

Abstract
Resveratrol, a polyphenolic-stilbene, has received increased attention in the last decade due to its wide range of biological activities. Beta(β)-thalassemias are inherited red cell disorders, found worldwide, characterized by ineffective erythropoiesis and red cell oxidative damage with reduced survival. We evaluated the effects of low-dose-resveratrol (5 μM) on in vitro human erythroid differentiation of CD34+ from normal and β-thalassemic subjects. We found that resveratrol induces accelerated erythroid-maturation, resulting in the reduction of colony-forming units of erythroid cells and increased intermediate and late erythroblasts. In sorted colony-forming units of erythroid cells resveratrol activates Forkhead-box-class-O3, decreases Akt activity and up-regulates anti-oxidant enzymes as catalase. In an in vivo murine model for β-thalassemia, resveratrol (2.4 mg/kg) reduces ineffective erythropoiesis, increases hemoglobin levels, reduces reticulocyte count and ameliorates red cell survival. In both wild-type and β-thalassemic mice, resveratrol up-regulates scavenging enzymes such as catalase and peroxiredoxin-2 through Forkhead-box-class-O3 activation. These data indicate that resveratrol inhibits Akt resulting in FoxO3 activation with upregulation of cytoprotective systems enabling the pathological erythroid precursors to resist the oxidative damage and continue to differentiate. Our data suggest that the dual effect of resveratrol on erythropoiesis through activation of FoxO3 transcriptional factor combined with the amelioration of oxidative stress in circulating red cells may be considered as a potential novel therapeutic strategy in treating β-thalassemia.
Introduction
Resveratrol, a polyphenolic-stilbene, has received increasing attention in the last decade due to its wide range of biological activities, which include antioxidant, anti-inflammatory and anti-tumoral effects.1–3 Although some progress has been made in the identification of the mechanism(s) underlying the various beneficial effects of resveratrol, much still remains to be investigated.3,4 Most of the studies carried out to evaluate the effects of resveratrol on erythropoiesis have been performed using K562 erythroleukemia cell lines5–7 and therefore very limited information is available regarding the effect of resveratrol on normal erythropoiesis.8 In these studies, resveratrol has been shown to increase fetal hemoglobin synthesis (50 μM resveratrol),6,9 to attenuate the TNF-α effects on erythropoiesis (0.4 μM, 10 – 30 μM resveratrol)8 and to block cell growth affecting cell cycle and redirecting cells towards either apopotosis or differentiation (60 μM resveratrol).5–7 Recently, the beneficial effects of resveratrol supplementation on pathological erythropoiesis have been reported in a mouse model for Fanconi’s anemia (FA), which is characterized by the hypersensitivity of FA cells to reactive oxygen species (ROS).10 The dynamic process of erythroid differentiation is characterized by the production of reactive oxygen species (ROS) both in response to erythropoietin signaling and to the large amount of iron imported into the cells during heme biosynthesis.11 The intracellular response to oxidative-stress in erythropoiesis involves the transcription factor, Forkhead box O3a (FOxO3), which controls pathway(s) regulating erythroid maturation and the levels of oxidative stress in murine erythropoiesis.12,13 FOxO3a is negatively regulated by the serine-threonine kinase Akt, which phosphorylates FOxO3a promoting its translocation from the nucleus to the cytoplasm and resulting in inhibition of FOxO3 transcriptional activity.12–14 Activation of FoxO3a has been proposed as a protective mechanism in pathological erythropoiesis characterized by abnormal ROS levels such as β-thalassemia.12
β-thalassemias (β-thal) are common inherited red cell disorders characterized by absent or reduced synthesis of β-globin chains. Despite extensive knowledge of the molecular defects causing β-thalassemia, less is known about the mechanisms responsible for the associated ineffective erythropoiesis and reduced red cell survival.11,15–20 Increased levels of reactive oxygen species (ROS) have been reported to contribute to the anemia of β-thalassemia, although the effects of ROS have not been fully defined.11,15–18 Exogenous anti-oxidant molecules might represent complementary therapeutic strategies to counteract the toxic effects of ROS in β-thalassemia. However, few of them have been shown to beneficially affect in vivo β-thalassemic red cell features and/or thalassemic ineffective erythropoiesis in vivo.16,21
Methods
In vitro erythropoiesis from CD34+ cells from peripheral circulation of normal and β-thalassemia-intermedia subjects
Regarding cell culture, phenotypic analysis and cell sorting strategy, peripheral blood from adult normal volunteers and from transfusion independent β-thalassemia patients (β-thalassemia intermedia) was collected, after obtaining informed consent according to the guidelines established by the Ethic Committee for human subject studies of the University of Milan and the principles of the Declaration of Helsinki. Approval by the Ethic Committee of the University of Milan for human erythroid precursors studies was obtained. We analyzed 20 erythroid cultures from the peripheral blood of different normal subjects and 20 erythroid cultures from 10 homozygous β-thalassemic intermedia patients (β0cod39).11,17 Details on cell cultures are reported in the Online Supplementary Appendix. The Resveratrol concentration (5 μM; Sigma Aldrich, St. Louis, MO, USA) used in this study was selected from dose-response studies (Online Supplementary Figure 1SA) and a review of the literature.3,5,6,8–10,22 The erythroid cell antigen profile and the sorting of erythroid precursors were carried out as reported by Merry-Weather Clarke et al.23 Details are reported in the Online Supplementary Appendix and Figure S1B).
Quantitative real-time PCR was carried out as previously described.24 Details are reported in the Online Supplementary Appendix. The primers used are listed in Online Supplementary Tables S1 and S2.
