

Abstract
We revisit the family Diademaceae based on available sequence data and morphology. Diademaceae is characterized by ascomata opening with a flat circular lid and fissitunicate, short orbicular frequently cylindrical, pedicellate asci. Ascospores are frequently circular in section but narrowing to one end with three or more transverse septa, without longitudinal septa, and mostly with a thick sheath. In recent treatments Clathrospora, Comoclathris, Diadema, Diademosa, and Graphyllium were placed in the family. Following molecular and morphological study, Clathrospora, Comoclathris, and Diademosa, are excluded from the family and referred to Pleosporaceae. Graphyllium is excluded from Diademaceae, based on hysterothecium-like ascomata with a longitudinal opening, and tentatively placed in Hysteriaceae with uncertainty; species with hysterothecia have now been accommodated in at least five families. The study accepts only Diadema in the family. The status of Diademaceae as a distinct family, based on the ascomata opening by a flat circular lid, is thought to be doubtful. Fresh collections of Diadema are needed for epitypification and to obtain sequence data to establish if this is a well-resolved family.
1. Introduction
Based on the ascomata opening by a flat circular lid, Shoemaker and Babcock [1] introduced Diademaceae, which they considered to be a unique family in the order Pleosporales. Initially five genera, that is, Clathrospora, Comoclathris, Diadema, Diademosa, and Macrospora, were included in the family [1]. Other than the ascomata opening by a flat circular lid, the family was characterized by bitunicate and fissitunicate, clavate or ellipsoidal, short pedicellate asci, and applanate or rarely cylindrical ascospores with three or more transverse septa with or without longitudinal septa and usually with a thick sheath and frequently circular in section but narrowing to one end [1, 2].
Species of the order Pleosporales with applanate ascospores can be found in three families (Diademaceae, Hysteriaceae, and Pleosporaceae), which differ in the way the ascomata open [1]. Ascomata openings by a flat circular lid are characteristic of Diademaceae. In species of Hysteriaceae ascomata open via a long narrow slit and species of Pleosporaceae open by a central pore [1]. Various authors have included and excluded different genera in Diademaceae by giving priority to different morphological characters [1, 3, 4]. Platyspora had been referred to this family by various authors [4] or was considered a synonym of Graphyllium [2, 4] or Comoclathris [5]. Lumbsch and Huhndorf [6] assigned Macrospora to Pleosporaceae, as the genus was considered to be a synonym of Pyrenophora and this treatment was followed by Zhang et al. [3, 4]. In the same study, Lumbsch and Huhndorf [6] had also referred Graphyllium to Diademaceae. Sequence data is now available for some of these genera thus the importance of their morphological characters and overall relationships can now be tested based on molecular phylogeny.
We have been studying the families of Pleosporales in order to provide a natural classification via morphological characterization together with molecular analysis [2–4, 7]. The family Diademaceae has been poorly studied and presently comprises five genera [6], but this has not changed since the family was introduced by Shoemaker and Babcock [1]. Given the considerable taxonomic confusion we revisited this family based on phylogenetic analyses of rDNA sequence data coupled with morphological characters. The aims of the study are to (i) discuss the familial placement of the genera in Diademaceae and assess whether they represent natural groups, (ii) determine which morphological characters are useful for generic delineation by observing the type species of each genera, and (iii) illustrate the genera to stimulate fresh collections being made so that molecular data can be used to resolve the systematic relationships of the family.
2. Materials and Methods
2.1. Specimen Examination
The basic methodology used in this study was the same as Ariyawansa et al. [7]. The type specimens were loaned from the US National Fungus Collections (BPI), Agriculture and Agri-Food Canada (DAOM), and New York Botanical Garden (NY). Ascomata were rehydrated in 5% KOH prior to examination and sectioning. Hand sections of the fruiting structures were mounted in water for microscopic studies and photomicrography. The fungus was examined in a Nikon ECLIPSE 80i compound microscope and photographed by a Cannon 450D digital camera fitted to the microscope. Measurements were made with the Tarosoft (R) Image Frame Work program and images used for figures were processed with Adobe Photoshop CS3 Extended version 10.0 software (Adobe Systems Inc., USA).
2.2. Phylogenetic Analysis
The large and small subunits of the nuclear ribosomal RNA genes (LSU, SSU) were included in the analysis. All sequences obtained from GenBank were used in Schoch et al. [8] and Zhang et al. [3] and are listed in Table 1. Sequences were aligned using Bioedit v7.2.0 version [9] and ClustalX v. 1.83 [10]. The alignments were checked visually and improved manually where necessary.
Table 1.
