

Abstract
The physiological responses of silicate-based bioactive glasses (BGs) are known to depend critically on both the P content (nP) of the glass and its silicate network connectivity (N̅BOSi). However, while the bioactivity generally displays a nonmonotonic dependence on nP itself, recent work suggest that it is merely the net orthophosphate content that directly links to the bioactivity. We exploit molecular dynamics (MD) simulations combined with 31P and 29Si solid-state nuclear magnetic resonance (NMR) spectroscopy to explore the quantitative relationships between N̅BO, nP, and the silicate and phosphate speciations in a series of Na2O–CaO–SiO2–P2O5 glasses spanning 2.1 ≤ N̅BOSi ≤ 2.9 and variable P2O5 contents up to 6.0 mol %. The fractional population of the orthophosphate groups remains independent of nP at a fixed N̅BO-value, but is reduced slightly as N̅BOSi increases. Nevertheless, P remains predominantly as readily released orthophosphate ions, whose content may be altered essentially independently of the network connectivity, thereby offering a route to optimize the glass bioactivity. We discuss the observed composition-structure links in relation to known composition-bioactivity correlations, and define how Na2O–CaO–SiO2–P2O5 compositions exhibiting an optimal bioactivity can be designed by simultaneously altering three key parameters: the silicate network connectivity, the (ortho)phosphate content, and the nNa/nCa molar ratio.
1. Introduction
Many melt-derived glasses of the Na2O–CaO–SiO2–P2O5 system, as well as other silicate-based analogues incorporating additional cations (e.g., Mg2+, Sr2+, Zn2+) and/or anions (e.g., F–) are termed “bioactive”, as when subjected to body fluids, they spontaneously integrate with the tissue through a hydroxy-carbonate apatite (HCA) surface layer.1−4 A few such bioactive glass (BG) options are widely exploited in orthopedic and periodontal surgery, e.g., the “45S5 Bioglass”.1−3
Silicate-based glasses only exhibit bioactivity over a relatively narrow span of compositions. However, despite vast efforts being spent for improving and tailoring these materials, encompassing bioactivity assessments (i.e., the rate of HCA formation) both in vitro and in vivo,2,3 very few quantitative composition/bioactivity relations are reported. The present insight thereof derives from empirical testing that typically involve systematic variations of the relative oxide contents, e.g., SiO2 and P2O5.2,3,5−9 Unfortunately, given the currently insufficient understanding of the underlying structure/composition relations that ultimately control the bioactivity, most empirically derived composition/bioactivity correlations have limited applicability to related BG formulations generated, for instance, by replacing glass modifier cations or by introducing additional glass network-forming elements, such as boron.
Considering the relatively modest progress made toward a rational design of BGs featuring optimal bioactivities via the relative oxide contents as direct variables, it is highly desirable to identify relatively simple descriptors10 that reasonably accurately capture the essential composition/bioactivity relations through the insight provided from the underlying structure/composition links. Two such parameters are employed,5,11,12 although relatively sparsely and not nearly to their full potential:
-
(i)
One descriptor is the silicate network connectivity,11−15 here denoted N̅BOSi and corresponding to the average number of bridging oxygen (BO) atoms per SiO4 group in the structure. QSi labels a SiO4 tetrahedron exhibiting n BO atoms and 4 – n nonbridging oxygen (NBO) ions. The negatively charged NBO species are balanced by the Na+/Ca2+ modifiers. The importance of the network connectivity for the bioactivity was first highlighted by Strnad;11 this concept was subsequently developed further and applied by several groups.7,8,12,14−25 The exact limits of N̅BOSi required for bioactive glass compositions remain unsettled,11,12,15,22 but ”favorable” values are commensurate with open structures primarily built by QSi and QSi3 groups that readily degrade when subjected to body fluids.
-
(ii)
The P content of the BG constitutes the second parameter for bioactivity predictions. Phosphorus is generally present as readily released orthophosphate anions (i.e., QP0 groups) dispersed across the glass matrix.26,27 Numerous investigations conducted both in vitro and in vivo reveal that the presence of P boosts the bioactivity.2,3,5−7,9 Dissolved Ca2+ and PO43– species increase the supersaturation of the HCA constituents in the body fluid, thereby accelerating the phosphate formation at the glass surface.2,3,5,28 The ion release also controls the local pH and avoids excessive acidity that inhibits bone bonding.29 However, if the glass-modifier content is insufficient for fully balancing all negative charges of the orthophosphate species, QP and higher-polymerization QPn (n = 2, 3) phosphate moieties form. They are believed to diminish the bioactivity, owing to their lower mobility and the higher energetic cost associated with their release compared with the orthophosphate groups.17,18 Yet, in modifier-rich BGs the existence and nature of the QP environments remains debated,14−19,30−35 in particular which compositional/structural parameters control their populations.
Given that the glass bioactivity is enhanced either by a relatively low silicate network polymerization or by large amounts of orthophosphate groups, designing BGs that combine these two beneficial features is an appealing perspective for optimizing the bioactivity. One option involves adding P to a base glass composition, while the silicate network connectivity is kept fixed at a “favorable” value. However, any potential bearings on N̅BOSi or the QP speciation by introducing high P contents must first be assessed. The extent to which additional P atoms enter the silicate network and distribute between ortho- and nonorthophosphate moieties dictates if the goal indicated above is practically feasible, i.e., whether it is possible to design low-connectivity BGs that feature substantial orthophosphate contents. To answer this key question, herein we quantitatively examine the relationships between the phosphate speciation, the silicate network connectivity, and the P content of the glass across wide compositional ranges.
Herein, we assess the structural alterations accompanying the variations of the P content and N̅BOSi by monitoring the sets of {QSi} and {QPn} populations that build each structure, as probed experimentally by magic-angle spinning (MAS) 29Si and 31P nuclear magnetic resonance (NMR) spectroscopy, as well as by molecular dynamics (MD) simulations.36 The observed composition–structure correlations are linked with composition–bioactivity relationships reported in the literature to advance the structure–bioactivity insight; we further suggest rational ways to combine the key descriptors toward an optimal bioactivity. The present direct comparison between the MD and NMR derived structural features is also the first attempt of its kind for a large set of BG samples (vide infra), which further span a wide range of (Na, Ca, Si, P) compositions. Our results confirm that both experimental and computational approaches reveal fully consistent qualitative trends. This mutual validation is also important for enabling discussions on specific MD-derived structural features that are not experimentally accessible.36
2. Glass Series Design
To elucidate the structural changes resulting from variations of either the P content or the silicate network connectivity while the other parameter remains fixed, we prepared a set of 13 BG specimens from the quaternary Na2O–CaO–SiO2–P2O5 system, each labeled BGp(N̅BOSi), with 0≤p≤6.0 representing the P2O5 content in mol % (see Table 1). Two glass series featuring nominal N̅BO values of N̅BOSi(nom)=2.5 and N̅BO(nom)=2.9, respectively, were derived by assuming that P is present solely as orthophosphate groups (QP0) in each glass structure, thereby requiring a known net amount of the Na+/Ca2+ network modifiers for charge balance. Both BG families exhibit a constant molar ratio nNa/nCa = 1.54, and the modifier reservoir consistently remains sufficiently large for simultaneously arranging the desired N̅BO-value together with the sole presence of QP0 groups. This BG design strategy, employed in a few recent studies,14,22,31 implies that the total nNa+nCa amount increases concurrently with nP across each BG(2.5) and BG(2.9) series, while that of Si decreases, approximately amounting to a net Si→P stoichiometric replacement (see Table 1).
Table 1. BG Sample Compositionsa.
| label | N̅BOSi(nom)b | aNa2O (mol %) | bCaO (mol %) | cSiO2 (mol %) | dP2O5 (mol %) | stoichiometric formula | y(CaO)c | nNa/nCad | ρ (g cm–3)e |
|---|---|---|---|---|---|---|---|---|---|
| BG2.6(2.1) | 2.11 | 0.244 (0.229) | 0.269 (0.255) | 0.461 (0.486) | 0.026 (0.030) | Na0.384Ca0.212Si0.363P0.041O1.233 | 0.52 | 1.81 | 2.704 |
| BG6.0(2.1) | 2.15 | 0.221 (0.207) | 0.324 (0.315) | 0.395 (0.412) | 0.060 (0.065) | Na0.345Ca0.253Si0.308P0.094O1.325 | 0.59 | 1.36 | 2.743 |
| BG0(2.5) | 2.50 | 0.186 (0.178) | 0.242 (0.231) | 0.572 (0.591) | 0.000 (0.000) | Na0.314Ca0.204Si0.482P0.000O1.325 | 0.57 | 1.54 | 2.673 |
| BG1.0(2.5) | 2.50 | 0.192 (0.181) | 0.249 (0.250) | 0.549 (0.559) | 0.010 (0.010) | Na0.319Ca0.207Si0.457P0.017O1.322 | 0.57 | 1.54 | 2.685 |
| BG2.0(2.5) | 2.50 | 0.197 (0.191) | 0.257 (0.258) | 0.526 (0.527) | 0.020 (0.024) | Na0.324Ca0.211Si0.432P0.033O1.320 | 0.57 | 1.54 | 2.691 |
| BG4.0(2.5) | 2.50 | 0.209 (0.193) | 0.271 (0.254) | 0.480 (0.509) | 0.040 (0.044) | Na0.334Ca0.217Si0.385P0.064O1.314 | 0.57 | 1.54 | 2.693 |
| BG6.0(2.5) | 2.50 | 0.219 (0.201) | 0.287 (0.274) | 0.434 (0.460) | 0.060 (0.065) | Na0.343Ca0.223Si0.340P0.094O1.310 | 0.57 | 1.54 | 2.707 |
| BG2.6(2.7) | 2.74 | 0.202 (0.194) | 0.222 (0.214) | 0.550 (0.560) | 0.026 (0.032) | Na0.329Ca0.181Si0.448P0.042O1.347 | 0.52 | 1.81 | 2.635 |
| BG0(2.9) | 2.93 | 0.151 (0.143) | 0.197 (0.183) | 0.652 (0.674) | 0.000 (0.000) | Na0.262Ca0.171Si0.566P0.000O1.435 | 0.57 | 1.53 | 2.600 |
| BG2.0(2.9) | 2.93 | 0.165 (0.151) | 0.215 (0.206) | 0.600 (0.623) | 0.020 (0.020) | Na0.278Ca0.182Si0.506P0.034O1.418 | 0.57 | 1.53 | 2.624 |
| BG3.0(2.9) | 2.93 | 0.172 (0.161) | 0.224 (0.226) | 0.574 (0.583) | 0.030 (0.030) | Na0.286Ca0.187Si0.477P0.050O1.409 | 0.57 | 1.53 | 2.625 |
| BG4.0(2.9) | 2.93 | 0.179 (0.162) | 0.233 (0.226) | 0.548 (0.572) | 0.040 (0.040) | Na0.294Ca0.191Si0.449P0.066O1.401 | 0.57 | 1.53 | 2.639 |
| BG6.0(2.9) | 2.93 | 0.193 (0.172) | 0.252 (0.244) | 0.495 (0.524) | 0.060 (0.060) | Na0.308Ca0.201Si0.396P0.096O1.385 | 0.57 | 1.53 | 2.661 |
{a, b, c, d} specify the nominally batched aNa2O–bCaO–cSiO2–dP2O5 oxide equivalents with a + b + c + d = 1; they were used for obtaining the stoichiometric formulae. Values within parentheses correspond to EDX-analyzed compositions.
