

Abstract
Purpose
To determine whether short-term pressure elevation affects complement gene expression in the retina in vitro and in vivo.
Methods
Muller cell (TR-MUL5) cultures and organotypic retinal cultures from adult mice and monkeys were subjected to either 24-h or 72-h of pressure at 0, 15, 30, and 45 mmHg above ambient. C57BL/6 mice were subjected to microbead-induced intraocular pressure (IOP) elevation for 7 days. RNA and protein were extracted and used for analysis of expression levels of complement component genes and complement component 1, q subcomponent (C1q) and complement factor H (CFH) immunoblotting.
Results
mRNA levels of complement genes and C1q protein levels in Muller cell cultures remained the same for all pressure levels after exposure for either 24 or 72 h. In primate and murine organotypic cultures, pressure elevation did not produce changes in complement gene expression or C1q and CFH protein levels at either the 24-h or 72-h time points. Pressure-related glial fibrillary acidic protein (GFAP) mRNA expression changes were detected in primate retinal organotypic cultures (analysis of variance [ANOVA]; p<0.05). mRNA expression of several other genes changed as a result of time in culture. Eyes subjected to microbead-induced IOP elevation had no differences in mRNA expression of complement genes and C1q protein levels (ANOVA; p>0.05 for both) with contralateral control and naïve control eyes.
Conclusions
Short-term elevation of pressure in vitro as well as short-term (1 week) IOP elevation in vivo does not seem to dramatically alter complement system gene expression in the retina. Prolonged expression to elevated pressure may be necessary to affect the complement system expression.
Introduction
High intraocular pressure (IOP) is an important risk factor for both the development and progression of glaucoma. In experimental animal models [1] as well as in humans [2] exposure to elevated IOP correlates with retinal ganglion cell (RGC) loss and optic nerve (ON) degeneration. In fact, lowering IOP is currently the only therapeutic means of slowing or halting the disease process.
Complement upregulation has been reported as an early event in the development of glaucoma. Studies in human glaucomatous tissue [3,4] and experimental animal models [5-7] have shown that complement system components are upregulated in the retina in both early and late stages of the disease.
The complement system is part of the innate immune system. Its functions are to opsonize antigens, recruit macrophages, and cause the lysis of bacterial pathogens by forming the terminal complement complex (TCC) [8,9]. The complement system comprises several dozen proteins that are synthesized mainly in the liver and circulate in the blood stream; some of the complement components can also be found on cell membranes. The circulating complement proteins do not cross the blood–brain and the blood–retina barriers because of their large size. To provide local protection, they are synthesized locally within the central nervous system (CNS). Several of the complement proteins, including complement component 1, q subcomponent (C1q) and complement component 3 (C3), have been shown to be expressed in the retina under normal conditions, albeit at low levels [4,10].
Locally produced complement components in the CNS may have additional roles besides their direct immunological functions. Recent studies have suggested that complement may participate in normal developmental processes, such as pruning of synapses [11] and other developmental and degenerative processes [9,12], implicating it in diseases such as schizophrenia [13,14], Alzheimer disease [15], Parkinson disease [16], and glaucoma [17].
Given the central role of IOP elevation in glaucomatous pathophysiology and the role of the complement system in the same process, we examined whether short-term increases in pressure affect complement expression in an immortalized Muller cell line and organotypic retinal cultures in vitro. We also examined whether complement is affected by relatively short (1 week) IOP elevation in vivo in a microbead-induced model of ocular hypertension in mice.
Methods
Cell cultures
Conditionally immortalized cell line of Muller cells (TR-MUL5; originally described in [18]) were used and maintained in culture as previously described [19]. The cell line was kindly provided by Dr. Thomas Weber (Mount Sinai School of Medicine, New York, NY).
TR-MUL5 cells were kept at 34 °C in high glucose Dulbecco's Modified Eagle Medium (DMEM; Gibco, Invitrogen, Carlsbad, CA), 10% heat-inactivated fetal bovine serum (FBS; Gibco, Invitrogen), penicillin and streptomycin (Gibco, Invitrogen) in noncoated culture flasks. Cells at approximately 30% confluence were used for pressurization. Immediately before pressurization the cells were transferred to serum-free media and kept there for the duration of the experiment.
Animals
All procedures were performed in accordance with the Association for Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research. All animal protocols in the current study and previous studies described were approved by the Institutional Animal Care and Use Committee (IACUC) committee of SUNY Downstate Medical Center (New York, NY).
A group of 3- to 5-month-old C57BL/6 mice from the colony that we maintain (SUNY Downstate Medical Center) was used in the experiments. Mice were kept in a 12 h:12 h light–dark cycle and fed ad libitum.
Retinas from four normally reared young adult (6–9-years old) female bonnet macaque (Macaca radiata) monkeys were obtained immediately after sacrifice. These monkeys were part of the control (untreated) group of an unrelated experiment (to study whether antidepressants affect neurogenesis), which was approved by a separate IACUC protocol. All monkeys were housed in accordance with the guidelines of the IACUC in a social pen containing eight subjects. For the duration of 10 weeks before sacrifice, the animals were sedated with ketamine once a week to receive a placebo via a gastric tube. No other experimental manipulations were performed. All monkeys received food and water ad libitum for the duration of the study. Prior to sacrifice, monkeys were anesthetized to a surgical depth with sodium pentobarbital (12.5 mg/kg intravenously [IV]) with the goal of maintaining a deep sedation and minimizing discomfort. The depth of anesthesia was confirmed as indicated by the absence of pain reflexes, which was assessed by compressing the tail and orbital fissure, and the absence of deep tendon reflexes of the knee. The heart was exposed and a cannula inserted through an incision in the left ventricle into the ascending aorta and secured with a Babcock clamp. The descending aorta was clamped with a hemostat and the right atrium incised. Using a peristaltic pump, cold saline was infused for 10 min to clear blood vessels.
Retinal organotypic cultures
Mouse retinal organotypic cultures were prepared from 3−5-month-old (n=40) C57BL/6 mice as previously described [20]. Briefly, animals were anesthetized using ketamine (400 mg/kg) and xylazine (80 mg/kg), and after cervical dislocation the eyes were immediately enucleated. The retinas were removed under sterile conditions in ice-cold Hank’s balanced salt solution (HBSS; Fisher, Pittsburgh PA). Relaxing incisions were made to create flatmounts that were placed in cell culture inserts (see below).
Primate organotypic cultures were similarly prepared after sacrifice following transcardial perfusion with ice-cold saline solution for 10 min. Immediately following the perfusion, eyes were enucleated and quickly dissected under sterile conditions in ice-cold HBSS. Four punches were made in each retina with a disposable skin biopsy punch (3 mm), one from each quadrant at the center of the quadrant between the optic nerve head and the edge of the retina.
Explants were mounted on cell culture inserts (BD Biosciences, San Jose, CA) with an 8-μm pore size and cultured overnight in N-2 (Gibco, Invitrogen), B-27 (Gibco, Invitrogen), and L-glutamine (Gibco, Invitrogen)-supplemented neurobasal medium (Gibco, Invitrogen), at 34 °C in 24-well plates (BD Biosciences). The following day, fresh medium was added to the cultures immediately before pressurization. After pressurization, the retinal cultures were immersed in Trizol reagent (Invitrogen, Carlsbad, CA) for RNA and protein extraction.