Taxa used in the phylogenetic analysis and their corresponding GenBank numbers. Culture and voucher abbreviations are indicated where available.
| Taxon | Culture | SSU | LSU |
|---|---|---|---|
| Aigialus grandis | JK 5244A | GU296131 |
GU301793 |
| Aigialus parvus | BCC 18403 | GU479743 | GU479778 |
| Alternaria alternata | CBS 916.96 | KC584507 | DQ678082 |
| Amniculicola immersa | CBS 123083 | GU456295 | FJ795498 |
| Amniculicola parva | CBS 123092 | GU296134 | FJ795497 |
| Ascocratera manglicola | JK 5262C | GU296136 | GU301799 |
| Bimuria novae-zelandiae | CBS 107.79 | AY016338 | AY016356 |
| Boeremia exigua | CBS 431.74 | EU754084 | EU754183 |
| Byssothecium circinans | CBS 675.92 | AY016339 | AY016357 |
| Clathrospora elynae | CBS 161.51 | KC584628 | KC584370 |
| Clathrospora elynae | CBS 196.54 | KC584629 | KC584371 |
| Clathrospora heterospora (Alternaria sp.) | CBS 175.52 | KC584577 | KC584320 |
| Cochliobolus heterostrophus | CBS 134.39 | AY544727 | AY544645 |
| Comoclathris compressa | CBS 156.53 | KC584630 | KC584372 |
| Comoclathris compressa | CBS 157.53 | KC584631 | KC584373 |
| Comoclathris magna (Alternaria sp.) | CBS 174.52 | KC584578 | DQ678068 |
| Didymella exigua | CBS 183.55 | EU754056 | EU754155 |
| Dothidea sambuci | DAOM 231303 | AY544722 | AY544681 |
| Dothidotthia aspera | CPC 12933 | EU673228 | EU673276 |
| Dothidotthia symphoricarpi | CPC 12929 | EU673224 | EU673273 |
| Halojulella avicenniae | BCC 18422 | GU371831 | GU371823 |
| Halojulella avicenniae | BCC 20173 | GU371830 | GU371822 |
| Helicascus nypae | BCC 36752 | GU479755 | GU479789 |
| Katumotoa bambusicola | MAFF 239641 | AB524454 | AB524595 |
| Lentithecium aquaticum | CBS 123099 | GU296156 | GU301823 |
| Lentithecium fluviatile | CBS 122367 | GU296158 | GU301825 |
| Leptosphaeria doliolum | CBS 505.75 | GU296159 | GU301827 |
| Leptosphaeria dryadis | CBS 643.86 | GU301828 | |
| Leptosphaeria maculans | DAOM 229267 | DQ470993 | DQ470946 |
| Leptosphaerulina australis | CBS 317.83 | GU296160 | GU301830 |
| Massarina eburnea | CBS 473.64 | GU296170 | GU301840 |
| Montagnula opulenta | CBS 168.34 | AF164370 | DQ678086 |
| Morosphaeria ramunculicola | BCC 18405 | GQ925839 | GQ925854 |
| Morosphaeria ramunculicola | JK 5304B | GU479760 | GU479794 |
| Neophaeosphaeria filamentosa | CBS 102202 | GQ387516 | GQ387577 |
| Neottiosporina paspali | CBS 331.37 | EU754073 | EU754172 |
| Ophiosphaerella herpotricha | CBS 240.31 | DQ678010 | DQ678062 |
| Phaeosphaeria eustoma | CBS 573.86 | DQ678011 | DQ678063 |
| Phoma radicina | CBS 111.79 | EU754092 | EU754191 |
| Pleomassaria siparia | CBS 279.74 | DQ678027 | DQ678078 |
| Pleospora betae | CBS 109410 | EU754079 | EU754178 |
| Pleospora calvescens | CBS 246.79 | EU754032 | EU754131 |
| Pleospora chenopodii | CBS 206.80 | JF740095 | JF740266 |
| Pleospora herbarum | CBS 191.86 | DQ247812 | DQ247804 |
| Pleospora incompta | CBS 467.76 | GU238220 | GU238087 |
| Pleospora typhicola | CBS 132.69 | JF740105 | JF740325 |
| Preussia terricola | DAOM 230091 | AY544726 | AY544686 |
| Prosthemium betulinum | CBS 127468 | AB553644 | AB553754 |
| Prosthemium canba | JCM 16966 | AB553646 | AB553760 |
| Pyrenophora phaeocomes | DAOM 222769 | DQ499595 | DQ499596 |
| Sporormiella minima | CBS 524.50 | DQ678003 | DQ678056 |
| Sporormiella minima | CBS 524.50 | DQ678003 | DQ678056 |
Maximum Likelihood analysis was performed in RAxML [11] implemented in raxmlGUIv.0.9b2 [12]. The search strategy was set to rapid bootstrapping and the analysis was carried out using the GTRGAMMAI model of nucleotide substitution. The number of replicates was automatically inferred using the stopping criterion [13]. Maximum Likelihood bootstrap values equal or greater than 50% are given below or above each node (Figure 1). Phylogenetic trees were drawn using Treeview v. 1.6.6 [Page 2001].
Figure 1.
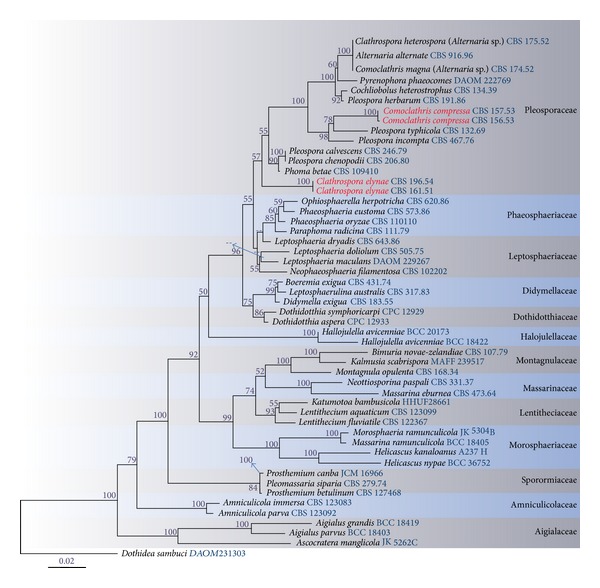
RAxML tree based on a combined dataset of SSU and LSU. Bootstrap support values >50% are shown above or below the branch. The putative strains of Clathrospora elynae (CBS 196.54 and CBS 161.51) and Comoclathris compressa (CBS 157.53 and CBS 156.53) are indicated in red. Dothidea sambuci is the out-group taxon. The original isolate numbers are noted after the species names.