For immunoblot-analysis of sorted human erythroid precursors and immunofluorescence assay for FoxO3a, 1 × 106 sorted CFU-E cells from both normal and β-thalassemic were solubilized as previously described.11,20 Details on immunoblot and immunofluorescence analysis are reported in the Online Supplementary Appendix. Whenever indicated, sorted CFU-Es were separated into cytosol and nuclear fractions as previously reported.25
The study was carried out in accordance with the Scientific Committee for Animal Experimentation (CIRSAL, University of Verona, Italy). C57B6/2J mice, wild-type controls (WT) and Hbbth3/+ mice were used as β-thalassemia models. Age- and sex-matched 2-month old mice (weight 20 g) were studied. The female/male ratio in the different groups was 1:1. Based on previous studies on resveratrol bioavailability in vivo,26–28 the mice were placed either on resveratrol (2.5 mg/kg incorporated into AIN-93G diet) or standard diet (AIN-93G diet). The mice were fed with resveratrol diet for six months. Hematologic parameters and red cell indices were determined as previously reported.16,29–32
For cytofluorimetric analysis of mouse bone-marrow precursors and cell sorting of murine erythroblasts, the cytofluorimetric analysis of erythroid precursors from bone marrow (BM) of both wild-type and β-thalassemic mice was carried out as previously described.33 Populations (pop.) II and III corresponding to basophilic erythroblasts and polychromatic erythroblasts (1 × 106) were sorted from BM of both mouse strains for RT-PCR and immunoblot analysis. Red cell survival was carried out using CFSE (10 μM; Molecular Probe, Invitrogen).4 Details are reported in the Online Supplementary Appendix.
Red cell ghost preparation30,36 and red cell membrane carbonyl group determination were carried out as previously reported36 (Online Supplementary Appendix).
FOXO-PRDX2 alignment analysis is described in the Online Supplementary Appendix, along with details of the statistical analysis carried out.
Results
Low concentration of resveratrol induces erythroid maturation during normal erythropoiesis
We first evaluated the effects of different doses of resveratrol on the production of normal erythroid cells generated from CD34+ cells isolated from peripheral blood. We found a dose-dependent decrease in cell production in agreement with earlier reports that show cellular toxic effects on differentiating cells at both moderate and high doses of resveratrol (Online Supplementary Figure S1A).5,6,8 We thus chose to work with a low dose of resveratrol (5 μM) and noted that even at this dose cell proliferation was decreased as compared to untreated cells (Figure 1A). Erythroid differentiation was assessed using a combination of three surface markers: the transferrin receptor (CD71) and Glycophorin A (GPA) and CD36,23 which enable the identification of erythroid cells at distinct stages of maturation (Figure 1B and Online Supplementary Figure S1B). Resveratrol induced a reduction of CFU-E cells but increased Int-E at Days 7 and 9 of culture (early erythropoiesis) as well as Int-E and Late-E at Days 11 and 14 of culture (late erythropoiesis) (Figure 1B). The accelerated maturation of erythroid cells induced by resveratrol was reflected by changes in cell morphology (Figure 1B), by increased surface expression of GPA, reduced expression of CD71 at an earlier time compared to untreated cells (Figure 2A), and early appearance of band 3 (Online Supplementary Figure S1C). No differences in the extent of cell apoptosis as monitored by the percentage of Annexin-V positive cells (7d: untreated 3.5±0.7% vs. resveratrol 3.1±0.4%, n= 6; NS; 11d: untreated 2.25±0.1% vs. resveratrol 1.9±0.8%, n=6; NS; 13 d: untreated 3.5±0.7% vs. resveratrol 3.1±0.4%, n= 6; NS; 11d: untreated 2.3±0.7% vs. resveratrol 2.0±0.1%, n=6; NS;) or in the expression of levels of gamma globin mRNA were noted in sorted cell erythroid populations from cultures with and without resveratrol (9d: HBG1 untreated 6.8±2.1 vs. resveratrol 9.3±0.9 n=6; NS; HBG2 untreated 8.2±2.1 vs. resveratrol 8.8±0.1 n=6; NS; 13d: HBG1 untreated 19.3±0.5 vs. resveratrol 17.2±2.7 n=6; NS; HBG2 untreated 19.7±0.6 vs. resveratrol 19.5±0.7 n=6; NS; HBG1 and HBG2 relative expression on GAPDH). These data imply that, while resveratrol inhibits proliferation of erythroid progenitors, it accelerates the terminal erythroid differentiation of proerythroblasts into late stage orthochromatic-erythroblasts.
Figure 1.

Low-dose resveratrol hamper s cell growth and affects the pattern of erythroid maturation in normal erythropoiesis. (A) Cell proliferation of erythroid precursors derived by in vitro liquid culture of CD34+ cells isolated from peripheral blood of normal (control cells) subjects with or without resveratrol (n=10). Arrows indicate when resveratrol 5 μM was added to the culture medium. Data are presented as means± SD; *P<0.05 compared to untreated cells. (B) (Left). Cytofluorimetric analysis of maturation pattern of erythroid precursors at different times of cell culture, 7, 9, 11, and 14 days (d) using the following surface marker s: CD36, glycophorin-A and CD71 (see also Online Supplementary Methods). This cytofluorimetric strategy allows the identification of the following homogenous cell populations: pro-erythroblasts (Pro-E), basophilic erythroblasts corresponding to intermediate erythroblasts (Int-E) and polychromatic and orthochromatic erythroblasts as late erythroblasts (Late). Data are expressed as percentages or as absolute cell counts and shown as means ± SD (n=10); *P< 0.05 compared to untreated cells (Right) morphology of erythroid precursors with or without resveratrol. Cytospins were stained with May-Grunwald-Giemsa. Cells were imaged under oil at 100× magnification using a Panfluor objective with 1.30 numeric aperture on a Nikon Eclipse DS-5M camera and processed with Digital Slide (DS-L1) Nikon. One representative image from a total of 10 for each condition at the different time points is shown.
Figure 2.