Pressure elevation
The cells and organotypic retinal cultures were subjected to different levels of pressure in a set of four polycarbonate pressure chambers that were kept in an incubator at 34 °C for either 24 or 72 h. Pressure was adjusted to 15 (±3) mmHg, 30 (±3) mmHg, and 45 (±3) mmHg above ambient in three of the chambers, while the fourth chamber was maintained at ambient pressure. Pressure was regulated with a custom-made active feedback electronic system that injected a mixture of 95% air/5% CO2 until the desired pressure was reached and maintained it with short bursts of gas. Pressure and temperature was continuously monitored and recorded in each chamber for the duration of the experiment by an in-house built data logger; data were downloaded to a personal computer for evaluation. All pressure exposure experiments were performed at least in triplicate for cells and quadruplicate for organotypic retinal cultures at each pressure level. Figure 1 shows a typical graph of pressure and temperature during a 72-h experiment.
Figure 1.
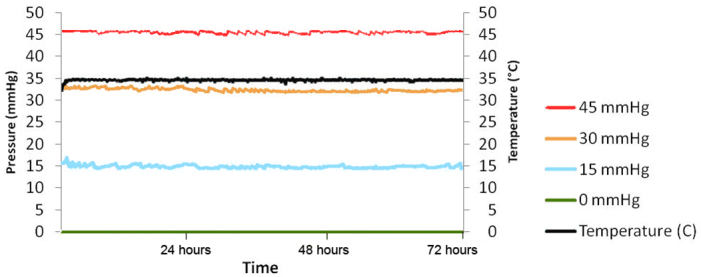
Typical pressure and temperature graph of a 72-h pressurization experiment. The four chambers were maintained at 0, 15, 33, and 46 mmHg while temperature was 34.5 °C.
Intraocular pressure elevation in vivo
IOP elevation was induced in one eye of sixteen 3-month-old male C57BL/6 mice by injection of 10-µm-diameter polystyrene beads (Invitrogen) as described previously [21]. Briefly, the animals were anesthetized as above, a small incision was made with a number 30G needle, and a pulled glass micropipette was used to inject 1–2 μl of the polystyrene beads into the anterior chamber (AC) of one eye. The contralateral eye remained uninjected and served as control. The animals were allowed to survive for 7 days, after which they were anesthetized as above, perfused intracardially with PBS (0.1 M, 137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4, pH 7.4), and the retinas were dissected out. In all animals baseline IOP was measured before treatment. After treatment, IOP was repeatedly measured and recorded every 2 days (totaling three times during the 7-day interval) using a prototype rebound tonometer as previously described [22]. Briefly, IOP was measured in awake, nonsedated mice that were restrained in a custom made restrainer that allows measurement of IOP without causing an increase in intrathoracic pressure. After mice were put in the restrainer, tetracaine 0.5% was applied to each eye prior to IOP measurements. Five measurements were taken from each eye and averaged; IOP was calculated using the tonometer calibration formula as previously described [22]. Only animals that demonstrated elevated IOP after the intraocular bead injection (n=10) were used in the experiments. Nine eyes from a separate group of untreated 3-month-old male mice (n=7) served as a naïve control group.
Cumulative IOP exposure was calculated as a sum of the products of a measured IOP value on a particular day and the number of days since the last IOP measurement. Peak IOP was defined as the highest IOP value of all measurements from one eye. Average IOP was calculated as the average of the three IOP values during the 7-day interval.
Real-time quantitative PCR
RNA was extracted from cells, retinal explants, and mouse retinas from sacrificed mice using Trizol reagent (Invitrogen) according to the manufacturer’s instructions. Cells, retinal explants, and mouse retinas from sacrificed mice were homogenized using a drive motor homogenizer Tissumizer SDT 181 (Tekmar, Cincinnati, OH) and total RNA was isolated using Trizol reagent (Invitrogen) according to the manufacturer’s instructions. The RNA was precipitated with isopropanol, washed in 70% ethanol, and column purified (RNAeasy mini kit; Qiagen, Valencia, CA). RNA quality was assessed by the 260/280 ratio using a Nanodrop ND-1000 spectrophotometer (Nanodrop Technologies; Wilmington, DE). It was then reverse transcribed to cDNA with a random primer as per the manufacturer’s instructions (Quantitect; Qiagen). 1000 ng of isolated total RNA was then reverse transcribed to cDNA using random hexamer primers following the manufacturer’s instructions (Quantitect; Qiagen). cDNA was processed for real-time quantitative PCR (RT-qPCR) with Power SYBR Green master mix (Applied Biosystems, Carlsbad, CA or Biobasic, Canada) and gene-specific primers. RT-qPCR was performed for the following mouse genes (gene name followed by GenBank accession number in parenthesis): complement component 1, sub component q (C1qbp; NM_007573.2), complement component 2 (C2; NM_013484.2), complement component 4B (C4b; NM_009780.2), complement factor H (CFH; NM_009888.3), thymus cell antigen 1, theta (Thy1; NM_009382.3), synuclein gamma (Sncg; NM_011430.2), and GFAP (NM_010277.3); RT-PCR was also performed for the following primate genes (gene name followed by GenBank accession number in parenthesis): C1qb (XM_001083650.2), CFH (XM_001111875.2), CD46 antigen, complement regulatory protein (CD46; AB172248.1), complement factor properdin (CFP; JU473400.1), complement regulator complement receptor 1-related gene/protein-y (CRRY; XM_002801945.1), decay accelerating factor for complement (DAF; XM_001112168.2), complement factor I (CFI; XM_001087512), serpin peptidase inhibitor, clade G (C1 inhibitor), member 1 (SERPING1; NC_007871.1), Thy1 (NM_001042638), and GFAP (XM_001102095) genes. All samples were run in triplicate and were analyzed on the Applied Biosystems 7900HT Fast Real-Time PCR System. Statistical analysis was performed using the delta-delta threshold cycle (Ct) method [23] after normalization using the mouse ribosomal protein S11 (RPS11) gene (GenBank accession number NM_013725.4) and primate RPS11 gene (GenBank accession number NM_001195754.1). Outliers were removed using Grubbs’ test [24].
Protein immunoblotting
Proteins were extracted from the organic phase of Trizol by dialyzing against three changes of 0.1% sodium dodecyl sulfate (SDS) at 4 °C for 48 h as previously described [25]. Protease inhibitors were added, and the samples were stored at −80 °C until used. Similarly, protein in the cell culture media was dialyzed as above and stored at −80 °C. Protein samples were concentrated using Amicon centrifugal filters (Millipore, Billerica, MA), diluted in Laemmli sample buffer (BioRad, Hercules, CA), separated in a 4%–20% SDS– polyacrylamide gel electrophoresis (PAGE) gel (Bio-Rad, Hercules, CA), and transferred to polyvinylidene fluoride (PVDF) membranes. The membranes were then blocked (Superblock; Thermo Fisher Scientific, Rockford, IL) and incubated with either a mouse monoclonal anti-C1q antibody (Abcam, Cambridge, MA; 1:2,000, overnight at 4 °C) or a goat polyclonal anti-CFH antibody (1:3,000, overnight at 4 °C; Quidel, catalog# A312; San Diego, CA), followed by incubation with a horseradish peroxidase-conjugated secondary antibody (Jackson Immunoresearch, West Grove, PA) for 2 h at room temperature. A rabbit anti-β-actin-specific antibody (1:5,000, overnight at 4 °C; Abcam) was used to detect β-actin, which served as a loading control.
