

Abstract
We evaluated the adjuvant effect of a modified glycoform of lipopolysaccharide (LPS) (LgtB-LpxL1) compared to that of the nonmodified glycoform Lpxl1 serogroup B meningococcal H44/76 native outer membrane vesicles (nOMVs) on immune responses to vaccination with the recombinant meningococcal protein, rPorA, tetanus toxoid, or meningococcal serogroup C capsular polysaccharide. We used LgtB-LpxL1 LPS because the disruption of the lgtB gene, which results in the exposure of N-acetylglucosamine-galactose-glucose residues in the LPS outer core, has been shown to enhance the activation of human dendritic cells in vitro. The responses were compared to those of a monophosphoryl lipid A (MPL)-based adjuvant and to an aluminum hydroxide suspension. The nOMVs induced blood serum IgG responses against each of the three antigens comparable to those obtained with MPL or aluminum salt. However, nOMVs elicited (i) a lower IgG1/IgG2a ratio against rPorA and (ii) serum bactericidal antibody titers superior to those achieved with aluminum salt, reaching similar titers to those obtained with MPL. Similarly, bactericidal antibody titers induced by immunization with meningococcal serogroup C polysaccharide and nOMVs were similar to those obtained using MPL but were better than those with aluminum salt. Immunization with tetanus toxoid and nOMVs resulted in tetanus toxoid-specific IgG responses similar to those obtained when adjuvanted with aluminum salt. These results highlight the potential utility of meningococcal LpxL1 LPS-containing nOMVs as an adjuvant for recombinant meningococcal protein vaccines and suggest their possible use with a variety of other antigens.
INTRODUCTION
Serogroup B Neisseria meningitidis (MenB) is the predominant cause of meningococcal disease in developed countries (1, 2). Unlike with serogroups A, C, Y, and W-135, the prevention of serogroup B is difficult to achieve using capsular polysaccharide vaccines, as the MenB polysaccharide closely resembles cell surface exposed structures, including those expressed on human neural cell adhesion molecules (3). It has been speculated that this leads to immunological tolerance and possibly to autoimmune reactivity induced by anti-MenB polysaccharide antibodies (3). An alternative approach to vaccine development has been the use of subcapsular antigens in the form of recombinant proteins or outer membrane vesicles (OMVs) (4, 5). Meningococcal OMVs consist of a phospholipid bilayer containing mainly outer membrane proteins, lipoproteins, and lipopolysaccharide (LPS), some of which can induce protective immune responses. OMV-based vaccines have been used on a large scale for the control of clonal outbreaks of meningococcal disease (5–7). Additionally, a recombinant protein-based vaccine containing OMV is currently in advanced clinical trials and was recently licensed in Europe (8). OMV-based vaccines are immunogenic; however, protection is restricted to the variants of the antigens in the vaccine, with the surface protein PorA being immunodominant and a major component of the OMVs. Therefore, the breadth of protection afforded by OMV vaccines against MenB disease largely depends on the variability of PorA, and to a certain extent, other outer membrane antigens expressed on the surfaces of the circulating target strains (9). Adjuvants can broaden the coverage of these vaccines (10), and the utilization of better adjuvants may ultimately be key to the successful development of broadly protective vaccines against MenB.
The adjuvants currently licensed for human use include aluminum salts, monophosphoryl lipid A (MPL), oil-in-water emulsions, and liposomes (11, 12). So far, aluminum salts have been employed in most meningococcal OMV and protein vaccines that have been developed. However, aluminum salts are poor adjuvants in many situations, especially when a cellular immune response is required, as they mainly induce a Th2-biased response (13). Interestingly, aluminum salts in OMV vaccines may contribute to reducing LPS-associated toxicity (14, 15). LPS has been suggested as an alternative adjuvant for meningococcal vaccines and can also act as a potential antigen (16–19). LPS is a strong adjuvant (20) and has been shown to skew T-cell responses toward a Th1-type immunity, which may be important for protection against meningococcal disease (21). The detergent extraction procedure used to produce OMV vaccines lowers LPS content and reduces reactogenicity but also minimizes the adjuvant effects of LPS on vaccine immunogenicity. The toxic and adjuvant effects of LPS are mainly mediated by its lipid A portion. A mutation in the lpxL1 gene results in penta-acylated LPS (LpxL1 LPS), which is less toxic but retains the immunostimulatory property of wild-type LPS (22), and thus allows the use of LPS as a potentially safe and effective adjuvant.
Upon recognition of the lipid A portion by the LPS-binding protein, LPS is transferred to CD14, which in turn delivers it to a Toll-like receptor 4 (TLR4)-MD2 complex present on the surface of antigen-presenting and particularly dendritic cells (DCs). This results in DC maturation and the activation of the adaptor proteins MyD88 and TIR-domain-containing adapter-inducing β interferon (TRIF), resulting in the release of proinflammatory cytokines. Recently, the core oligosaccharide portion of LPS was shown to mediate interaction with DCs independently of TLR4 (23). Since DCs play a central role in the initiation of immune responses, an alteration in the sugar composition in the outer core of LPS may enhance its adjuvant effect. In particular, the disruption of lgtB, preventing the addition of galactose and further extensions, allows a strong interaction with human DC-specific intercellular adhesion molecule 3-grabbing nonintegrin (DC-SIGN), leading to better antigen capture, uptake, and processing by DCs (24).
We hypothesized that the disruption of lgtB results in an LPS molecule with a stronger adjuvant effect than an lpxl1 mutation only. The lpxl1 mutation enabled us to utilize native OMVs (nOMVs) that were produced without the use of detergent, thereby maintaining a high level of LPS in its natural membrane-bound conformation. In this study, we evaluated the adjuvant effects of nOMVs containing LpxL1 LPS and LgtB-LpxL1 LPS derived from the MenB strain H44/76 using a recombinant meningococcal protein antigen, rPorA P1.7-2,4. We also evaluated their adjuvant effects on a nonmeningococcal protein antigen (tetanus toxoid) and a meningococcal nonprotein antigen (MenC polysaccharide).
MATERIALS AND METHODS
Bacterial strains and growth conditions.
The strains used for nOMV production were derived from N. meningitidis strains H44/76 (B:15:P1.7,16, immunotype L3,7,9) and MC58 (B:15:P17, 16b, immunotype L3) containing a disrupted lgtB gene (MC58-lgtB) (25). Serum bactericidal antibody (SBA) assays were performed using N. meningitidis strains BZ198 (B:NT:P1.7-2,4) and C11 (C:16:P1.7,1). The bacteria were grown on brain heart infusion (BHI) agar (Merck, Darmstadt, Germany) supplemented with Levinthal's base (10% vol/vol) in a humidified atmosphere containing 5% CO2 at 37°C. Where required, the medium was supplemented with kanamycin (100 μg/ml) or tetracycline (2 μg/ml) (Sigma-Aldrich, Gillingham, United Kingdom).
Construction of isogenic mutant strains expressing different LPS glycoforms.
H44/76-lgtB was constructed by transforming H44/76 with chromosomal DNA from MC58-lgtB using the spot transformation technique as previously described (26–28). The transformants were identified on media as described above, supplemented with the appropriate antibiotics, and characterized using PCR and tricine-SDS-PAGE (TSDS-PAGE). To disrupt lpxL1 in both the wild-type H44/76 and H44/76-lgtB strains, the 5′ and 3′ ends of the lpxL1 gene were first amplified from H44/76 chromosomal DNA by PCR using two primer pairs: lpxL1-for (5′-ATCCTTCGGGGATGCAGG-3′) and lpxL1-rev (5′-TAATCGTTGCCGTCTGAATACCG-3′), and lpxL1-XmaI-for (5′-TTGGTACCCGGGTGCCGGAC-3′) and lpxL1-XmaI-rev (5′-GTCCGGCGGGCCCGTACCAA-3′). The products were cloned separately into the TA vector pCR2.1-TOPO (Invitrogen, Paisley, United Kingdom) before being ligated together to create a novel XmaI restriction site within the gene. A tetracycline resistance cassette excised from pUC4NmDUS (29) using XmaI was used to disrupt the lpxL1gene, which was similarly prepared. The resulting plasmid was used to transform N. meningitidis strains H44/76 and H44/76-lgtB to produce H44/76-lpxL1 and H44/76-lgtB-lpxL1.
nOMV production.