3. Results
3.1. Molecular Phylogeny Based on Combined nrSSU and nrLSU
The combined 18 S and 28 S nrDNA data set comprised 52 taxa including strains of Clathrospora elynae (CBS 196.54 and CBS 161.51), Comoclathris magna (CBS 174.52), Clathrospora heterospora (CBS 175.52), and Comoclathris compressa (CBS 157.53 and CBS 156.53) with Dothidea sambuci as the out-group taxon. The 52 taxa analyzed in the cladogram formed 13 familial clades. Maximum Likelihood analysis used 1000 bootstrap replicates and yielded a tree with the likelihood value of ln:-9930.285726 and the following model parameters: alpha: 0.512987 and invar: 0.499567; Π(A): 0.259512, Π(C): 0.207265, Π(G): 0.277826, and Π(T): 0.255397. Phylogenetic trees obtained from maximum likelihood analyses yielded trees with similar overall topology at family and generic relationship in agreement with previous work [3, 4, 8].
3.2. Molecular Phylogeny of Diademaceae
Two putative strains of Clathrospora elynae (CBS 196.54 and CBS 161.51) which had been previously referred to Diademaceae by Lumbsch and Huhndorf [6] and Shoemaker and Babcock [1] were clustered in the family Pleosporaceae but separated from other genera of the family with a relatively high bootstrap value (55%). The type species of Comoclathris, C. lanata, was not available for study, but the two Comoclathris compressa strains cluster in a well supported clade within the Pleosporaceae, outside the Alternaria complex. Therefore we confer with Zhang et al. [4] and Woudenberg et al. [14] in transferring these two genera to Pleosporaceae. Two putative strains of Comoclathris magna (CBS 174.52) and Clathrospora heterospora (CBS 175.52) were clustered within the Alternaria complex as in Woudenberg et al. [14]. Woudenberg et al. [14] have tentatively considered Comoclathris magna (CBS 174.52) and Clathrospora heterospora (CBS 175.52) as Alternaria species. There is, however, confusion concerning the CBS 175.52 strain, because Dong et al. [15] used the name Comoclathris baccata in their paper for strain CBS 175.52 but submitted sequences to GenBank under the name Clathrospora diplospora [14]. In their study, Woudenberg et al. [14] have synonymised Comoclathris baccata with C. heterospora. We could not locate the type species of Diadema, Diadema tetramerum, and Diademosa, Diademosa californiana, for phylogenetic analysis due to the unavailability of sequence data. Therefore recollection, epitypification, and sequence data of Diadema, Diadema tetramerum, and Diademosa, Diademosa californiana, are necessary to validate Diademaceae genera and species relationships.
3.3. Taxonomy
Diademaceae. Shoemaker & C.E. Babc., Can. J. Bot. 70(8): 1618 (1992), MycoBank: MB 81955.
Parasitic or saprobic in stems and leaves. Sexual state: Ascomata subepidermal or subcuticular and later become superficial, globose, opening via flat circular lid, dark brown to black. Peridium thin, consisting of small pigmented thick-walled cells of textura angularis. Hamathecium of dense cellular pseudoparaphyses. Asci 8-spored, bitunicate, fissitunicate, clavate or ellipsoidal, short orbicular pedicel, without an ocular chamber. Ascospores partially overlapping to biseriate, fusiform, brown, with three or more transverse septa without longitudinal septa, mostly terete (cylindrical; frequently circular in section but narrowing to one end), mostly with a thick sheath. Asexual state: Unknown.
Type: Diadema. Shoemaker & C.E. Babc.
Shoemaker and Babcock [1] introduced Diademaceae which they considered to be a distinctive family comprising Clathrospora, Comoclathris, Diadema, Diademosa, and Macrospora whose species have ascomata opening by a flat circular lid [1]. The feature of ascomata opening via a flat circular lid was considered to be an adaptation to the alpine habitat [16]. Ascospores are fusiform, brown, with three or more transverse septa, with or without longitudinal septa, and frequently terete, usually with a thick sheath [1, 2, 4].
Lumbsch and Huhndorf [6] excluded Macrospora from Diademaceae and assigned it to Pleosporaceae, as it was considered to be a synonym for Pyrenophora. We have seen type material of Macrospora scirpicola and it is neither diademaceous nor pleosporaceous and therefore will be considered as subject of a future paper. Lumbsch and Huhndorf [6] also included Graphyllium in the family Diademaceae, but this classification has not been followed by many authors. Shoemaker and Babcock [1] and Zhang et al. [4] referred Graphyllium to the family Hysteriaceae based on its hysterothecium-like ascomata forming a longitudinal, slit-like opening. Shoemaker and Babcock [1] assigned Clathrospora to Diademaceae based on ascomata opening with an intraepidermal discoid lid and muriform applanate ascospores with more than one row of longitudinal septa. Ascomata, however, have slightly papillate ostioles and Alternaria-like asexual morphs, and recent molecular data shows that Clathrospora has an affinity with the family Pleosporaceae [4, 14]. Platyspora has been referred to Diademaceae [4] and was considered a synonym of Graphyllium [2, 4] or as a synonym of Comoclathris [5].