Antibody binding was visualized with a chemiluminescence kit (ECL; Pierce, Rockford, IL) on an image station (Kodak 440CF; Kodak, Boston, MA) and quantified with ImageJ image-analysis software (developed by Wayne Rasband, National Institutes of Health, Bethesda, MD). For C1q, the sum of the normalized integrated density of all the bands was used for quantification.
Statistical analysis
The difference between Ct values of each gene under investigation and RPS11 (40S ribosomal protein S11) housekeeping gene was calculated (dCt) and normalized by subtraction of the Ct values of the samples in the 0 mmHg group (ddCt). Results were subjected to two-way analysis of variance (ANOVA) for treatment (pressure level) and length of incubation (24 or 72 h) for cultures using NCSS (Kaysville, UT). Fold changes were calculated (as 2–ddCt) for each replicate and then averaged among samples in each treatment group for graphing. For IOP experiments, t test or one-way ANOVA was used where appropriate. Significant differences (p<0.05) were further explored with Tukey–Kramer post hoc analysis.
Protein levels were normalized to the levels of β-actin and were subjected to statistical analysis using ANOVA. Tables were prepared using Microsoft Excel (Microsoft, Seattle, WA).
Results
Effect of elevated pressure on isolated retinal cell cultures
Expression levels of C1q, C2, C4, CFH, and CFI mRNA were not significantly different (ANOVA; p>0.05) in TR-MUL5 cells in cultures subjected to elevated pressure for either 24 or 72 h (data not shown). Levels of C3 mRNA were low and could not be reliably assessed in the samples analyzed.
Immunoblotting for C1q also did not detect any significant differences in the protein levels after 24 and 72 hours exposure to elevated pressure (ANOVA; p>0.05) in TR-MUL5 cell cultures (Figure 2).
Figure 2.
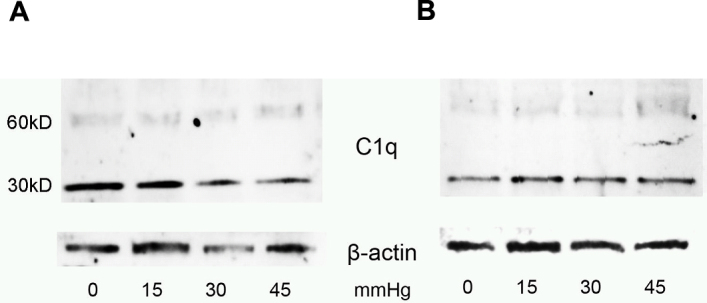
Extracellular complement component 1, q subcomponent (C1q) protein levels in Muller cell line TR-MUL5 cultures subjected to different pressure levels. Shown are representative immunoblots of extracellular C1q levels in TR-MUL5 cell cultures subjected to various levels of pressure (0, 15, 30, and 45 mmHg above ambient) for either (A) 24 or (B) 72 h.
Effect of elevated pressure on organotypic retinal cultures
No significant changes were detected in gross morphology or retinal cytoarchitecture after culturing either murine or primate retinal explants for 3 days. The effects of pressure on mRNA expression of various complement components in mouse and primate retinal organotypic cultures are presented in Figure 3 and Figure 4, respectively.
Figure 3.

Real-time reverse transcription polymerase chain reaction (RT–PCR) analysis of mouse retinal organotypic cultures subjected to different pressure levels. RT–PCR assays were performed to detect changes in mRNA expression levels in mouse retinal organotypic cultures subjected to different levels of pressure for 24 h (A) and 72 h (B). Fold changes are expressed in comparison to the average mRNA levels in cultures exposed to ambient atmospheric pressure (0 mmHg; thus the 0 mmHg group would have a mean of 1). Error bars represent standard error of the mean.
Figure 4.

Real-time reverse transcription polymerase chain reaction (RT–PCR) analysis of primate retinal organotypic cultures subjected to different pressure levels. RT–PCR assays were performed to detect changes in mRNA expression levels in primate retinal organotypic cultures subjected to different levels of pressure for 24 h (A) and 72 h (B). Fold changes are expressed in comparison to the average mRNA levels in cultures exposed to ambient atmospheric pressure (0 mmHg; thus the 0 mmHg group would have a mean of 1). Error bars represent standard error of the mean.
Levels of mRNA of complement genes did not differ across groups with different pressure exposure for either 24 h or 72 h of pressurization (two-way ANOVA comparison by pressure exposure; p>0.05) for both mouse and primate retinal explants. In addition, none of the other proteins studied showed any pressure effect in mouse cultures. Pressure caused a significant effect on GFAP mRNA expression (ANOVA; p=0.027) in primate retinal organotypic cultures after 24 h.
mRNA levels of Thy 1, GFAP, C2, and C4b were significantly affected by time in culture (but not by pressure exposure; two-way ANOVA; p<0.0002, 0.0057, 0.012, and 0.00006, respectively) in organotypic retinal cultures from C57BL/6 mice. At the same time, for primate retinas mRNA levels of Thy1, GFAP, SerpinG1, and CFI were also significantly affected by time in culture (but not by pressure exposure; two-way ANOVA; p<0.00001, p<0.005, p<0.00001, and p=0.028, respectively).
Protein levels of C1q (as determined by immunoblotting) remained unchanged (ANOVA; p>0.05) by exposure to various pressure levels after 24 and 72 h in both mouse (data not shown) and monkey (Figure 5A,B) organotypic retinal cultures. CFH immunoreactive protein also did not show any significant differences (ANOVA; p>0.05) between primate retinal organotypic cultures subjected to different pressure levels for 72 h (Figure 5C).
Figure 5.
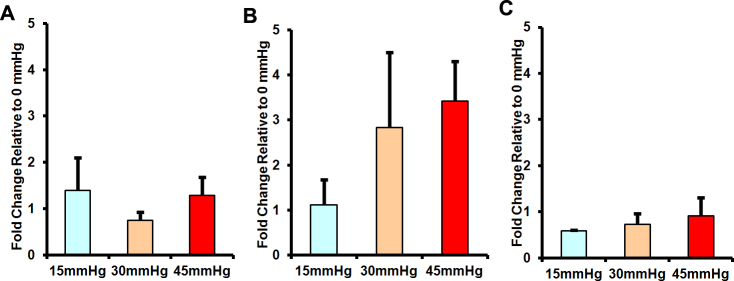
Complement component 1, q subcomponent (C1q) and complement factor H (CFH) protein levels in mouse and primate retinal organotypic cultures subjected to different pressure levels. Shown are quantifications of immunoblotting analysis of protein levels of C1q in primate organotypic retinal cultures subjected to different levels of pressure for either 24 (A) or 72 (B) hours. Quantification of immunoblotting analysis of CFH protein levels in primate retinal organotypic cultures subjected to different pressure levels for 72 h are presented in (C). Protein levels are normalized to the average level of the respective protein from cultures exposed to ambient atmospheric pressure (0 mmHg; thus the 0 mmHg group would have a mean of 1). Error bars represent standard error of the mean. (ANOVA; p>0.05).