The nOMVs were produced as previously described (30) but with some modifications. Briefly, bacteria were grown to confluence and resuspended into 20 ml of 0.05 M Tris-HCl, 0.15 M NaCl, 0.01 M EDTA, and 0.01% thimerosal (pH 7.4). The suspension was incubated at 56°C for 1 h and sonicated with 10 15-s bursts. Following two rounds of centrifugation of the sonicated cell mass at 16,000 × g for 15 min at 4°C, OMVs were harvested from the supernatant and purified twice by ultracentrifugation at 100,000 × g for 2 h at 4°C. The final OMV pellet was resuspended in 3% sucrose containing 0.01% thimerosal and stored at 4°C. The protein contents of the OMVs were quantified using a Micro-Lowry assay (Sigma-Aldrich), and the protein and LPS profiles were assessed by SDS-PAGE (31) and TSDS-PAGE (32), respectively.
Vaccines.
Recombinant PorA P1.7-2,4 was prepared following expression of porA cloned into apET30-EkLIC plasmid in Escherichia coli strain T7 Express (New England BioLabs, Hitchin, United Kingdom). The cells were harvested, disrupted by sonication, and the inclusion bodies isolated as previously described (33) and resuspended in a buffer of Tris (10 mM [pH 7.5]), EDTA (1 mM), and urea (8 M). Following centrifugation at 14,000 × g for 20 min to remove the cell debris, the supernatant was added while stirring rapidly to Tris (20 mM [pH 7.9]), NaCl (1 M), and 2% (wt/vol) Zwittergent 3-14 (ZW 3-14) in a 1:1 ratio before being dialyzed against Tris (20 mM [pH 7.9]), NaCl (0.5 M), and 0.05% (wt/vol) ZW 3-14 for two changes of 6 to 8 h at 4°C. After dialysis, the rPorA was filtered through a 0.45-μm syringe filter and then applied to a HisTrap HP column (GE Healthcare) in Tris (20 mM [pH 7.9]), NaCl (0.5 M), 0.1% (vol/vol) lauryldimethylamine oxide, and imidazole (10 mM). The column was washed with the same buffer containing 40 mM, and then 500 mM imidazole was utilized to elute the rPorA, which was then dialyzed against Tris (10 mM [pH 7.9]), NaCl (150 mM), and 0.05% (wt/vol) ZW 3-14. A circular dichroism spectrum was run using a Jasco J-10 spectrometer and a 0.05-cm path length quartz cell and showed near-complete folding. MenC polysaccharide (MenC-PS) and tetanus toxoid (TT) were obtained from the National Institute for Biological Standards and Control (NIBSC) (Potters Bar, United Kingdom). The aluminum hydroxide wet gel suspension was obtained from Brenntag Biosector, Denmark, and the MPL-based Sigma adjuvant system was obtained from Sigma-Aldrich. This adjuvant, referred to as MPL, is a stable oil-in-water emulsion (2% squalene oil-in-water), which can be used as an alternative to the classical Freund's water-in-oil emulsion. Each vial contains 0.5 mg monophosphoryl lipid A (isolated from Salmonella minnesota) and 0.5 mg synthetic trehalose dicorynomycolate in 44 μl of squalene oil, 0.2% Tween 80, and water. The Sigma adjuvant system has been tested for adjuvanticity and safety by Sigma and is approved for use in mice.
Murine immunization.
The animal studies were conducted according to the United Kingdom Home Office regulations and were approved by the local ethics committee at the NIBSC (Home Office Project license no. 80/2157). The samples were obtained following the use of terminal general anesthesia, and all efforts were made to minimize suffering. Groups of 10 6- to 8-week-old National Institutes of Health (NIH) inbred mice (NIH/OlaHSD) obtained from Harlan, United Kingdom, were injected subcutaneously on days 0 and 28 with rPorA (5 μg), MenC-PS (10 μg), or TT (1 flocculation unit [Lf], approximately 5 μg/ml), in combination with either LpxL1 nOMVs (2.5 μg), LgtB-LpxL1 nOMVs (2.5 μg), MPL (41.7 μg), or aluminum hydroxide (330 μg), as described in Table 1. Blood samples were obtained by cardiac puncture on day 42, and the blood serum was separated and stored at −80°C.
TABLE 1.
Murine immunization groups
| Group no. | Antigen amount and type | Adjuvant amount and type |
|---|---|---|
| 1 | 5.0 μg rPorA P1.7-2,4 | 2.5 μg H44/76-lpxL1 nOMVs |
| 2 | 5.0 μg rPorA P1.7-2,4 | 2.5 μg H44/76-lgtB-lpxL1 nOMVs |
| 3 | 5.0 μg rPorA P1.7-2,4 | 41.7 μg MPL |
| 4 | 5.0 μg rPorA P1.7-2,4 | 330 μg aluminum hydroxide |
| 5 | 10 μg MenC polysaccharide | 2.5 μg H44/76-lpxL1 nOMVs |
| 6 | 10 μg MenC polysaccharide | 2.5 μg H44/76-lgtB-lpxL1 nOMVs |
| 7 | 10 μg MenC polysaccharide | 41.7 μg MPL |
| 8 | 10 μg MenC polysaccharide | 330 μg aluminum hydroxide |
| 9 | 1 Lf tetanus toxoid | 2.5 μg H44/76-lpxL1 nOMVs |
| 10 | 1 Lf tetanus toxoid | 2.5 μg H44/76-lgtB-lpxL1 nOMVs |
| 11 | 1 Lf tetanus toxoid | 41.7 μg MPL |
| 12 | 1 Lf tetanus toxoid | 330 μg aluminum hydroxide |
Analysis of antibody responses by ELISA.
Recombinant PorA P1.7-2,4 (0.4 μg/ml) in a buffer containing 15 mM sodium carbonate and 35 mM sodium bicarbonate (pH 9.6) was used to coat 96-well microtiter plates overnight at 4°C. After washing with Tris-buffered saline (5 mM Tris, 15 mM NaCl [pH 7.6]), the plates were blocked with 0.5% bovine serum albumin (BSA) (Sigma-Aldrich) at 37°C for 1 h. The serum samples were diluted from 1:2,000 to 1:256,000 in 0.5% BSA before being added to the wells (100 μl/well) and were incubated at 4°C for 16 to 18 h. The plates were washed before the addition of either anti-mouse IgG (1:5,000) (Sigma), anti-mouse IgG1 (1:2,500) (Southern Biotechnology), or anti-mouse IgG2a (1:2,500) (Southern Biotechnology), conjugated to alkaline phosphatase and diluted in 0.5% BSA. The plates were incubated at 37°C for 2 h, washed, and 100 μl of p-nitrophenyl phosphate (pNPP) (Sigma) diluted in pNPP substrate buffer (Zymed) was added to each well. Following incubation in the dark at room temperature for 50 min, the reaction was stopped by adding 50 μl of 2-M NaOH per well, and the absorbance at 405 nm was measured (with a 620-nm reference) using a Multiskan EX microplate reader (Thermo Scientific).
An anti-MenC-PS enzyme-linked immunosorbent assay (ELISA) was performed as described above using Immulon 2HB plates (Fisher Scientific) coated with a mixture of MenC-PS and methylated human serum albumin (NIBSC) diluted to a final concentration of 5 μg/ml each in phosphate-buffered saline (PBS). The mouse serum samples were diluted 1:50 in PBS containing 5% newborn bovine serum (Sigma).
