

Abstract
In a recent paper,1 we examined how experience of repeatedly flying with a specific partner influences pigeons’ subsequent navigational decision-making in larger flocks. We found that pairs develop into a “behavioral unit” through their shared experience of joint flights, acquiring a single idiosyncratic route during training, and then forming spatially distinct subgroups when flying with other pairs. Further, differences between the route preferences of different pairs appear to be reconciled through the same mechanisms as those that apply to individuals. Here we examine in more detail the development of route preferences in pairs, as an example of “collective learning.” We find that pairs acquire routes more quickly, but with less precision, than individuals. We use these results to hypothesize on the advantages and limitations of solving problems collectively.
Keywords: collective learning, route recapitulation, homing, social navigation, Columba livia
Many animals travel in groups and derive a range of benefits from the presence of their conspecifics.2 For example, group travel can have anti-predatory advantages by increasing the probability of detecting attackers,3 and can improve navigation by increasing the accuracy of orientation.4 Homing pigeons (Columba livia) exhibit a strong tendency to travel in flocks, and GPS tracking studies have confirmed that groups, from pairs to larger flocks, indeed home more efficiently than individuals.5,6 In addition, while individual pigeons acquire idiosyncratic and faithfully recapitulated routes through repeated flights from a given release site,7 pairs of birds with conflicting route preferences tend to prefer to remain together, and resolve differences either through compromising over route choice or through one bird assuming the lead.5 Interestingly, less experienced pigeons tend to follow the path of those more experienced, especially when their own knowledge is low,8 and initially naïve birds are able to learn homing routes from knowledgeable partners through repeated joint flights.9 However, one aspect of such co-navigation by pairs that remains open concerns what happens if both individuals are naïve in terms of local navigational experience. In essence, what happens when neither bird has prior knowledge of a specific navigational problem, and they have to solve it collectively? Does the presence of another individual during learning help or hinder the development of idiosyncratic routes?
The training phase of our study1 provides data with which we can begin to answer this question. The experimental birds were between 3 and 7 y old, and had homing experience but had never been released from the sites used in the present study. We used a mixture of males and females, but none of the birds was raising chicks at the time of the experiment, reducing the likelihood that motivational differences due to breeding state would have influenced results. We trained our 16 subjects by releasing them 17 times in succession from 2 different release sites. At one site all birds were released singly, and at the other they were always released in pairs (with each bird always paired with the same partner). This allowed us to compare the development of homing routes in the same birds under 2 conditions. As described in our paper, we found that pairs developed idiosyncratic homing routes; hence, flying with a partner did not inhibit the establishment of faithfully recapitulated routes (Fig. 1). We also showed that the general course of route development in pairs closely resembled what we observed in individuals. However, one interesting aspect of our results that deserves further attention is that pairs seemed to profit from joint flights by acquiring route knowledge faster. During both solo and pairwise training, individual homing efficiency increased as training progressed (nonlinear regression, Solo flights: r2 = 0.62, Paired flights: r2 = 0.47, Fig. 2A), but the corresponding regression curves differed significantly (F-test, F(2,540) = 3.34, P = 0.036). Also, there was a significant difference between the 2 training protocols at the beginning of training (2-way repeated measures ANOVA, F(1,255) = 4.46, P = 0.036, Fig. 2A), when the rate of efficiency change was at its maximum level, with the birds undergoing pairwise training showing a higher level of efficiency already during their second training flight (Bonferroni post hoc test, P < 0.001).

Figure 1. Examples of the last 3 of 17 training flights performed by homing pigeons trained (A and C) solo and (B and D) pairwise. Panels A and B and panels C and D show data from the same individual. Solo-training was conducted from the College Farm release site (direction and distance to home: 74°, 7.0 km); pair-training from Church Hanborough (direction and distance to home: 129°, 6.14 km). In both panels the release point is indicated by a white dot, the home loft by a gray dot. Blue grid is 1 × 1 km. Tracks were recorded using miniature GPS logging devices.