Low-dose resveratrol induces early erythroid maturation, activates FOxO3a and inhibits Akt pathway (A) (Upper panel). Flow cytometric analysis of expression of transmembrane glycophorin-A (GPA) CD71 during erythropoiesis at days (d) 7, 9, 11, and 14 of culture with or without resveratrol. Note the early appearance of GPA and the early reduction of CD71 in resveratrol-treated cells compared to untreated cells. One representative image from a total of 10 for each condition different time points is shown. Lower panel. Kinetic of GPA appearance and reduction of CD71 in resveratrol treated cells compared to untreated ones. Data are presented as means ± SD (n=10); *P< 0.05 compared to untreated cells. (B) FoxO3 immunostaining of CFU-E cells at Day 7 (7d) of culture with or without resveratrol. Cells were FACS-sorted, cytospun onto glass slides and immunostained with anti-FoxO3a antibody and counterstained with DAPI. The mean fluorescence was measured in the nucleus of 30 cells using Image J software. *P<0.05 compared to untreated cells (n=6). (C) Immunoblot analysis of FoxO3a on nucleus of sorted CFU-E at Day 7 of culture. Histone-H3 was used as loading control. representative gel from the other 6 with similar results is presented. (Right). Relative quantification of immunoreactivity of FoxO3 and Histone-sorted CFU-E cells. Data are presented as FoxO3/Histone-H3 ratio and shown as means ±SD (n=6). *P<0.05 compared to untreated cells. (D) Western-blot (Wb) analysis of phospho-Akt (p-Akt) and Akt in sorted CFU-E cells at Day 7 of culture with resveratrol (Resv) or without (control, C). Tubulin was used as protein loading control. One representative gel from the other 6 with similar results is presented./Right). Relative quantification of immunoreactivity of phospho-Akt (p-Akt), Akt and tubulin in sorted CFU-E cells. Data are presented as p-Akt/tubulin or Akt/tubulin ratio and shown as means ±SD (n=6); *P<0.05 compared to untreated cells.
Since we recently reported that resveratrol targets the transcription factor, forkhead box O3a (FOxO3a),4 which plays a key role in erythropoiesis,12 we evaluated FOxO3a and Akt activation in sorted CFU-E cells.37,38
Resveratrol enhances the expression of FOxO3a and inhibits Akt activity in sorted CFU-E
In sorted CFU-E cells, at Day 7 resveratrol up-regulates FOXO3 expression without a significant change in FOXO1 mRNA levels (Online Supplementary Figure S2A).
Nuclear localization of FoxO3a was used as a surrogate assay for FoxO3a activity. In sorted CFU-E cells, we found that resveratrol increased the overall expression of FoxO3a protein in the nucleus (Figure 2B), which was confirmed by immunoblot analysis (Figure 2C). FoxO3a function is modulated by Akt activity, which phosphorylates FoxO3a, thereby sequestrating it into the cytoplasm and blocking its transcriptional activity.12,39,40 We then evaluated Akt activity in CFU-E cells sorted in the presence and absence of resveratrol. We found reduced phosphorylation of Akt in resveratrol treated CFU-E obtained from cultures at Day 7 compared to untreated cultures (Figure 2D). These data suggest that resveratrol enhances FOxO3 expression in CFU-E, inhibits Akt and modulates the activity of FoxO3a, sustaining erythroid precursors during accelerated erythropoiesis (Figure 2). The activation of FoxO3a is also supported by the finding of upregulation of catalase, a scavenging enzyme (Online Supplementary Figure S2B), whose transcription is promoted by FoxO3a.12
Low concentration of resveratrol hampers cell proliferation and induces cell differentiation of human β-thalassemic-erythroid cells in vitro
To evaluate the effect of resveratrol on disordered erythropoiesis, we chose β-thalassemia as a model since its erythroid differentiation is characterized by blocked cell maturation, oxidative stress and ineffective erythropoiesis.11,15,17,41,42 CD34+ cells derived from peripheral blood of β-thalassemia intermedia subjects were studied in vitro (Online Supplementary Figure S3A and B). This culture system recapitulates the ineffective erythropoiesis observed in vivo in β-thalassemia.11,17 Resveratrol significantly reduced cell proliferation of β-thalassemic erythroid precursors during the early phase of erythropoiesis (7d) (Figure 3A), with a reduction in the percentage of CFU-E cells and an increase of Int-E cells. At late stage of erythropoiesis (14d) there was a reduction of basophilic erythroblasts (Int-E) with a concomitant increase of polychromatophilic- and orthochromatic-erythroblasts (late-E), indicating a shift toward normal maturation and erythropoiesis (Figure 3B). The increased surface expression of GPA and reduced expression of CD71 at an earlier time of culture of β-thalassemic cells in the presence of resveratrol compared to untreated cells also imply accelerated terminal erythroid differentiation (Online Supplementary Figure S3C). No changes in the gamma-globin-chain mRNA levels were detected in sorted β-thalassemic cells with or without resveratrol (data not shown).
Figure 3.
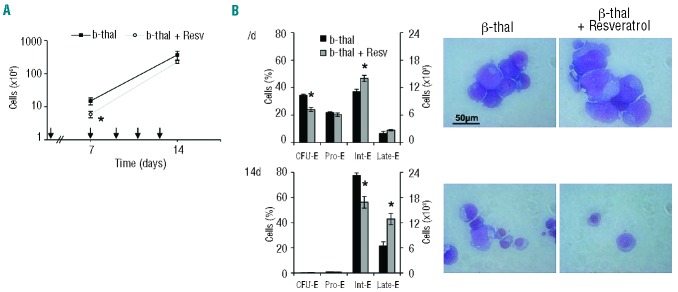
β-thalassemic erythropoiesis is affected by low-dose resveratrol, which induces early erythroid maturation. (A) Cell proliferation of β-thalassemic (b-thal) erythroid precursors derived by in vitro liquid culture of CD34+ cells isolated from peripheral blood of β-thalassemic (b-thal) subjects with or without resveratrol (Resv) (n=10). Arrows indicated when resveratrol 5 μM was added to the culture medium. Data are presented as means± SD; *P<0.05 compared to untreated β-thalassemic cells. (B) Cytofluorimetric analysis of maturation pattern of β-thalassemic erythroid precursors at different times of cell culture, 7, and 14 days (d), using the following surface markers: CD36, glycophorin-A and CD71(23) (Online Supplementary Appendix). This cytofluorimetric strategy allows the identification of the following homogenous cell populations: pro-erythroblasts (Pro-E), basophilic erythroblasts corresponding to intermediate erythroblasts (Int-E) and polychromatic and orthochromatic erythroblasts as late erythroblasts (Late E). Data are expressed as percentages or as absolute cell counts and shown as means ± SD (n=10); *P< 0.05 compared to untreated β-thalassemic cells. (Right) morphology of β-thalassemic (b-thal) erythroid precursors with or without resveratrol (Resv). Cytospins were stained with May-Grunwald-Giemsa. Cells were imaged under oil at 100× magnification using a Panfluor objective with 1.30 numeric aperture on a Nikon Eclipse DS-5M camera and processed with Digital Slide (DS-L1) Nikon. One representative image of the other 10 for each condition at the different time points is shown.