Anti-TT ELISA was performed as previously described (34), with slight modifications. The plates (Immulon 2HB) were coated with TT (NIBSC) diluted to 2 Lf/ml in sterile water. Blocking was performed using skimmed milk (Fluka) diluted in 1% (wt/vol) PBS. The mouse serum samples were diluted to 1:1,000 in PBS containing 0.5% BSA. The pNPP substrate was incubated for 50 min (IgG1 and IgG2a) or 60 min (IgG).
An in-house standard serum pool was prepared by pooling the serum samples of randomly selected mice. Variable starting dilutions between 1:50 and 1:2,000 (depending on the assay) were used to generate standard curves against which the antigen-specific IgG concentration of each serum sample was quantified. Each standard curve was fitted using a four-parametric sigmoid logistic model, as previously described (35).
To enable calculations of IgG1/IgG2a ratios, individual IgG1 and IgG2a concentrations were expressed as the log10 of the titers, defined as the reciprocal serum dilution that gave 50% of the maximum absorbance.
Serum bactericidal antibody assay.
The SBA assay was performed as previously described (36). Briefly, a serum pool was made for each immunization group, using an equal volume of serum from each mouse, which was then incubated at 56°C for 30 min to inactivate endogenous complement. Pooled serum samples diluted 1:4 in Hanks' balanced salt solution (HBSS) with 0.5% glucose were serially diluted 2-fold in 96-well plates (Greiner). The target strains were grown overnight on agar plates and subplated for 4 h before suspensions of these strains were diluted and added at 1,000 CFU/well, and they were incubated with the mouse serum for 10 min at 37°C at 5% CO2. Baby rabbit complement at 25% of the total volume was added, and the microtiter plate was incubated at 37°C at 5% CO2 for 1 h. Part of the reaction mixture was spread onto BHI agar plates, incubated overnight, and counted. The SBA titer was defined as the reciprocal of the highest serum dilution that resulted in ≥50% bacterial killing.
A competitive inhibition SBA assay was performed as previously described (37). Briefly, rPorA P1.7-2,4 or P1.7,16 was added to a final concentration of 500 μg/ml to the decomplemented pooled serum of mice. The mixture was incubated for 16 to 18 h at 4°C before being tested in the SBA assay against strain BZ198, as described above.
Statistical analysis.
ELISA results were analyzed using GraphPad Prism 5 (GraphPad Software, Inc.). If the data were shown to follow a normal distribution as assessed by the Shapiro-Wilk test, a one-way analysis of variance (ANOVA) was performed to look for an overall difference between the groups. This was followed by a post hoc Bonferroni's multiple comparison test. If the data did not follow a normal distribution, a nonparametric Kruskal-Wallis test, followed by the Mann-Whitney U test, was performed.
RESULTS
Protein and LPS composition of H44/76-LpxL1 and H44/76-lgtB-LpxL1nOMVs.
Characterization of the protein and LPS contents of LpxL1 and LgtB-LpxL1 nOMVs by SDS-PAGE and TSDS-PAGE showed that the protein profiles were comparable, with the exception of lpxL1 nOMVs having an additional band consistent with a class 5 protein, i.e., Opa or OpcA (Fig. 1A). LpxL1 nOMVs also possessed two distinct LPS bands (Fig. 1B), while a single band was observed in the LgtB-LpxL1 nOMVs (Fig. 1B). The positions of the LgtB-LpxL1 band confirmed the successful truncation of the LPS molecule. The upper band of LpxL1 nOMVs was at a similar level to the full-length LpxL1 cell lysate control; therefore, the lower band in the truncated lpxL1-LPS implies the presence of an additional truncated form of LPS in the LpxL1 nOMVs.
FIG 1.
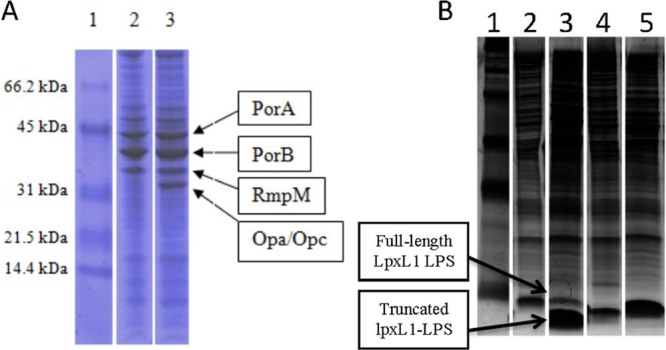
(A) SDS-PAGE of LpxL1 and LgtB-LpxL1 nOMVs. Lane 1, low-molecular-weight protein standard; lane 2, LgtB-LpxL1 nOMVs; lane 3, LpxL1 nOMVs. The identity of the protein represented in each band is also shown. (B) Silver-stained TSDS-PAGE of LpxL1 and LgtB-lpxL1 nOMVs and cell lysates. Lane 1, low-molecular-weight protein standard; lane 2, H44/76-LpxL1 cell lysates; lane 3, H44/76-LpxL1 nOMVs; lane 4, H44/76-LgtB-LpxL1 cell lysates; lane 5, H44/76-LgtB-LpxL1 nOMVs.
The adjuvant effect of nOMVs on IgG responses is antigen dependent.
The adjuvant effect of nOMVs, MPL, and aluminum hydroxide differed depending on the immunizing antigen. Groups of mice were immunized twice with one of the 3 antigens at 4-week intervals, and immunogenicity was analyzed using blood serum samples collected 2 weeks after the last injection. Following 2 immunizations with rPorA, MPL induced a higher IgG response in mice than either nOMV (P < 0.0001) or aluminum hydroxide (P < 0.05), as tested 2 weeks after the last injection (Fig. 2A). In addition, aluminum hydroxide stimulated greater IgG production than LgtB-LpxL1 nOMV (P < 0.05). A similar pattern was observed following 2 immunizations with TT (Fig. 2B), as MPL elicited a significantly higher TT-specific IgG response than the three other adjuvants (P < 0.0001 for LpxL1 nOMVs, P < 0.001 for LgtB-LpxL1 nOMVs, and P < 0.01 for aluminum hydroxide) (Fig. 2B). In this case, however, there was no significant difference between aluminum hydroxide and the nOMVs. When MenC-PS was used as the antigen (Fig. 2C), all four adjuvants had a similar effect on the production of total IgG (Fig. 2C). In all cases, there was no significant difference between the two nOMVs.
FIG 2.
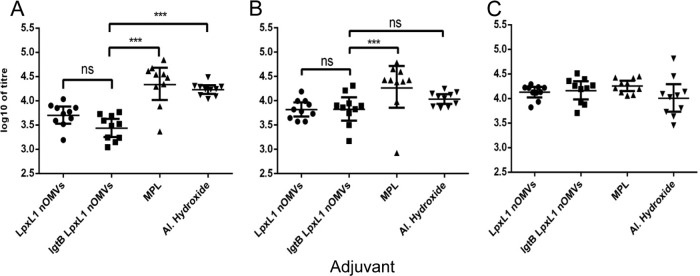
Antigen-specific IgG levels in the blood serum samples of mice immunized with rPorA (A), tetanus toxoid (B), or MenC polysaccharide (C) in combination with lpxL1 nOMVs, lgtB-lpxL1 nOMVs, MPL, or aluminum hydroxide. The antigens used to coat the ELISA plates were recombinant rPorA P1.7-2,4 (A), unadsorbed tetanus toxoid (B), and MenC polysaccharide (C). The mice were immunized twice at 4-week intervals, and the serum samples were collected 2 weeks after the last injection. Each graph shows individual IgG levels, the geometric mean, and 95% confidence interval. IgG concentrations are expressed as log10 of titer, defined as the reciprocal serum dilution giving 50% of maximum absorbance. ns, nonsignificant; ***, statistically significant at P < 0.0001.
nOMVs bias PorA-specific IgG subclass production toward a Th1-type response.