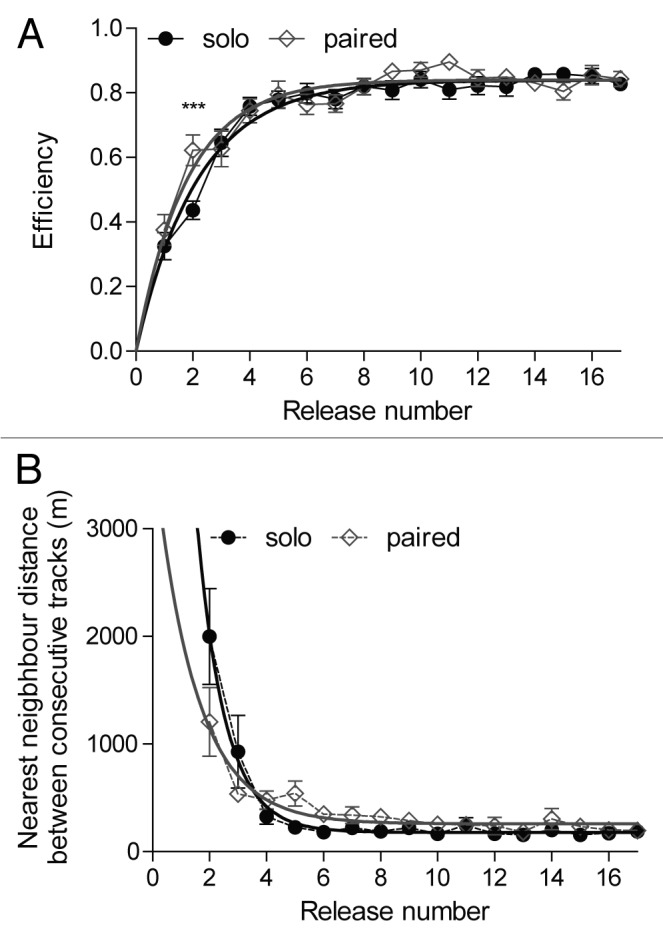
Figure 2. Training data from homing pigeons released 17 times consecutively from the same release site, either singly or in pairs. (A) Homing efficiency (mean ± S.E.M.) and (B) nearest neighbor distance (mean ± S.E.M.) between consecutive tracks as a function of training progression. Black circles indicate group averages for solo training; white diamonds correspond to paired training. Solid lines show nonlinear regression curves fitted to the data (see main text). Asterisks represent significant differences (Bonferroni post hoc test, P < 0.001).
Next, we used nearest neighbor analysis to evaluate how the precision with which individuals recapitulated their idiosyncratic routes changed as training progressed. We measured the similarity between consecutive tracks flown by a given bird by computing the spatial proximity between these tracks. For each point along the focal track, we determined the distance to the nearest neighboring point along the previous (comparison) track, and used the average distance across all pairs of points as a measure of track similarity (where lower values correspond to higher similarity, i.e., lower variation). During both individual and paired training, track variation decreased with training (nonlinear regression, Solo flights: r2 = 0.23, Paired flights: r2 = 0.13, Fig. 2B), with significantly different curves between the 2 conditions (F-test, F(2,506) = 8.9, P < 0.001).
Hence, pairwise trained homing pigeons were not only able to develop joint stereotyped homing routes, but their acquisition was also characterized by a faster increase in homing efficiency than that observed in individually established routes. Similarly, at an early stage of route development, birds exhibited less variation between consecutive tracks when trained with a partner than when trained solo. However, by the end of training, birds recapitulated routes slightly less accurately after learning in pairs than after learning individually, suggesting that the initial advantage of faster learning in pairs is offset by less accurate acquisition (or less accurate execution) of the task. Nonetheless, an important caveat to consider is that in order to be able to use the same birds in both the pair- and solo-training conditions, we necessarily had to train them from 2 different sites for our 2 treatments, making it essential to conduct further tests to rule out the effect of release site on the group differences in the speed and accuracy of learning we detected. One approach would be to train 2 different groups of subjects from the same release site, with one group trained individually and the other in pairs. In that case, the disadvantage of not being able to conduct within-subject analyses may be outweighed by the benefit of removing any potential release site effect.
Two additional, interesting questions concern the relative contributions of the 2 birds of a pair to the problem-solving process, and the relative degrees of retention they each achieve. At one extreme, it may be that pairs develop routes because both birds contribute to route choice during early flights and both acquire route memories as a result of their experiences (which we would call “collective learning”). At the other extreme, it may be that one bird (a leader) is responsible for making navigational choices throughout, and only it, but not the follower, acquires a memory of the route. The latter possibility is unlikely to be the case (at least in such an extreme sense): we know that birds that follow others can learn from the experience.9 Nonetheless, what indeed happens during problem-solving and learning in pairs has implications for how we interpret what we observed in the test phase of our study,1 where we released trained pairs together with other pairs. Did the routes taken by those flocks of 4 birds represent the outcome of 4 birds reconciling conflict over route choice, or effectively just of the 2 leaders, each carrying with it its own follower? Our future experiments will examine this issue by testing pair-trained birds individually immediately after their last paired flight, to ascertain their respective levels of acquisition achieved during the collective learning task. We will then be able to use these data to refine our understanding of decision-making outcomes in larger flocks.