The silicate network connectivity, i.e., average number of bridging oxygen atoms per SiO4 tetrahedron, calculated by accounting for the modifier cation-consumption of the orthophosphate species in each batched glass composition, according to the procedure of Edén.15,23
Molar fraction of CaO out of the glass modifier oxides; y(CaO) = n(CaO)/[n(CaO) + n(Na2O)].
Molar ratio between Na+ and Ca2+ cations.
Densities (accurate within ±0.003 g cm–3) were determined by the Archimedes method in water at 22 °C.
Besides each BG(2.5) and BG(2.9) branch (where we drop the p subscript when collectively referring to all its members), two additional samples—BG2.6(2.1) and BG2.6(2.7)—form a set of glasses exhibiting roughly constant P2O5 contents (2–3 mol %), but with N̅BOSi increasing from 2.1 to 2.9. This series enables us to selectively probe the structural changes resulting when solely the silicate network-connectivity varies. Note that the BG2.6(2.1) specimen corresponds to the widely utilized “45S5 Bioglass”.1−3
The primary aim of the present work is to enhance the structure–bioactivity insight by exploring the phosphate and silicate speciations of both bioactive and nonbioactive glass compositions that are selected based on the following hypotheses, inferred and discussed by Edén15 on the basis of in vitro bioactivity tests reported in the literature:6,7 (A) A silicate network connectivity conforming to the range 2.0 ≲ N̅BOSi ≲ 2.7 is a prerequisite for bioactivity; it is optimized for values around 2.5–2.6.15 (B) Provided that requirement A is fulfilled, the bioactivity grows monotonically with the net orthophosphate content
| 1 |
where nP is the stoichiometric amount of P (see Table 1). Here and onward, xPn and xSi denote the fractional population of QPn and QSi groups out of the total {QPn} and {QSi} speciation, respectively. In this work, we examine and discuss hypotheses A and B further.
3. Materials and Methods
3.1. Sample Preparations and Characterization
The BGs were prepared from precursors of NaH2PO4 (99.99%; Merck), and Na2CO3 (99.9%), CaCO3 (99.9%), and SiO2 (99.99%) from ChemPur. To accelerate spin–lattice relaxation for the NMR experimentation, 0.1 wt % of Fe2O3 was added to each batch. Each precursor-mixture (6.0 g) was ball-milled for 12 h and subsequently placed in a Pt crucible that was heated for 4 h in an electric furnace at 950 °C for CO2 removal. The temperature was set to a final value in the range of 1350–1620 °C, with the highest temperatures required by compositions exhibiting a large P-content and/or high NBOSi-value. Each melt was held for 4 h prior to its quenching by immersing the bottom of the crucible in water.
Powder X-ray diffraction with a PANalytical X’pert PRO MPD diffractometer and Cu Kα1 radiation revealed no crystalline phases (detection limit ≲1%). Here, 2θ ranged between 10° and 70°. Scanning electron microscopy (SEM) with a JSM 7000F (JEOL) microscope in backscatter electron imaging mode (20 kV acceleration voltage) evidenced that each specimen constitutes a homogeneous glass phase, except for BG6.0(2.9) that manifested tendencies of amorphous phase-separation. Cation contents were estimated by a LINK INCA (Oxford instruments) energy-dispersive X-ray (EDX) detector. Each reported composition resulted from averaging over ten analyses from distinct fragments, with the respective O content calculated to charge-balance the cations. The nominal and EDX-analyzed glass compositions agree overall well (see Table 1), where the deviations remain within the uncertainty of the analysis. Hence, the sample nomenclature and structural analysis assume the nominal glass compositions. See Mathew et al.25 for further synthesis/characterization details.
3.2. Solid-State NMR Experiments
All MAS NMR experimentation utilized a Bruker Avance-III spectrometer operating at 9.4 T; i.e., the 31P and 29Si Larmor frequencies are −162.0 MHz and 79.47 MHz, respectively. Finely ground glass powders were packed in 7 mm zirconia rotors and spun at 7.00 kHz. The NMR acquisitions utilized single pulses with flip angles, rf nutation frequencies, and relaxation delays of {70°, 62.5 kHz, 120 s} and {90°, 81 kHz, 40 s} for 29Si and 31P, respectively, with the corresponding number of accumulated signal transients depending on the content of the detected nucleus and ranging over 400–900 and 256–640. Separate T1 relaxation measurements verified that these relaxation delays provide quantitative NMR spectra. No signal apodization was employed in the data processing. 31P and 29Si chemical shifts are quoted relative to 85% H3PO4(aq) and neat tetramethylsilane (TMS), respectively.
3.3. Molecular Dynamics Simulations
Classical MD simulations were performed with the DLPOLY3 package37,38 for NVT ensembles, where 6000–12000 {Na, Ca, Si, P, O} atoms were placed in a cubic box with periodic boundary conditions and a size within 4.2–5.1 nm; these numbers varied slightly to match each nominal BG composition and experimental density (see Table 1 and the Supporting Information). The melt-quench simulation started from a random atom configuration, equilibrated for 100 ps at 3500 K, followed by a 10 K/ps cooling procedure down to 300 K. A final NVT run at 300 K was then performed for 200 ps, of which the last 150 ps were used for the structural analysis. For each glass composition, this protocol was completed 2–4 times with different initial configurations (see the Supporting Information). The average value and uncertainty of each reported structural parameter were derived from these independent samples.
A polarizable shell-model potential, developed for modeling multicomponent glasses,17−19,39 was employed throughout. Each cation carries its full formal charge. O2– species are represented as core (OC) and shell (OS) units coupled by a 300 THz harmonic oscillator and bearing charges of zC = 0.8482e and zS = −2.8482e, respectively (i.e., zC + zS = −2e constitutes the formal charge). The short-range OS–OS and cation–OS interactions were parametrized by a Buckingham potential,17−19,39 evaluated for all pairs up to 0.8 nm. Long-range Coulombic interactions among all charged particles were calculated by a smoothed particle mesh Ewald summation38 with a 1.2 nm real-space cutoff and an accuracy of 10–6. The intratetrahedral O–Si–O and O–P–O bond angles were constrained by using three-body truncated harmonic potentials.38 The equations of motion were integrated in time-steps of 0.2 fs by the velocity Verlet integrator approach. The temperature was controlled by a Berendsen thermostat with a 1.0 ps relaxation time constant. See refs (17−19 and 39) for further details on the simulation procedures, and the Supporting Information for additional discussions on the convergence of the MD data with respect to the system size and the cooling rate of the simulation.
4. Results
4.1. 31P NMR
The 31P MAS NMR spectra recorded from the BG(2.5) and BG(2.9) series are displayed in Figure 1. All are representative of phosphate groups in strongly disordered structures, as reflected by NMR signals associated with 7.5–8.0 ppm full-width at half-maximum (fwhm) height. Onward disregarding the BG6.0(2.9) sample that is not fully homogeneous (see section 3.1), all specimens within each fixed-N̅BOSi series exhibit essentially equal 31P NMR peak maxima (δmax) and fwhm values, regardless of the P content. The observed peak maxima in the range 6–9 ppm are consistent with QP (orthophosphate) environments charge-balanced by both Na+ and Ca2+.26,27,40
Figure 1.

Experimental 31P MAS NMR spectra recorded from Na2O–CaO–SiO2–P2O5 glasses, where the BG(2.5) and BG(2.9) series are displayed in the left and right columns, respectively, and ordered according to increasing P2O5 contents from top to bottom. The peak maximum is specified at the outermost portion of each spectrum; it is mainly dictated by the 31P resonances from QP0 groups, whose traces are not indicated, except for the NMR spectrum from BG6.0(2.9) that required deconvolutions into two distinct 31P signals. Shaded areas mark signal contributions from QP moieties, whose relative populations (in %) are indicated. The curve beneath each spectrum reveals the difference between the experimental and best-fit spectra.
The NMR peakshape recorded from each BG(2.5) glass is nearly Gaussian, but it becomes progressively more asymmetric when the silicate-network connectivity increases: this growing “tail” stems from QP1 groups.16,20,25−27,31−33,41,42 A main goal of the present work is to quantify these populations and rationalize their dependence on the silicate network connectivity and P content of the BG. The Supporting Information discusses the problems of detecting low amounts of QP groups by solid-state 31P NMR on BGs (such as the most thoroughly studied “45S5” composition; e.g., see Pedone et al.30), as well as highlighting their assisted probing by exploiting information from the spinning sidebands in the NMR spectrum. While we for brevity denote the n ≥ 1 phosphate moieties by QPn, their BO atoms all involve linkages to Si rather than P, as highlighted previously in numerous MD studies,17−19,30 as well as experimentally by through-space and through-bond coupling-based NMR techniques.33,41 We note that standard 31P MAS NMR experimentation cannot discriminate between 31P in Si–O–P and P–O–P motifs.33,41 The MD-derived fractions of P–O–P bonds in the present glasses remain very low (≲ 3% out of all P–O–Si and P–O–P motifs) and are only present in the P-richest samples featuring ≥4 mol % P2O5.