We analyzed the presence of the IgG1 and IgG2a antibody subclasses against rPorA and TT in order to assess the effects of the different adjuvants on T helper cell differentiation. In mice, IgG1 is produced predominantly by a Th2-associated response, whereas IgG2a is produced by a Th1-associated response (38, 39). Hence, a low IgG1/IgG2a ratio is indicative of a Th1-predominant response in mice. Immunizations with all adjuvants resulted in similar levels of IgG2a against rPorA, with the exception of MPL, which elicited significantly more of this antibody type than the LgtB-LpxL1 nOMV (Fig. 3A). However, aluminum hydroxide mediated the induction of significantly higher levels of IgG1 than either nOMV (P < 0.0001), and there was no significant difference between the two nOMVs (Fig. 3B). Consequently, both nOMVs resulted in significantly lower IgG1/IgG2a ratios than aluminum hydroxide (P < 0.0001). MPL also induced a lower IgG1/IgG2a ratio than aluminum hydroxide (P = 0.0288) (Fig. 3C). With the TT antigen, although MPL elicited a stronger IgG1 response than LpxL1 nOMVs (P = 0.0029), LgtB-LpxL1 nOMVs (P = 0.0021), and aluminum hydroxide (P = 0.0021), there was no significant difference between the four adjuvants in terms of IgG2a levels or the IgG1/IgG2a ratios (Fig. 4C).
FIG 3.
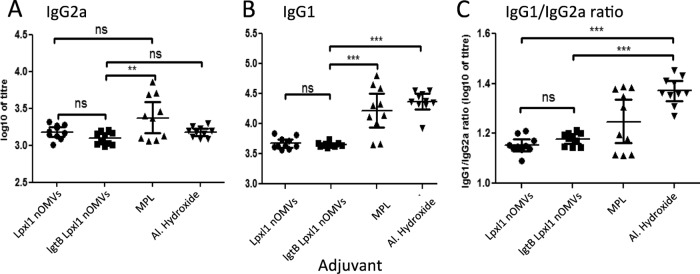
Recombinant rPorA P1.7-2,4-specific antibody subclasses. The serum samples of recombinant rPorA P1.7-2,4-immunized mice (ten per group) were analyzed by ELISA for the presence of recombinant rPorA P1.7-2,4-specific IgG2a (A) or IgG1 (B). (C) The IgG1/IgG2a ratio was then calculated. Each graph shows the antibody level or IgG1/IgG2a ratio for each serum sample, the geometric mean of each group, and the 95% confidence interval. The IgG1 and IgG2a concentrations are expressed as log10 of titer, defined as the reciprocal serum dilution giving 50% of maximum absorbance. ns, nonsignificant; **, statistical significance with P < 0.001; ***, statistically significant at P < 0.0001.
FIG 4.

Tetanus toxoid-specific antibody subclasses. The serum samples of tetanus toxoid-immunized mice (ten per group) were analyzed by ELISA for the presence of tetanus toxoid-specific IgG2a (B) and IgG1 (A). (C) The IgG1/IgG2a ratio was then calculated. Each graph shows the antibody level or IgG1/IgG2a ratio for each serum, the geometric mean of each group, and the 95% confidence interval. The IgG1 and IgG2a concentrations are expressed as log10 of titer, defined as the reciprocal serum dilution giving 50% of maximum absorbance. Since the calculation of antibody titers became inaccurate when the highest optical density at 405 nm (OD405) was <0.5, some sera were repeated at lower dilutions. If the OD405 was still <0.5 even at a 1:5 dilution, the sera were assigned an arbitrary titer of 1. This applied to a total of 9 sera in the IgG2a ELISA (2 from the TT + LpxL1 group, 3 from the TT + LgtB-LpxL1 group, 1 from the TT + MPL group, and 3 from the TT + aluminum hydroxide group). ns, nonsignificant; **, statistically significant at P < 0.001.
nOMVs promote strong anti-meningococcal serum bactericidal activity following immunization with rPorA.
In mice immunized with rPorA combined with an nOMV variant, both nOMV variants helped induce SBA titers comparable to those with the MPL adjuvant. Immunization using either MPL or nOMVs resulted in positive titers between 1:32 and 1:128 (Table 2). By comparison, the combination of rPorA with aluminum hydroxide was unable to mediate the induction of bactericidal antibodies, resulting in an SBA titer of <1:4. To test the function of the antibodies produced following immunization with MenC-PS, SBA against MenC strain C11 (C:16:P1.7,1) was also performed (Table 2). SBA titers of 1:128 were obtained with MPL, 1:64 with H44/76-LgtB-LpxL1 nOMVs, and 1:32 using H44/76-lpxL1 nOMVs. No detectable bactericidal antibodies were induced in mice immunized with aluminum hydroxide alone (titer < 1:4). Although the SBA responses to PS immunization are known to largely exceed the responses induced by subcapsular proteins, it should be noted that the H44/76 nOMVs used as the adjuvant share the same VR1 epitope with C11 (P1.7), hence favoring the nOMV-induced response in this setting.
TABLE 2.
SBA titers following immunization with rPorA or MenC-PS against strains BZ198 (B:NT:P1.7-2,4) (following rPorA immunization) or C11 (C:16:P1.7,1) (following MenC immunization)
| Adjuvant | Titer for antigena: |
|
|---|---|---|
| PorA P1.7-2,4 | MenC-PS | |
| H44/76-lpxL1 nOMVs | 1:32 | 1:32 |
| H44/76-lgtB-lpxL1 nOMVs | 1:128 | 1:64 |
| MPL | 1:128 | 1:128 |
| Aluminum hydroxide | <1:4 | <1:4 |
Titers represent the reciprocal of the last dilution at which ≥50% killing was achieved. Serum samples whose first dilution yielded <50% killing of the number of bacteria used were assigned a titer of <1:4.
There was also a possibility of cross-reactivity in the bactericidal antibodies against PorA when nOMVs were used as the adjuvant, since they contain PorA P1.7,16, which has a VR1 sequence that closely resembles the VR1 used as the immunizing antigen, P1.7-2,4. The P1.7-specific mouse monoclonal antibody cross-reacts with P1.7-2 (40). Therefore, a competitive inhibition SBA assay was performed to investigate whether the bactericidal antibodies were specific for PorA P1.7-2,4. Incubation of the mouse serum samples with rPorA P1.7-2,4 protein prior to SBA showed that this removed all bactericidal activity of the sera when analyzed against BZ198 (which expresses P1.7-2,4) as the target strain. A similar adsorption level of serum with the recombinant PorA P1.7,16 protein led to a reduction in bactericidal activity by 75%.
DISCUSSION
LPS is a potent inducer of the innate immune system through activation of the TLR-4/MD2 complex, and as such, it is a promising adjuvant candidate for many vaccines if it can be rendered less toxic (41). The recent discovery that certain genetic modifications, such as the disruption of lpxL1 and lgtB, result in a safer and possibly more stimulatory molecule has underlined the potential of LPS as an adjuvant for human vaccines. In this study, we provide evidence that meningococcal nOMVs containing LpxL1 or LgtB-LpxL1 LPS are potent adjuvants for meningococcal protein antigens. Their activities were comparable to that of MPL with regard to the bactericidal response to meningococcal antigens. With the view of the potential future use of nOMVs in human vaccines, this is encouraging, given that the comparator MPL is a component of adjuvants that have already been licensed for human use (AS01, AS02, and AS04) and have been shown to augment responses to a number of vaccine antigens (4, 42, 43). The protein antigens induced more variable IgG responses when adjuvanted with MPL than with nOMVs or aluminum hydroxide. We suspect that the mixing of MPL (oil) with the antigens (in saline buffers) may have been less homogeneous than with the nOMVs and aluminum hydroxide.
