

Abstract
Background
The interest on magnesium (Mg) has grown since clinical studies have shown the efficacy of Mg-containing phosphate binders. However, some concern has arisen for the potential effect of increased serum Mg on parathyroid hormone (PTH) secretion. Our objective was to evaluate the direct effect of Mg in the regulation of the parathyroid function; specifically, PTH secretion and the expression of parathyroid cell receptors: CaR, the vitamin D receptor (VDR) and FGFR1/Klotho.
Methods
The work was performed in vitro by incubating intact rat parathyroid glands in different calcium (Ca) and Mg concentrations.
Results
Increasing Mg concentrations from 0.5 to 2 mM produced a left shift of PTH–Ca curves. With Mg 5 mM, the secretory response was practically abolished. Mg was able to reduce PTH only if parathyroid glands were exposed to moderately low Ca concentrations; with normal–high Ca concentrations, the effect of Mg on PTH inhibition was minor or absent. After 6-h incubation at a Ca concentration of 1.0 mM, the expression of parathyroid CaR, VDR, FGFR1 and Klotho (at mRNA and protein levels) was increased with a Mg concentration of 2.0 when compared with 0.5 mM.
Conclusions
Mg reduces PTH secretion mainly when a moderate low calcium concentration is present; Mg also modulates parathyroid glands function through upregulation of the key cellular receptors CaR, VDR and FGF23/Klotho system.
Keywords: calcium-sensing receptor, magnesium, parathyroid glands, PTH, vitamin D receptor
INTRODUCTION
The production of parathyroid hormone (PTH) is essential for calcium (Ca) homeostasis and the maintenance of normal mineral metabolism. Parathyroid cells have cell-surface calcium-sensing receptors (CaRs), which enable them to respond to changes in extracellular Ca [1, 2]; thus, even small changes in extracellular Ca induce rapid changes in PTH secretion. Calcitriol and FGF23 also decrease parathyroid function through their specific parathyroid cell receptors: the vitamin D receptor (VDR) and the FGFR-Klotho receptor system, respectively. In vivo and in vitro studies have also shown a direct effect of phosphate on PTH synthesis and secretion, although a specific cell sensor for phosphate has not been identified, yet.
The activation of the CaR results in Gq/11-dependent activation of phosphatidylinositol-specific phospholipase C, cytosolic Ca increase, protein kinase C (PKC) activation [3] and triggering of the PLA2-AA-MAPKase pathway [4, 5] that regulates PTH synthesis and other essential proteins such as CaR, VDR, Klotho and FGFR1 [6–9].
Although Ca is the main CaR agonist, other divalent and trivalent cations are also able to activate the CaR [2]. For example, an increase in extracellular magnesium (Mg) inhibits PTH secretion and reduces agonist-stimulated cAMP accumulation from parathyroid cells [10–13]. In dialysis patients, high serum Mg concentrations have been associated with low serum PTH levels. This inverse relationship between extracellular levels of Mg and PTH led to the notion of an inhibitory effect of Mg on PTH secretion in these patients [2, 14–17]. However, the PTH secretory response to changes in extracellular Mg concentration has not been formally evaluated in patients.
The interest on Mg has grown since clinical studies have shown the efficacy of Mg-containing phosphate binders [18], and some concern has arisen related to the potential effect of a moderate increase in serum Mg on PTH secretion. In patients with normal serum Ca concentration and controlled PTH levels, it is not clear whether an increase in Mg may elicit an over suppression of PTH. Therefore, one specific question that should be addressed is whether changes in serum Mg have the same impact on PTH secretion when parathyroid cells are exposed to low, normal or high serum Ca concentrations.
The aim of this in vitro study was to evaluate the direct effect of Mg on the regulation of the parathyroid function using intact rat parathyroid glands. Specifically, the present work investigates the effect of Mg at different Ca concentrations on PTH secretion and on the expression of the parathyroid cell receptors CaR, VDR and FGFR1/Klotho.
MATERIALS AND METHODS
Animals
Wistar rats (250 g body wt) were fed a diet containing 1.1% Ca and 0.8% phosphorus and vitamin D (1 IU/g). All animals received optimal care in compliance with the ‘Principles of Laboratory Animal Care,’ formulated by the Spanish National Society for Medical Research and the ‘Guide for the Care and Use of Laboratory Animals’, prepared by the National Academy of Sciences. The experimental protocols were reviewed and approved by the Ethics Committee for Animal Research of the Universidad de Cordoba.
Parathyroid gland culture
Rat parathyroid glands were obtained as previously described [19]. Briefly, rats were anaesthetized with thiopental sodium (50 mg/kg ip). Blood was drained by aortic puncture, and within 2 min the parathyroid glands were dissected free of the thyroid gland and removed.