Species of the order Pleosporales with applanate ascospores were previously separated into three families (Diademaceae, Hysteriaceae, and Pleosporaceae) which differ in the way the ascomata open [2]. Ascomata openings by a flat circular lid were characteristic of the family Diademaceae, while species of Hysteriaceae open via a long narrow slit and species of Pleosporaceae open by a central pore [1]. Based on the above discussion we exclude Clathrospora, Comoclathris, Diademosa, and Graphyllium from the Diademaceae. Based on morphology and/or molecular data and at this time, we accept only Diadema, which has mostly terete ascospores (except D. obtusa which has flattened ascospores), in the family. Diademaceae is, however, not supported by molecular data, but no sequence data is available for the generic type Diadema. Further studies are required to resolve the phylogenetic relationship in the Pleosporales. In the light of all of the above, we retain the Diademaceae to include a single genus Diadema which has immersed, intraepidermal ascomata, opening via a flat circular lid, and asci with a short orbicular pedicel without an ocular chamber and ascospores are reddish-brown, usually cylindrical, and frequently circular in section but narrowing to one end with a distinct, mucilaginous sheath.
3.4. Accepted Genus in Diademaceae
Diadema. Shoemaker & C.E. Babc., Can. J. Bot. 67(5): 1349 (1989), MycoBank: MB 25293.
Saprobic on culms of grasses (Poaceae). Sexual state: Ascomata scattered, immersed, intra-epidermal, globose to subglobose, black to brown, smooth-walled and opening via a flat circular lid. Peridium 1-layered, composed of small pigmented thick walled compressed cells, base composed of small pigmented thick-walled cells of textura angularis. Hamathecium of dense, numerous, septate, hyaline, cellularpseudoparaphyses. Asci 8-spored, numerous, bitunicate, fissitunicate, broadly-clavate, with a short orbicular pedicel, without an ocular chamber. Ascospores obliquely biseriate, broadly fusiform, usually cylindrical; frequently circular in section but narrowing to one end, brown to reddish-brown, without longitudinal septa, guttulate, smooth-walled or finely punctate, with wide, distinct mucilaginous sheath. Asexual state: Unknown.
Type Species: Diadema tetramerum. Shoemaker & C.E. Babc. [as “tetramera”], Can. J. Bot. 67(5): 1354 (1989), MycoBank: MB 136222 (see Figure 2).
Figure 2.

Diadema tetramerum (holotype). (a) Ascomata on substrate opening via a flat circular lid. (b) Vertical section of ascoma. (c) Closeup of the peridium. (d) Hyaline, septate pseudoparaphyses. (e) Apical part of the asci, ((f)-(g)) Asci with short orbicular pedicel. ((h)–(j)) Reddish-brown ascospores with broad sheath. Scale bars: (b) = 100 μm, (c) = 10 μm, ((d)–(g)) = 60 μm, and ((h)–(j)) = 30 μm.
Saprobic on culms of grasses (Poaceae). Sexual state: Ascomata 170–200 × 150–270 μm ( μm, n = 10), scattered, immersed, intra-epidermal, globose to subglobose, black to brown, smooth-walled and opening via a flat circular lid. Peridium 10–22 μm (, n = 20), 1-layered, composed of small, pigmented, thick-walled, compressed cells, base composed of small, pigmented, thick-walled cells of textura angularis. Hamathecium of dense, 2-3 μm diam (, n = 20), numerous, septate, hyaline, cellularpseudoparaphyses. Asci 100–150 × 20–25 μm ( μm, n = 20), 8-spored, numerous, bitunicate, fissitunicate, broadly-clavate, with a short orbicular pedicel, rounded at apex without an ocular chamber. Ascospores 30–48 × 14–20 μm ( μm, n = 40), obliquely biseriate, broadly fusiform, brown to reddish-brown, 3-transseptate, without longitudinal septa, guttulate, smooth-walled or finely punctate, with a distinct, 4-5 μm wide, mucilaginous sheath. Asexual state: Unknown.
Material Examined. USA, California, Mt. Shasta, ridge south of Horse Camp, elevation 8250 ft, on culms of Trisetum spicatum (L.) Richter, 2 July 1947 W.B. Cooke 20223 (DAOM, holotype).
Shoemaker and Babcock [16] introduced Diadema and characterized the genus by large ascospores without longitudinal septa with a distinct mucilaginous sheath and ascomata with a circular lid-like opening. Currently eight species of Diadema are listed in Index Fungorum [5]. Six species were included when the genus was introduced and another two species (Diadema ahmadii, Kaz. Tanaka & S.H. Iqbal, and Diadema sieversiae (Peck) Huhndorf) were later added [4, 16]. The nature of the ascomata appears to be an important character of this genus and family. Except D. obtusa all other species of Diadema have terete; that is, ascospores are cylindrical, frequently circular in section but narrowing to one end. We observed D. tetramerum, the generic type of Diadema and besides ascomata opening via a circular lid, asci with the short orbicular pedicel without an ocular chamber and trans-septate, ascospores, lacking longitudinal septa, and surrounded by a very broad sheath narrowed to a waist near the middle septum are considered to be significant for the genus.