The immune responses elicited with adjuvants were not compared with ones elicited by the antigens alone in the absence of adjuvant. Purified proteins notoriously induce poor immune response on their own, and in this study, modified OMVs were compared with the adjuvant currently used in humans, aluminum hydroxide. However, previous data showed that the immunization of mice with rPorA on its own induced low antigen-specific antibody titers (mean of 21.6 U/ml, ranging from 0.97 to 57.7), while immunization with rPorA plus aluminum hydroxide generated responses on average 12 times higher (mean, 165 U/ml; range, 34.5 to 377 U/ml; P < 0.0001) (our unpublished data).
In addition to the LPS, other components of the nOMVs might also have adjuvant properties for both meningococcal and nonmeningococcal antigens. For example, bacterial lipoproteins are known agonists for TLR2, which may subsequently act as an adjuvant for Th1 immune responses (44). This may influence the induction of bactericidal properties, as discussed below.
The demonstration of bactericidal antibodies following the immunization of mice is the currently accepted animal model used to indicate the likelihood of inducing SBA antibody responses in humans (45). These data demonstrate that nOMVs, containing genetically detoxified LpxL1 LPS, are able to stimulate a functional response against a meningococcal protein antigen, similar to that produced by MPL (26, 46). Of greater importance is our demonstration that both nOMVs were superior to aluminum hydroxide. Aluminum salts are the current adjuvant in two meningococcal protein vaccines that are in phase III clinical trials (47–49), one of which was recently licensed in Europe. Therefore, there is the possibility that the inclusion of nOMVs in future protein vaccines may result in superior SBA activity, with the added benefit of the inclusion of additional antigens contained within the nOMV. The advantage of nOMVs over depleted OMVs (dOMVs) is that they retain some of the proteins that may be lost during detergent extraction, such as factor H binding protein and class 5 proteins, which may be important for stimulating a protective immune response (49).
The inhibition SBA assay confirmed that most of the bactericidal antibodies were targeted against PorA P1.7-2,4, given the complete abrogation of bactericidal activity against a P1.7-2,4-expressing strain when serum samples were preincubated with the same rPorA variant. However, bacteriolysis was also partially inhibited by rPorA P1.7,16, suggesting some cross-reactivity between the two PorA variants. This might be due to the same VR1 family being present in both sequences. However, P1.7-2 is a shorter VR1 sequence, also referred to as a masked or inaccessible subtype epitope, because it only becomes available to the P1.7 monoclonal antibody when it is denatured (40). Most SBA activity against PorA P1.7-2,4 is thus thought to be directed against the VR2 P1,4 sequence.
Native OMVs helped stimulate SBA responses that were similar to those observed with MPL, despite inducing similar or lower levels of antigen-specific total IgG. Only specific IgG subsets can initiate bacteriolysis via the complement cascade. In mice, a lower IgG1/IgG2a ratio indicates a Th1-biased response (50), which is thought to favor the production of bactericidal antibodies (51, 52). The finding that nOMVs resulted in a lower IgG1/IgG2a ratio than MPL or aluminum hydroxide with PorA was consistent with the SBA data, and it is in agreement with the notion that TLR4 agonists and aluminum salts are adjuvants that bias toward Th1 and Th2 responses, respectively (13). Nevertheless, aluminum hydroxide failed to induce any detectable bactericidal activity against BZ198, despite raising a similar level of recombinant PorA P1.7-2,4-specific IgG2a as the other LPS-based adjuvants. This suggests that bactericidal activity is not only dependent on the quantity of IgG2a but also on other factors, such as antibody avidity (53), epitope-specificity of the antibodies (54), and possibly the relative amount of other IgG subclasses.
Purified LpxL1 LPS has been shown to inhibit TLR4-dependent cytokine production in humans but has agonist activity in mice (55, 56). The disruption of lpxL1, which reduces LPS toxicity, may therefore also abrogate any LPS-stimulated adjuvant effect in humans. However, a recent phase I study has shown that LpxL1 LPS contained in a nOMV-based vaccine is able to elicit bactericidal responses in humans (16).
The disruption of lgtB prevents the addition of a galactose (Gal) residue to N-acetylglucosamine (GlcNAc), resulting in the exposure of GlcNAc(β1-3)-Gal(β1-4)-Glc-R in the oligosaccharide outer core of LPS. Steeghs et al. (24) showed that the exposed GlcNAc, attached to the aforementioned oligosaccharide backbone, interacted with the carbohydrate recognition domain of DC-SIGN on the surface of human DCs. This is believed to facilitate the activation of DCs and presentation to cells of the adaptive immune system. There is evidence that the murine homologue of DC-SIGN, mSIGNR1, interacts with similar residues in the outer core of Salmonella LPS (57), which also enhances the activation of the innate immune system (58). In addition, LgtB LPS has also been shown to skew T cells toward a Th1-type response through a currently unknown mechanism (24). However, mSIGNR1 has a very different cellular expression pattern from human DC-SIGN; therefore, the immunological outcome of LgtB LPS injection in mice may not be similar to that in humans (59).
Nevertheless, we did not find convincing evidence of the superiority of lgtB nOMVs. Although the nOMVs were engineered to differ in their LPS glycoforms, the final preparations did contain a number of other differences, namely, the presence of Opa or OpcA and a truncated form of LPS in the H44/76-LpxL1 nOMVs. The presence of the truncated LPS was possibly due to the L3 to L8 immunotype transition due to phase variation of lgtA, which has been shown to occur in both H44/76 and MC58 (60, 61), and the alteration in Opa/OpcA expression was likely also due to phase variation occurring during strain construction (60).
In addition to their potential use in future vaccines consisting of meningococcal protein antigens, these nOMVs elicited similar levels of tetanus toxoid-specific IgG as aluminum hydroxide, which has been employed successfully in human diphtheria-pertussis-tetanus vaccines (62). Therefore, this leads to the possibility that meningococcal nOMVs may be an alternative adjuvant for nonmeningococcal protein antigens, with the additional benefit of providing some protection against meningococcal disease.
We demonstrated that LpxL1 LPS formulated in nOMVs had a significant adjuvant effect with meningococcal PorA, TT, and MenC-PS in mice. These effects were similar to those seen with MPL, which is a potent TLR4 agonist, but which is not approved for use in humans. However, nOMVs have already been tested in phase I trials and were well tolerated (16). The superiority of nOMVs over aluminum hydroxide in this study supports the possibility that nOMVs are a better adjuvant than aluminum salts in novel meningococcal protein vaccines, which are able to skew the immune response toward the production of bactericidal antibodies. They would have the further benefit of providing additional vaccine antigens, which might broaden the coverages of these vaccines. These findings warrant further characterization of these nOMVs and investigation into their safety and effectiveness in humans.
ACKNOWLEDGMENTS
J.C.N. and O.H.M. are supported by funding from the Faculty of Medicine, Dentistry and Health Sciences of the University of Melbourne. C.S.R. is supported by research grants from Meningitis UK, Action Medical Research, SPARKS, and by the NIHR Biomedical Research Centre, Oxford. She is an Oxford Martin fellow. M. Sadarangani was supported through a Research Training Fellowship (no. RTF1263) awarded by Action Medical Research, United Kingdom. J.C.H. was supported by the NIHR Biomedical Research Centre, Oxford. J.P.D. and M. Saleem were funded by a Wellcome Trust Translation award. A.J.P. is supported by the NIHR Oxford Biomedical Research Centre, and he is a Jenner Investigator and James Martin Senior Fellow.
A.J.P. conducts clinical trials on behalf of Oxford University funded by manufacturers of meningococcal vaccines, including Novartis Vaccines, GlaxoSmithKline, Pfizer, and Sanofi Pasteur. He does not receive any personal payments from them or travel reimbursements or honoraria. His department has received unrestricted educational grants from vaccine manufacturers for organization of courses and symposia. I.F. and A.J.P. are named on patents in the field of group B meningococcal vaccines. E.R.M. is a member of the Scientific Advisory Board of Novartis Vaccines and Diagnostics and of the Scientific Advisory Board of GlycoVaxyn.