The 31P NMR results of Figure 2 from glasses featuring a nearly constant P content (2–3 mol % P2O5) but variable N̅BOSi values between 2.1 and 2.9, suggest a strict relationship between the fraction of QP groups in the BG structure and its silicate network connectivity. To quantify these relationships further, we deconvoluted each 31P NMR signal into two Gaussian peak components stemming from QP0 and QP species by constrained iterative fitting (see the Supporting Information). Each peak is represented by the parameter-triplet (δPn, WP, xPn), corresponding to the mean 31P chemical shift, the fwhm, and the fractional population, respectively, of the given QP species. The best-fit results are listed in Table 2. They reveal no significant variations across the set of BGs, as anticipated from the very similar experimental 31P NMR spectra. Owing to the higher 31P shielding resulting from Qn → Qn+1 conversions, the chemical shifts associated with QP0 and QP groups are separated by ≈8 ppm, with those from QP0 moieties appearing within a narrow window of 6.4–8.7 ppm, whereas their QP counterparts span between −1.7 ppm and 0.4 ppm. As discussed further in section 4.3, the results from BGs associated with four distinct N̅BOSi-values evidence a concurrent growth of the relative fraction of the QP environments from ≈0.05 to ≈0.20 when N̅BOSi is increased between 2.1 and 2.9, whereas the xP values are independent of the P content within each BG(N̅BOSi) series.
Figure 2.

Experimental 31P (left column) and 29Si (right column) NMR spectra, plotted with black traces and acquired from the as-indicated BGp(N̅BOSi) samples. The spectra are listed according to increasing silicate network connectivity from top to bottom. Each 31P NMR contribution from the QP groups is marked by a shaded area (left column), with the respective population (in %) specified. The gray traces in the right column represent 29Si NMR peaks stemming from the various QSin groups, as obtained by spectral deconvolution. The curve beneath each NMR spectrum represents the difference between the experiment and its respective best-fit.
Table 2. 31P NMR Resultsa.
| net peak |
QP0 |
QP1 |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| label | δmax (ppm) | W (ppm) | δ0 (ppm) | W0 (ppm) | x0 | δ1 (ppm) | W1 (ppm) | x1 | N̅BOSi(nom) | N̅BOSi(NMRP)b |
| BG2.6(2.1) | 8.8 | 7.51 | 8.7 | 7.4 | 0.959 | 0.4 | 7.3 | 0.041 | 2.11 | 2.11 |
| BG6.0(2.1) | 7.3 | 7.48 | 7.3 | 7.4 | 0.955 | –0.8 | 7.4 | 0.045 | 2.15 | 2.14 |
| BG0(2.5) | – | – | – | – | – | – | – | – | 2.50 | 2.50 |
| BG1.0(2.5) | 7.4 | 7.77 | 7.4 | 7.6 | 0.899 | –0.6 | 8.1 | 0.101 | 2.50 | 2.50 |
| BG2.0(2.5) | 7.3 | 7.72 | 7.4 | 7.5 | 0.896 | –0.6 | 8.3 | 0.104 | 2.50 | 2.50 |
| BG4.0(2.5) | 7.5 | 7.57 | 7.5 | 7.4 | 0.931 | –0.6 | 7.9 | 0.069 | 2.50 | 2.49 |
| BG6.0(2.5) | 7.2 | 7.55 | 7.2 | 7.4 | 0.902 | –0.8 | 8.2 | 0.098 | 2.50 | 2.48 |
| BG2.6(2.7) | 7.5 | 7.72 | 7.5 | 7.4 | 0.838 | –0.6 | 8.4 | 0.162 | 2.74 | 2.73 |
| BG0(2.9) | – | – | – | – | – | – | – | – | 2.93 | 2.93 |
| BG2.0(2.9) | 6.5 | 7.96 | 6.4 | 7.5 | 0.795 | –1.7 | 8.6 | 0.205 | 2.93 | 2.92 |
| BG3.0(2.9) | 6.7 | 7.97 | 6.8 | 7.5 | 0.814 | –1.2 | 8.9 | 0.186 | 2.93 | 2.91 |
| BG4.0(2.9) | 6.4 | 7.85 | 6.5 | 7.4 | 0.805 | –1.5 | 8.7 | 0.195 | 2.93 | 2.90 |
| BG6.0(2.9)c | 5.7 | 7.47 | 7.0 | 7.1 | 0.549 | –0.6 | 9.2 | 0.179 | 2.93 | 2.89 |
| 4.8 | 5.7 | 0.272 | ||||||||
The data involves the net NMR chemical shift (δmax; uncertainty ±0.1 ppm) and fwhm height (W; ±0.15 ppm) of the 31P NMR signal, as well as the chemical shift (δn; ±0.3 ppm), fwhm height (Wn; ±0.5 ppm), and fractional population (xn; ±0.02) of each QPn contribution extracted by spectra deconvolution.
Vaules corrected for the NMR-derived minor QP1 contributions.
This sample exhibits phase separation and its 31P NMR spectrum required two QP0 signals for deconvolution.
For a fixed number of NBO ions at the phosphate group, its 31P chemical shift is dictated mainly by the nature and location of the surrounding glass-modifier cations.16,27,31,40 Previous 31P NMR reports on Na–Ca–Si–P–O glasses16,25,27,30−32,43 observed δP0 values that are intermediate of the shifts associated with the orthophosphate groups present in polymorphs of Na3PO4 and Ca3(PO4)2 that resonate around 12–15 ppm and 0–3 ppm, respectively. A linear relationship between δP and y(CaO) = n(CaO)/[n(CaO) + n(Na2O)] is reported for Na2O–CaO–SiO2–P2O5 glasses.16,27,31 The present 31P NMR results for BGs featuring a nearly constanty(CaO) value accord with those findings: the following weighted average,
| 2 |
with y(CaO) = 0.57 for the BG(2.5) and BG(2.9) series (see Table 1), predicts a chemical-shift range of 5.2 ≤ (δ̅P0/ppm) ≤ 8.2 for the QP groups when δA and δB are selected over the typical shift-spans of crystalline Na and Ca based orthophosphates, i.e., 12–15 ppm and 0–3 ppm, respectively. Consequently, our observed δP0 values in the range of 6.5–7.5 ppm are overall consistent with earlier NMR work reporting an essentially statistical Na/Ca distribution around the orthophosphate species;16,27,31,43 see the Supporting Information for further discussions.
The present NMR analysis cannot discriminate between a strict statistical (Na,Ca)–P association and minor preferences of either cation to charge-balance the phosphate moieties. A more accurate 17O NMR analysis applied to the 45S5 BG, revealed a slight depletion of Na+–PO43– contacts,30 while previous MD simulations also suggested a strong preference for Ca2+–PO43– rather than Na+–PO43– associations,17,44−46 as also observed in our modeled BG2.6(2.1) structure. The MD-derived Na/Ca partitioning among Si/P for the entire glass series will be presented together with those from a more extended NMR study in a forthcoming paper.
4.2. 29Si NMR
For Na–Ca–Si–P–O glasses, the mean 29Si chemical shift (δSin) of a QSi species depends principally on the same structural factors as 31P, i.e., the number of BO atoms (n) at the SiO4 tetrahedron, and the Na+/Ca2+ constellation for local charge-balance.16,27,40,47,48 All glasses within a given BG(N̅BOSi) branch exhibit a constant nNa/nCa ratio. Consequently, the center-of-gravity shift (δCG) of the 29Si NMR peak and the accompanying peakshape are expected to depend predominantly on the precise set of fractional populations {xSi} and their associated {δSin} values, where 29Si nuclei of QSi moieties typically resonate in the range from −112 ppm to −100 ppm and the shift increases by roughly 7–12 ppm per QSin → QSi conversion.40,47,48 Hence, for the present glasses, identical 29Si NMR responses are expected throughout a given BG(N̅BOSi) series, whereas a gradual net displacement toward more negative shifts should occur when N̅BO increases. The latter trend is witnessed by the 29Si MAS NMR spectra presented in Figure 2. In contrast, very similar spectra are observed across each BG(2.5) and BG(2.9) branch, regardless of the P content of the BG; see the top row of Figure 3. Furthermore, Table 3 reveals very similar sets of 29Si NMR best-fit parameters {δSin, WSi, xSin} within each N̅BO-branch.
Figure 3.
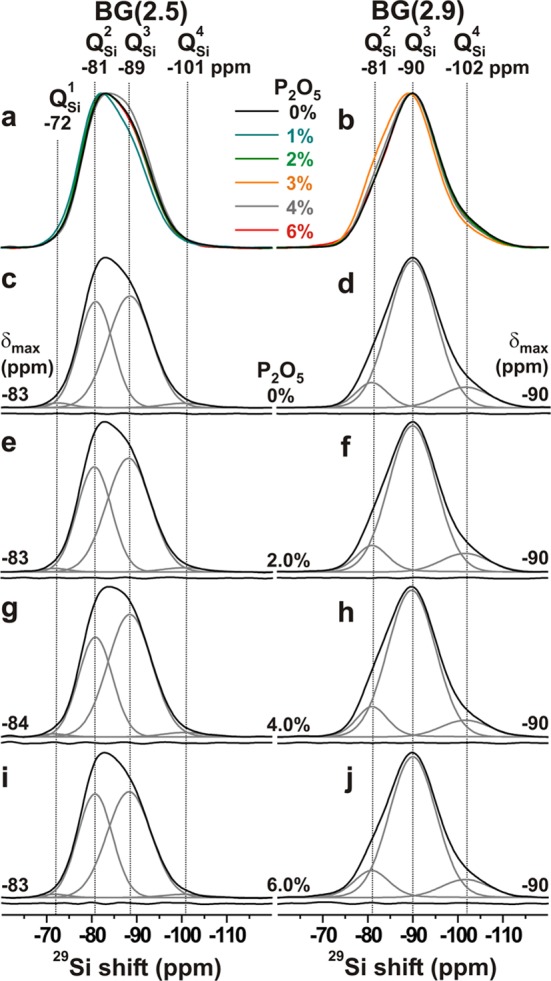
29Si MAS NMR spectra obtained from the BG(2.5) (left column) and BG(2.9) (right column) series of glasses. (a, b) Superimposed NMR spectra associated with the as-indicated glasses. (c–j) Experimental spectra (black traces) displayed together with deconvoluted peak-components (gray traces). They are assigned at the top of each column. The curves beneath the spectra constitute differences between experiments and best-fits.