Intact rat parathyroid glands were placed inside a nylon basket in individual wells containing 2 mL of buffered (pH 7.4) incubation medium (125 mM NaCl, 5.9 mM KCl, 1 mM sodium pyruvate, 4 mM glutamine, 12 mM glucose and 25 mM HEPES with 0.1 IU/mL human insulin, 0.1% bovine serum albumin, 100 IU/mL penicillin G, and 100 mg/mL streptomycin). A phosphate concentration of 1 mM was achieved by addition of 1:2 NaH2PO4–Na2HPO4. Calcium concentrations ranging from 0.8 to 1.5 mM (measured by a selective electrode Spotlyte Ca2+/pH Analyzer, Menarini Diagnostics, Barcelona, Spain) was achieved by addition of CaCl2. Mg was added as MgCl2·6H20 to achieve Mg concentrations ranging from 0.5 to 5.0 mM. The glands were maintained at 37°C with constant rocking and shaking (model AOS-0, SBS Instruments, Badalona, Spain). Cell viability after the experiment was >80%. All chemical products were obtained from Sigma (St Louis, MO).
PTH secretion
After extraction, rat parathyroid glands were stabilized in incubation medium at 1.25 mM Ca for 6 h. Then, the combined effect of changes in Mg and Ca concentrations on PTH secretion was determined. The parathyroid glands were consecutively incubated for a 1 h period in Ca concentrations of 0.8, 1.0, 1.2 and 1.5 mM. The same experiment was repeated using Mg concentrations of 0.5, 1.0, 2.0 and 5.0 mM. At the end of the incubation period, an aliquot of the medium was frozen at −20°C for PTH measurement. Intact rat PTH levels were quantified using an Elisa kit (Immutopics, San Clemente, CA). Set points of the PTH-Ca curve were calculated as the concentration of Ca corresponding to the midrange between the values of maximal and minimal PTH secretion.
mRNA expression
Parathyroid glands were incubated for 6 h at 0.8, 1.0 and 1.5 mM Ca in presence of physiological (0.5 mM) or elevated (2 mM) Mg level. At the end of the incubation period, the glands were freeze-dried and stored at −80°C for later PTH mRNA, VDR mRNA, CaR mRNA, FGFR mRNA and klotho mRNA measurements. For RNA isolation, 1 mL of phenol-guanidine isothiocyanate solution (Tri-Reagent, Sigma) was added to the glands.
The glands were exposed to ultrasound for 5 min at 4°C to allow for complete cell rupture. Thereafter, total RNA was extracted following a modification of the previous protocol [20]. Extracted total RNA was dissolved in nuclease-free water (Sigma, St. Louis, MO) and heated for 10 min at 60°C. Total RNA was quantified by spectrophotometry (NanoDrop, Wilmington, DE). CaR, VDR, FGFR and Klotho mRNA expression were determined by real-time RT-PCR, and beta-actin mRNA levels were assessed as internal controls (SensiMix SYBR No-ROX One-Step Kit, Bioline, London, UK). Quantitative real-time reverse transcriptase-PCR (Light cycler; Roche Diagnostics, Basel, Switzerland) was performed using specific primers and 50 ng of total RNA/sample. Table 1 shows the sequences of the primers.
Table 1.
Primers used for qRT-PCR
| Primer | Forward | Reverse |
|---|---|---|
| CaR | 5′-TGGAGAGACAGATGCGAGTG-3′ | 5′-GTC CAC GCC AGA AAC TCA AT-3′ |
| VDR | 5′-ACAGTCTGAGGCCCAAGCTA-3′ | 5′-TCCCTGAAG TCAGCGTAGGT-3′ |
| Β-actin | 5′-TGTCACCAACTGGGACGATATGGA G-3′ | 5′-ACAATGCCAGTGGTACGACCAGA-3′ |
| PTH | 5′-TTGTCTCCTTACCCAGGCAGAT-3′ | 5′-TTTGCCCAGGTTGTGCATAA-3′ |
| FGFR1 | 5′-CAATGTCTCAGATGCACTGCCA-3′ | 5′-ACAGGCCTACGGTTTGGTTTG-3′ |
| Klotho | 5′-GAAAATGGCTGGTTTGTCTCG-3′ | 5′-CCTGATGGCTTTTAAGCTTTC-3′ |
Immunohistochemistry
After 6 h of incubation with normal (0.5 mM) or high (2 mM) Mg and 0.8, 1.0 or 1.5 mM Ca, the parathyroid glands were fixed in 4% formalin. Three-micrometre sections were deparaffinized and incubated in 0.3% H2O2 in methanol for 30 min. Then, sections were microwave-treated in 0.01 mmol/L citrate buffer (pH 6) for 20 min. Sections were blocked with goat serum 10% for 40 min, then the sections were incubated overnight at 4°C in a humidified chamber with primary affinity-purified rat anti-VDR mAb (Chemicon Int., Temecula, CA; 1:100 dilution), mouse anti-FGFR1 mAb (GeneTex, Irvine, CA; 1:200 dilution), mouse anti-CaR mAb (Abcam, Cambridge, UK; 1:500 dilution) or rabbit anti-Klotho antibody (Alpha Diagnostic, Int., San Antonio, TX; 1:50 dilution). After rinsing, the sections were incubated for 40 min at room temperature with a peroxidase-labelled polymer conjugated to goat antimouse/rabbit Igs and treated with 3,3′ diaminobenzidine-tetrachloride chromogen solution for 10 to 20 min (En-Vision + System HRP [DAB]; DakoCytomation, Glostrup, Denmark; 1:100 dilution). Every step was followed by three washes with phosphate-buffered saline for 5 min. Sections were counterstained with haematoxylin (Dako). Immunoreactivity was assessed using NIH image freeware 1.62. Distinct positive staining was quantified in randomly selected areas on each specimen, in a blinded manner, over a minimum of five fields in more than three sections. The expression of CaR, VDR, Klotho and FGR1 was measured by calculating the average optical density (OD) per section of tissue by dividing the sum integrated OD by the sum area (background staining was subtracted from this value). To avoid the interference of nonspecific positive staining, plots of <30 pixels were excluded.