Effect of elevated intraocular pressure elevation in vivo
Of the 16 mice injected with microbeads, ten demonstrated increased IOP during the time of the experiment. Only those mice were included in the final analysis. Mean (±standard error of the mean [SEM]) cumulative IOP exposure was 137.66±8.18 mmHg ×days for the eyes injected with microbeads, 106.84±5.42 mmHg*days for contralateral eyes of the same animals, and 108.39±3.88 mmHg*days for the eyes of the naïve mice.
Peak (mean±SEM) IOP was 25.12±2.54 mmHg for eyes injected with microbeads, 18.38±1.19 mmHg for contralateral eyes, and 15.48±0.55 mmHg for naïve eyes.
Average (mean±SEM) IOP of all mice was 18.72±1.56 mmHg for eyes injected with microbeads, 14.13±0.61 mmHg for contralateral eyes, and 15.48±0.55 mmHg for naïve eyes.
Cumulative IOP exposure for the eyes injected with microbeads was statistically significantly higher (ANOVA; p<0.007) compared to nontreated contralateral eyes and eyes from naïve animals (Tukey–Kramer post hoc analysis; p<0.05; Figure 6). Microbead-injected eyes also had statistically significantly higher peak IOP (ANOVA; p<0.013) and average IOP (ANOVA; p<0.025) values compared to the nontreated contralateral and naïve eyes.
Figure 6.
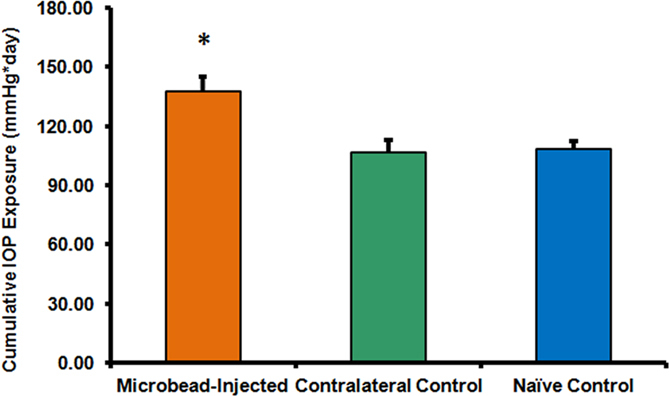
Microbead-injected eyes develop higher intraocular pressure (IOP). Eyes exposed to microbead injections in the anterior chamber developed significantly higher cumulative IOP (ANOVA; p<0.0065) compared to the contralateral eyes and eyes of the naive animals. Error bars represent standard error of the mean, *p<0.05.
mRNA expression levels of the genes studied were found to be similar in the retinas from the eyes subjected to increased IOP versus retinas from contralateral eyes (Figure 7A) when normalized to the mRNA levels from the retinas of naïve animals (t test; p>0.05).
Figure 7.
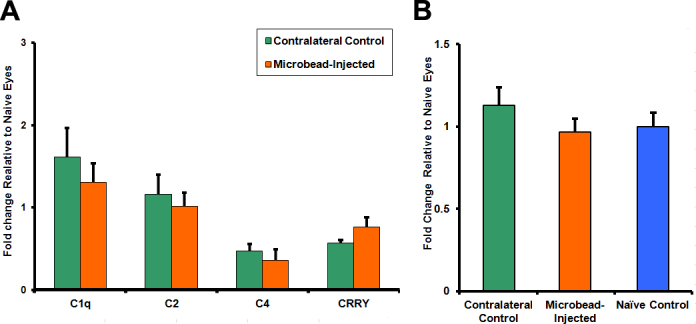
Effect of short-term microbead-induced intraocular pressure elevation on the complement system in the retina of C57BL/6 mice. Short-term (7 days) microbead-induced intraocular pressure (IOP) elevation did not produce changes in either complement component mRNA levels (A; t test; p>0.05) or complement component 1, q subcomponent (C1q) protein (B) expression levels (ANOVA; p>0.05) in the retinas of C57BL/6 mice. mRNA and C1q protein levels are normalized to the average level of the respective gene and C1q protein from retinas of naïve control mice (thus the naïve control group would have a mean of 1). Error bars represent standard error of the mean.
C1q immunoblotting did not reveal any differences in the levels of C1q protein (ANOVA; p>0.05) between retinas subjected to IOP elevation, contralateral control retinas of the same animals, or retinas of naïve mice (Figure 7B).
Discussion
Circulating complement has traditionally been thought of as an arm of the innate immune system, charged with opsonizing and, in some instances, lysing invading pathogens. More recently, it has emerged that complement has often immunomodulatory functions. It is therefore not surprising that complement appears to be involved in several systemic autoimmune conditions [26,27].
Up until the early 1990s, it was not well appreciated that CNS not only has the capability but actually produces most, if not all, complement components. Since that time, complement has been implicated in several neurodegenerative conditions [28,29]. At the same time, recent results have suggested that certain complement proteins may facilitate neuroprotection [30]. Similarly to the CNS, complement is also locally produced in the eye [5,31]. It has been postulated that locally produced complement plays a protective role in this immune-privileged organ [32]. However, in the past few years, several reports have linked complement with several degenerative eye conditions—most notably age-related macular degeneration [33] and glaucoma [4,5,34].
The involvement of complement in glaucomatous RGC loss and axonal degeneration is intriguing. Complement upregulation in the DBA/2J model of glaucoma occurs early, before the pathology becomes apparent, and continues even after significant RGC loss has occurred [4]. Similar observations were made in a rat glaucomatous model and in human glaucoma specimens [5]. Proteomic analysis in human tissue showed upregulation of both C1q and C3 but also the downregulation of CFH in human glaucomatous retina [3].
It has been suggested that complement might have different roles in glaucomatous pathology based on the stage of the disease [17]. Early in the disease, deposition of the complement could be mediating removal of RGC dendritic synapses [11], thus lowering RGC electrical activation and promoting their survival. As cell death becomes inevitable and some of the RGCs start undergoing apoptosis, complement might be promoting clearance of the cell debris and by doing so attenuate the inflammatory responses [17]. On the other hand, it was suggested that complement deposited on RGCs may not only target the compromised cells but also cause a bystander lysis of healthy RGCs and exacerbate the pathology [5]. Additionally, deposition of the complement may in fact activate adjacent glial cells and cause them to become reactive and start synthesizing various inflammatory mediators, including cytokines and superoxide radicals. Synthesis of these molecules by adjacent astroglial and microglial cells could further damage RGCs. In fact, glial cell activation has been recently demonstrated to be one of the pathological mechanisms that operate in glaucomatous disease [35,36].
It is clear that the role of the complement system in glaucoma is a complex one. It thus requires careful dissection of potential factors and mechanisms that could be responsible for its activation. Given the impact of IOP on glaucoma pathogenesis, we attempted to examine whether pressure elevation can affect the complement system.
Cell lines are a powerful tool for dissecting responses of individual components of a complex tissue to different experimental conditions. In recent years several retinal cell lines have been established, including a rat Muller cell line (TR-MUL5) [18,37] and a human Muller cell line (MIO-M1) [38], among others. Those lines have been extensively used to understand behavior of particular retinal cell types under various conditions. For example, Muller cell lines were used in studies that looked at the effects of excess extracellular glutamate [39] as well as of osmotic stress [40].