No molecular data is available for the type or other species of Diadema. Therefore recollection, epitypification, and sequence data is essential to establish family and species relationships.
3.5. Excluded Genera
Clathrospora. Rabenh., Hedwigia 1(18): 116 (1857).
Saprobic on wood and stems. Sexual state: Ascomata semi-immersed, scattered on putrid host stems and foliage, brown to blackish brown, subglobose or nearly globose, with a central sunken ostiole open via a circular lid, asci and pseudoparaphyses forming at the base of the peridium. Peridium composed of 3–5 layers of brown, relatively thick-walled cells of textura angularis, inner cells flattened, thin-walled and lighter. Hamathecium composed of dense, hyaline, filiform, pseudoparaphyses which are longer than the asci. Asci 8-spored, bitunicate, fissitunicate, thick-walled, cylindrical to clavate, with a short pedicle and shallow ocular chamber. Ascospores biseriate, fusiform 7-transseptate, two or many rows of longitudinal septa, muriform, constricted only at the central septum, dark brown to brown, surrounded by a thin, hyaline mucilaginous sheath. Asexual State: Alternaria-like.
Type Species: Clathrospora elynae. Rabenh., Hedwigia 1 : 116 (1857) (see Figure 3).
Figure 3.

Clathrospora elynae (isotype). (a) Herbarium material. (b) Closeup of ascomata. (c) Section of the ascomata. (d) Closeup of the peridium (e) Hyaline, filiform, and pseudoparaphyses. ((f)–(h)) Cylindrical to clavate asci with a short pedicle and ocular chamber. ((i)–(k)) Dark brown to brown muriform ascospores surrounded by a thin, hyaline mucilaginous sheath. Scale bars: (b) = 100 μm, (c) = 10 μm, ((d)–(g)) = 60 μm, and ((h)–(j)) = 30 μm.
Saprobic on wood and stems. Sexual State: Ascomata 140 × 220–145 × 175 μm ( μm, n = 10), semi-immersed, scattered on the putrid host stems and foliage, subglobose or nearly globose, brown to blackish brown, with a central sunken ostiole open via a circular lid, asci and pseudoparaphyses forming on the base of the peridium. Peridium 20–55 μm (, n = 20), composed of 3–5 layers of brown, relatively thick-walled cells of textura angularis, inner cells flattened, thin-walled and lighter. Hamathecium composed of dense, 2-3 μm diam (, n = 20), hyaline, filiform, pseudoparaphyses, longer than the asci. Asci 160–230 × 24–48 μm ( μm, n = 20), 8-spored, bitunicate, fissitunicate, thick-walled, cylindrical to clavate, with a short pedicle and ocular chamber. Ascospores 40–65 × 18–27 μm ( μm, n = 40), biseriate, fusiform, 7-transseptate, two or many rows of longitudinal septa, muriform, constricted only at the central septum, dark brown to brown, surrounded by a thin, hyaline mucilaginous sheath. Asexual State: Alternaria-like.
Material Examined. Switzerland, on the stem of Carex curvula, September 1898, Winter (BPI 627748, isotype).
Shoemaker and Babcock [1] assigned Clathrospora to Diademaceae and included an additional nine species and provided a key to the genus based on the number of septa and length of ascospores. Clathrospora was characterized by circular lid-like opening and applanate, muriform ascospores. Currently, 50 Clathrospora species are listed in the genus in Index Fungorum [5]. Molecular studies based on combine gene analysis showed that two putative strains of Clathrospora, C. elynae (CBS 196.54) and C. diplospora (IMI 68086), were clustered in Pleosporaceae [4, 8]. We obtained similar results in the phylogenetic tree produced from combined nrLSU and nrSSU sequence analysis (Figure 1). Clathrospora elynae the type of Clathrospora formed a separate clade with relatively high bootstrap support (55%) within Pleosporaceae. Based on the phylogenetic result together with the morphological characters (slightly papillate ostiole and Alternaria-like asexual morph) we refer Clathrospora to Pleosporaceae.
Comoclathris. Clem., Gen. fung. (Minneapolis): 37, 173 (1909) ≡ Platyspora Wehm., World Monograph of the Genus Pleospora and its Segregates: 254 (1961).
Habitat saprobic on dead wood or stems. Sexual state: Ascomata semi-immersed to superficial, scattered or aggregated, subglobose or nearly globose, brown to blackish brown coriaceous, ascomata opening via a large circular aperture or lid. Peridium comprising 3-4 layers of brown, relatively thick-walled cells of textura angularis. Hamathecium composed of dense, hyaline, filiform, septate pseudoparaphyses. Asci 8-spored, bitunicate, fissitunicate, cylindrical to cylindro-clavate, with an ocular chamber. Ascospores uniseriate or partially overlapping, fusiform, muriform, brown to reddish-brown, surrounded by a thick, hyaline, mucilaginous sheath. Asexual State: Alternaria-like
Type Species: Comoclathris lanata. Clem. [as “Comochlatris”], Gen. fung. (Minneapolis): 1–227 (1909). MycoBank: MB 209341.