Resveratrol significantly increased FOXO3 expression in β-thal CFU-E cells compared to untreated β-thalassemic cells with no effects on FOXO1 mRNA levels (Online Supplementary Figure S4A). Since the number of cells from β-thalassemic erythroid precursors is limited due to the ineffective erythropoiesis and the effect of resveratrol on FoxO3a that has been shown in healthy cells by immunofluorescence and immunoblot analysis, we studied FoxO3a localization by immunofluorescence on sorted β-thalassemic CFU-E cells. We found that FoxO3a was similarly localized in the nucleus in both treated and untreated cells, but that FoxO3 nuclear expression was increased at 11 days of culture in resveratrol treated β-thalassemic CFU-E cells compared to untreated cells (Figure 4A). We confirmed the reduced activation of Akt in resveratrol treated β-thalassemic CFU-E similar to that observed in control CFU-Es, validating a role for resveratrol on the Akt signaling pathway (Figure 4B). In β-thalassemic CFU-E, we noted upregulation of catalase through FoxO3a activation (Online Supplementary Figure S4B). Thus, resveratrol induces erythroid differentiation by inhibiting Akt and enhancing FoxO3 activity in normal and disordered erythropoiesis, suggesting a potential role for resveratrol in reducing ineffective erythropoiesis in β-thalassemia by upregulation of the anti-oxidant gene as catalase.
Figure 4.

β-thalassemic sorted CFU-E from cells cultured with low dose resveratrol results in the upregulation and activation of FoxO3a and inhibition of Akt. (A) FoxO3 immunostaining of β-thalassemic CFU-E cells at Days 7 (7d) and 11 (11d) of culture with and without resveratrol. Cells are FACS-sorted, cytospun onto glass slides, and immunostained with anti-FoxO3a antibody and counterstained with DAPI. (Right panel). The mean fluorescence was measured in the nucleus of 30 cells using Image J software. Data are presented as means ±SD; *P<0.05 compared to untreated cells (n=5). (B) Western-blot (Wb) analysis of phospho-Akt (p-Akt) and Akt in sorted β-thalassemic CFU-E cells at Days 7 and of culture with resveratrol (Resv) or without (control, C). Tubulin was used as protein loading control. One representative gel from the other 6 with similar results is presented. (Right panel). Relative quantification of immunoreactivity of phospho-Akt (p-Akt), Akt and tubulin in sorted CFU-E cells. Data are presented as p-Akt/tubulin or Akt/tubulin ratio and shown as means ±SD (n=6). *P<0.05 compared to untreated cells.
Resveratrol decreases ineffective erythropoiesis and anemia in a in vivo mouse model of β-thalassemia
To evaluate the impact of resveratrol treatment in vivo, we used β-thalassemic mice (Hbbth3/+), a model which resembles in severity human β-thalassemia intermedia. In resveratrol treated β-thalassemic mice, we observed significant increases in hematocrit, hemoglobin levels, MCV and MCH, which were associated with a significant decrease in reticulocyte count (Table 1) and a decrease in total bilirubin (WT 0.15±0.04 vs. resveratrol 0.2±0.06 n=6; Hbbth3/+ 0.7±0.1 vs. resveratrol 0.2±0.01 mg/dL; n=6 P<0.05). β-thalassemic mice treated with resveratrol exhibited a significant reduction in spleen size, a sign of extramedullar erythropoiesis (Online Supplementary Figure S5A). Erythroid precursors were evaluated by flow cytometric analysis using the specific markers CD44 and TER119 (Online Supplementary Figure S5B).33 In the BM of resveratrol-treated wild-type mice, we observed a decrease in polychromatic-erythroblasts (pop. III) and a decrease in orthochromatic-erythroblasts compared to untreated wild-type mice (pop. IV; Figure 5A and Online Supplementary Figure S5C). In the BM of β-thalassemic mice, resveratrol induced a decrease in basophilic erythroblasts (pop. II) and an increase in orthochromatic erythroblasts (pop. IV; Figure 5A and Online Supplementary Figure S5C) suggesting that resveratrol decreases the extent of ineffective erythropoiesis of β-thalassemic mice.
Table 1.
Effect of resveratrol on hematologic parameters and red cell index in β thalassemic (hbb3th/+) mice.
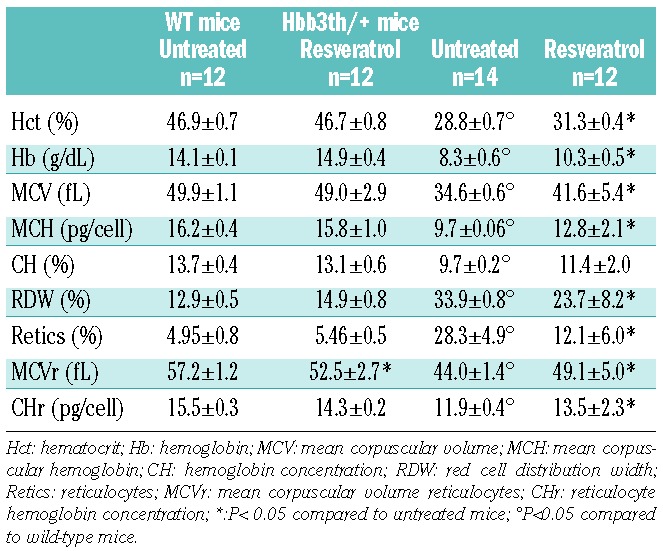
Figure 5.