Recent studies have suggested that Muller cells are capable of producing complement system components and may regulate complement activation within the retina [41]. In addition, our earlier work [4] has suggested that Muller cells and astrocytes are responsible for complement production in the retina in glaucoma. In this study, we examined the effect of pressure elevation on Muller cells by using a TR-MUL5 cell culture. Similar models have been used in the past to identify molecular changes that occur in response to high pressure. For example, they have been used to show that elastin synthesis by glial cells is affected by high pressure [42] and that elevated pressure induces apoptosis [43] and oxidative stress in cultured RGCs [44].
We did not detect any significant changes in the levels of complement components, at least within the timeframe of this experiment and for the pressure levels studied. These pressure levels were selected to represent normal (15 mmHg), moderately high (30 mmHg), and very high (45 mmHg) pressure levels that can be encountered by retinal cells in vivo. Other studies that used such an ex vivo system have often looked at pressure levels that are outside the physiologic range [43-47]. Although the lack of effect of elevated pressure on complement expression by Muller cells can be interpreted as an inherent lack of responsiveness, other potential explanations could be that additional cells or factors are required, as has been reported for other cell types in the retina [48], or that immortalized cell lines have lost such a capacity. Yet another possible explanation is that a longer exposure to the elevated pressure is needed to induce detectable upregulation of complement components. Of course we cannot completely rule out small changes in complement system expression that were below the detection limit of our methodology.
In contrast to cell lines, organotypic cultures allow evaluation of responses of the entire tissue to different conditions under investigation. At the same time, they retain much of the flexibility and scalability of an in vitro system. Retinal organotypic cultures have been used to assess the regeneration potential of RGCs [49], the effects of stem cell transplantation [50], glutamate excitotoxicity [51], as well as the effects of drug treatment [52]. Recently, it has also been shown that organotypic retinal cultures can be used for gene transfer studies [53].
In this study we employed retinal organotypic cultures from mice and monkeys to assess the effects of pressure elevation. Experiments with murine explants allowed direct comparison of the results from some of the cell culture experiments with the rodent cell lines as well as with in vivo experiments in mice (see below). On the other hand, using primate tissue allowed us to address potential interspecies variability. It also allows us to relate the findings to human disease.
It has been shown that rodent organotypic retinal cultures can be maintained in culture up to 17 days without significant alterations in laminar architecture of the retina [20]. Other groups have observed that organotypic tissue cultures could be kept morphologically and structurally intact for at least 4 days [53]. In the current study we used a protocol that allows long survival of the tissue in culture [20]. We used the same protocol and media for the monkey retinal explants. Under the same conditions, organotypic cultures from donor postmortem human eyes have been maintained for up to 96 h with preservation of human retinal architecture and no notable decrease in thickness/density of the nuclear layers [54]. Inline with the above reports, we did not observe any morphological alterations in the retinal tissue for culture periods up to 72 h.
Neither murine nor primate explants exhibited changes in the expression of complement genes after exposure to elevated pressure for either 24 or 72 h. Similarly, the levels of C1q and CFH proteins remained unchanged over the range of pressures, further supporting the conclusion that short-term elevation of pressure does not affect locally synthesized complement in the retina.
It has been suggested that elevation of pressure does not cause stress to the cells resting on a rigid support to any appreciable degree; even pressure elevations of 50 mmHg are calculated to cause relative deformations of cellular components of less than 0.005% [55]. Nonetheless, several studies employing different cell lines resting on a rigid support that were subjected to different levels of pressure have documented various biologic responses to pressure elevation [43,56].
The discrepancy between the lack of substantial cell deformation and observed major biologic responses to pressure elevation has been puzzling. It has been recently argued [57] that most of the studies in which biologic response to pressure were observed have actually been documenting responses to differences in oxygen (and other gasses) tension that were not accounted for in the designs of those experiments. Use of a vertical column of medium to exert elevated pressure hydrostatically on the cells located at the bottom of the column causes the cells at the bottom to experience decreased oxygen (and increased CO2) tensions because of differences in diffusion of gases through variable amounts of fluid. If oxygen tension were controlled, astrocytes under different levels of pressure did not show significant differences in migration [57]. Another potential confounding factor is bottle-to-bottle differences in gas tension of the medium, which might also contribute to different oxygen (or CO2) tensions that the cells experience in the incubator atmosphere [57].
On the other hand, a recent study that looked at mechanosensitive channel TRPV1 in cultured RGC-5 cells using methodology that eliminates diffusion issues has offered a potential explanation of how pressure alone could trigger various biologic responses [58]. It was shown that activation of TRPV1 contributes to the apoptotic cell death of ganglion cells induced by pressure elevation [58]. The authors discussed several ways that elevated pressure could bring about activation of these channels and induce the downstream apoptotic pathways. In particular, cell membrane compression due to elevated pressure could disrupt the cytoskeletal scaffolding, and this, in turn, could affect the conductance of channels sensitive to mechanical tension.
The design of our experiment as well as utilization of an active pressurization and feedback system similar to the one used in this study [58] addresses some of the issues mentioned above when using columns of fluid to affect pressure hydrostatically. First, cell cultures and organotypic retinal cultures are maintained under ambient pressure in the incubator overnight before they are subjected to elevated pressure the following day. This allows equilibration with the incubator gases and minimizes potential variability of gas tensions in different bottles of medium. Second, an active pressurization system eliminates differences that could arise due to oxygen diffusion through the column of medium because under all conditions the column of the medium is the same (and is also negligible, only around 1–2 mm tall), whereas pressure elevation is achieved by a compressed gaseous mixture above the medium in the enclosed chamber.
According to Henry's law, the amount of gas dissolved in a liquid is directly proportional to the partial pressure of that gas in equilibrium with the liquid. As such, an increase in pressure of oxygen in the enclosed chamber in our system will lead to an increase of the partial pressure of oxygen in the culture medium. However, those effects are small as the difference between chambers represents only an approximately 5% variation from atmospheric pressure (atmospheric pressure can range up to 15% at sea level). Thus, it is most likely that the difference in gene and/or protein expression between cultures exposed to different pressure levels is the direct result of pressure itself.
Similarly, the issue of how IOP causes damage to the retina and optic nerve has not been resolved. IOP is the pressure inside the eye. As such, it applies a force that acts vertical to the retina and a tangential strain. As the retina is not a rigid structure, the pressure drop across the retina is minimal (as experiments involving simultaneous cannulation of the posterior choroid and vitreous have shown [59]). Although there is undoubtedly some strain applied to the retina with changing IOP, scleral expansion (which would allow for force transfer to the retina by making the surface that the retina occupies larger) is minimal (less that 5 µm or about 0.007% of the eye circumference). Thus, if there is an effect of IOP on the retina itself (rather than the optic nerve), this effect is likely directly related to the actual pressure that the retina is subjected to.
As such, it is interesting that primate organotypic retinal cultures showed a pressure-related change in expression of GFAP. GFAP is upregulated early in response to injury within the CNS [60] and retina [61,62] and can be thought of as a marker of tissue “stress.” Despite the fact that GFAP expression is affected by pressure, none of the complement genes studied showed any change in expression in either cultured cells or retinas. At the same time, one has to keep in mind that in the organotypic culture model there is inherent and unavoidable tissue damage, which may precipitate generalized complement activation that may mask small changes caused by the pressure level difference. Inspection of the RT-qPCR dissociation curves from organotypic cultures and retinas from the in vivo experiments suggests that this is not the case, but such a possibility cannot be completely excluded.