Table 3. 29Si NMR Resultsa.
| net NMR peak |
QSi1 |
QSi2 |
QSi3 |
QSi4 |
|||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| label | –δmax (ppm) | –δCG (ppm) | W (ppm) | –δ1 (ppm) | W1 (ppm) | x1 | –δ2 (ppm) | W2 (ppm) | x2 | –δ3 (ppm) | W3 (ppm) | x3 | –δ4 (ppm) | W4 (ppm) | x4 | N̅BOSi(nom) | N̅BOSi(NMRSi) |
| BG2.6(2.1) | 79.5 | 81.0 | 11.12 | 72.5 | 7.5 | 0.066 | 79.5 | 9.1 | 0.724 | 87.8 | 9.3 | 0.210 | – | – | – | 2.11 | 2.14 |
| BG6.0(2.1) | 80.9 | 81.7 | 13.66 | 73.4 | 7.1 | 0.056 | 80.2 | 9.2 | 0.658 | 87.5 | 11.0 | 0.286 | – | – | – | 2.15 | 2.20 |
| BG0(2.5) | 83.0 | 85.3 | 15.68 | 72.5 | 7.0 | 0.015 | 80.9 | 8.9 | 0.409 | 88.5 | 11.5 | 0.555 | 100.5 | 11.0 | 0.021 | 2.50 | 2.58 |
| BG1.0(2.5) | 82.1 | 84.0 | 15.12 | 72.4 | 7.0 | 0.017 | 80.5 | 9.1 | 0.455 | 88.0 | 11.5 | 0.512 | 100.5 | 11.0 | 0.016 | 2.50 | 2.53 |
| BG2.0(2.5) | 82.9 | 85.1 | 15.80 | 71.8 | 7.0 | 0.012 | 80.6 | 8.9 | 0.405 | 88.2 | 11.5 | 0.563 | 100.5 | 11.0 | 0.020 | 2.50 | 2.59 |
| BG4.0(2.5) | 84.0 | 85.6 | 15.83 | 71.6 | 7.0 | 0.010 | 80.8 | 8.9 | 0.372 | 88.5 | 11.5 | 0.598 | 100.5 | 11.0 | 0.020 | 2.50 | 2.63 |
| BG6.0(2.5) | 82.8 | 85.0 | 15.75 | 71.9 | 7.0 | 0.010 | 80.7 | 9.0 | 0.423 | 88.3 | 11.5 | 0.551 | 100.5 | 11.0 | 0.016 | 2.50 | 2.57 |
| BG2.6(2.7) | 88.2 | 87.6 | 15.56 | 72.3 | 5.2 | 0.006 | 81.0 | 8.4 | 0.242 | 88.9 | 11.0 | 0.677 | 98.1 | 12.0 | 0.075 | 2.74 | 2.82 |
| BG0(2.9) | 89.8 | 90.1 | 14.70 | – | – | – | 80.9 | 8.2 | 0.094 | 89.9 | 12.5 | 0.793 | 101.7 | 12.8 | 0.113 | 2.93 | 3.02 |
| BG2.0(2.9) | 89.9 | 90.1 | 14.57 | – | – | – | 81.0 | 8.2 | 0.108 | 89.9 | 12.4 | 0.791 | 101.6 | 12.5 | 0.101 | 2.93 | 2.99 |
| BG3.0(2.9) | 89.2 | 89.0 | 15.53 | – | – | – | 80.9 | 8.4 | 0.144 | 89.3 | 12.5 | 0.779 | 101.4 | 12.9 | 0.077 | 2.93 | 2.93 |
| BG4.0(2.9) | 89.7 | 89.9 | 14.46 | – | – | – | 81.0 | 8.1 | 0.118 | 89.7 | 12.5 | 0.791 | 101.7 | 12.7 | 0.091 | 2.93 | 2.97 |
| BG6.0(2.9) | 90.0 | 90.1 | 14.41 | – | – | – | 80.8 | 8.1 | 0.121 | 90.0 | 12.2 | 0.778 | 101.7 | 12.6 | 0.101 | 2.93 | 2.98 |
The data involve the peak maximum (δmax; uncertainty ±0.2 ppm), the center-of-gravity shift (δCG; ±0.3 ppm) and the fwhm height (W; ±0.3 ppm) of the net 29Si NMR signal, and the chemical shift (δn), fwhm height (Wn), and fractional population (xn) of each QSin peak contribution resulting from spectral deconvolution. N̅BO(nom) and N̅BOSi(NMRSi) represent the silicate network connectivity obtained from the BG composition and by eq 3, respectively.
The net NMR responses derive from superimposed signals centered at the chemical shifts {δSi4, δSi, δSi2, δSi} that appear around {−101, −88, −81, −72} ppm, respectively (see Figures 2 and 3). However, typically only three components are significant across the entire N̅BOSi-span of the present glasses, of which two moieties dominate: QSi and QSi3. The networks of the BG(2.9) glasses are primarily built by QSi groups (≈78% out of the total SiO4 speciation), as expected, with the remaining constituting roughly equal amounts ≈10% of QSi4 and QSi groups. While minor QSi2 contributions must be present because N̅BO < 3.0, the NMR spectra unambiguously also reveal the presence of QSi4 groups. This feature stems from disproportionation equilibria, such as 2QSi ↔ QSin+1 + QSi.40,47,48 They produce deviations from a strict binary distribution that predicts the coexistence of only two tetrahedral species, QSin and QSi, except when N̅BOSi equals any member of the set {0, 1, 2, 3, 4}, when one sole QSi group associated with n = N̅BOSi results. A similar trend is observed for the BG(2.5) series for which a binary distribution predicts equal {xSi, xSi3} values, whereas the NMR-derived QSi populations (Table 3) reveal higher (≈0.55) and lower (≈0.41) xSi3 and xSi fractions, respectively; those NMR spectra further comprise weak signals from QSi4 and QSi tetrahedra (xSin ≲ 0.02).
From each set {xSin}, we calculated the 29Si NMR-derived value N̅BO(NMRSi), according to
| 3 |
Table 4 compares these results with their nominal counterparts [N̅BOSi(nom)], as well as those obtained by correcting the latter values for the observed non-negligible amounts of QP groups (whose presence releases Na+/Ca2+ modifiers that become available for depolymerizing the silicate glass network), which provides a set of 31P NMR-derived values, denoted {N̅BOSi(NMRP)}. Owing to the overall low amounts of P, each of N̅BO(NMRP) remains close to its N̅BOSi(nom) analogue (within ≤1% deviation throughout). Whereas the N̅BO(NMRSi) values stay fairly constant across each BG(2.5) and BG(2.9) branch, they are consistently 2–4% larger than their nominal counterparts. Yet, it is gratifying that the deviations between the respective N̅BOSi data obtained by 31P and 29Si NMR typically only amount to ≈3%, with the largest discrepancy (≈5%) observed for the BG4.0(2.5) glass.
Table 4. Fractional populations of QSin Groups derived by MD simulations and NMRa.
|
xPn populationsb |
xSin populationsb |
silicate network connectivity |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| label | QP0 | QP1 | QSi1 | QSi2 | QSi3 | QSi4 | N̅BOSi(nom) | N̅BOSi(NMRP)c | N̅BOSi(NMRSi)d | N̅BOSi(MD)e |
| BG2.6(2.1) | 0.834 (0.959) | 0.166 (0.041) | 0.186 (0.066) | 0.517 (0.724) | 0.266 (0.210) | 0.021 (0.000) | 2.11 | 2.11 | 2.14 | 2.10 |
| BG6.0(2.1) | 0.798 (0.955) | 0.200 (0.045) | 0.197 (0.056) | 0.514 (0.658) | 0.252 (0.286) | 0.027 (0.000) | 2.15 | 2.14 | 2.20 | 2.09 |
| BG0(2.5) | – | – | 0.068 (0.015) | 0.428 (0.409) | 0.425 (0.555) | 0.076 (0.021) | 2.50 | 2.50 | 2.58 | 2.50 |
| BG1.0(2.5) | 0.670 (0.899) | 0.318 (0.101) | 0.073 (0.017) | 0.426 (0.455) | 0.435 (0.512) | 0.065 (0.016) | 2.50 | 2.50 | 2.53 | 2.49 |
| BG2.0(2.5) | 0.721 (0.896) | 0.267 (0.104) | 0.080 (0.012) | 0.418 (0.405) | 0.440 (0.563) | 0.060 (0.020) | 2.50 | 2.50 | 2.59 | 2.48 |
| BG4.0(2.5) | 0.622 (0.931) | 0.364 (0.069) | 0.084 (0.010) | 0.443 (0.372) | 0.409 (0.598) | 0.063 (0.020) | 2.50 | 2.49 | 2.63 | 2.45 |
| BG6.0(2.5) | 0.690 (0.902) | 0.304 (0.098) | 0.087 (0.010) | 0.457 (0.423) | 0.392 (0.551) | 0.062 (0.016) | 2.50 | 2.48 | 2.57 | 2.42 |
| BG2.6(2.7) | 0.517 (0.838) | 0.475 (0.162) | 0.041 (0.006) | 0.336 (0.242) | 0.507 (0.677) | 0.115 (0.075) | 2.74 | 2.73 | 2.82 | 2.70 |
| BG0(2.9) | – | – | 0.016 (0.000) | 0.225 (0.094) | 0.571 (0.793) | 0.189 (0.113) | 2.93 | 2.93 | 3.02 | 2.93 |
| BG2.0(2.9) | 0.417 (0.795) | 0.553 (0.205) | 0.017 (0.000) | 0.246 (0.108) | 0.553 (0.791) | 0.183 (0.101) | 2.93 | 2.92 | 2.99 | 2.90 |
| BG3.0(2.9) | 0.444 (0.814) | 0.528 (0.186) | 0.020 (0.000) | 0.252 (0.144) | 0.562 (0.779) | 0.166 (0.077) | 2.93 | 2.91 | 2.93 | 2.87 |
| BG4.0(2.9) | 0.440 (0.805) | 0.554 (0.195) | 0.022 (0.000) | 0.269 (0.118) | 0.539 (0.791) | 0.169 (0.091) | 2.93 | 2.90 | 2.97 | 2.86 |
| BG6.0(2.9) | 0.460 (0.821) | 0.527 (0.179) | 0.028 (0.000) | 0.295 (0.121) | 0.529 (0.778) | 0.149 (0.101) | 2.93 | 2.89 | 2.98 | 2.79 |
Values within parentheses correspond to the populations obtained by NMR. Typical standard deviations of the MD-generated xPn and xSi populations are σ = 0.034 and σ = 0.011, respectively, as estimated from all simulations that involved ≈6000 atoms (see the Supporting Information).