Comoclathris, typified by Comoclathris lanata, was introduced by Clements (1909). The genus is characterized by ascomata with circular lid-like openings and applanate reddish-brown to dark reddish-brown, muriform ascospores, with single longitudinal septa [1]. Zhang et al. [4] tentatively placed Comoclathris in the Pleosporaceae based on Alternaria-like asexual morphs and this was followed by Woudenberg et al. [14]. Comoclathris shares common characters with Pleospora herbarum, the type of Pleospora, in having cylindrical to cylindroclavate asci with an ocular chamber and muriform, brown or pale brown, with or without sheath ascospores. Comoclathris and Pleospora differ in the opening of ascomata (opening via a large circular aperture or lid versus open by a central pore). Comoclathris and Pleoseptum share similar characters in having globose, black, ascomata, and cylindrical to cylindroclavate asci with muriform, yellowish to dark brown ascospores. Comoclathris differs from Pleoseptum in having superficial ascomata with circular lid-like openings composed of comparatively thin peridium and applanate and fusiform ascospores surrounded by a distinct hyaline, mucilaginous thick sheath [3, 4]. In Pleoseptum ascomata are immersed, usually with a papillate apex, with a relatively broad peridium and ovoid to fusoid ascospores [2, 3]. Comoclathris was considered to differ from Clathrospora as in the latter genus species have two or more rows of longitudinal septa as compared with a single row in Comoclathris [3]. Shoemaker and Babcock [1] provided a key to 21 species of Comoclathris. Presently 32 epithets are listed for Comoclathris in Index Fungorum [5]. Molecular data for Comoclathris lanata, the type species of Comoclathris, is not available. Two strains of Comoclathris compressa (CBS 157.53 and CBS 156.53), however, cluster together in a well-supported clade within the family Pleosporaceae [14]. Based on the phylogenetic result coupled with the morphological characters (Alternaria-like asexual morph) we agreed with Zhang et al. [4] and Woudenberg et al. [14] to place Comoclathris in Pleosporaceae. This is, however, based on a species and recollection of the type species is essential to establish the correct placement of the genus.
Diademosa. Shoemaker & C.E. Babc., Can. J. Bot. 70(8): 1641 (1992).
Saprobic on stems and wood. Sexual state: Ascomata immersed, initially erumpent becoming superficial, scattered, depressed-globose, some flattened at the base, opening a disc-like lid of brown prismatic cells with setae. Peridium composed of brown pseudoparenchyma cells of textura angularis. Hamathecium of numerous, dense, septate, hyaline, cellularpseudoparaphyses. Asci 8-spored, bitunicate, fissitunicate, clavate with short narrow pedicel and minute ocular chamber. Ascospores biseriate, partially overlapping, fusiform, straight, frequently circular in section but narrowing to one end, with transverse and vertical septa, pale brown to dark brown, smooth walled. Asexual state: Unknown.
Type Species: Diademosa californiana. (M.E. Barr) Shoemaker & C.E. Babc. [as “californianum”], Can. J. Bot. 70(8): 1641 (1992) ≡ Graphyllium californianum M.E. Barr, Mem. N. Y. bot. Gdn 62: 40 (1990) (see Figure 4).
Figure 4.
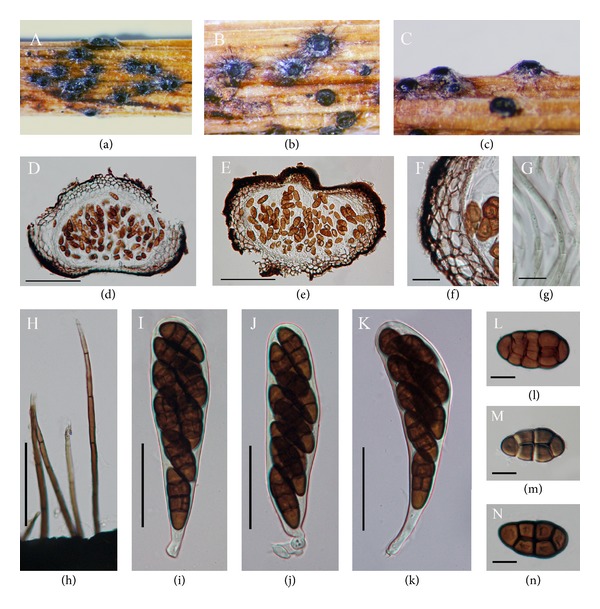
Diademosa californiana (holotype). ((a)-(b)) Ascomata on host substrate. (c) Side view of the ascomata. ((d)-(e)) Section of ascomata. (f) Section of peridium. (g) Septate, hyaline, and cellularpseudoparaphyses. (h) Light to dark brown seta. ((i)–(k)) Ascus with minute pedicel bearing irregularly arranged 8 ascospores. (l)–(n) Ascospores. Scale bars: ((d)-(e)) = 200 μm, ((f)–(h)) = 10 μm, ((i)–(k)) = 50 μm, and (l)–(n) = 10 μm.
Saprobic on stem and wood. Sexual state: Ascomata 200–365 × 240–425 μm ( μm, n = 10), immersed, initially erumpent becoming superficial, scattered, depressed-globose, some flattened at the base, opening a disc-like lid of brown prismatic cells with setae. Peridium 24–59 μm (, n = 20), composed brown pseudoparenchyma cells of textura angularis. Hamathecium of dense, 2-3 μm diam (, n = 20), numerous, septate, hyaline, cellularpseudoparaphyses. Asci 140–175 × 24–28 μm ( μm, n = 20), 8-spored, bitunicate, fissitunicate, clavate with short narrow pedicel and minute ocular chamber. Ascospores 50–65 × 26–32 μm ( μm, n = 40), biseriate or discontinuously arranged, partially overlapping, fusiform, straight, cylindrical; frequently circular in section but narrowing to one end, with transverse and vertical septa, muriform, constricted at first septum, pale brown to dark brown, smooth walled. Asexual state: Unknown.