Statistical analysis
Results are expressed as the mean ± SEM (n = 9 in each group: three different experiments with three repetitions). The difference between mean values for three or more groups was assessed by one-way analysis of variance (ANOVA) followed by post hoc Duncan analysis. The difference between means for two different groups was determined by t test. A P value of <0.05 was considered significant. These analyses were performed with the assistance of a computer program (SPSS 15.0, Chicago, IL).
RESULTS
The effect of magnesium on PTH secretion in vitro
Results from in vitro testing of PTH secretory response to changes in Ca and Mg are presented in Figure 1. Figure 1A shows the PTH results in pg/mL. Furthermore, in order to standardize the change in PTH and avoid small differences in PTH values attributable to minimal differences in the total amount of parathyroid tissue in the 10 glands of each well, the results are also presented as percent versus basal calcium in Figure 1B. Incubation of parathyroid glands with increasing Ca concentrations produced a concentration-dependent decrease in PTH values. Increasing Mg concentrations from 0.5 to 1 and 2 mM produced a left shift of the PTH-Ca curves. Thus, the set points of the PTH curves were 1.12; 1.02 and 0.91 mM Ca for Mg concentrations of 0.5; 1 and 2, respectively. With the extreme Mg concentration of 5 mM, the secretory response was practically abolished at all Ca concentrations.
FIGURE 1:

Effect of calcium on PTH secretion at different magnesium concentrations in vitro. Intact, normal rat parathyroid glands were incubated at basal 1.25 mM Ca, and then, they were incubated for 1 h consecutive periods at Ca concentrations of 0.8, 1.0, 1.2 and 1.5 mM. The same experiment was repeated with each of the following Mg concentrations: 0.5, 1.0, 2.0 and 5.0 mM. PTH results are shown in pg/mL (A) and as % versus basal calcium (B) to standardize the change in PTH, avoiding differences in the total amount of parathyroid tissue. Values are expressed as means ± SEM (n = 9 in each group: three different experiments with three repetitions). (B) Data of PTH secretion at each different levels of calcium. Values are expressed means ± SEM (n = 9 in each group: three different experiments with three repetitions). *P < 0.05 versus Mg 0.5 mM in the same Ca group.
The addition of Mg to the medium reduced PTH secretion, but the degree of inhibition varied according to the Ca concentration. This is shown in Figure 1C, which depicts the data in a different way so that differences become more clearly visible. At the lowest Ca concentration (0.8 mM) with normal 0.5 mM Mg, maximal PTH secretion was observed; PTH increased to more than twice as with normal 1.25 mM Ca and 0.5 mM Mg. Mg concentrations of 1.0 and 2.0 mM had no significant effect on PTH secretion. Only an extremely high Mg concentration of 5.0 mM produced a significant 62% decrease in PTH secretion to values slightly lower than the PTH observed with normal Ca and Mg of 0.5 mM. At a Ca concentration of 1.0 mM with 0.5 mM Mg, PTH secretion was almost twice as high as with normal Ca. Here, the addition of both Mg 2.0 and 5.0 mM significantly decreased PTH secretion by 38 and 68%, respectively, to reach levels that were 120 and 60% of the PTH observed with normal Ca and Mg of 0.5 mM (Figure 1C); Mg 1.0 mM did not significantly reduce PTH secretion in this setting. At a normal 1.2 mM Ca concentration, Mg concentrations of 1.0 mM and 2.0 mM did not reduce PTH secretion significantly. Only an extremely high Mg concentration (5.0 mM) produced a significant 54% reduction in PTH secretion to levels that were 56% of that with normal Ca and Mg of 0.5 mM. Finally, when parathyroid glands were incubated with high Ca (1.5 mM), only the concentration of 5.0 mM Mg was able to decrease PTH secretion (by 37%) to levels that were 43% of normal Ca and 0.5 mM Mg, while no effect was detected with Mg 1.0 and 2.0 mM (Figure 1C).
The effect of magnesium on parathyroid CaR, VDR, FGFR1 and Klotho expression
From previous experiments, it became apparent that the inhibition of PTH by Mg was observed mainly when the serum Ca was 1.0 mM. The next set of experiments evaluating the effect of Mg on parathyroid CaR, VDR, FGFR1 and Klotho expression were performed using a Ca concentration of 1.0 mM and a Mg concentration of 0.5 or 2.0 mM.