The MD-derived structures also comprise minor amounts of QP2 (xP ≲ 0.03) and QSi0 (xSi ≲ 0.01) groups that are not listed.
Corrected for the NMR-derived minor QP1 contributions.
Obtained from the 29Si NMR-derived {xSin} sets via eq 3.
Obtained from the MD data.
4.3. Silicate and Phosphate Speciations: MD Simulations versus NMR
Here, we present the MD-modeled {QSin} and {QP} speciations and contrast their predictions with the experimental data obtained by MAS NMR.
4.3.1. NBO Distribution among Phosphate Groups
Some of the sparse experimental reports on (semi)quantitative {xP0, xP} populations of Na2O–CaO–SiO2–P2O5 structures concluded that the QP1 groups generally increase with the P2O5 content.16,26,32 In contrast, Grussaute et al.31 reported that the QP populations are independent thereof, at least for x(P2O5) ≤ 2.4 mol % and high silicate network connectivities (2.65–3.0). However, inspection of their 31P NMR data reveals a clear growth of xP1 when N̅BO increases, but this trend was not commented. Mercier et al.32 highlighted a dependence of the QP1 fraction on the SiO2 content of the glass, further inferring that it increased concomitantly with the silicate network polymerization; yet, their glass-series design did not allow for reaching a quantitative xP/N̅BOSi correlation. Such a relationship is established herein.
Table 4 lists the sets of MD-generated {xP0, xP} populations for the present structures. It is gratifying that the following qualitative trends fully accord with the experimental results of section 3.1: (i) The QP1 population grows steadily as N̅BO increases, with the extreme values of xP1 =0.17 and xP ≈0.55 observed for the BG2.6(2.1) and BG(2.9) structures, respectively. (ii) At a fixed silicate network connectivity, the QP1 population remains essentially constant when the P content varies (Table 4). The primary discrepancy between the {xP} sets obtained from 31P NMR and the MD calculations is the significantly lower orthophosphate populations in the modeled structures, whereas the QP1 contributions are consistently 2.5–3 times higher than their experimental counterparts. Noteworthy, the deviations constitute a straightforward scaling, essentially independent of the N̅BO value. Previous MD studies of the Na2O–CaO–SiO2–P2O5 system have focused almost exclusively on the “45S5” composition [i.e., BG2.6(2.1)], where consistently higher xP1 values were observed17,18,44−46,49,50 compared to those (≈0) estimated by 31P NMR.24,30,43 This discrepancy likely reflects slight xP overestimations in MD simulations, and a corresponding potential underestimation by 31P MAS NMR whenever the QP1 population is at the detection level of the technique, such as for 45S5 (see the Supporting Information). The present MD-generated phosphate speciation of BG2.6(2.1) confirm previously modeled results on 45S5,17,18,46,49,50 while our remaining specimens complement the very few MD reports from other Na–Ca–Si–P–O compositions17,18 by systematically exploring a significantly wider compositional range than previously considered.
Because the NMR-derived phosphate speciations solely comprise QP0 and QP groups, there is a direct mapping between xP1 and N̅BO (i.e., the average number of BO atoms per PO4 tetrahedron), whereas the modeled counterparts additionally comprise minor xP2 populations (≲0.03; see the Supporting Information). To gain quantitative insight into the relationship between the QP population and the silicate network polymerization, being of direct concern for designing BG specimens with optimal bioactivity, we fitted the {N̅BOP} set against {N̅BO}. For both the NMR and MD generated data, Figure 4 reveals an approximately linear relationship over the present parameter space {2.1 ≤ N̅BOSi ≤ 2.9; p ≤ 6.0}. This trend persists regardless of whether each NMR/MD derived set {N̅BO} is correlated with the composition-dictated N̅BOSi(nom) values, or with those obtained from either of 29Si NMR [N̅BO(NMRSi)] or MD simulations [N̅BOSi(MD)] (the latter are discussed below). As phase separation tendencies were observed for the BG6.0(2.9) glass (see section 3.1), the linear N̅BO/N̅BOSi correlation likely breaks down for higher values of N̅BO and/or x(P2O5).
Figure 4.
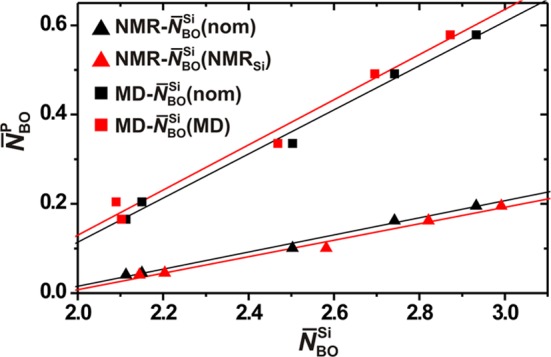
Average number of BO atoms at the phosphate groups (N̅BOP) in the BG structures, plotted against its Si analogue (N̅BO), as obtained either by 29Si NMR experiments [N̅BOSi(NMRSi)], or by MD simulations [N̅BO(MD)]. Averages were taken over all N̅BOP-values within each series of constant N̅BO(nom). Given that the experimental and modeled N̅BOSi values generally differ slightly from their nominal counterparts [N̅BO(nom)], the results of correlating the {N̅BOP(NMR)} and {N̅BO(MD)} set with N̅BOSi(nom) are also provided (red symbols). Straight lines represent best-fit results of each N̅BO/N̅BOSi correlation and method {MD-N̅BO(nom), MD-N̅BOSi(MD), NMR-N̅BO(nom), NMR-N̅BOSi(NMRSi)}; they are associated with the respective R2 correlation coefficients of {0.988, 0.977, 0.989, 0.981}.
4.3.2. NBO Distribution among Silicate Groups
We now focus on the BO/NBO partitioning among the network-forming SiO4 groups, i.e., the n-distribution of {QSin}. The MD-derived {QSi} speciations are contrasted with their experimental counterparts in Table 4. As required and also in full accordance with 29Si NMR, the modeled structures manifest essentially equal {QSin} sets within each glass family of constant N̅BO(nom)-value (regardless of the P content), whereas the QSin distribution shifts progressively toward higher n for increasing silicate network polymerization. For each silicate network, its average number of BO atoms, N̅BO(MD), was calculated according to eq 3. The {N̅BOSi(MD)} set accords well with its nominal {N̅BO(nom)} counterpart, besides a slight but consistent decrease of the MD-generated results when the P content increases; this is readily explained by the presence of significant fractions of QP1 groups that release some Na+/Ca2+ cations relative to the as-assumed scenario of solely orthophosphate species when the present glass compositions were devised (see section 2). However, owing to the overall low P2O5 contents (≤6 mol %), the observed reduction of N̅BO remains very minor.
The major distinction between the experimental and modeled data is the significantly wider {QSin} distributions observed from the latter (Table 4). The NMR/MD derived {QSi} sets are displayed in Figure 5, which also includes results by assuming either a “binary” or “random” (statistical) BO/NBO distribution among the SiO4 groups. The 29Si NMR-derived silicate speciation generally comprises three terms, {QSin–1, QSi, QSin+1}, in agreement with some recent NMR reports on BGs,30,32,33 although earlier studies generally employed spectra deconvolutions into two signals.16,24,27,43 The modeled/experimental {QSi} populations and the binary/random distribution scenarios are next compared by their respective distribution widths σ. As very similar σ-values are observed among the various glass structures for a given distribution type (“NMR/MD/random”; except for the binary scenario that anyway obeys σ ≤ 0.5), only averages over the entire set of samples are considered, which compare as follows:
| 4 |
with their (approximate) values given within parentheses.
Figure 5.
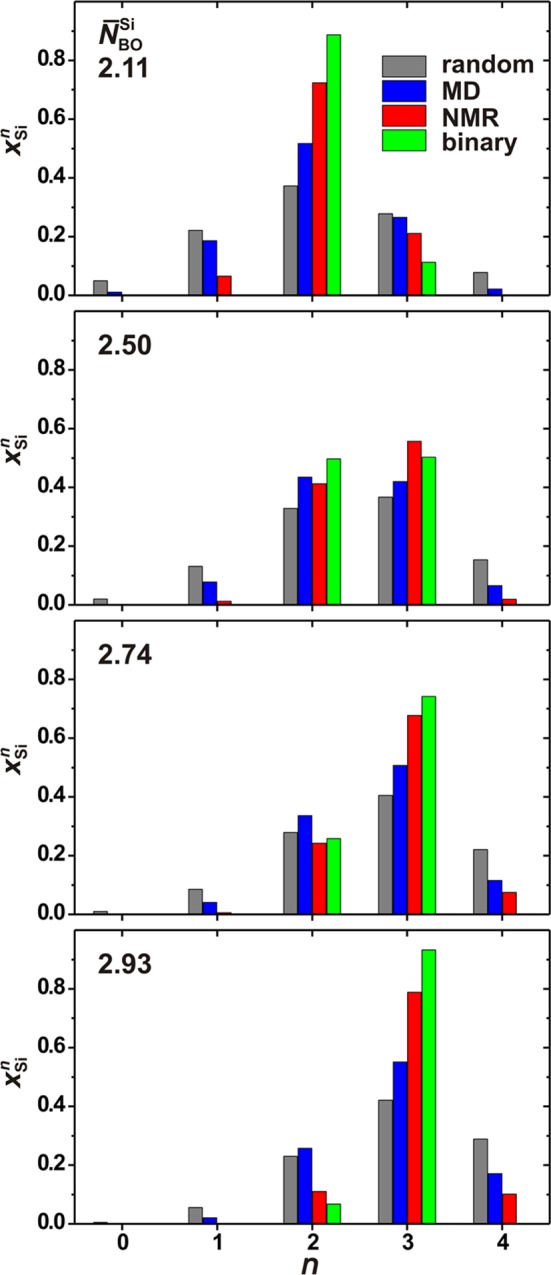
QSin fractional populations determined either by 29Si NMR or MD simulations and plotted versus the number of BO atoms (n) at the SiO4 group. For the glass series BG(2.5) and BG(2.9) that comprise several samples, the {xSi} values represent averages over all members within the series. The results are compared with the predictions from binary and random NBO/BO distributions (calculated from the expressions of ref (21)) for each as-indicated N̅BOSi(nom) value.