Despite the appeal of in vitro experiments, it is in vivo studies that ultimately allow us to answer whether results from in vitro experiments are relevant to actual biologic processes occurring in the organism. Inducible models of glaucoma in rodents have advantages of precise control of pathology onset. The recently developed microbead occlusion model has been shown to induce RGC damage in mice mediated by increased IOP [63]. In this model appreciable RGC loss could be detected as early as 2 weeks after the onset of IOP elevation [21]. The experiments described above examined whether short-term elevation of IOP caused by microbead injection in the anterior chamber of the eye leads to upregulation of the complement system in the mouse retina. The results suggest that IOP elevation for up to 7 days in C57BL/6 mice does not produce changes in complement expression for any of the genes studied and in C1q protein levels. This finding is surprising given the potential role of complement in the pathogenesis of glaucoma. It is, however, possible that IOP elevation in these mice was not of sufficient duration to cause complement upregulation. We are presently studying complement expression in animals exposed to elevated IOP for longer periods of time.
In the present study, we attempted to investigate the effect of short-term pressure elevation on complement gene expression in the retina by using a multifaceted approach that combined various in vitro and in vivo models. Studies in TR-MUL5 cell lines did not detect significant changes in cell responses to elevated pressure for up to 3 days in culture. Using an in vitro system in which entire retinal tissue is subjected to elevated pressure, i.e., organotypic retinal cultures, we similarly did not detect any significant changes in complement upregulation as a function of elevated pressure for up to 3 days. Those results were mirrored by the data from an in vivo model in which we subjected mouse retinas to experimental IOP elevation for 7 days. Combined, our results show that short-term exposure to elevated pressure alone does not dramatically affect complement gene expression in rodents and in organotypic cultures in primates.
This study was designed to test the null hypothesis that pressure does not affect complement expression in the retina. As such, the results of this study cannot be used as proof that such changes do not occur. To do that, a study with much higher power testing and an alternative null hypothesis would be technically needed. However, the absence of changes in complement activation across all the models employed in the current study strongly suggests that reported involvement of the complement system in glaucoma is independent of short-term changes of IOP. Either long-term elevation in IOP leads to pronounced cumulative effects of pressure or an additional stimulus is required to cause complement upregulation and activation, like the one seen in DBA/2J mice [4], laser-treated rats [5], or patients having glaucoma [3-5]. Additional studies are needed to elucidate the role of complement activation in glaucoma.
Acknowledgments
We would like to thank Dr. Thomas Weber of Mount Sinai School of Medicine for providing us with the TR-MUL5 cell line. Support: R01EY015224, Unrestricted Challenge grant and student fellowship grants (CD and GK) from Research to Prevent Blindness, Inc.
References
- 1.Chauhan BC, Pan J, Archibald ML, LeVatte TL, Kelly ME, Tremblay F. Effect of intraocular pressure on optic disc topography, electroretinography, and axonal loss in a chronic pressure-induced rat model of optic nerve damage. Invest Ophthalmol Vis Sci. 2002;43:2969–76. [PubMed] [Google Scholar]
- 2.Quigley HA, Addicks EM, Green WR, Maumenee AE. Optic nerve damage in human glaucoma. II. The site of injury and susceptibility to damage. Arch Ophthalmol. 1981;99:635–49. doi: 10.1001/archopht.1981.03930010635009. [DOI] [PubMed] [Google Scholar]
- 3.Tezel G, Yang X, Luo C, Kain AD, Powell DW, Kuehn MH, Kaplan HJ. Oxidative stress and the regulation of complement activation in human glaucoma. Invest Ophthalmol Vis Sci. 2010;51:5071–82. doi: 10.1167/iovs.10-5289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Stasi K, Nagel D, Yang X, Wang RF, Ren L, Podos SM, Mittag T, Danias J. Complement component 1Q (C1Q) upregulation in retina of murine, primate, and human glaucomatous eyes. Invest Ophthalmol Vis Sci. 2006;47:1024–9. doi: 10.1167/iovs.05-0830. [DOI] [PubMed] [Google Scholar]
- 5.Kuehn MH, Kim CY, Ostojic J, Bellin M, Alward WL, Stone EM, Sakaguchi DS, Grozdanic SD, Kwon YH. Retinal synthesis and deposition of complement components induced by ocular hypertension. Exp Eye Res. 2006;83:620–8. doi: 10.1016/j.exer.2006.03.002. [DOI] [PubMed] [Google Scholar]
- 6.Jha P, Banda H, Tytarenko R, Bora PS, Bora NS. Complement mediated apoptosis leads to the loss of retinal ganglion cells in animal model of glaucoma. Mol Immunol. 2011;48:2151–8. doi: 10.1016/j.molimm.2011.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Howell GR, Macalinao DG, Sousa GL, Walden M, Soto I, Kneeland SC, Barbay JM, King BL, Marchant JK, Hibbs M, Stevens B, Barres BA, Clark AF, Libby RT, John SW. Molecular clustering identifies complement and endothelin induction as early events in a mouse model of glaucoma. J Clin Invest. 2011;121:1429–44. doi: 10.1172/JCI44646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Song WC, Sarrias MR, Lambris JD. Complement and innate immunity. Immunopharmacology. 2000;49:187–98. doi: 10.1016/s0162-3109(00)80303-3. [DOI] [PubMed] [Google Scholar]
- 9.Ricklin D, Hajishengallis G, Yang K, Lambris JD. Complement: a key system for immune surveillance and homeostasis. Nat Immunol. 2010;11:785–97. doi: 10.1038/ni.1923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Luo C, Chen M, Xu H. Complement gene expression and regulation in mouse retina and retinal pigment epithelium/choroid. Mol Vis. 2011;17:1588–97. [PMC free article] [PubMed] [Google Scholar]
- 11.Stevens B, Allen NJ, Vazquez LE, Howell GR, Christopherson KS, Nouri N, Micheva KD, Mehalow AK, Huberman AD, Stafford B, Sher A, Litke AM, Lambris JD, Smith SJ, John SW, Barres BA. The classical complement cascade mediates CNS synapse elimination. Cell. 2007;131:1164–78. doi: 10.1016/j.cell.2007.10.036. [DOI] [PubMed] [Google Scholar]