Footnotes
Published ahead of print 18 December 2013
REFERENCES
- 1.Brunelli B, Del Tordello E, Palumbo E, Biolchi A, Bambini S, Comanducci M, Muzzi A, Pizza M, Rappuoli R, Donnelly JJ, Giuliani MM, Serruto D. 2011. Influence of sequence variability on bactericidal activity sera induced by Factor H binding protein variant 1.1. Vaccine 29:1072–1081. 10.1016/j.vaccine.2010.11.064 [DOI] [PubMed] [Google Scholar]
- 2.Cartwright K, Noah N, Peltola H, Meningococcal Disease Advisory Board 2001. Meningococcal disease in Europe: epidemiology, mortality, and prevention with conjugate vaccines. Report of a European advisory board meeting Vienna, Austria, 6–8 October, 2000. Vaccine 19:4347–4356. 10.1016/S0264-410X(01)00205-5 [DOI] [PubMed] [Google Scholar]
- 3.Finne J, Leinonen M, Mäkelä PH. 1983. Antigenic similarities between brain components and bacteria causing meningitis. Implications for vaccine development and pathogenesis. Lancet 2:355–357 [DOI] [PubMed] [Google Scholar]
- 4.Fries LF, Gordon DM, Richards RL, Egan JE, Hollingdale MR, Gross M, Silverman C, Alving CR. 1992. Liposomal malaria vaccine in humans: a safe and potent adjuvant strategy. Proc. Natl. Acad. Sci. U. S. A. 89:358–362. 10.1073/pnas.89.1.358 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Giuliani MM, Adu-Bobie J, Comanducci M, Aricò B, Savino S, Santini L, Brunelli B, Bambini S, Biolchi A, Capecchi B, Cartocci E, Ciucchi L, Di Marcello F, Ferlicca F, Galli B, Luzzi E, Masignani V, Serruto D, Veggi D, Contorni M, Morandi M, Bartalesi A, Cinotti V, Mannucci D, Titta F, Ovidi E, Welsch JA, Granoff D, Rappuoli R, Pizza M. 2006. A universal vaccine for serogroup B meningococcus. Proc. Natl. Acad. Sci. U. S. A. 103:10834–10839. 10.1073/pnas.0603940103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ruijne N, Lea RA, O'Hallahan J, Oster P, Martin D. 2006. Understanding the immune responses to the meningococcal strain-specific vaccine MeNZB measured in studies of infants. Clin. Vaccine Immunol. 13:797–801. 10.1128/CVI.00038-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sierra GV, Campa HC, Varcacel NM, Garcia IL, Izquierdo PL, Sotolongo PF, Casanueva GV, Rico CO, Rodriguez CR, Terry MH. 1991. Vaccine against group B Neisseria meningitidis: protection trial and mass vaccination results in Cuba. NIPH Ann. 14:195–207; discussion 208–110 [PubMed] [Google Scholar]
- 8.Gorringe AR, Pajón R. 2012. Bexsero: a multicomponent vaccine for prevention of meningococcal disease. Hum. Vaccin. Immunother. 8:174–183. 10.4161/hv.18500 [DOI] [PubMed] [Google Scholar]
- 9.Tappero JW, Lagos R, Ballesteros AM, Plikaytis B, Williams D, Dykes J, Gheesling LL, Carlone GM, Høiby EA, Holst J, Nøkleby H, Rosenqvist E, Sierra G, Campa C, Sotolongo F, Vega J, Garcia J, Herrera P, Poolman JT, Perkins BA. 1999. Immunogenicity of 2 serogroup B outer-membrane protein meningococcal vaccines: a randomized controlled trial in Chile. JAMA 281:1520–1527 [DOI] [PubMed] [Google Scholar]
- 10.Verheul AF, Van Gaans JA, Wiertz EJ, Snippe H, Verhoef J, Poolman JT. 1993. Meningococcal lipopolysaccharide (LPS)-derived oligosaccharide-protein conjugates evoke outer membrane protein- but not LPS-specific bactericidal antibodies in mice: influence of adjuvants. Infect. Immun. 61:187–196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.MacLeod MK, McKee AS, David A, Wang J, Mason R, Kappler JW, Marrack P. 2011. Vaccine adjuvants aluminum and monophosphoryl lipid A provide distinct signals to generate protective cytotoxic memory CD8 T cells. Proc. Natl. Acad. Sci. U. S. A. 108:7914–7919. 10.1073/pnas.1104588108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rappuoli R, Mandl CW, Black S, De Gregorio E. 2011. Vaccines for the twenty-first century society. Nat. Rev. Immunol. 11:865–872. 10.1038/nri3085 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Moingeon P, Haensler J, Lindberg A. 2001. Towards the rational design of Th1 adjuvants. Vaccine 19:4363–4372. 10.1016/S0264-410X(01)00193-1 [DOI] [PubMed] [Google Scholar]
- 14.Holst J, Martin D, Arnold R, Huergo CC, Oster P, O'Hallahan J, Rosenqvist E. 2009. Properties and clinical performance of vaccines containing outer membrane vesicles from Neisseria meningitidis. Vaccine 27(Suppl 2):B3–B12. 10.1016/j.vaccine.2009.04.071 [DOI] [PubMed] [Google Scholar]
- 15.Rosenqvist E, Høiby EA, Bjune G, Aase A, Halstensen A, Lehmann AK, Paulssen J, Holst J, Michaelsen TE, Nøkleby H, Frøholm LO, Closs O. 1998. Effect of aluminium hydroxide and meningococcal serogroup C capsular polysaccharide on the immunogenicity and reactogenicity of a group B Neisseria meningitidis outer membrane vesicle vaccine. Dev. Biol. Stand. 92:323–333 [PubMed] [Google Scholar]
- 16.Keiser PB, Biggs-Cicatelli S, Moran EE, Schmiel DH, Pinto VB, Burden RE, Miller LB, Moon JE, Bowden RA, Cummings JF, Zollinger WD. 2011. A phase 1 study of a meningococcal native outer membrane vesicle vaccine made from a group B strain with deleted lpxL1 and synX, over-expressed factor H binding protein, two PorAs and stabilized OpcA expression. Vaccine 29:1413–1420. 10.1016/j.vaccine.2010.12.039 [DOI] [PubMed] [Google Scholar]
- 17.Jäkel A, Plested JS, Hoe JC, Makepeace K, Gidney MA, Lacelle S, St Michael F, Cox AD, Richards JC, Moxon ER. 2008. Naturally-occurring human serum antibodies to inner core lipopolysaccharide epitopes of Neisseria meningitidis protect against invasive meningococcal disease caused by isolates displaying homologous inner core structures. Vaccine 26:6655–6663. 10.1016/j.vaccine.2008.09.041 [DOI] [PubMed] [Google Scholar]
- 18.Plested JS, Harris SL, Wright JC, Coull PA, Makepeace K, Gidney MA, Brisson JR, Richards JC, Granoff DM, Moxon ER. 2003. Highly conserved Neisseria meningitidis inner-core lipopolysaccharide epitope confers protection against experimental meningococcal bacteremia. J. Infect. Dis. 187:1223–1234. 10.1086/368360 [DOI] [PubMed] [Google Scholar]
- 19.Plested JS, Makepeace K, Jennings MP, Gidney MA, Lacelle S, Brisson J, Cox AD, Martin A, Bird AG, Tang CM, Mackinnon FM, Richards JC, Moxon ER. 1999. Conservation and accessibility of an inner core lipopolysaccharide epitope of Neisseria meningitidis. Infect. Immun. 67:5417–5426 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Beutler B, Rietschel ET. 2003. Innate immune sensing and its roots: the story of endotoxin. Nat. Rev. Immunol. 3:169–176. 10.1038/nri1004 [DOI] [PubMed] [Google Scholar]