Material Examined. USA, Bump-Cold Boiling Lake Trail, Lassen Volcanic National Park, Shasta, California, on branch of Wyethia, 12 July 1966, W.B. Cooke & D.L. Hawksworth. (NY, holotype).
Diademosa was established by Shoemaker and Babcock [1] and typified by D. californiana, based on the ascoma opening via a circular lid and ascospores being frequently circular in section, but narrowing to one end. Diademosa californiana was initially introduced as Graphyllium californianum by Barr [17] and referred to Hysteriaceae based on the pore or slit like opening. Reexamination of the type specimens by Shoemaker and Babcock [1] concluded that Diademosa opened by a flat lid similar to Diadema and assigned it into Diademaceae. The lid is hard to observe in sections unless they are mounted directly in lactic acid because excessive swelling occurs in water [1]. Diademosa differs from Comoclathris in having cylindrical, frequently circular in section, but narrowing to one end ascospores compared with flattened ascospores of Comoclathris. Diademosa and the generic type of Pleosporaceae, Pleospora share common characters. Both Diademosa and Pleospora comprise narrowly oblong ascomata with cellular pseudoparaphyses and cylindrical to clavate asci with muriform, brown or pale brown ascospores. However, Diademosa differs from Pleospora in having an ascomata opening via a circular lid, covered with setae and asci with short narrow pedicel, while Pleospora species have ascomata opening by a central pore without setae and asci with a short, thick, furcated pedicel. Except the ascomata opening via disc-like lid, Diademosa resembles some characters of Pyrenophora. That is, both Diademosa and Pyrenophora have superficial ascomata with setae and muriform, smooth-walled, light brown to dark brown ascospores. Currently four species of Diademosa are listed in Index Fungorum [5], but no molecular data is available for the genus. We place Diademosa in Pleosporaceae because of its similarities with other genera in this family, but confirmation of the phylogenetic status of this genus depends on recollecting the fungus and epitypification with molecular sequences.
Graphyllium. Clem., Botanical Survey of Nebraska 5: 6 (1901).
Habitat saprobic on woody stems. Sexual state: Ascomata semi-immersed, hysteriform, black to brown, subglobose to ovoid. Peridium comprising 2-3 layers of brown, relatively thick cells of textura angularis, inner cells flattened, thin-walled and lighter. Asci 8 spored, bitunicate, fissitunicate, clavate. Ascospores biseritate overlapping, muriform, applanate, obpyriform, straight, with 3-4 transverse septa, 1–2 longitudinal septa or no longitudinal septa, brown to olive green. Asexual state: Unknown.
Type Species: Graphyllium chloës. Clem., Bot. Surv. Nebraska 5: 6 (1901) ≡ Pleospora chloës (Clem.) Petr., Sydowia 6(5-6): 337 (1952).
Initially Graphyllium was placed in the Hypodermiaceae by Clémencet (1901) and described as “Hysterothecium innate, then erumpent, linear, simple, membranaceous-plectenchymatous, black; asci ovoid or cylindrical-clavate, 8-spored; spores brown, elliptical to oblong, with transverse-and longitudinal septa, but not muriform; pseudoparaphyses simple or branched, septate, forming an epithecium." Later Barr [17] transferred the genus to order Pleosporales and referred to Phaeosphaeriaceae. Platyspora was considered as a synonym of Graphyllium [4]. Shoemaker and Babcock [1] assigned Graphyllium to Hysteriaceae considering the ascomatal characters along with applanate ascospores that are at least 3-septate in side view and have some longitudinal septa in front view. Lumbsch and Huhndorf [6] included Graphyllium in the family Diademaceae, but Zhang et al. [4] referred to Hysteriaceae. We examined the generic type of Graphyllium, G. chloës we also agreed to refer Graphyllium tentatively in Hysteriaceae because of its hysterothecium-like ascomata forming a longitudinal opening which is clearly deviated from the lid-like opening in Diademaceae. However the correct placement of this taxon still depends on epitypification with molecular data.
4. Concluding Remarks
The importance of molecular data in determining the importance of morphological characters and relationship of microfungi cannot be overstressed and has proved significant at establishing genus and species relationships [18, 19] and resolving cryptic species in important plant pathogenic genera, for example, Diaporthe [20] and Pestalotiopsis [21]. Shoemaker and Babcock [1] introduced Diademaceae which they considered to be a distinctive family based on the ascomata opening via a flat, circular lid and comprising Clathrospora, Comoclathris, Diadema, Diademosa, and Macrospora [1]. Recent studies based on molecular phylogeny [4, 14], including this study, conclude that Clathrospora and Comoclathris clustered within Pleosporaceae. Molecular data, however, is not available for Diadema and Diademosa. Graphyllium is placed in Hysteriaceae because of its hysterothecium-like ascomata with a slit like opening; this clearly diverges from the lid like opening in Diademaceae. The characteristic feature of ascomata opening via a flat circular lid is considered as an adaptation to the alpine habitat [16]. It is, however, doubtful if this character is significant and whether Diademaceae is a separate family in the order Pleosporales. Until further molecular data becomes available we maintain Diademaceae with a single genus Diadema based on its large transseptate ascospores surrounded by a distinct mucilage sheath and ascomata with a circular, lid-like opening [16].