In vivo supplementation with resveratrol ameliorates β-thalassemic ineffective erythropoiesis, up-regulates Foxo3 and peroxiredoxin-2 (Prdx2). (A) Cytofluorimetric analysis of maturation pattern of wild-type (wt) and β-thalassemic (b-thal) erythroid precursors from the bone marrow of mice with or without resveratrol supplementation using the following surface markers: CD44 and TER119 (Online Supplementary Appendix and Figure S5B). This cytofluorimetric strategy allows the identification of the following homogenous cell populations: population I corresponding to pro-erythroblasts, population II corresponding to basophilic erythroblasts, population III corresponding to polychromatic erythroblasts and population IV corresponding to orthochromatic erythroblsts. Data presented as means ± SD (absolute cell counts are shown in Online Supplementary Figure S5C) (n=10); *P<0.05 compared to untreated mice. (B) (Left): RT-PCR expression of Foxo3 and peroxiredoxin-2 (Prdx2) on sorted basophilic erythroblasts from bone marrow of mice with or without resveratrol (Resv) supplementation. Sorted basophilic erythroblasts and polychromatic erythroblasts from 10 different mice from each mouse group were analyzed. Experiments were performed in triplicate. Error bars represent standard deviations (mean ± SD; *P<0.05 compared to untreated mice, n=10). (Right): RT-PCR expression of peroxiredoxin-2 (Prdx2) on sorted polychromatic erythroblasts from bone marrow of mice with or without resveratrol (Resv) supplementation. Sorted polychromatic erythroblasts from 10 different mice from each group were analyzed. Experiments were performed in triplicate. Error bars represent the standard deviations (mean ± SD; *P<0.05 compared to untreated mice; n=10). (D) Immunoblot analysis of PRDX2 in sorted basophilic and polychromatic erythroblasts of wild-type (wt) and β-thalassemic mice with or without resveratrol (Resv) supplementation. Band 3 was used as loading control. One representative experiment of 4 others with similar results. (Right): relative quantification of immunoreactivity of peroxiredoxin-2 (Prdx2) and band 3 in sorted polychromatic erythroblasts of wild-type (wt) and β-thalassemic (b-thal) mice with or without resveratrol (Resv) supplementation. Data expressed as Prdx2/band 3 ratio and presented as means ±SD (n=5); *P<0.05 compared to untreated mice.
Resveratrol sustains in vivo β-thalassemic erythropoiesis by up-regulating foxo3 and peroxiredoxin-2
In sorted basophilic erythroblasts as seen with human cells in vitro Foxo3 expression was increased in both control and β-thalassemic mice treated with resveratrol (Figure 5B). In sorted basophilic- and polychromatic-erythroblasts we found upregulation of catalase (Cat) in both mouse strains supplemented with resveratrol similarly to that observed in cultured human cells (Online Supplementary Figure S5D). Among the scavenging enzymes with cytoprotective function, we have recently reported that peroxiredoxin-2 (Prdx2) plays a crucial role in β-thalassemic red cells and proposed a role of Prdx2 in erythropoiesis.30,35 Upregulation of both Foxo3 and Prdx2 have been recently described in Drosophila neuronal cells and in mammalian cell lines exposed to oxidative stress.43,44 By analyzing nucleotide sequences of Prdx2 and Foxo DNA binding motifs, we found a conserved FOXO binding site in the core promoter region (−15 to −8 bp) of Prdx2. We then evaluated Prdx2 expression in sorted basophilic erythroblasts and polychromatic erythroblasts and found upregulation of Prdx2 in basophilic erythroblasts from wild-type mice and in polychromatic erythroblasts in both mouse groups treated with resveratrol (Figure 5B). The immunoblot analysis of Prdx2 in sorted polychromatic erythroblasts confirmed the upregulation of Prdx2 in resveratrol treated mice (Figure 5C). These data suggest that Foxo3a might promote Prdx2 transcription.
Resveratrol improves red cell survival and decreases red cell membrane oxidative damage in β-thalassemic mice
Resveratrol treatment resulted in significant hematologic changes in β-thalassemic mice (Table 1). Hct and Hb levels increased significantly along with increased MCV and MCH, while the reticulocyte count and total bilirubin significantly decreased. In resveratrol-treated β-thalassemic mice, red cell morphology ameliorated (Figure 6A) and erythrocyte survival studies showed a significant improvement, whereas no significant changes were noted in treated wild-type mice (Figure 6B). We also evaluated the extent of red cell membrane oxidative damage by quantifying carbonyl groups present on red cell membrane. At baseline, markedly increased membrane oxidant damage was noted in β-thalassemic mouse red cells compared to wild-type red cells (Figure 6C).18,30,45 Resveratrol supplementation significantly reduced carbonyl groups in the red cell membrane of β-thalassemic mice compared to untreated β-thalassemic mice (Figure 6C). No differences were noted between red cells from untreated and treated wild-type mice (Figure 6C).
Figure 6.
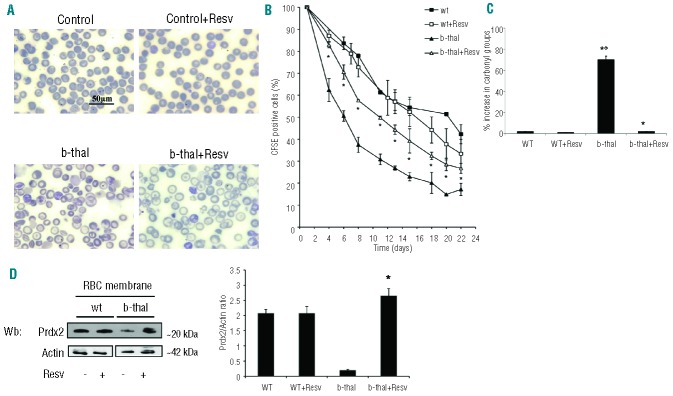
In vivo supplementation with resveratrol ameliorates red cell morphology, increases red cell lifespan and reduces red cell membrane oxidative damage in β-thalassemic mice. (A) Morphology of red cells from wild-type (wt) and β-thalassemic (b-thal) mice with or without resveratrol supplementation. Cells were stained with May-Grunwald-Giemsa. Cells were imaged under oil at 100× magnification using a Panfluor objective with 1.30 numeric aperture on a Nikon Eclipse DS-5M camera and processed with Digital Slide (DS-L1) Nikon. We show one representative image from a total of 12 for each condition. (B) Red cell survival (see also Methods) of CFSE labeled red cells from wild-type (wt) and β-thalassemic (b-thal) mice with or without resveratrol (Resv) supplementation. Data presented as means ± SD (n=4) from each mouse group; *P<0.05 compared to untreated mice. (C) Percentage of carbonyl groups from red cell membranes from wild-type (wt) and β-thalassemic (b-thal) mice with or without resveratrol (Resv) supplementation. Data are presented as means ± SD (n= 6) from each group; *P<0.05 compared to untreated mice; °P<0.05 compared to wild-type mice. (D) Peroxiredoxin-2 (Prdx2) membrane association in wild-type (wt) and β-thalassemic (b-thal) mice with or without resveratrol (Resv) supplementation. Actin was used as loading control protein. (Right): relative quantification of immunoreactivity of peroxiredoxin-2 (Prdx2) and actin in red cell membrane from wild-type (wt) and β-thalassemic (b-thal) mice with or without resveratrol (Resv) supplementation. Data are presented as means ±SD (n=6); *P<0.05 compared to untreated mice.