- 12.Mastellos D, Germenis AE, Lambris JD. Complement: an inflammatory pathway fulfilling multiple roles at the interface of innate immunity and development. Curr Drug Targets Inflamm Allergy. 2005;4:125–7. doi: 10.2174/1568010053622993. [DOI] [PubMed] [Google Scholar]
- 13.Hakobyan S, Boyajyan A, Sim RB. Classical pathway complement activity in schizophrenia. Neurosci Lett. 2005;374:35–7. doi: 10.1016/j.neulet.2004.10.024. [DOI] [PubMed] [Google Scholar]
- 14.Mayilyan KR, Arnold JN, Presanis JS, Soghoyan AF, Sim RB. Increased complement classical and mannan-binding lectin pathway activities in schizophrenia. Neurosci Lett. 2006;404:336–41. doi: 10.1016/j.neulet.2006.06.051. [DOI] [PubMed] [Google Scholar]
- 15.Fonseca MI, Ager RR, Chu SH, Yazan O, Sanderson SD, LaFerla FM, Taylor SM, Woodruff TM, Tenner AJ. Treatment with a C5aR antagonist decreases pathology and enhances behavioral performance in murine models of Alzheimer's disease. J Immunol. 2009;183:1375–83. doi: 10.4049/jimmunol.0901005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Loeffler DA, Camp DM, Conant SB. Complement activation in the Parkinson's disease substantia nigra: an immunocytochemical study. J Neuroinflammation. 2006;3:29. doi: 10.1186/1742-2094-3-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ren L, Danias J. A role for complement in glaucoma? Adv Exp Med Biol. 2010;703:95–104. doi: 10.1007/978-1-4419-5635-4_7. [DOI] [PubMed] [Google Scholar]
- 18.Sarthy VP, Brodjian SJ, Dutt K, Kennedy BN, French RP, Crabb JW. Establishment and characterization of a retinal Muller cell line. Invest Ophthalmol Vis Sci. 1998;39:212–6. [PubMed] [Google Scholar]
- 19.Tomi M, Funaki T, Abukawa H, Katayama K, Kondo T, Ohtsuki S, Ueda M, Obinata M, Terasaki T, Hosoya K. Expression and regulation of L-cystine transporter, system xc-, in the newly developed rat retinal Muller cell line (TR-MUL). Glia. 2003;43:208–17. doi: 10.1002/glia.10253. [DOI] [PubMed] [Google Scholar]
- 20.Johnson TV, Martin KR. Development and characterization of an adult retinal explant organotypic tissue culture system as an in vitro intraocular stem cell transplantation model. Invest Ophthalmol Vis Sci. 2008;49:3503–12. doi: 10.1167/iovs.07-1601. [DOI] [PubMed] [Google Scholar]
- 21.Chen H, Wei X, Cho KS, Chen G, Sappington R, Calkins DJ, Chen DF. Optic neuropathy due to microbead-induced elevated intraocular pressure in the mouse. Invest Ophthalmol Vis Sci. 2011;52:36–44. doi: 10.1167/iovs.09-5115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Danias J, Kontiola AI, Filippopoulos T, Mittag T. Method for the noninvasive measurement of intraocular pressure in mice. Invest Ophthalmol Vis Sci. 2003;44:1138–41. doi: 10.1167/iovs.02-0553. [DOI] [PubMed] [Google Scholar]
- 23.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Methods (San Diego, Calif) 2001;25:402–8. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 24.Grubbs F. Procedures for Detecting Outlying Observations in Samples. Technometrics. 1969;11:1–21. [Google Scholar]
- 25.Panagis L, Zhao X, Ge Y, Ren L, Mittag TW, Danias J. Gene expression changes in areas of focal loss of retinal ganglion cells in the retina of DBA/2J mice. Invest Ophthalmol Vis Sci. 2010;51:2024–34. doi: 10.1167/iovs.09-3560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Manderson AP, Botto M, Walport MJ. The role of complement in the development of systemic lupus erythematosus. Annu Rev Immunol. 2004;22:431–56. doi: 10.1146/annurev.immunol.22.012703.104549. [DOI] [PubMed] [Google Scholar]
- 27.Skerka C, Licht C, Mengel M, Uzonyi B, Strobel S, Zipfel PF, Jozsi M. Autoimmune forms of thrombotic microangiopathy and membranoproliferative glomerulonephritis: Indications for a disease spectrum and common pathogenic principles. Mol Immunol. 2009;46:2801–7. doi: 10.1016/j.molimm.2009.05.018. [DOI] [PubMed] [Google Scholar]
- 28.Panaro MA, Cianciulli A. Current opinions and perspectives on the role of immune system in the pathogenesis of Parkinson's disease. Curr Pharm Des. 2012;18:200–8. doi: 10.2174/138161212799040574. [DOI] [PubMed] [Google Scholar]
- 29.Crehan H, Holton P, Wray S, Pocock J, Guerreiro R, Hardy J. Complement receptor 1 (CR1) and Alzheimer's disease. Immunobiology. 2012;217:244–50. doi: 10.1016/j.imbio.2011.07.017. [DOI] [PubMed] [Google Scholar]
- 30.Benoit ME, Tenner AJ. Complement protein C1q-mediated neuroprotection is correlated with regulation of neuronal gene and microRNA expression. J Neurosci. 2011;31:3459–69. doi: 10.1523/JNEUROSCI.3932-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Vogt SD, Barnum SR, Curcio CA, Read RW. Distribution of complement anaphylatoxin receptors and membrane-bound regulators in normal human retina. Exp Eye Res. 2006;83:834–40. doi: 10.1016/j.exer.2006.04.002. [DOI] [PubMed] [Google Scholar]
- 32.Bora NS, Jha P, Bora PS. The role of complement in ocular pathology. Semin Immunopathol. 2008;30:85–95. doi: 10.1007/s00281-008-0110-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Haines JL, Hauser MA, Schmidt S, Scott WK, Olson LM, Gallins P, Spencer KL, Kwan SY, Noureddine M, Gilbert JR, Schnetz-Boutaud N, Agarwal A, Postel EA, Pericak-Vance MA. Complement factor H variant increases the risk of age-related macular degeneration. Science. 2005;308:419–21. doi: 10.1126/science.1110359. [DOI] [PubMed] [Google Scholar]
- 34.Miyahara T, Kikuchi T, Akimoto M, Kurokawa T, Shibuki H, Yoshimura N. Gene microarray analysis of experimental glaucomatous retina from cynomologous monkey. Invest Ophthalmol Vis Sci. 2003;44:4347–56. doi: 10.1167/iovs.02-1032. [DOI] [PubMed] [Google Scholar]
- 35.Bosco A, Steele MR, Vetter ML. Early microglia activation in a mouse model of chronic glaucoma. J Comp Neurol. 2011;519:599–620. doi: 10.1002/cne.22516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ebneter A, Casson RJ, Wood JP, Chidlow G. Microglial activation in the visual pathway in experimental glaucoma: spatiotemporal characterization and correlation with axonal injury. Invest Ophthalmol Vis Sci. 2010;51:6448–60. doi: 10.1167/iovs.10-5284. [DOI] [PubMed] [Google Scholar]
- 37.Akanuma S, Soutome T, Hisada E, Tachikawa M, Kubo Y, Hosoya K. Na+-independent nucleoside transporters regulate adenosine and hypoxanthine levels in Muller cells and the inner blood-retinal barrier. Invest Ophthalmol Vis Sci. 2013;54:1469–77. doi: 10.1167/iovs.12-10905. [DOI] [PubMed] [Google Scholar]
- 38.Limb GA, Salt TE, Munro PM, Moss SE, Khaw PT. In vitro characterization of a spontaneously immortalized human Muller cell line (MIO-M1). Invest Ophthalmol Vis Sci. 2002;43:864–9. [PubMed] [Google Scholar]