We conclude that the experimentally determined {xSin} sets, typically comprising three (significant) contributions, is slightly wider than a binary n-distribution, whereas the MD-derived counterpart is intermediate between the two limiting models, meaning that it is significantly wider than the scenarios of either a binary distribution or that estimated by NMR, but markedly more ordered than predicted from a statistical BO/NBO partitioning among the SiO4 groups.
5. Discussion
5.1. Orthophosphate Content versus Silicate Network Connectivity
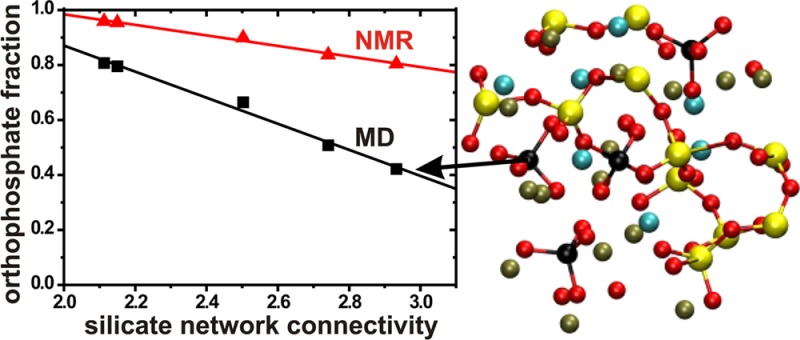
For a wide range of Na2O–CaO–SiO2–P2O5 glass compositions, the present experimental and modeling results (Table 4) firmly establish and extend the inferences by Grussaute et al.31 that for a constant network polymerization, the QP1 population is independent of the P2O5 content of the glass (provided that y(CaO) is fixed31). Hence, the total orthophosphate population [nP; see eq 1] is directly proportional to the net stoichiometric amount of P in the structure, as evidenced by Figure 6 that plots each experimental and modeled number density, ρN and ρN0 of the P atoms and the QP species, respectively, against the P2O5 content of the glass. Two important inferences may be made, both having bearings on future BG glass design:
-
(i)
Considering previous demonstrations that the bioactivity increases with the P content of the BG,6,7,9,22 our present results evidence a direct correlation between the bioactivity and the orthophosphate content of the structure. Further, from the direct nP/bioactivity correlation inferred in refs (15 and 22) follows that the bioactivity increases monotonically with nP0 [see eq 1], thereby establishing the validity of hypothesis B in section 2.
-
(ii)
One may readily design a BGp(N̅BOSi) composition that combines a favorable silicate network polymerization with a high P content, at least for p ≤ 6.0. Hence, for the range of suitable silicate network connectivities 2.0 ≲ N̅BO ≲ 2.6 (discussed further in section 5.2), an optimal bioactivity is attained by arranging the highest possible P content, without altering the N̅BOSi-value significantly, as the majority of the phosphate species are not forming bonds to other SiO4 or PO4 groups, but enter the structure as isolated orthophosphate ions (provided that the glass modifier content is increased accordingly; see section 2 and Table 1).
Figure 6.
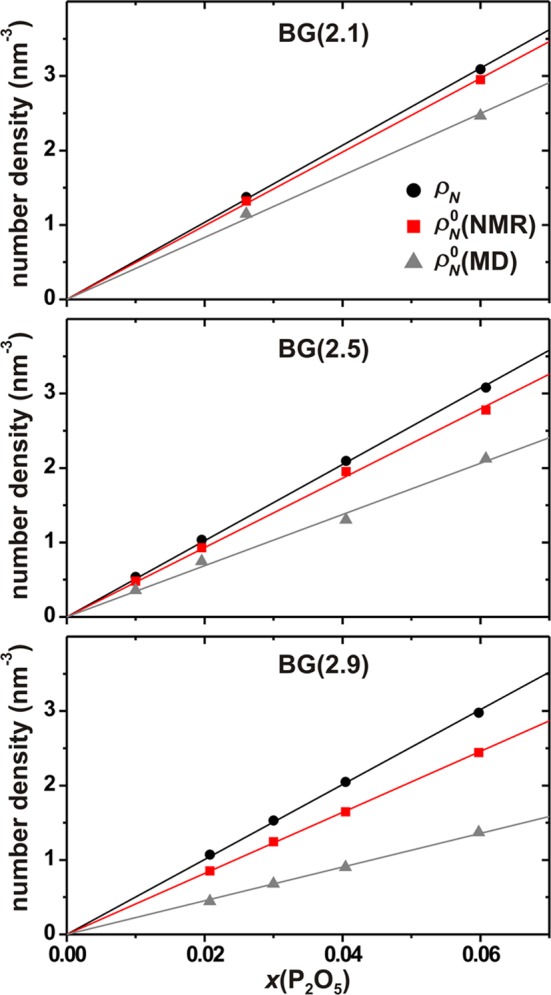
Number density (number of species per nm3) of P atoms (ρN) and orthophosphate groups (ρN0), plotted against the molar fraction of P2O5 for the as-indicated BG(N̅BO) series of glasses. ρN was obtained from the stoichiometric glass composition and its accompanying experimental density ρ (see Table 1), whereas ρN0 = xPρN involves the fractional population xP0 obtained either by 31P NMR or by MD simulations. Straight lines represent best-fit results.
5.2. Bioactivity versus Silicate Network Connectivity
Early work concluded that a bioactive glass composition required a “network connectivity <3”.11,12 This originated from the incorrect assumption of a nonpreferential BO/NBO distribution among Si and P; because P is mainly present as orthophosphate ions, their Na+/Ca2+ consumption then leads to a higher than predicted N̅BOSi-value; to avoid such confusions, we refer specifically to the silicate network connectivity, i.e., N̅BO.15 Yet, the precise N̅BOSi-range that optimizes the bioactivity remains unsettled in the current absence of a systematic investigation.
On the basis of published data from a large series of Na2O–CaO–SiO2–P2O5 compositions,6 we proposed N̅BOSi ≲ 2.7 as a necessary criterion for bioactivity, with optimal N̅BO values predicted at the upper range of 2.5–2.6, beyond which the bioactivity diminishes rapidly.15 According to these predictions, the bioactivity increases from the BG(2.1) to the BG(2.5) glass branch—with the fastest HCA growth expected from the BG6.0(2.5) specimen due to its highest P content—whereas BG2.6(2.7) is anticipated to display a low bioactivity and no member of the BG(2.9) series should give physiological responses. While bioactivity testing is underway for the current glasses, the hypothesis that N̅BOSi ≈ 2.55 provides optimum conditions is supported further by the very recent work of Duée et al.9 They employed “mixture designs” to identify optimal BG candidates with x(P2O5)≤0.05; it amounted in two “best” glass compositions, both incidentally exhibiting N̅BO = {2.56; 2.60}, despite that the silicate network connectivity was not a targeted parameter in their assessments. Noteworthy, the “D47” composition9 (0.215Na2O–0.265CaO–0.470SiO2–0.050P2O5) is almost identical to a “BG5.0(2.5)” glass in our nomenclature, i.e., 0.214Na2O–0.278CaO–0.458SiO2–0.050P2O5. The onset of HCA formation from D47 was twice as rapid as for the “45S5 Bioglass”,9 hence providing further support for our stated optimal N̅BOSi-range.
Another question is if the bioactivity-increase across the range 2.0 ≲ N̅BOSi ≲ 2.6 and its sharp reduction as N̅BO grows further can be rationalized from a structural viewpoint. The {QSin} speciations of Table 3 reveal that the most bioactive glasses exhibit structures built primarily by QSi and QSi3 groups, i.e., representing interconnected chain/ring motifs with a significant cross-linking: if the latter is negligible, then the BG degrades too rapidly in its contact with body fluids, thereby preventing significant HCA formation.6,9 In contrast, a too high polymerization of the network impedes its degradability in solutions due to the large number of Si–O–Si bonds requiring hydrolysis, coupled with a diminished Na+/Ca2+ reservoir and accompanying reduced ion dissolution associated with the (too) Si-rich composition. Apparently, the best compromise between these two limits occurs when the QSi and QSi3 populations are nearly matched, i.e., when N̅BO≈2.5 (see Table 4), as opposed to the (perhaps more intuitive) scenario of N̅BOSi ≈ 2.0 that reveals predominantly QSi moieties. This is one reason why the link between the glass solubility and the bioactivity is not strict.
As the network polymerization increases across the range 2.1–2.9, the QSi3 population grows steadily at the expense of its QSi counterpart; while this effect alone cannot explain the nonmonotonic dependence of the bioactivity on N̅BOSi, the bioactivity might reduce when QSi reaches above some ”critical” level (≥70%, according to Table 3). Yet, the emergence of QSi4 structural groups is apparently a more accurate indicator for the onset of “non-bioactivity”. While QSi motifs are essentially absent in the BG structures associated with N̅BOSi ≤ 2.5, their population increases steeply for more polymerized networks; they constitute ≈10% out of the total SiO4 speciation for the nonbioactive BG(2.9) compositions (Table 3). The loss of bioactivity may be understood from the significantly higher cross-linking invoked by the QSi groups and the accompanying local depletion of glass modifier ions; altogether these features create structural portions that more strongly resists degradation in aqueous medium compared to the chain/sheet-like motifs prevailing in the bioactive glass structures. These inferences were made from the NMR-derived {QSin} speciations; however, as in the case of the QP populations, this trend is also mirrored by the modeled structures (Table 4).