Acknowledgments
We are grateful to the Mushroom Research Foundation, Chiang Rai, Thailand, for supporting this research. MFLU Grant no. 56101020032 is thanked for supporting studies on Dothideomycetes. Hiran A. Ariyawansa and Ji Chuan Kang are grateful to the International collaboration plan of Science and Technology at Guizhou Province (Contract no. [2012] 7006) and the construction of innovation talent team of Science and Technology at Guizhou Province (Contract no. [2012] 4007). Hiran Ariyawansa is grateful to A. D Ariyawansa and D. M. K Ariyawansa for their valuable suggestions.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of the paper.
References
- 1.Shoemaker RA, Babcock CE. Applanodictyosporous pleosporales: Clathrospora, Comoclathris, Graphyllium, Macrospora, and Platysporoides . Canadian Journal of Botany. 1992;70(8):1617–1618. [Google Scholar]
- 2.Hyde KD, Jones EBG, Liu JK, Ariyawansa H, et al. Families of dothideomycetes. Fungal Diversity. 2013;63:1–313. [Google Scholar]
- 3.Zhang Y, Crous PW, Schoch CL, Hyde KD. Pleosporales. Fungal Diversity. 2012;53:1–221. doi: 10.1007/s13225-011-0117-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Zhang Y, Koko TW, Hyde KD. Towards a monograph of Dothideomycetes: studies on Diademaceae . Cryptogamie Mycologie. 2011;32(2):115–126. [Google Scholar]
- 5. http://www.indexfungorum.org/Names/Names.asp.
- 6.Lumbsch HT, Huhndorf SM. Outline of Ascomata—2009. Myconet. 2010;14:1–64. [Google Scholar]
- 7.Ariyawansa HA, Maharachchikumbura SSN, Karunarathne SC, et al. Deniquelata barringtoniae gen. et sp. nov., associated with leaf spots of Barringtonia asiatica . Phytotaxa. 2013;105(1):11–20. [Google Scholar]
- 8.Schoch CL, Crous PW, Groenewald JZ, et al. A class-wide phylogenetic assessment of Dothideomycetes . Studies in Mycology. 2009;64:1–15. doi: 10.3114/sim.2009.64.01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hall TA. (Nucleic Acids Symposium).BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. 1999;41 [Google Scholar]
- 10.Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The CLUSTAL X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research. 1997;25(24):4876–4882. doi: 10.1093/nar/25.24.4876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Stamatakis A. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. 2006;22(21):2688–2690. doi: 10.1093/bioinformatics/btl446. [DOI] [PubMed] [Google Scholar]
- 12.Silvestro D, Michalak I. raxmlGUI: a graphical front-end for RAxML. 2012, http://sourceforge.net/projects/raxmlgui/.
- 13.Pattengale ND, Alipour M, Bininda-Emonds ORP, Moret BME, Stamatakis A. How many bootstrap replicates are necessary? (Lecture Notes in Computer Science).Research in Computational Molecular Biology. 2009;5541:184–200. doi: 10.1089/cmb.2009.0179. [DOI] [PubMed] [Google Scholar]
- 14.Woudenberg JHC, Groenewald JZ, Binder M, Crous PW. Alternaria redefined. Mycology. 2013;75:171–212. doi: 10.3114/sim0015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Dong J, Chen W, Crane JL. Phylogenetic studies of the Leptosphaeriaceae, Pleosporaceae and some other Loculoascomycetes based on nuclear ribosomal DNA sequences. Mycological Research. 1998;102:151–156. [Google Scholar]
- 16.Shoemaker RA, Babcock CE. Diadema. Canadian Journal of Botany. 1989;67(5, article 1349) [Google Scholar]
- 17.Barr ME. Prodromus to Class Loculoascomycetes. Amherst, Mass, USA: Hamilton I. Newell; 1987. [Google Scholar]
- 18.Dai D, Bhat DJ, Liu JK, Chukeatirote E, Zhao R, Hyde KD. Bambusicola, a new genus from bamboo with asexual and sexual morphs. Cryptogami Mycologie. 2012;33(3):363–379. [Google Scholar]
- 19.Zhang H, Hyde KD, Mckenzie EHC, Bahkali AH, Zhou D. Sequence data reveals phylogenetic affinities of Acrocalymma aquatica sp. nov., Aquasubmersa mircensis gen. et sp. nov. and Clohesyomyces aquaticus (freshwater coelomycetes) Cryptogamie Mycologie. 2012;33(3):333–346. [Google Scholar]
- 20.Udayanga D, Liu X, McKenzie EHC, Chukeatirote E, Bahkali AHA, Hyde KD. The genus Phomopsis: biology, applications, species concepts and names of common phytopathogens. Fungal Diversity. 2011;50:189–225. [Google Scholar]
- 21.Maharachchikumbura SSN, Guo LD, Cai L, et al. A multi-locus backbone tree for Pestalotiopsis, with a polyphasic characterization of 14 new species. Fungal Diversity. 2012;56:95–129. [Google Scholar]