Since we previously showed that Prdx2 membrane association in β-thalassemic red cell is reduced, we evaluated the Prdx2 membrane binding in red cells from resveratrol-treated mice. The amount of Prdx2 associated with the membrane was significantly increased in β-thalassemic mouse red cells (Figure 6D) suggesting that resveratrol increases red cell lifespan in association with a decrease in red cell membrane oxidant damage in β-thalassemic mice, which in combination with beneficial effects on ineffective erythropoiesis, reduces the severity of anemia in murine β-thalassemia.
Discussion
We show that low-dose resveratrol induces early maturation of normal erythroid precursors by activation of the FoxO3a transcriptional factor, inhibition of Akt and upregulation of antioxidant response genes such as catalase. The effects of resveratrol on cell maturation are highly dependent on resveratrol concentration and on cell types.1,5–8 The findings from previous studies are not directly applicable to ours due to either the use of erythroid leukemia cell lines or study of primary erythroid cells without detailed characterization of their stage of cell differentiation.5–7 Here, we observed that CFU-E cells are the most susceptible erythroid cell population to the effects of low dose resveratrol (Figures 1 and 2). Since resveratrol has no effect on the expression of erythropoietin receptors during erythroid differentiation,8 we propose that resveratrol might hamper cell proliferation and induce cell maturation as supported by the early expression of GPA and band 3 in resveratrol-treated cells and by similar observations in other cell models.7,22 We recently showed that resveratrol targets FoxO3 a key transcriptional factor in erythropoiesis involved in upregulation of scavenging enzymes.4,12 We explored the possibility of resveratrol playing a pivotal role as an exogenous anti-oxidant agent and as a modulator of endogenous anti-oxidant systems. In resveratrol-treated CFU-E, FoxO3 was up-regulated and became predominantly localized in the nucleus, suggesting an effect of resveratrol on FoxO3a activation. This is also supported by the resveratrol inhibition of Akt, which regulates FoxO3a translocation to the nucleus.12,13 The resveratrol-induced FoxO3a activation results in upregulation of antioxidant response genes such as catalase which protects erythroid cells from the cellular stress associated with accelerated erythroid maturation. We hypothesized that these effects of resveratrol might be relevant in disordered erythropoiesis as seen in β-thalassemia that is characterized by ineffective erythropoiesis and oxidative stress. Previous studies have evaluated the impact of different anti-oxidant molecules on anemia of β-thalassemia, but only a few of them have been shown to ameliorate β-thalassemic ineffective erythropoiesis.16,21,46 In resveratrol-treated β-thalassemic erythroid precursors, we once again observed a reduction of CFU-E cells with an increase in Int-Es (Figure 3B), suggesting that resveratrol accelerates β-thalassemic cell maturation through the same mechanism found in normal erythroid precursors and sustains β-thalassemic erythropoiesis by upregulation of anti-oxidant response systems such as catalase (Online Supplementary Figure S4B). We, therefore, carried out in vivo studies using a murine β-thalassemic model to determine whether resveratrol administration could modify in vivo β-thalassemic anemia. In β-thalassemic mice, resveratrol decreased ineffective erythropoiesis seen by the reduction in spleen size and normalization of the pattern of β-thalassemic mouse erythroid differentiation. Interestingly, similar results on ineffective erythropoiesis were also obtained in the same β-thalassemic mouse model treated with Jak2 inhibitor.42 Although resveratrol has been described as not affecting the EPO pathway in normal erythroid precursors,8 we cannot exclude a possible effect of resveratrol on the up-regulated Jak2 pathway in β-thalassemic erythroid precursors.42
The beneficial effects of resveratrol on the β-thalassemic hematologic phenotype was also supported by amelioration of red cell morphology, increased red cell survival, decreased anemia as reflected by increased hemoglobin values with reduced reticulocytosis and decreased levels of total bilirubin, a marker of hemolysis. The effect of resveratrol on mature red cells is supported by the reduction in the extent of red cell membrane oxidative damage and the increased membrane association of Prdx2.1,2,16,47
The in vivo studies confirmed our findings from the in vitro model: resveratrol favored cell maturation and upregulation of FOxO3a in sorted basophilic erythroblasts. We found that catalase and Prdx2 are also up-regulated in this erythroid population in wild-type mice and in polychromatic erythroblasts from both mouse models. This suggests that Prdx2 might be an additional scavenging enzyme whose expression may be induced by FoxO3a as supported by the presence of a FOXO consensus binding site in the promoter region of Prdx2. Thus, resveratrol may confer resistance to oxidative stress in β-thalassemic erythropoiesis and thereby reduce the extent of ineffective erythropoiesis of β-thalassemia. The dose of resveratrol chosen for the in vivo mouse β-thalassemic study had a resveratrol plasma concentration of 2.6 μM; close to that obtained in human subjects treated with 25 mg resveratrol.26–28 Since resveratrol bioavailability is similar in rodents and humans, our study suggests that this polyphenol might be considered a new possible complementary tool in the treatment of anemia of β-thalassemia given its dual role through a novel mechanism by promoting terminal erythroid differentiation with activation of FoxO3a and the upregulation of anti-oxidant systems as catalase and Prdx2. In addition, by clarifying the FoxO3a signaling pathways affected by resveratrol in β-thalassemia, we identified novel opportunities for targeted pharmacological interventions with compound others than resveratrol such as specific FoxO3 activators.4
Supplementary Material
Footnotes
The online version of this article has a Supplementary Appendix.
Funding
This work was supported by grants from FUR2010-2011, University of Verona, Italy, to MB and LDF, PRIN 2008 to LDF and AI and NIH/NIA grants and support from CONACYT to SM and Glenn Foundation for Medical Research to DS.