The mRNA levels of CaR, VDR, FGFR1 and Klotho exhibited a 2.2, 1.6, 3.9 and 4.6-fold increase, respectively, at a Mg concentration of 2.0 mM when compared with 0.5 mM Mg (P < 0.05 in all the comparisons) (Figure 2). As the expression of the different mRNAs (CaR, VDR, FGFR1 and Klotho) changed in the same direction, we evaluated the expression of PTH mRNA to prove that the control gene performance was stable in this in vitro setting. PTH mRNA levels lowered to a 73 ± 5% at Mg 2.0 mM when compared with Mg 0.5 mM, P < 0.05 (Figure 2E).
FIGURE 2:

The effect of magnesium on the mRNA expression of parathyroid CaR, VDR, FGF23 and Klotho in vitro. Intact, normal rat parathyroid glands were incubated for 6 h with normal (0.5 mM) or high (2.0 mM) Mg concentration in the medium with Ca 1.0 mM. CaR, VDR and FGF23/Klotho mRNA levels were measured by quantitative real-time RT-PCR (versus β-actin mRNA). The expression of PTH mRNA (E) was also measured to prove that the control gene performance was stable in this in vitro setting. Values are expressed as means ± SEM (n = 9 in each group: three different experiments with three repetitions). *P < 0.05 versus Mg 0.5 mM.
The expression of these receptors at protein level was also evaluated by immunohistochemistry; protein content paralleled mRNA expression levels. Parathyroid CaR staining showed a 1.4-fold increase at 2.0 mM Mg when compared with 0.5 mM Mg (P < 0.05) (Figure 3A). Parathyroid VDR protein expression was also upregulated at 2.0 mM Mg by 1.2-fold with respect to the levels found at 0.5 mM Mg (P < 0.05) (Figure 3B). Finally, immunohistochemical analysis revealed that both FGFR1 and Klotho staining were 1.8- and 1.4-fold greater, respectively, at 2.0 mM than at 0.5 mM Mg (P < 0.05).
FIGURE 3:
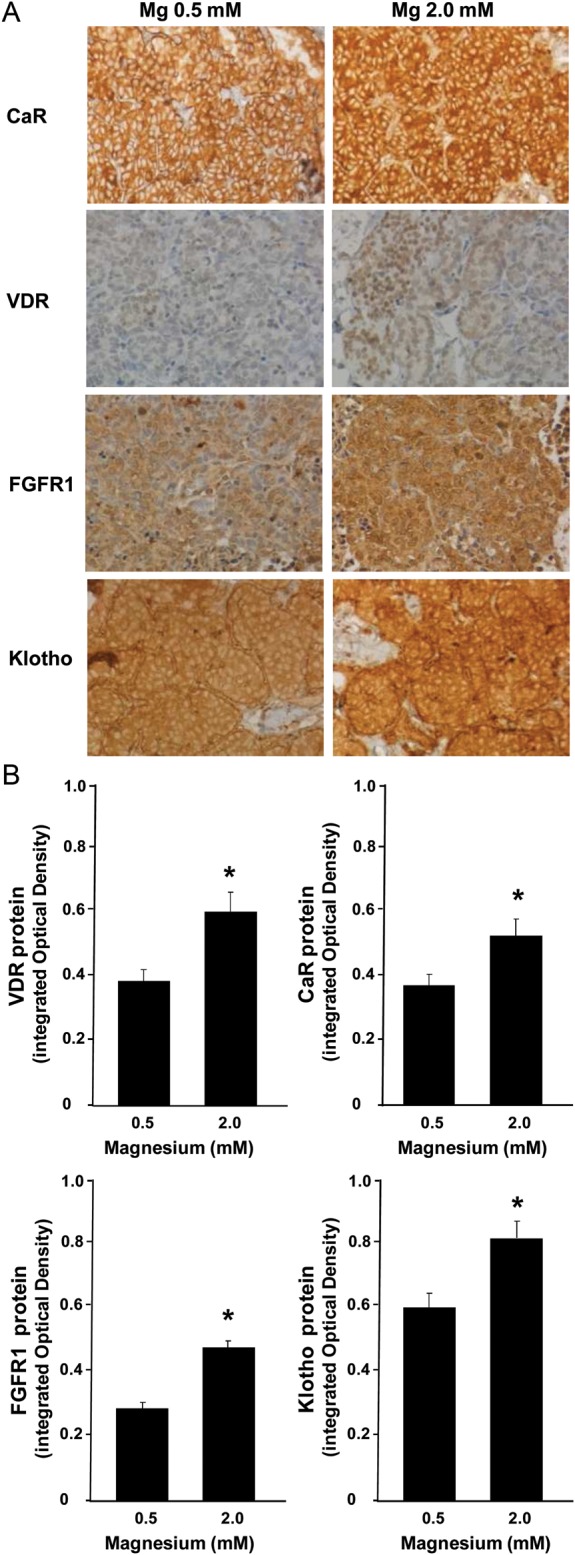
The effect of magnesium on the protein expression of parathyroid CaR, VDR, FGF23 and Klotho in vitro. Intact, normal rat parathyroid glands were incubated for 6 h with normal (0.5 mM) or high (2.0 mM) Mg concentration in the medium with low Ca 1.0 mM. Parathyroid glands were processed for immunohistochemical evaluation of CaR, VDR, FGF23 and Klotho protein. (A) Images are representative of four glands from three different experiments. Positive reaction (brown deposits) was revealed by the diaminobenzidine-tetrachloride system. Sections were counterstained with hematoxylin. Original magnification ×200. (B) Quantification of positive staining. It was performed in a minimum of five fields in more than three sections per specimen. *P < 0.05 versus Mg 0.5 mM.