- 39.Taylor S, Calder CJ, Albon J, Erichsen JT, Boulton ME, Morgan JE. Involvement of the CD200 receptor complex in microglia activation in experimental glaucoma. Exp Eye Res. 2011;92:338–43. doi: 10.1016/j.exer.2011.01.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ando D, Kubo Y, Akanuma S, Yoneyama D, Tachikawa M, Hosoya K. Function and regulation of taurine transport in Muller cells under osmotic stress. Neurochem Int. 2012;60:597–604. doi: 10.1016/j.neuint.2012.02.018. [DOI] [PubMed] [Google Scholar]
- 41.Williams JA, Greenwood J, Moss SE. Retinal changes precede visual dysfunction in the complement factor H knockout mouse. PLoS ONE. 2013;8:e68616. doi: 10.1371/journal.pone.0068616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hernandez MR, Pena JD, Selvidge JA, Salvador-Silva M, Yang P. Hydrostatic pressure stimulates synthesis of elastin in cultured optic nerve head astrocytes. Glia. 2000;32:122–36. doi: 10.1002/1098-1136(200011)32:2<122::aid-glia20>3.0.co;2-j. [DOI] [PubMed] [Google Scholar]
- 43.Agar A, Li S, Agarwal N, Coroneo MT, Hill MA. Retinal ganglion cell line apoptosis induced by hydrostatic pressure. Brain Res. 2006;1086:191–200. doi: 10.1016/j.brainres.2006.02.061. [DOI] [PubMed] [Google Scholar]
- 44.Liu Q, Ju WK, Crowston JG, Xie F, Perry G, Smith MA, Lindsey JD, Weinreb RN. Oxidative stress is an early event in hydrostatic pressure induced retinal ganglion cell damage. Invest Ophthalmol Vis Sci. 2007;48:4580–9. doi: 10.1167/iovs.07-0170. [DOI] [PubMed] [Google Scholar]
- 45.Tezel G, Wax MB. Increased production of tumor necrosis factor-alpha by glial cells exposed to simulated ischemia or elevated hydrostatic pressure induces apoptosis in cocultured retinal ganglion cells. J Neurosci. 2000;20:8693–8700. doi: 10.1523/JNEUROSCI.20-23-08693.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wax MB, Tezel G, Kobayashi S, Hernandez MR. Responses of different cell lines from ocular tissues to elevated hydrostatic pressure. Br J Ophthalmol. 2000;84:423–8. doi: 10.1136/bjo.84.4.423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Sappington RM, Chan M, Calkins DJ. Interleukin-6 protects retinal ganglion cells from pressure-induced death. Invest Ophthalmol Vis Sci. 2006;47:2932–42. doi: 10.1167/iovs.05-1407. [DOI] [PubMed] [Google Scholar]
- 48.Juel HB, Kaestel C, Folkersen L, Faber C, Heegaard NH, Borup R, Nissen MH. Retinal pigment epithelial cells upregulate expression of complement factors after co-culture with activated T cells. Exp Eye Res. 2011;92:180–8. doi: 10.1016/j.exer.2011.01.003. [DOI] [PubMed] [Google Scholar]
- 49.Bähr M, Vanselow J, Thanos S. In vitro regeneration of adult rat ganglion cell axons from retinal explants. Exp Brain Res. 1988;73:393–401. doi: 10.1007/BF00248232. [DOI] [PubMed] [Google Scholar]
- 50.Sergeev SA, Khramova YV, Semenova ML, Saburina IN, Kosheleva NV. Behavior of Transplanted Multipotent Cells after in Vitro Transplantation into the Damaged Retina. Acta Naturae. 2011;3:66–72. [PMC free article] [PubMed] [Google Scholar]
- 51.Xin H, Yannazzo JA, Duncan RS, Gregg EV, Singh M, Koulen P. A novel organotypic culture model of the postnatal mouse retina allows the study of glutamate-mediated excitotoxicity. J Neurosci Methods. 2007;159:35–42. doi: 10.1016/j.jneumeth.2006.06.013. [DOI] [PubMed] [Google Scholar]
- 52.Kaempf S, Johnen S, Salz AK, Weinberger A, Walter P, Thumann G. Effects of bevacizumab (Avastin) on retinal cells in organotypic culture. Invest Ophthalmol Vis Sci. 2008;49:3164–71. doi: 10.1167/iovs.07-1265. [DOI] [PubMed] [Google Scholar]
- 53.Moritoh S, Tanaka KF, Jouhou H, Ikenaka K, Koizumi A. Organotypic tissue culture of adult rodent retina followed by particle-mediated acute gene transfer in vitro. PLoS ONE. 2010;5:e12917. doi: 10.1371/journal.pone.0012917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Niyadurupola N, Sidaway P, Osborne A, Broadway DC, Sanderson J. The development of human organotypic retinal cultures (HORCs) to study retinal neurodegeneration. Br J Ophthalmol. 2011;95:720–6. doi: 10.1136/bjo.2010.181404. [DOI] [PubMed] [Google Scholar]
- 55.Ethier CR, Read AT, Chan DW. Effects of latrunculin-B on outflow facility and trabecular meshwork structure in human eyes. Invest Ophthalmol Vis Sci. 2006;47:1991–8. doi: 10.1167/iovs.05-0327. [DOI] [PubMed] [Google Scholar]
- 56.Salvador-Silva M, Aoi S, Parker A, Yang P, Pecen P, Hernandez MR. Responses and signaling pathways in human optic nerve head astrocytes exposed to hydrostatic pressure in vitro. Glia. 2004;45:364–77. doi: 10.1002/glia.10342. [DOI] [PubMed] [Google Scholar]
- 57.Lei Y, Rajabi S, Pedrigi RM, Overby DR, Read AT, Ethier CR. In vitro models for glaucoma research: effects of hydrostatic pressure. Invest Ophthalmol Vis Sci. 2011;52:6329–39. doi: 10.1167/iovs.11-7836. [DOI] [PubMed] [Google Scholar]
- 58.Sappington RM, Sidorova T, Long DJ, Calkins DJ. TRPV1: contribution to retinal ganglion cell apoptosis and increased intracellular Ca2+ with exposure to hydrostatic pressure. Invest Ophthalmol Vis Sci. 2009;50:717–28. doi: 10.1167/iovs.08-2321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Gerometta R, Escobar D, Candia OA. An hypothesis on pressure transmission from anterior chamber to optic nerve. Med Hypotheses. 2011;77:827–31. doi: 10.1016/j.mehy.2011.07.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Herx LM, Yong VW. Interleukin-1 beta is required for the early evolution of reactive astrogliosis following CNS lesion. J Neuropathol Exp Neurol. 2001;60:961–71. doi: 10.1093/jnen/60.10.961. [DOI] [PubMed] [Google Scholar]
- 61.Humphrey MF, Chu Y, Mann K, Rakoczy P. Retinal GFAP and bFGF expression after multiple argon laser photocoagulation injuries assessed by both immunoreactivity and mRNA levels. Exp Eye Res. 1997;64:361–9. doi: 10.1006/exer.1996.0219. [DOI] [PubMed] [Google Scholar]
- 62.Panagis L, Thanos S, Fischer D, Dermon CR. Unilateral optic nerve crush induces bilateral retinal glial cell proliferation. Eur J Neurosci. 2005;21:2305–9. doi: 10.1111/j.1460-9568.2005.04046.x. [DOI] [PubMed] [Google Scholar]
- 63.Sappington RM, Carlson BJ, Crish SD, Calkins DJ. The microbead occlusion model: a paradigm for induced ocular hypertension in rats and mice. Invest Ophthalmol Vis Sci. 2010;51:207–16. doi: 10.1167/iovs.09-3947. [DOI] [PMC free article] [PubMed] [Google Scholar]