009;20:477–485. doi: 10.1111/j.1399-3038.2009.00852.x. [DOI] [PubMed] [Google Scholar]
- Stockman LJ, Curns AT, Anderson LJ, Fischer-Langley G. Respiratory syncytial virus-associated hospitalizations among infants and young children in the United States, 1997–2006. Pediatr Infect Dis J. 2012;31:5–9. doi: 10.1097/INF.0b013e31822e68e6. [DOI] [PubMed] [Google Scholar]
- Strahle L, Marq JB, Brini A, Hausmann S, Kolakofsky D, Garcin D. Activation of the beta interferon promoter by unnatural Sendai virus infection requires RIG-I and is inhibited by viral C proteins. J Virol. 2007;81:12227–12237. doi: 10.1128/JVI.01300-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun J, Cardani A, Sharma AK, Laubach VE, Jack RS, Muller W, Braciale TJ. Autocrine regulation of pulmonary inflammation by effector T-cell derived IL-10 during infection with respiratory syncytial virus. PLoS Pathog. 2011;7:e1002173. doi: 10.1371/journal.ppat.1002173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swedan S, Andrews J, Majumdar T, Musiyenko A, Barik S. Multiple functional domains and complexes of the two nonstructural proteins of human respiratory syncytial virus contribute to interferon suppression and cellular location. J Virol. 2011;85:10090–10100. doi: 10.1128/JVI.00413-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan YR, Peng D, Chen CM, Qin XQ. Nonstructural protein-1 of respiratory syncytial virus regulates HOX gene expression through interacting with histone. Mol Biol Rep. 2013;40:675–679. doi: 10.1007/s11033-012-2107-9. [DOI] [PubMed] [Google Scholar]
- Thomas KW, Monick MM, Staber JM, Yarovinsky T, Carter AB, Hunninghake GW. Respiratory syncytial virus inhibits apoptosis and induces NF-kappa B activity through a phosphatidylinositol 3-kinase-dependent pathway. J Biol Chem. 2002;277:492–501. doi: 10.1074/jbc.M108107200. [DOI] [PubMed] [Google Scholar]
- Tliba O, Damera G, Banerjee A, Gu S, Baidouri H, Keslacy S, Amrani Y. Cytokines induce an early steroid resistance in airway smooth muscle cells: Novel role of interferon regulatory factor-1. Am J Respir Cell Mol Biol. 2008;38:463–472. doi: 10.1165/rcmb.2007-0226OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tran KC, He B, Teng MN. Replacement of the respiratory syncytial virus nonstructural proteins NS1 and NS2 by the V protein of parainfluenza virus 5. Virology. 2007;368:73–82. doi: 10.1016/j.virol.2007.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright PF, Karron RA, Madhi SA, Treanor JJ, King JC, O’Shea A, Ikizler MR, Zhu Y, Collins PL, Cutland C, Randolph VB, Deatly AM, Hackell JG, Gruber WC, Murphy BR. The interferon antagonist NS2 protein of respiratory syncytial virus is an important virulence determinant for humans. J Infect Dis. 2006;193:573–581. doi: 10.1086/499600. [DOI] [PubMed] [Google Scholar]