We hypothesize that the faster increase in efficiency we observed in pair-trained pigeons (if confirmed by further experiments that rule out a release site confound) might derive from a “many wrongs”-like effect,10 where partners’ different navigational skills, experiences, and individual errors are averaged out to give a better estimate of the home direction than what single birds are able to achieve. Each bird may also memorize different parts of the route more or less accurately, and this may allow them to rely on each other’s distinct experiences, pool complementary pieces of information, and thereby settle on routes faster. Alternatively, higher rates of increase in efficiency may also arise from a reduced need for vigilance in groups.11 Birds with partners need to attend to a smaller proportion of their immediate surroundings, which in turn may facilitate landmark recognition and retention. On the other hand, pairwise trained birds must also divide their attention between 2 processes: 1) maintaining cohesion with their partner by responding to its movements, and 2) establishing a route by memorizing landmarks. Such a trade-off may explain the fact that birds trained in pairs exhibit more track variability in the final stages of training than solo-trained individuals, pointing to both benefits and limitations to learning and executing tasks in groups. In summary, our initial work has opened up several interesting questions and alternative interpretations, which we hope to resolve by further experimentation to better understand the phenomenon of collective problem-solving.
Acknowledgments
We thank Julian Howe for technical assistance. Flack A was supported by Microsoft Research, Cambridge, and Biro D by a Royal Society University Research Fellowship.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Footnotes
Previously published online: www.landesbioscience.com/journals/cib/article/26521
References
- 1.Flack A, Freeman R, Guilford T, Biro D. Pairs of pigeons act as behavioural units during route learning and co-navigational leadership conflicts. J Exp Biol. 2013;216:1434–8. doi: 10.1242/jeb.082800. [DOI] [PubMed] [Google Scholar]
- 2.Krause J, Ruxton G. Living in Groups. USA: Oxford University Press, 2002. [Google Scholar]
- 3.Elgar MA. Predator vigilance and group size in mammals and birds: a critical review of the empirical evidence. Biol Rev Camb Philos Soc. 1989;64:13–33. doi: 10.1111/j.1469-185X.1989.tb00636.x. [DOI] [PubMed] [Google Scholar]
- 4.Bergman G, Donner KO. An analysis of the spring migration of the common scoter and the long-tailed duck in southern Finland. Acta Zool Fenn. 1964;105:1–59. [Google Scholar]
- 5.Biro D, Sumpter DJT, Meade J, Guilford T. From compromise to leadership in pigeon homing. Curr Biol. 2006;16:2123–8. doi: 10.1016/j.cub.2006.08.087. [DOI] [PubMed] [Google Scholar]
- 6.Dell’Ariccia G, Dell’Omo G, Wolfer DP, Lipp H-P. Flock flying improves pigeons’ homing: GPS track analysis of individual flyers versus small groups. Anim Behav. 2008;76:1165–72. doi: 10.1016/j.anbehav.2008.05.022. [DOI] [Google Scholar]
- 7.Meade J, Biro D, Guilford T. Homing pigeons develop local route stereotypy. Proc Biol Sci. 2005;272:17–23. doi: 10.1098/rspb.2004.2873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Flack A, Pettit B, Freeman R, Guilford T, Biro D. What are leaders made of? The role of individual experience in determining leader–follower relations in homing pigeons. Anim Behav. 2012;83:703–9. doi: 10.1016/j.anbehav.2011.12.018. [DOI] [Google Scholar]
- 9.Pettit B, Flack A, Freeman R, Guilford T, Biro D. Not just passengers: pigeons, Columba livia, can learn homing routes while flying with a more experienced conspecific. Proc Biol Sci. 2013;280:20122160. doi: 10.1098/rspb.2012.2160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Simons AM. Many wrongs: the advantage of group navigation. Trends Ecol Evol. 2004;19:453–5. doi: 10.1016/j.tree.2004.07.001. [DOI] [PubMed] [Google Scholar]
- 11.Jackson AL, Ruxton GD. Toward an individual-level understanding of vigilance: the role of social information. Behav Ecol. 2006;17:532–8. doi: 10.1093/beheco/arj060. [DOI] [Google Scholar]