Authorship and Disclosures
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Bauer JH, Helfand SL. New tricks of an old molecule: lifespan regulation by p53. Aging cell. 2006;5(5):437–40 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Armour SM, Baur JA, Hsieh SN, Land-Bracha A, Thomas SM, Sinclair DA. Inhibition of mammalian S6 kinase by resveratrol suppresses autophagy. Aging. 2009;1(6):515–28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Cucciolla V, Borriello A, Oliva A, Galletti P, Zappia V, Della Ragione F. Resveratrol: from basic science to the clinic. Cell Cycle. 2007; 6(20):2495–510 [DOI] [PubMed] [Google Scholar]
- 4.Hubbard BP, Gomes AP, Dai H, Li J, Case AW, Considine T, et al. Evidence for a common mechanism of SIRT1 regulation by allosteric activators. Science. 2013;339(6124):1216–9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ferry-Dumazet H, Garnier O, Mamani-Matsuda M, Vercauteren J, Belloc F, Billiard C, et al. Resveratrol inhibits the growth and induces the apoptosis of both normal and leukemic hematopoietic cells. Carcinogenesis. 2002;23(8):1327–33 [DOI] [PubMed] [Google Scholar]
- 6.Rodrigue CM, Arous N, Bachir D, Smith-Ravin J, Romeo PH, Galacteros F, et al. Resveratrol, a natural dietary phytoalexin, possesses similar properties to hydroxyurea towards erythroid differentiation. Br J Haematol. 2001;113(2):500–7 [DOI] [PubMed] [Google Scholar]
- 7.Ragione FD, Cucciolla V, Criniti V, Indaco S, Borriello A, Zappia V. p21Cip1 gene expression is modulated by Egr1: a novel regulatory mechanism involved in the resveratrol antiproliferative effect. J Biol Chem. 2003;278 (26):23360–8 [DOI] [PubMed] [Google Scholar]
- 8.Jeong JY, Silver M, Parnes A, Nikiforow S, Berliner N, Vanasse GJ. Resveratrol ameliorates TNFalpha-mediated suppression of erythropoiesis in human CD34(+) cells via modulation of NF-kappaB signalling. Br J Haematol. 2011;155(1):93–101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Fibach E, Prus E, Bianchi N, Zuccato C, Breveglieri G, Salvatori F, et al. Resveratrol: Antioxidant activity and induction of fetal hemoglobin in erythroid cells from normal donors and beta-thalassemia patients. Int J Mol Med. 2012;29(6):974–82 [DOI] [PubMed] [Google Scholar]
- 10.Zhang QS, Marquez-Loza L, Eaton L, Duncan AW, Goldman DC, Anur P, et al. Fancd2−/− mice have hematopoietic defects that can be partially corrected by resveratrol. Blood. 2010;116(24):5140–8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.De Franceschi L, Bertoldi M, De Falco L, Santos Franco S, Ronzoni L, Turrini F, et al. Oxidative stress modulates heme synthesis and induces peroxiredoxin-2 as a novel cytoprotective response in beta-thalassemic erythropoiesis. Haematologica. 2011;96(11): 1595–604 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Marinkovic D, Zhang X, Yalcin S, Luciano JP, Brugnara C, Huber T, et al. Foxo3 is required for the regulation of oxidative stress in erythropoiesis. J Clin Invest. 2007;117(8):2133–44 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ghaffari S, Kitidis C, Zhao W, Marinkovic D, Fleming MD, Luo B, et al. AKT induces erythroid-cell maturation of JAK2-deficient fetal liver progenitor cells and is required for Epo regulation of erythroid-cell differentiation. Blood. 2006;107(5):1888–91 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo P, Hu LS, et al. Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell. 1999;96(6):857–68 [DOI] [PubMed] [Google Scholar]
- 15.Rund D, Rachmilewitz E. Beta-thalassemia. N Engl J Med. 2005;353(11):1135–46 [DOI] [PubMed] [Google Scholar]
- 16.de Franceschi L, Turrini F, Honczarenko M, Ayi K, Rivera A, Fleming MD, et al. In vivo reduction of erythrocyte oxidant stress in a murine model of beta-thalassemia. Haematologica. 2004;89(11):1287–98 [PubMed] [Google Scholar]
- 17.De Franceschi L, Ronzoni L, Cappellini MD, Cimmino F, Siciliano A, Alper SL, et al. K-CL co-transport plays an important role in normal and beta thalassemic erythropoiesis. Haematologica. 2007;92(10):1319–26 [DOI] [PubMed] [Google Scholar]
- 18.Olivieri O, De Franceschi L, Capellini MD, Girelli D, Corrocher R, Brugnara C. Oxidative damage and erythrocyte membrane transport abnormalities in thalassemias. Blood. 1994; 84(1):315–20 [PubMed] [Google Scholar]
- 19.Ginzburg Y, Rivella S. β-thalassemia: a model for elucidating the dynamic regulation of ineffective erythropoiesis and iron metabolism. Blood. 2011;118(16):4321–30 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Liu J, Zhang J, Ginzburg Y, Li H, Xue F, De Franceschi L, et al. Quantitative analysis of murine terminal erythroid differentiation in vivo: novel method to study normal and disordered erythropoiesis. Blood. 2013;121(8): e43–9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Fibach E, Rachmilewitz E. The role of oxidative stress in hemolytic anemia. Curr Mol Med. 2008;8(7):609–19 [DOI] [PubMed] [Google Scholar]
- 22.Ragione FD, Cucciolla V, Borriello A, Pietra VD, Racioppi L, Soldati G, et al. Resveratrol arrests the cell division cycle at S/G2 phase transition. Bioch Biophys Res Com. 1998;250 (1):53–8 [DOI] [PubMed] [Google Scholar]
- 23.Merryweather-Clarke AT, Atzberger A, Soneji S, Gray N, Clark K, Waugh C, et al. Global gene expression analysis of human erythroid progenitors. Blood. 2011;117(13): e96–108 [DOI] [PubMed] [Google Scholar]
- 24.Capasso M, Avvisati RA, Piscopo C, Laforgia N, Raimondi F, de Angelis F, et al. Cytokine gene polymorphisms in Italian preterm infants: association between interleukin-10 -1082 G/A polymorphism and respiratory distress syndrome. Ped Res. 2007;61(3):313–7 [DOI] [PubMed] [Google Scholar]