DISCUSSION
The present study evaluates the combined effect of changes in Ca and Mg on parathyroid gland function. The work was performed by incubating whole parathyroid glands in Ca concentrations ranging from 0.8 to 1.5 mM; the experiments were repeated at different Mg concentrations. Our results show that Mg reduces PTH secretion mainly if serum Ca is somewhat below normal. In addition, Mg upregulates pivotal parathyroid CaR, VDR and FGFRI/Klotho at mRNA and protein levels.
In vivo animal and clinical studies [2, 14–17, 21–23] have shown that high Mg concentrations inhibit PTH secretion. Clinical studies on patients on haemodialysis and peritoneal dialysis showed that serum Mg was inversely and independently associated with PTH concentrations, even after adjusting for Ca and phosphorus levels [16, 17]. In an attempt to isolate the specific effect of Mg on PTH secretion, in vitro studies were also performed many years ago [10–13]. However, these were carried out in isolated parathyroid cells that rapidly lose CaR [24]. Thus, it appeared worthy to evaluate this effect in an in vitro model of intact parathyroid glands as this allows the elucidation of a direct effect of phosphorus on PTH secretion [19], which had been elusive in isolated parathyroid cells.
In the present study, we found that Mg reduced PTH secretion in an in vitro model of intact rat parathyroid glands. This is in accordance with previous studies in dispersed human and bovine parathyroid cells [10–13, 25]. However, we found that high Mg was able to reduce PTH only when parathyroid glands were exposed to moderately low Ca concentrations. The PTH–Ca curves obtained with progressively increasing Mg concentrations (from 0.5 to 5 mM) were consistently shifted to the left; thus, there was a decrease in the set point of Ca for PTH secretion. With normal to high Ca concentrations, only an extremely high Mg concentration of 5.0 mM was able to decrease PTH secretion; the effect of lower Mg concentrations was minor or absent. However, at 1 mM Ca (8 mg/dL total Ca), a serum concentration at the lower end in dialysis patients, the inhibitory effect of a Mg concentration of 2.0 mM was clearly evidenced. Interestingly, at 0.8 mM Ca the inhibitory effect of Mg was not observed anymore. It appears that in the setting of marked low calcium concentration, the parathyroid glands are strongly stimulated, so that only very high doses of Mg are able to decrease PTH secretion. In the general dialysis population, low Ca concentration is not the major concern; rather high Ca concentrations have been described to be associated with negative outcomes. But at high Ca concentrations, high Mg concentrations do not seem to pose a problem according to our experiments.
The relative potencies of Ca and Mg in inhibiting PTH secretion were previously addressed by Habener and Potts [10] and Brown's group [12, 13], who reported that Ca was about three times more potent than Mg in reducing PTH secretion in vitro. Our results allow the following conclusion: at a Mg concentration of 0.5 mM an increase in Ca from 1 to 1.5 mM (delta of 0.5 mM) reduces PTH by 57%; on the other hand, if Ca is maintained at 1 mM and the Mg concentration is increased by 0.5 mM (from 0.5 to 1 mM), the PTH secretion decreases by 14%. The ratio 57/14% is 4.0, indicating that Ca is four times more potent than Mg in suppressing PTH. However, the ability of Mg to decrease PTH is constant at all calcium concentrations. At 1 mM Ca, an increase in Mg concentration from 0.5 to 2.0 mM (delta of 1.5 mM) reduces PTH by 38%. An equal 38% decrease in PTH secretion is obtained with only a small increase in Ca from 1.0 to 1.2 mM (see Figure 1). Thus, the relative potency of Ca and Mg in reducing PTH secretion varies according to the basal serum Ca concentration and the degree of change in both Mg and Ca. Still, the potency of Ca in suppressing PTH is at least four times greater than that of Mg, which is close to what has been published before.
It is known that, like Ca, Mg is able to activate the CaR; though binding sites for Ca and Mg seem to be different. However, differences between Ca and Mg-elicited actions may be further related to mechanisms other than how Mg interacts with the CaR [2]. Mg and Ca may activate different intracellular signalling pathways. The paradoxical decrease in PTH secretion observed at any given Ca concentration under Mg deficiency in vivo, in vitro and in patients with severe hypomagnesaemia [26–28] was explained by a mechanism involving an intracellular increase in the activity of G alpha subunits of heterotrimeric G-proteins [29]. Besides, as deduced from a study by the group of Fitzpatrick et al. [30], addition of extracellular Mg to single isolated bovine parathyroid cells was able to affect PTH secretion through changes in intracellular Ca concentrations via two different CaR-independent mechanisms.