- 21.Naess LM, Aarvak T, Aase A, Oftung F, Høiby EA, Sandin R, Michaelsen TE. 1999. Human IgG subclass responses in relation to serum bactericidal and opsonic activities after immunization with three doses of the Norwegian serogroup B meningococcal outer membrane vesicle vaccine. Vaccine 17:754–764. 10.1016/S0264-410X(98)00259-X [DOI] [PubMed] [Google Scholar]
- 22.van der Ley P, Steeghs L, Hamstra HJ, ten Hove J, Zomer B, van Alphen L. 2001. Modification of lipid A biosynthesis in Neisseria meningitidis lpxL mutants: influence on lipopolysaccharide structure, toxicity, and adjuvant activity. Infect. Immun. 69:5981–5990. 10.1128/IAI.69.10.5981-5990.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhang P, Snyder S, Feng P, Azadi P, Zhang S, Bulgheresi S, Sanderson KE, He J, Klena J, Chen T. 2006. Role of N-acetylglucosamine within core lipopolysaccharide of several species of gram-negative bacteria in targeting the DC-SIGN (CD209). J. Immunol. 177:4002–4011 [DOI] [PubMed] [Google Scholar]
- 24.Steeghs L, van Vliet SJ, Uronen-Hansson H, van Mourik A, Engering A, Sanchez-Hernandez M, Klein N, Callard R, van Putten JP, van der Ley P, van Kooyk Y, van de Winkel JG. 2006. Neisseria meningitidis expressing lgtB lipopolysaccharide targets DC-SIGN and modulates dendritic cell function. Cell. Microbiol. 8:316–325. 10.1111/j.1462-5822.2005.00623.x [DOI] [PubMed] [Google Scholar]
- 25.Jennings MP, Hood DW, Peak IR, Virji M, Moxon ER. 1995. Molecular analysis of a locus for the biosynthesis and phase-variable expression of the lacto-N-neotetraose terminal lipopolysaccharide structure in Neisseria meningitidis. Mol. Microbiol. 18:729–740. 10.1111/j.1365-2958.1995.mmi_18040729.x [DOI] [PubMed] [Google Scholar]
- 26.Borrow R, Aaberge IS, Santos GF, Eudey TL, Oster P, Glennie A, Findlow J, Høiby EA, Rosenqvist E, Balmer P, Martin D. 2005. Interlaboratory standardization of the measurement of serum bactericidal activity by using human complement against meningococcal serogroup b, strain 44/76-SL, before and after vaccination with the Norwegian MenBvac outer membrane vesicle vaccine. Clin. Diagn. Lab. Immunol. 12:970–976. 10.1128/CDLI.12.8.970-976.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Criss AK, Kline KA, Seifert HS. 2005. The frequency and rate of pilin antigenic variation in Neisseria gonorrhoeae. Mol. Microbiol. 58:510–519. 10.1111/j.1365-2958.2005.04838.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gunn JS, Stein DC. 1996. Use of a non-selective transformation technique to construct a multiply restriction/modification-deficient mutant of Neisseria gonorrhoeae. Mol. Gen. Genet. 251:509–517. 10.1007/BF02173639 [DOI] [PubMed] [Google Scholar]
- 29.Sadarangani M, Hoe JC, Callaghan MJ, Jones C, Chan H, Makepeace K, Daniels-Treffandier H, Deadman ME, Bayliss C, Feavers I, Van der Ley P, Pollard AJ. 2012. Construction of Opa-positive and Opa-negative strains of Neisseria meningitidis to evaluate a novel meningococcal vaccine. PLoS One 7:e51045. 10.1371/journal.pone.0051045 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Zollinger WD, Mandrell RE, Griffiss JM, Altieri P, Berman S. 1979. Complex of meningococcal group B polysaccharide and type 2 outer membrane protein immunogenic in man. J. Clin. Invest. 63:836–848. 10.1172/JCI109383 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Vipond C, Wheeler JX, Jones C, Feavers IM, Suker J. 2005. Characterization of the protein content of a meningococcal outer membrane vesicle vaccine by polyacrylamide gel electrophoresis and mass spectrometry. Hum. Vaccin. 1:80–84. 10.4161/hv.1.2.1651 [DOI] [PubMed] [Google Scholar]
- 32.Lesse AJ, Campagnari AA, Bittner WE, Apicella MA. 1990. Increased resolution of lipopolysaccharides and lipooligosaccharides utilizing tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis. J. Immunol. Methods 126:109–117. 10.1016/0022-1759(90)90018-Q [DOI] [PubMed] [Google Scholar]
- 33.Saleem M, Moore J, Derrick JP. 2012. Expression, purification, and crystallization of neisserial outer membrane proteins. Methods Mol. Biol. 799:91–106. 10.1007/978-1-61779-346-2_6 [DOI] [PubMed] [Google Scholar]
- 34.Manghi MA, Pasetti MF, Brero ML, Deluchi S, di Paola G, Mathet V, Eriksson PV. 1994. Development of an ELISA for measuring the activity of tetanus toxoid in vaccines and comparison with the toxin neutralization test in mice. J. Immunol. Methods 168:17–24. 10.1016/0022-1759(94)90204-6 [DOI] [PubMed] [Google Scholar]
- 35.Plikaytis BD, Turner SH, Gheesling LL, Carlone GM. 1991. Comparisons of standard curve-fitting methods to quantitate Neisseria meningitidis group A polysaccharide antibody levels by enzyme-linked immunosorbent assay. J. Clin. Microbiol. 29:1439–1446 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Borrow R, Carlone GM. 2001. Serogroup B and C serum bactericidal assays, p 289–304 In Pollard AJ, Maiden MC. (ed), Meningococcal vaccines. Methods and protocols. Humana Press, Inc., Totowa, New Jersey: [DOI] [PubMed] [Google Scholar]
- 37.Giuliani MM, Biolchi A, Serruto D, Ferlicca F, Vienken K, Oster P, Rappuoli R, Pizza M, Donnelly J. 2010. Measuring antigen-specific bactericidal responses to a multicomponent vaccine against serogroup B meningococcus. Vaccine 28:5023–5030. 10.1016/j.vaccine.2010.05.014 [DOI] [PubMed] [Google Scholar]
- 38.Coffman RL, Seymour BW, Lebman DA, Hiraki DD, Christiansen JA, Shrader B, Cherwinski HM, Savelkoul HF, Finkelman FD, Bond MW. 1988. The role of helper T cell products in mouse B cell differentiation and isotype regulation. Immunol. Rev. 102:5–28. 10.1111/j.1600-065X.1988.tb00739.x [DOI] [PubMed] [Google Scholar]
- 39.DeKruyff RH, Mosmann RR, Umetsu DT. 1990. Induction of antibody synthesis by CD4+ T cells: IL 5 is essential for induction of antigen-specific antibody responses by TH2 but not TH1 clones. Eur. J. Immunol. 20:2219–2227. 10.1002/eji.1830201010 [DOI] [PubMed] [Google Scholar]
- 40.Wedege E, Dalseg R, Caugant DA, Poolman JT, Frøholm LO. 1993. Expression of an inaccessible P1.7 subtype epitope on meningococcal class 1 proteins. J. Med. Microbiol. 38:23–28. 10.1099/00222615-38-1-23 [DOI] [PubMed] [Google Scholar]
- 41.Steinhagen F, Kinjo T, Bode C, Klinman DM. 2011. TLR-based immune adjuvants. Vaccine 29:3341–3355. 10.1016/j.vaccine.2010.08.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.GlaxoSmithKline Vaccine HPV-007 Study Group, Romanowski B, de Borba PC, Naud PS, Roteli-Martins CM, De Carvalho NS, Teixeira JC, Aoki F, Ramjattan B, Shier RM, Somani R, Barbier S, Blatter MM, Chambers C, Ferris D, Gall SA, Guerra FA, Harper DM, Hedrick JA, Henry DC, Korn AP, Kroll R, Moscicki AB, Rosenfeld WD, Sullivan BJ, Thoming CS, Tyring SK, Wheeler CM, Dubin G, Schuind A, Zahaf T, Greenacre M, Sgriobhadair A. 2009. Sustained efficacy and immunogenicity of the human papillomavirus (HPV)-16/18 AS04-adjuvanted vaccine: analysis of a randomised placebo-controlled trial up to 6.4 years. Lancet 374:1975–1985. 10.1016/S0140-6736(09)61567-1 [DOI] [PubMed] [Google Scholar]