- 25.Montesano Gesualdi N, Chirico G, Pirozzi G, Costantino E, Landriscina M, Esposito F. Tumor necrosis factor-associated protein 1 (TRAP-1) protects cells from oxidative stress and apoptosis. Stress. 2007;10(4):342–50 [DOI] [PubMed] [Google Scholar]
- 26.Walle T, Hsieh F, DeLegge MH, Oatis JE, Jr, Walle UK. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab Dispos. 2004;32(12):1377–82 [DOI] [PubMed] [Google Scholar]
- 27.Baur JA, Sinclair DA. Therapeutic potential of resveratrol: the in vivo evidence. Nat Rev Drug Discov. 2006;5(6):493–506 [DOI] [PubMed] [Google Scholar]
- 28.Soleas GJ, Angelini M, Grass L, Diamandis EP, Goldberg DM. Absorption of trans-resveratrol in rats. Methods Enzymol. 2001;335:145–54 [DOI] [PubMed] [Google Scholar]
- 29.Matte A, Bertoldi M, Mohandas N, An X, Bugatti A, Brunati AM, et al. Membrane association of peroxiredoxin-2 in red cells is mediated by the N-terminal cytoplasmic domain of band 3. Free Radic Biol Med. 2013;55:27–35 [DOI] [PubMed] [Google Scholar]
- 30.Matte A, Low PS, Turrini F, Bertoldi M, Campanella ME, Spano D, et al. Peroxiredoxin-2 expression is increased in beta-thalassemic mouse red cells but is displaced from the membrane as a marker of oxidative stress. Free Radic Biol Med. 2010; 49(3):457–66 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.De Franceschi L, Brugnara C, Beuzard Y. Dietary magnesium supplementation ameliorates anemia in a mouse model of beta-thalassemia. Blood. 1997;90(3):1283–90 [PubMed] [Google Scholar]
- 32.De Franceschi L, Daraio F, Filippini A, Carturan S, Muchitsch EM, Roetto A, et al. Liver expression of hepcidin and other iron genes in two mouse models of beta-thalassemia. Haematologica. 2006;91(10):1336–42 [PubMed] [Google Scholar]
- 33.Chen K, Liu J, Heck S, Chasis JA, An X, Mohandas N. Resolving the distinct stages in erythroid differentiation based on dynamic changes in membrane protein expression during erythropoiesis. PNAS. 2009;106(41): 17413–8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Foller M, Sopjani M, Koka S, Gu S, Mahmud H, Wang K, et al. Regulation of erythrocyte survival by AMP-activated protein kinase. FASEB J. 2009;23(4):1072–80 [DOI] [PubMed] [Google Scholar]
- 35.De Franceschi L, Tomelleri C, Matte A, Brunati AM, Bovee-Geurts PH, Bertoldi M, et al. Erythrocyte membrane changes of chorea-acanthocytosis are the result of altered Lyn kinase activity. Blood. 2011;118 (20):5652–63 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Fields R, Dixon HB. Micro method for determination of reactive carbonyl groups in proteins and peptides, using 2,4-dinitrophenyl-hydrazine. Biochem J. 1971;121(4):587–9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Roy SK, Chen Q, Fu J, Shankar S, Srivastava RK. Resveratrol inhibits growth of orthotopic pancreatic tumors through activation of FOXO transcription factors. PloS one. 2011; 6(9):e25166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Seoane J, Le HV, Shen L, Anderson SA, Massague J. Integration of Smad and forkhead pathways in the control of neuroepithelial and glioblastoma cell proliferation. Cell. 2004;117(2):211–23 [DOI] [PubMed] [Google Scholar]
- 39.Nakamura N, Ramaswamy S, Vazquez F, Signoretti S, Loda M, Sellers WR. Forkhead transcription factors are critical effectors of cell death and cell cycle arrest downstream of PTEN. Mol Cell Biol. 2000;20(23):8969–82 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Van Der Heide LP, Hoekman MF, Smidt MP. The ins and outs of FoxO shuttling: mechanisms of FoxO translocation and transcriptional regulation. Biochem J. 2004;380(Pt 2):297–309 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Mathias LA, Fisher TC, Zeng L, Meiselman HJ, Weinberg KI, Hiti AL, et al. Ineffective erythropoiesis in beta-thalassemia major is due to apoptosis at the polychromatophilic normoblast stage. Exp Hematol. 2000;28(12): 1343–53 [DOI] [PubMed] [Google Scholar]
- 42.Libani IV, Guy EC, Melchiori L, Schiro R, Ramos P, Breda L, et al. Decreased differentiation of erythroid cells exacerbates ineffective erythropoiesis in beta-thalassemia. Blood. 2008;112(3):875–85 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Miyamoto N, Izumi H, Miyamoto R, Kubota T, Tawara A, Sasaguri Y, et al. Nipradilol and timolol induce Foxo3a and peroxiredoxin 2 expression and protect trabecular meshwork cells from oxidative stress. Invest Ophthal Mol Vis Sci. 2009;50(6):2777–84 [DOI] [PubMed] [Google Scholar]
- 44.Lee KS, Iijima-Ando K, Iijima K, Lee WJ, Lee JH, Yu K, et al. JNK/FOXO-mediated neuronal expression of fly homologue of peroxiredoxin II reduces oxidative stress and extends life span. J Biol Chem. 2009;284 (43):29454–61 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.de Franceschi L, Shalev O, Piga A, Collell M, Olivieri O, Corrocher R, et al. Deferiprone therapy in homozygous human beta-thalassemia removes erythrocyte membrane free iron and reduces KCl cotransport activity. J Lab Clin Med. 1999;133(1):64–9 [DOI] [PubMed] [Google Scholar]
- 46.Fibach E, Rachmilewitz EA. The role of antioxidants and iron chelators in the treatment of oxidative stress in thalassemia. Ann NY Acad Sci. 2010;1202:10–6 [DOI] [PubMed] [Google Scholar]
- 47.Vitrac X, Desmouliere A, Brouillaud B, Krisa S, Deffieux G, Barthe N, et al. Distribution of [14C]-trans-resveratrol, a cancer chemopreventive polyphenol, in mouse tissues after oral administration. Life Sci. 2003;72(20): 2219–33 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.