Ca also regulates parathyroid function by modulating the expression of key inhibitory receptors such as the CaR, VDR and FGFR/Klotho. In the present study we addressed whether Mg is also able to regulate the CaR expression. We did this by incubating parathyroid glands at a Ca concentration of 1.0 mM and Mg concentrations of 0.5 or 2.0 mM, which are inside the physiologically possible range. The expression of CaR, measured both at mRNA and protein levels, was upregulated by high Mg concentration. Thus, this distinct effect of Mg on the CaR expression represents an additional contribution of this cation to reduce PTH secretion. From previous own experiments [9, 31], we know that the minimal incubation period required for seeing a response in gene expression is 6 h. Therefore, the inhibitory effect of Mg on PTH secretion observed in our in vitro experiments cannot be attributed to changes in parathyroid CaR expression because the PTH secretion was evaluated after a 1 h of incubation, which is not enough time to allow for changes in CaR expression.
The effect of Mg on the expression of the parathyroid VDR, another key receptor in the regulation of PTH secretion, was also evaluated. The expression of VDR is upregulated by Ca [6] and calcimimetics [31]. Here, we show that Mg upregulates the VDR expression at mRNA and protein levels. This effect of Mg would favour the inhibition of PTH secretion by vitamin D.
Recent studies have shown that FGF23 regulates parathyroid function. FGF23 exerts an inhibitory effect on PTH secretion and parathyroid cell proliferation by binding to the receptor FGFR1 in the presence of its co-receptor Klotho [9, 32]. Klotho and FGFR are abundantly expressed in normal parathyroid cells, but in hyperplastic parathyroid glands the expression of FGFR1 and Klotho is markedly reduced, which likely makes the tissue resistant to the action of FGF23 [9]. Both FGFR1 and Klotho gene expression are upregulated by Ca [9]. Thus, we assessed whether Mg is also able to modulate the expression of these receptors. The results of this study show higher levels of both receptors in the parathyroid glands incubated with high (2.0 mM) Mg concentration when compared with those observed with normal (0.5 mM) Mg concentration. Therefore, the increased levels of expression of FGFR1 and Klotho may be considered as an additional mechanism underlying the downregulation of PTH secretion by Mg.
The capacity of Mg to modulate the expression of the CaR and, subsequently, of the VDR and the FGFR1/Klotho at long term suggests that this cation could be considered as a wide-range inhibitor of the whole parathyroid function. Furthermore, the inhibitory effects of Mg on parathyroid function, mainly when Ca is moderately low, could help to control secondary hyperparathyroidism. Interestingly, Mg also would favour the efficiency of therapeutic molecules targeting the parathyroid CaR, VDR or FGF23/Klotho. On the other hand, it is also important to emphasize that the decrease in PTH secretion did not result in an oversuppression of PTH at moderately elevated Mg concentrations. Our data certainly suggest that hypomagnesaemia should be avoided. In those cases of suppression of PTH by high Ca, the increased levels of Mg had limited additional suppressive effect on PTH. In the present study, performed with parathyroid glands from rats with normal renal function, we observed that Mg induced up-regulation of CaR, VDR and FGFR/Klotho. However, since the effect of Mg is through the activation of the CaR, and this receptor is usually down-regulated in CKD, results could be different in hyperplastic parathyroid glands from uraemic rats. Although this surely deserves to be evaluated in future studies, it could be assumed that higher levels of Mg should be needed to maintain comparable levels of CaR activity as well as the corresponding upregulation of the other receptors. Thus, avoiding hypomagnesaemia (and hypocalcaemia) in CKD prevents downregulation of key parathyroid receptors (CaR, VDR, Klotho FGFR1).
In conclusion, the results of this study show that parathyroid glands are sensitive to an inhibitory effect of Mg only when a moderate low calcium concentration is present. Mg also modulates the function of parathyroid glands through upregulation of the key cellular receptors CaR, VDR and the FGF23/Klotho system.
CONFLICT OF INTEREST STATEMENT
M.E.R.-O., A.C., C.H., J.M.-M.A.Z., A.P.-R., P.P.-M., J.F.N.-G., J.R.M.-C. and Y.A. have nothing to declare. M.R. has received research grants from Amgen and Fresenius and lecture fees from Amgen, Abbott, Shire and Fresenius. M.P., K.G. and S.S. are employees of Fresenius Medical Care Deutschland GmbH. JP-D is a consultant of Fresenius Medical Care Deutschland GmbH.
CONFLICT OF INTEREST STATEMENT
The results presented in this paper have not been published previously in whole or part, except in abstract format.
ACKNOWLEDGEMENTS
The study has been financed in part by Fresenius Medical Care Deutschland GmbH. Financial support was also obtained from Instituto de Salud Carlos III (FIS 10/10311, FIS 11/02055), Consejeria de Salud (JA 0127/2008), Consejeria de Innovacion, Ciencia y Empresa (CTS-5205) of Junta de Andalucia and SysKid European Proyect and Eutox. Y.A. is a senior researcher supported by the Nicolás Monardes Programme, Consejeria de Salud-SAS (Junta de Andalucia).