5.3. The Roles of Na and Ca
Once fixating both the P content and silicate network connectivity, the nNa/nCa molar ratio constitutes the remaining free parameter for a given Na2O–CaO–SiO2–P2O5 glass formulation. Despite influencing the bioactivity to a lesser extent than the {N̅BOSi, nP} pair, the relative Na/Ca content is worth discussing as its bearing on the bioactivity yet remains to be systematically assessed, particularly considering indications that the bioactivity-boost for increasing P content may be strictly monotonic only over a range of relative Na/Si amounts within 0.8 ≲ nNa/nSi ≲ 1.2.15
Keeping either a large Na or Ca content has both its pros and cons. Large amounts of Ca are beneficial for primarily three reasons: (i) Being a component of HCA, higher dissolution rates of Ca improves the apatite supersaturation.2,3,5 (ii) The {xP0, xP} values depend to some extent on the nNa/nCa ratio, with the desirable orthophosphate fraction increasing concurrently with the Ca content.31 (iii) Large amounts of Na+ ions induces cytotoxicity.9,51 In contrast, arranging a large Na content is favorable for (i) facilitating the preparation of P-rich glasses by lowering the melting temperature, and particularly, (ii) enhancing the glass solubility; the higher Na+ mobility relative to Ca2+ overall accelerates the surface reactions and elevates the local pH, altogether stimulating the calcium phosphate formation.2,3,5,29
Noteworthy, as both the Ca2+ and PO43– ions are dispersed throughout the glass matrix,24−27 an increased glass degradation also facilitates their release (vide infra). We therefore hypothesize that a relatively high nNa/nCa ratio within 1.0–2.0 (i.e., 0.3≤y(CaO)≤0.5) may release an otherwise stricter lower bound on the N̅BOSi-value, thereby allowing the combination of a high bioactivity with a relatively condensed silicate network exhibiting N̅BO ≈2.5 (see section 5.2). Furthermore, given that each Na+ and Ca2+ species associate with both Si and P (section 4.1), altering the nNa/nCa ratio offers a route to tune the glass–surface reactivity, and thereby the BG degradation, without any significant bearings on the N̅BOSi-value. Note, however, that increasing the relative amount of Ca (i.e., decreasing nNa/nCa) may be favorable as it elevates xPslightly,31 and thereby the net orthophosphate content [see eq 1].
5.4. Recommendations for Bioactivity Optimizations
The present results suggest that future bioactivity-composition assessments should target the parameter-triplet {N̅BOSi, nP, nNa/nCa} in the search for optimal BG compositions. Our suggested {N̅BO, nP, nNa/nCa} parametrization of the glass composition provides more transparent insight into the composition-bioactivity relationships compared with the standard formulations expressed as oxide equivalents. The bioactivity is mainly dictated by the {N̅BOSi, nP} pair, and to a lesser extent by the exact nNa/nCa ratio. Hence, it is expected to be optimized at the highest incorporable P2O5 content around the parameter space {N̅BO ≈ 2.55, 1 ≲ nNa/nCa ≲ 2}.15 The feasibility of preparing P-richer compositions than x(P2O5) > 0.06 needs to be tested. Future work must also more quantitatively define our suggested weak interdependence between these bioactivity descriptors. Additionally, the “optimal parameter-spaces” stated herein are strictly only applicable to Na–Ca–Si–P–O glasses (and limiting systems thereof), and their transferability to related M–M′–Si–P–O glasses remains to be explored.
6. Conclusions
The alterations of the QSin and QP speciations in a series of 13 glasses were for the first time explored systematically over a wide compositional range within the Na2O–CaO–SiO2–P2O5 system, by using a combination of atomistic MD simulations and 31P/29Si solid-state NMR experiments; both techniques generally revealed equivalent qualitative trends. Our glass series design allowed for an independent probing of the structural changes accompanying a variation in either the P content of the BG or its silicate network connectivity; the series encompassed both bioactive (2.1 ≤ N̅BOSi ≤ 2.5) and nonbioactive glass compositions (N̅BO > 2.7). When N̅BOSi increases from 2.1 to 2.9, both the MD/NMR-derived {QSi} speciations reveal net QSi2 → QSi conversions (as expected). However, despite that the network polymerization increases accordingly throughout the entire range of bioactive glass compositions (N̅BOSi ≲ 2.7), the progressive formation of QSi groups itself does not obviously correlate with the transition from bioactive to nonbioactive compositions, which merely coincides with the emergence of non-negligible (≳10%) contributions of QSi4 groups; the accompanying markedly enhanced structural cross-linking from four BO atom per SiO4 tetrahedron (as opposed to 1–3 BO) together with a local depletion of readily released Na+/Ca2+ cations strongly diminishes the glass degradation in aqueous media.
If the silicate network connectivity of the BG remains constant, the {QP0, QP} fractional populations are independent of the amount of P2O5 (at least for x(P2O5) ≤ 0.06), thereby providing a direct link between the orthophosphate content and nP. In contrast, if nP remains constant, the fraction of QP0 species decreases linearly as N̅BO increases. Fortunately, for the network connectivity-range 2.0 ≲ N̅BOSi ≲ 2.6 that encompasses nearly all bioactive glass compositions, QP moieties constitute ≳80% of the total phosphate speciation, thereby rationalizing earlier statements15,22 that the bioactivity increases monotonically with the P content of the BG (provided that the glass modifier reservoir is sufficiently large to charge-balance all QP0 groups). By showing that the QP concentration increases with the P2O5 molar fraction for a fixed silicate network connectivity, the present results prove it possible to design highly bioactive glasses that combine a favorable network connectivity with large amounts of readily released orthophosphate ions, thereby promoting both a rapid degradation of the glass network and a fast dissolution of biologically active ions.52,53 The structural role of P is the key factor: the majority of all P species enter the structure as orthophosphate groups detached from the glass network; their fast dissolution enhances the bioactivity relative to a glass with the same network connectivity but a lower P content.
Future composition/bioactivity assessments/optimizations should target the parameter-set {N̅BOSi, nP, nNa/nCa}. Each such triplet translates into a unique Na2O–CaO–SiO2–P2O5 glass composition, whose bioactivity may be roughly assessed by inspection, as each parameter influences the glass bioactivity in a predictable manner, while their effects may be tuned almost independently from each other. However, future studies must better quantify the expectedly weak correlation between the {xP, xP1} populations and the nNa/nCa ratio,31 as well as exploring the degree of correlation between the bioactivity of the glass and its solubility. As the latter may be changed by varying either the silicate network connectivity or the Na content of the BG, whose effects are likely synergetic, the N̅BO-range providing high bioactivities is presumably not completely decoupled from the nNa/nCa ratio: rather, we suggest that the increased solubility associated with Na-rich BGs may (slightly) alter the N̅BOSi-values defining each transition between high/low/nonbioactive compositions. These ideas are currently being explored.
Acknowledgments
This work was supported by the Swedish Research Council (contract 2010-4943) and the Faculty of Sciences at Stockholm University. A.T. acknowledges support (University Research Fellowship) from the U.K.’s Royal Society. NMR equipment Grants from the Swedish Research Council, and the Knut and Alice Wallenberg Foundation are gratefully acknowledged. We thank Shahriar Iftekhar and Kirill Okhotnikov for their initial help and input regarding glass synthesis and MD simulations, respectively.
Supporting Information Available
Further information about the NMR spectra and the deconvolution procedures, discussion about the detection of QP1 groups by 31P NMR, as well as additional MD data and discussions thereof. This material is available free of charge via the Internet at http://pubs.acs.org.
The authors declare no competing financial interest.
Supplementary Material
References
- Hench L. L.; Splinter R. J.; Allen W. C.; Greenlee T. K. Bonding Mechanisms at the Interface of Ceramic Prosthetic Materials. J. Biomed. Mater. Res. 1971, 2, 117–141. [Google Scholar]
- Hench L. L. Bioceramics–from Concept to Clinic. J. Am. Ceram. Soc. 1991, 74, 1487–1510. [Google Scholar]
- Jones J. R. Review of Bioactive Glass: From Hench to Hybrids. Acta Biomater. 2013, 9, 4457–4486. [DOI] [PubMed] [Google Scholar]
- Hench L. L.; Polak J. M. Third-Generation Biomedical Materials. Science 2002, 295, 1014–1017. [DOI] [PubMed] [Google Scholar]
- Andersson Ö. H.; Karlsson K. H. On the Bioactivity of Silicate Glass. J. Non-Cryst. Solids 1991, 129, 145–151. [Google Scholar]
- Lebecq I.; Désanglois F.; Leriche A.; Follet-Houttemane C. Compositional Dependence on the In Vitro Bioactivity of Invert or Conventional Bioglasses in the Si-Ca-Na-P System. J. Biomed. Mater. Res. A 2007, 83, 156–168. [DOI] [PubMed] [Google Scholar]
- O’Donnell M. D.; Watts S. J.; Hill R. G.; Law R. V. The Effect of Phosphate Content on the Bioactivity of Soda-Lime-Phosphosilicate Glasses. J. Mater. Sci: Mater. Med. 2009, 20, 1611–1618. [DOI] [PubMed] [Google Scholar]