- 43.Vandepapelière P, Horsmans Y, Moris P, Van Mechelen M, Janssens M, Koutsoukos M, Van Belle P, Clement F, Hanon E, Wettendorff M, Garçon N, Leroux-Roels G. 2008. Vaccine adjuvant systems containing monophosphoryl lipid A and QS21 induce strong and persistent humoral and T cell responses against hepatitis B surface antigen in healthy adult volunteers. Vaccine 26:1375–1386. 10.1016/j.vaccine.2007.12.038 [DOI] [PubMed] [Google Scholar]
- 44.Sieling PA, Chung W, Duong BT, Godowski PJ, Modlin RL. 2003. Toll-like receptor 2 ligands as adjuvants for human Th1 responses. J. Immunol. 170:194–200 [DOI] [PubMed] [Google Scholar]
- 45.Beninati C, Midiri A, Mancuso G, Biondo C, Arigò M, Gerace E, Papasergi S, Gambuzza M, Boretti M, Magliani W, Conti S, Polonelli L, Teti G. 2006. Antiidiotypic DNA vaccination induces serum bactericidal activity and protection against group B meningococci. J. Exp. Med. 203:111–118. 10.1084/jem.20051540 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Borrow R, Richmond P, Kaczmarski EB, Iverson A, Martin SL, Findlow J, Acuna M, Longworth E, O'Connor R, Paul J, Miller E. 2000. Meningococcal serogroup C-specific IgG antibody responses and serum bactericidal titres in children following vaccination with a meningococcal A/C polysaccharide vaccine. FEMS Immunol. Med. Microbiol. 28:79–85. 10.1111/j.1574-695X.2000.tb01460.x [DOI] [PubMed] [Google Scholar]
- 47.Jiang HQ, Hoiseth SK, Harris SL, McNeil LK, Zhu D, Tan C, Scott AA, Alexander K, Mason K, Miller L, DaSilva I, Mack M, Zhao XJ, Pride MW, Andrew L, Murphy E, Hagen M, French R, Arora A, Jones TR, Jansen KU, Zlotnick GW, Anderson AS. 2010. Broad vaccine coverage predicted for a bivalent recombinant factor H binding protein based vaccine to prevent serogroup B meningococcal disease. Vaccine 28:6086–6093. 10.1016/j.vaccine.2010.06.083 [DOI] [PubMed] [Google Scholar]
- 48.Lee AW, Vesikari T, Gilbert CL, Klopfer SO, Schödel FP, Bhuyan PK. 2011. Immunogenicity and safety of a Haemophilus influenzae B (Hib)-hepatitis B vaccine with a modified process hepatitis B component administered with concomitant pneumococcal conjugate vaccine to infants. Vaccine 29:7942–7948. 10.1016/j.vaccine.2011.08.071 [DOI] [PubMed] [Google Scholar]
- 49.Koeberling O, Giuntini S, Seubert A, Granoff DM. 2009. Meningococcal outer membrane vesicle vaccines derived from mutant strains engineered to express factor H binding proteins from antigenic variant groups 1 and 2. Clin. Vaccine Immunol. 16:156–162. 10.1128/CVI.00403-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Arigita C, Luijkx T, Jiskoot W, Poelen M, Hennink WE, Crommelin DJ, Ley P, Els C, Kersten GF. 2005. Well-defined and potent liposomal meningococcal B vaccines adjuvated with LPS derivatives. Vaccine 23:5091–5098. 10.1016/j.vaccine.2005.06.001 [DOI] [PubMed] [Google Scholar]
- 51.Hoogerhout P, Donders EM, van Gaans-van den Brink JA, Kuipers B, Brugghe HF, van Unen LM, Timmermans HA, ten Hove GJ, de Jong AP, Peeters CC. 1995. Conjugates of synthetic cyclic peptides elicit bactericidal antibodies against a conformational epitope on a class 1 outer membrane protein of Neisseria meningitidis. Infect. Immun. 63:3473–3478 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Mosmann TR, Coffman RL. 1989. TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu. Rev. Immunol. 7:145–173. 10.1146/annurev.iy.07.040189.001045 [DOI] [PubMed] [Google Scholar]
- 53.Granoff DM, Maslanka SE, Carlone GM, Plikaytis BD, Santos GF, Mokatrin A, Raff HV. 1998. A modified enzyme-linked immunosorbent assay for measurement of antibody responses to meningococcal C polysaccharide that correlate with bactericidal responses. Clin. Diagn. Lab. Immunol. 5:479–485 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Martin DR, Ruijne N, McCallum L, O'Hallahan J, Oster P. 2006. The VR2 epitope on the PorA P1.7-2,4 protein is the major target for the immune response elicited by the strain-specific group B meningococcal vaccine MeNZB. Clin. Vaccine Immunol. 13:486–491. 10.1128/CVI.13.4.486-491.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Sprong T, Ley P, Abdollahi-Roodsaz S, Joosten L, Meer Jv, Netea M, Deuren Mv. 2011. Neisseria meningitidis lipid A mutant LPSs function as LPS antagonists in humans by inhibiting TLR 4-dependent cytokine production. Innate Immun. 17:517–525. 10.1177/1753425910383999 [DOI] [PubMed] [Google Scholar]
- 56.Steeghs L, Keestra AM, van Mourik A, Uronen-Hansson H, van der Ley P, Callard R, Klein N, van Putten JP. 2008. Differential activation of human and mouse Toll-like receptor 4 by the adjuvant candidate LpxL1 of Neisseria meningitidis. Infect. Immun. 76:3801–3807. 10.1128/IAI.00005-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lindberg AA, Hellerqvist CG. 1980. Rough mutants of Salmonella Typhimurium: immunochemical and structural analysis of lipopolysaccharides from rfaH mutants. J. Gen. Microbiol. 116:25–32 [DOI] [PubMed] [Google Scholar]
- 58.Nagaoka K, Takahara K, Tanaka K, Yoshida H, Steinman RM, Saitoh S, Akashi-Takamura S, Miyake K, Kang YS, Park CG, Inaba K. 2005. Association of SIGNR1 with TLR4-MD-2 enhances signal transduction by recognition of LPS in gram-negative bacteria. Int. Immunol. 17:827–836. 10.1093/intimm/dxh264 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kang YS, Yamazaki S, Iyoda T, Pack M, Bruening SA, Kim JY, Takahara K, Inaba K, Steinman RM, Park CG. 2003. SIGN-R1, a novel C-type lectin expressed by marginal zone macrophages in spleen, mediates uptake of the polysaccharide dextran. Int. Immunol. 15:177–186. 10.1093/intimm/dxg019 [DOI] [PubMed] [Google Scholar]
- 60.de Vries FP, van Der Ende A, van Putten JP, Dankert J. 1996. Invasion of primary nasopharyngeal epithelial cells by Neisseria meningitidis is controlled by phase variation of multiple surface antigens. Infect. Immun. 64:2998–3006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Jennings MP, Srikhanta YN, Moxon ER, Kramer M, Poolman JT, Kuipers B, van der Ley P. 1999. The genetic basis of the phase variation repertoire of lipopolysaccharide immunotypes in Neisseria meningitidis. Microbiology 145(Pt 11):3013–3021 [DOI] [PubMed] [Google Scholar]
- 62.Tritto E, Mosca F, De Gregorio E. 2009. Mechanism of action of licensed vaccine adjuvants. Vaccine 27:3331–3334. 10.1016/j.vaccine.2009.01.084 [DOI] [PubMed] [Google Scholar]