REFERENCES
- 1.Brown EM, Gamba G, Riccardi D, et al. Cloning and characterization of an extracellular Ca(2+)-sensing receptor from bovine parathyroid. Nature. 1993;366:575–580. doi: 10.1038/366575a0. [DOI] [PubMed] [Google Scholar]
- 2.Brown EM, MacLeod RJ. Extracellular calcium sensing and extracellular calcium signaling. Physiol Rev. 2001;81:239–297. doi: 10.1152/physrev.2001.81.1.239. [DOI] [PubMed] [Google Scholar]
- 3.Brown EM, Pollak M, Hebert SC. Sensing of extracellular Ca2+ by parathyroid and kidney cells: cloning and characterization of an extracellular Ca(2+)-sensing receptor. Am J Kidney Dis. 1995;25:506–513. doi: 10.1016/0272-6386(95)90118-3. [DOI] [PubMed] [Google Scholar]
- 4.Bourdeau A, Souberbielle JC, Bonnet P, et al. Phospholipase-A2action and arachidonic acid metabolism in calcium-mediated parathyroid hormone secretion. Endocrinology. 1992;130:1339–1344. doi: 10.1210/endo.130.3.1537295. [DOI] [PubMed] [Google Scholar]
- 5.Almadén Y, Canalejo A, Ballesteros E, et al. Regulation of arachidonic acid production by intracellular calcium in parathyroid cells: effect of extracellular phosphate. J Am Soc Nephrol. 2002;13:693–698. doi: 10.1681/ASN.V133693. [DOI] [PubMed] [Google Scholar]
- 6.Garfia B, Cañadillas S, Canalejo A, et al. Regulation of parathyroid vitamin D receptor expression by extracellular calcium. J Am Soc Nephrol. 2002;13:2945–2952. doi: 10.1097/01.asn.0000037676.54018.cb. [DOI] [PubMed] [Google Scholar]
- 7.Mendoza FJ, Lopez I, Canalejo R, et al. Direct upregulation of parathyroid calcium-sensing receptor and vitamin D receptor by calcimimetics in uremic rats. Am J Physiol Renal Physiol. 2009;296:F605–F613. doi: 10.1152/ajprenal.90272.2008. [DOI] [PubMed] [Google Scholar]
- 8.Cañadillas S, Canalejo R, Rodriguez-Ortiz ME, et al. Upregulation of parathyroid VDR expression by extracellular calcium is mediated by ERK1/2-MAPK signaling pathway. Am J Physiol Renal Physiol. 2010;298:F1197–F1204. doi: 10.1152/ajprenal.00529.2009. [DOI] [PubMed] [Google Scholar]
- 9.Canalejo R, Canalejo A, Martinez-Moreno JM, et al. FGF23 fails to inhibit uremic parathyroid glands. J Am Soc Nephrol. 2010;21:1125–1135. doi: 10.1681/ASN.2009040427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Habener JF, Potts JT., Jr Relative effectiveness of calcium and magnesium in the secretion and biosynthesis of parathyroid hormone in vitro. Endocrinology. 1976;98:197–202. doi: 10.1210/endo-98-1-197. [DOI] [PubMed] [Google Scholar]
- 11.Brown EM. Relationship of 3, 5′-adenosine monophosphate accumulation to parathyroid hormone release in dispersed cells from pathological human parathyroid tissue. J Clin Endocrinol Metab. 1981;52:961–968. doi: 10.1210/jcem-52-5-961. [DOI] [PubMed] [Google Scholar]
- 12.Brown EM, Thatcher JG, Watson EJ, et al. Extracellular calcium potentiates the inhibitory effects of magnesium on parathyroid function in dispersed bovine parathyroid cells. Metabolism. 1984;33:171–176. doi: 10.1016/0026-0495(84)90131-8. [DOI] [PubMed] [Google Scholar]
- 13.Shoback DM, Thatcher JG, Brown EM. Interaction of extracellularcalcium and magnesium in the regulation of cytosolic calcium and PTH release in dispersed bovine parathyroid cells. Mol Cell Endocrinol. 1984;38:179–186. doi: 10.1016/0303-7207(84)90116-3. [DOI] [PubMed] [Google Scholar]
- 14.McGonigle RJ, Weston MJ, Keenan J, et al. Effect of hypermagnesemia on circulating plasma parathyroid hormone in patients on regular hemodialysis therapy. Magnesium. 1984;3:1–7. [PubMed] [Google Scholar]
- 15.Takahashi S, Okada K, Yanai M. Magnesium and parathyroid hormone changes to magnesium-free dialysate in continuous ambulatory peritoneal dialysis patients. Perit Dial Int. 1994;14:75–78. [PubMed] [Google Scholar]
- 16.Navarro JF, Mora C, Jiménez A, et al. Relationship between serum magnesium and parathyroid hormone levels in hemodialysis patients. Am J Kidney Dis. 1999;34:43–48. doi: 10.1016/s0272-6386(99)70106-x. [DOI] [PubMed] [Google Scholar]