- Watts S. J.; Hill R. G.; O’Donnell M. D.; Law R. V. Influence of Magnesia on the Structure and Properties of Bioactive Glasses. J. Non-Cryst. Solids 2010, 356, 517–524. [Google Scholar]
- Duée C.; Grattepanche-Lebecq I.; Désanglois F.; Follet-Houttemane C.; Chai F.; Hildebrand H. F. Predicting Bioactive Properties of Phosphosilicate Glasses Using Mixture Designs. J. Non-Cryst. Solids 2013, 362, 47–55. [Google Scholar]
- Christie J. K.; Tilocca A. Molecular Dynamics Simulations and Structural Descriptors of Radioisotope Glass Vectors for in Situ Radiotherapy. J. Phys. Chem. B 2012, 116, 12614–12620. [DOI] [PubMed] [Google Scholar]
- Strnad Z. Role of the Glass Phase in Bioactive Glass-Ceramics. Biomaterials 1992, 13, 317–321. [DOI] [PubMed] [Google Scholar]
- Hill R. An Alternative View of the Degradation of Bioglass. J. Mater. Sci. Lett 1996, 15, 1122–1125. [Google Scholar]
- Stevels J. M. Neue Erkenntnisse Uber Die Struktur Des Glases. Philips Techn. Rdsch 1960, 9/10, 337–349. [Google Scholar]
- O’Donnell M. D.; Watts S. J.; Law R. V.; Hill R. G. Effect of P2O5 Content in Two Series of Soda Lime Phosphosilicate Glasses on Structure and Properties - Part I: NMR. J. Non-Cryst. Solids 2008, 354, 3554–3560. [Google Scholar]
- Edén M. The Split Network Analysis for Exploring Composition-Structure Correlations in Multi-Component Glasses: I. Rationalizing Bioactivity-Composition Trends of Bioglasses. J. Non-Cryst. Solids 2011, 357, 1595–1602. [Google Scholar]
- Elgayar I.; Aliev A. E.; Boccaccini A. R.; Hill R. G. Structural Analysis of Bioactive Glasses. J. Non-Cryst. Solids 2005, 351, 173–183. [Google Scholar]
- Tilocca A.; Cormack A. N. Structural Effects of Phosphorus Inclusion in Bioactive Silicate Glasses. J. Phys. Chem. B 2007, 111, 14256–14264. [DOI] [PubMed] [Google Scholar]
- Tilocca A.; Cormack A. N.; de Leeuw N. H. The Structure of Bioactive Silicate Glasses: New Insight from Molecular Dynamics Simulations. Chem. Mater. 2007, 19, 95–103. [Google Scholar]
- Tilocca A. Structural Models of Bioactive Glasses from Molecular Dynamics Simulations. Proc. R. Soc. A 2009, 465, 1003–1027. [Google Scholar]
- Gunawidjaja P. N.; Lo A. Y. H.; Izquierdo-Barba I.; García A.; Arcos D.; Stevensson B.; Grins J.; Vallet-Regí M.; Edén M. Biomimetic Apatite Mineralization Mechanisms of Mesoporous Bioactive Glasses as Probed by Multinuclear 31P, 29Si, 23Na and 13C Solid-State NMR. J. Phys. Chem. C 2010, 114, 19345–19356. [Google Scholar]
- Edén M.; Sundberg P.; Stålhandske C. The Split Network Analysis for Exploring Composition-Structure Correlations in Multi-Component Glasses: II. Multinuclear NMR Studies of Alumino-Borosilicates and Glass-Wool Fibers. J. Non-Cryst. Solids 2011, 357, 1587–1594. [Google Scholar]
- Hill R. G.; Brauer D. S. Predicting the Bioactivity of Glasses Using the Network Connectivity or Split Network Models. J. Non-Cryst. Solids 2011, 357, 3884–3887. [Google Scholar]
- Gunawidjaja P. N.; Mathew R.; Lo A. Y. H.; Izquierdo-Barba I.; García A.; Arcos D.; Vallet-Regí M.; Edén M. Local Structures of Mesoporous Bioactive Glasses and Their Surface Alterations In Vitro: Inferences from Solid-State Nuclear Magnetic Resonance. Philos. Trans. R. Soc. A 2012, 370, 1376–1399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin R. A.; Twyman H. L.; Rees G. J.; Smith J. M.; Barney E. R.; Smith M. E.; Hanna J. V.; Newport R. J. A Structural Investigation of the Alkali Metal Site Distribution within Bioactive Glass Using Neutron Diffraction and Multinuclear Solid State NMR. Phys. Chem. Chem. Phys. 2012, 14, 12105–12113. [DOI] [PubMed] [Google Scholar]
- Mathew R.; Turdean-Ionescu C.; Stevensson B.; Izquierdo-Barba I.; García A.; Arcos D.; Vallet-Regí M.; Edén M. Direct Probing of the Phosphate-Ion Distribution in Bioactive Silicate Glasses by Solid-State NMR: Evidence for Transitions between Random/Clustered Scenarios. Chem. Mater. 2013, 25, 1877–1885. [Google Scholar]
- Dupree R.; Holland D.; Mortuza M. G.; Collins J. A.; Lockyer M. W. G. An MAS NMR Study of Network - Cation Coordination in Phosphosilicate Glasses. J. Non-Cryst. Solids 1988, 106, 403–407. [Google Scholar]
- Lockyer M. W. G.; Holland D.; Dupree R. NMR Investigation of the Structure of some Bioactive and Related Glasses. J. Non-Cryst. Solids 1995, 188, 207–219. [Google Scholar]
- Ebisawa Y.; Kokubo T.; Ohura K.; Yamamuro T. Bioactivity of CaO·SiO2-Based Glasses: In Vitro Evaluation. J. Mater. Sci: Mater. Med. 1990, 1, 239–244. [Google Scholar]
- Karlsson K. H.; Fröberg K.; Ringbom T. A Structural Approach to Bone Adhering of Bioactive Glasses. J. Non-Cryst. Solids 1989, 112, 69–72. [Google Scholar]
- Pedone A.; Charpentier T.; Malavasi G.; Menziani M. C. New Insights into the Atomic Structure of 45S5 Bioglass by Means of Solid-State NMR Spectroscopy and Accurate First-Principles Simulations. Chem. Mater. 2010, 22, 5644–5652. [Google Scholar]
- Grussaute H.; Montagne L.; Palavit G.; Bernard J. L. Phosphate Speciation in Na2O-CaO-P2O5-SiO2 and Na2O-TiO2-P2O5-SiO2 Glasses. J. Non-Cryst. Solids 2000, 263, 312–317. [Google Scholar]
- Mercier C.; Follet-Houttemane C.; Pardini A.; Revel B. Influence of P2O5 Content on the Structure of SiO2-Na2O-CaO-P2O5 Bioglasses by 29Si and 31P MAS-NMR. J. Non-Cryst. Solids 2011, 357, 3901–3909. [Google Scholar]
- Fayon F.; Duée C.; Poumeyrol T.; Allix M.; Massiot D. Evidence of Nanometric-Sized Phosphate Clusters in Bioactive Glasses as Revealed by Solid-State 31P NMR. J. Phys. Chem. C 2013, 117, 2283–2288. [Google Scholar]
- Tilocca A. Structure and Dynamics of Bioactive Phosphosilicate Glasses and Melts from Ab Initio Molecular Dynamics Simulations. Phys. Rev. B 2007, 76, 224202. [Google Scholar]
- Fujikura K.; Karpukhina N.; Kasuga T.; Brauer D. S.; Hill R. G.; Law R. V. Influence of Strontium Substitution on Structure and Crystallisation of Bioglass 45S5®. J. Mater. Chem. 2012, 22, 7395–7402. [Google Scholar]
- Tilocca A. Models of Structure, Dynamics and Reactivity of Bioglasses: A Review. J. Mater. Chem 2010, 20, 6848–6858. [Google Scholar]
- Smith W.; Forester T. R. DL_POLY_2.0: A General-Purpose Parallel Molecular Dynamics Simulation Package. J. Mol. Graphics 1996, 14, 136–141. [DOI] [PubMed] [Google Scholar]
- Todorov I. T.; Smith W.; Trachenko K.; Dove M. T. DL_POLY_3: New Dimensions in Molecular Dynamics Simulations via Massive Parallelism. J. Mater. Chem. 2006, 16, 1911–1918. [Google Scholar]
- Tilocca A.; de Leeuw N. H.; Cormack A. N. Shell-Model Molecular Dynamics Calculations of Modified Silicate Glasses. Phys. Rev. B 2006, 73, 104209. [Google Scholar]
- MacKenzie K. J. D.; Smith M. E.. Multinuclear Solid-State NMR of Inorganic Materials; Pergamon Press: Amsterdam, 2002. [Google Scholar]
- Leonova E.; Izquierdo-Barba I.; Arcos D.; Lopez-Noriega A.; Hedin N.; Vallet-Regí M.; Edén M. Multinuclear Solid-State NMR Studies of Ordered Mesoporous Bioactive Glasses. J. Phys. Chem. C 2008, 112, 5552–5562. [Google Scholar]
- Mathew R.; Gunawidjaja P. N.; Izquierdo-Barba I.; Jansson K.; García A.; Arcos D.; Vallet-Regí M.; Edén M. Solid-State 31P and 1H NMR Investigations of Amorphous and Crystalline Calcium Phosphates Grown Biomimetically from a Mesoporous Bioactive Glass. J. Phys. Chem. C 2011, 115, 20572–20582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FitzGerald V.; Pickup D. M.; Greenspan D.; Sarkar G.; Fitzgerald J. J.; Wetherall K. M.; Moss R. M.; Jones J. R.; Newport R. J. A Neutron and X-Ray Diffraction Study of Bioglass® with Reverse Monte Carlo Modelling. Adv. Funct. Mater. 2007, 17, 3746–3753. [Google Scholar]
- Xiang Y.; Du J. Effect of Strontium Substitution on the Structure of 45S5 Bioglasses. Chem. Mater. 2011, 23, 2703–2717. [Google Scholar]
- Du J.; Xiang Y. Effect of Strontium Substitution on the Structure, Ionic Diffusion and Dynamic Properties of 45S5 Bioactive Glasses. J. Non-Cryst. Solids 2012, 358, 1059–1071. [Google Scholar]
- Malavasi G.; Pedone A.; Menziani M. C. Study of the Structural Role of Gallium and Aluminum in 45S5 Bioactive Glasses by Molecular Dynamics Simulations. J. Phys. Chem. B 2013, 117, 4142–4150. [DOI] [PubMed] [Google Scholar]
- Murdoch J. B.; Stebbins J. F.; Carmichael I. S. E. High-Resolution 29Si NMR-Study of Silicate and Aluminosilicate Glasses - the Effect of Network-Modifying Cations. Am. Mineral. 1985, 70, 332–343. [Google Scholar]
- Maekawa H.; Maekawa T.; Kawamura K.; Yokokawa T. The Structural Groups of Alkali Silicate-Glasses Determined from 29Si MAS-NMR. J. Non-Cryst. Solids 1991, 127, 53–64. [Google Scholar]
- Tilocca A. Short- and Medium-Range Structure of Multicomponent Bioactive Glasses and Melts: An Assessment of the Performances of Shell-Model and Rigid-Ion Potentials. J. Chem. Phys. 2008, 129, 084504. [DOI] [PubMed] [Google Scholar]
- Pedone A.; Malavasi G.; Menziani M. C. Computational Insight into the Effect of CaO/MgO Substitution on the Structural Properties of Phospho-Silicate Bioactive Glasses. J. Phys. Chem. C 2009, 113, 15723–15730. [Google Scholar]
- Wallace K. E.; Hill R. G.; Pembroke J. T.; Brown C. J.; Hatton P. V. Influence of Sodium Oxide Content on Bioactive Glass Properties. J. Mater. Sci: Mater. Med. 1999, 10, 697–701. [DOI] [PubMed] [Google Scholar]
- Xynos I. D.; Edgar A. J.; Buttery L. D. K.; Hench L. L.; Polak J. M. Gene-Expression Profiling of Human Osteoblasts Following Treatment with the Ionic Products of Bioglass 45S5 Dissolution. J. Biomed. Mater. Res. 2001, 55, 151–157. [DOI] [PubMed] [Google Scholar]
- Tilocca A.; Cormack A. N. The Initial Stages of Bioglass Dissolution: A Car-Parrinello Molecular-Dynamics Study of the Glass-Water Interface. Proc. R. Soc. A 2011, 467, 2102–2111. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.