- 17.Navarro JF, Mora C, Macia M, et al. Serum magnesium concentration is an independent predictor of parathyroid hormone levels in peritoneal dialysis patients. Perit Dial Int. 1999;19:455–461. [PubMed] [Google Scholar]
- 18.de Francisco AL, Leidig M, Covic AC, et al. Evaluation of calcium acetate/magnesium carbonate as a phosphate binder compared with sevelamer hydrochloride in haemodialysis patients: a controlled randomized study (CALMAG study) assessing efficacy and tolerability. Nephrol Dial Transplant. 2010;25:3707–3717. doi: 10.1093/ndt/gfq292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Almadén Y, Canalejo A, Ballesteros E, et al. Effect of high extracellular phosphate concentration on arachidonic acid production by parathyroid tissue in vitro. J Am Soc Nephrol. 2000;11:1712–1718. doi: 10.1681/ASN.V1191712. [DOI] [PubMed] [Google Scholar]
- 20.Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-cloroform extraction. Anal Biochem. 1987;162:156–159. doi: 10.1006/abio.1987.9999. [DOI] [PubMed] [Google Scholar]
- 21.Massry SG, Coburn JW, Kleeman CR. Evidence for suppression of parathyroid gland activity by hypermagnesemia. J Clin Invest. 1970;49:1619–1629. doi: 10.1172/JCI106379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cholst IN, Steinberg SF, Tropper PJ, et al. The influence of hypermagnesemia on serum calcium and parathyroid hormone levels in human subjects. N Engl J Med. 1984;310:1221–1225. doi: 10.1056/NEJM198405103101904. [DOI] [PubMed] [Google Scholar]
- 23.Ferment O, Garnier PE, Touitou Y. Comparison of the feedback effect of magnesium and calcium on parathyroid hormone secretion in man. J Endocrinol. 1987;113:117–122. doi: 10.1677/joe.0.1130117. [DOI] [PubMed] [Google Scholar]
- 24.Brown AJ, Zhong M, Ritter C, et al. Loss of calcium responsiveness in cultured bovine parathyroid cells is associated with decreased calcium receptor expression. Biochem Biophys Res Commun. 1995;212:861–867. doi: 10.1006/bbrc.1995.2048. [DOI] [PubMed] [Google Scholar]
- 25.Chattopadhyay N, Mithal A, Brown EM. The calcium-sensing receptor: a window into the physiology and pathophysiology of mineral ion metabolism. Endocr Rev. 1996;17:289–307. doi: 10.1210/edrv-17-4-289. Review. Erratum in: Endocr Rev 1996; 17: 517. [DOI] [PubMed] [Google Scholar]
- 26.Inaba M, Okuno S, Imanishi Y, et al. Magnesium deficiency enhances secretion of parathyroid hormone in normal and 5/6-nephrectomized uremic rats. J Endocrinol Invest. 1992;15:135–142. [PubMed] [Google Scholar]
- 27.Mahaffee DD, Cooper CW, Ramp WK, et al. Magnesium promotes both parathyroid hormone secretion and adenosine 3′,5′-monophosphate production in rat parathyroid tissues and reverses the inhibitory effects of calcium on adenylate cyclase. Endocrinology. 1982;110:487–495. doi: 10.1210/endo-110-2-487. [DOI] [PubMed] [Google Scholar]
- 28.Duran MJ, Borst GC, III, Osburne RC, et al. Concurrent renal hypomagnesemia and hypoparathyroidism with normal parathormone responsiveness. Am J Med. 1984;76:151–154. doi: 10.1016/0002-9343(84)90764-2. [DOI] [PubMed] [Google Scholar]
- 29.Quitterer U, Hoffmann M, Freichel M, et al. Paradoxical block of parathormone secretion is mediated by increased activity of G alpha subunits. J Biol Chem. 2001;276:6763–6769. doi: 10.1074/jbc.M007727200. [DOI] [PubMed] [Google Scholar]
- 30.Miki H, Maercklein PB, Fitzpatrick LA. Effect of magnesium on parathyroid cells: evidence for two sensing receptors or two intracellular pathways? Am J Physiol. 1997;272:E1–E6. doi: 10.1152/ajpendo.1997.272.1.E1. [DOI] [PubMed] [Google Scholar]
- 31.Rodriguez ME, Almaden Y, Cañadillas S, et al. The calcimimetic R-568 increases vitamin D receptor expression in rat parathyroid glands. Am J Physiol Renal Physiol. 2007;292:F1390–F1395. doi: 10.1152/ajprenal.00262.2006. [DOI] [PubMed] [Google Scholar]
- 32.Ben-Dov IZ, Galitzer H, Lavi-Moshayoff V, et al. The parathyroid is a target organ for FGF23 in rats. J Clin Invest. 2007;117:4003–4008. doi: 10.1172/JCI32409. [DOI] [PMC free article] [PubMed] [Google Scholar]