

Abstract
The migration of endothelial cells (ECs) plays an important role in vascular remodeling and regeneration. ECs are constantly subjected to shear stress resulting from blood flow and are able to convert mechanical stimuli into intracellular signals that affect cellular behaviors and functions. The aim of this study is to elucidate the effects of Rac1, which is the member of small G protein family, on EC migration under different laminar shear stress (5.56, 10.02, and 15.27 dyn/cm2). The cell migration distance under laminar shear stress increased significantly than that under the static culture condition. Especially, under relative high shear stress (15.27 dyn/cm2) there was a higher difference at 8 h (P < 0.01) and 2 h (P < 0.05) compared with static controls. RT-PCR results further showed increasing mRNA expression of Rac1 in ECs exposed to laminar shear stress than that exposed to static culture. Using plasmids encoding the wild-type (WT), an activated mutant (Q61L), and a dominant-negative mutant (T17N), plasmids encoding Rac1 were transfected into EA.hy 926 cells. The average net migration distance of Rac1Q61L group increased significantly, while Rac1T17N group decreased significantly in comparison with the static controls. These results indicated that Rac1 mediated shear stress-induced EC migration. Our findings conduce to elucidate the molecular mechanisms of EC migration induced by shear stress, which is expected to understand the pathophysiological basis of wound healing in health and diseases.
Keywords: Rac1, shear stress, endothelial cell migration
Introduction
Vascular endothelial cell (EC) migration is an essential process in angiogenesis, wound healing, vessel remodeling, and reendothelialization. During physiological and pathologic courses, many effectors, such as growth factors, chemokines, and mechanical stimulation, can regulate the migration of vascular ECs through signaling transduction pathway. The intracellular signaling pathways mediating cell migration have to respond to diverse extracellular cues and translate them into finely regulated cellular activities.1-3
ECs lining the interior walls of blood vessels alter the expression of “stress-sensitive” genes in response to shear flow in blood.4 There is considerable evidence that fluid shear stress functions as a significant regulatory factor in regulating EC migration through mechano-chemical transduction.5-9 Studies on intracellular signaling events have shown that shear stress activated multiple signaling molecules, including protein kinase C, focal adhesion kinases (FAK), Rho family GTPases, phosphatidylinositol 3-kinase (PI3K), and mitogen-activated protein kinases (MAPKs).10,11 Thus, shear stress can activate multiple mechano-sensing molecules and lead to initiation and propagation of signals through networks of pathways.12 These molecules lead to the activation of various signaling cascades, contributing to a highly coordinated and orchestrated mechano-chemical transduction signaling. The activated signal transduction pathway is not simply linear, but involves in cross-talk, feedback, and bidirectional communication to form a signaling network.13 Rho GTPases act as molecular switches to control signal transduction pathways by cycling between an active GTP-bound form and an inactive GDP-bound form. In the GTP-bound form, they interact with downstream targets to elicit a variety of intracellular responses.12,14 Rho family small GTPases belong to the Ras superfamily, including Cdc42, Rac, and Rho, playing important roles in regulating the actin-based cytoskeletal structure and intracellular signaling. The current studies showed that Rac1 was required at the front of the migrating cell to regulate actin polymerization to form lamellipodial and membrane protrusion.15,16 In addition, the activation of Rac1 can activate RhoA to affect the formation of stress fibers and cell contraction, thereby regulate EC migration,17 which plays an important role in vascular endothelial growth factor-induced EC migration.18,19 In addition, shear stress has been proven to be an important modulator of cellular function in cell migration process.9,20 However, most experiments in previous research were performed under a certain shear stress.8 There is little study that compares the role of Rac1 in EC migration under different levels of shear stress.
Our previous studies suggested that Rac1 induced by interleukin-8 (IL-8) could mediate the migration of ECs,21 and CXCR1/2 as mechano-sensors could mediate laminar shear stress-induced endothelial cell migration.22 In this study, we further examine whether Rac1 mediates the migration of ECs induced by shear stress and elucidate the underlying molecular mechanisms, i.e., (1) three levels of shear stress were applied to investigate EC migration compared with static condition. (2) EA.hy 926 cells transfected with cDNA encoding the WT of Rac1 (Rac1WT), a dominant active mutant of Rac1 (Rac1Q61L), and a dominant negative mutant of Rac1 (Rac1T17N) were subjected to three levels of shear stress to evaluate the role of Rac1 in shear stress-induced EC migration. Elucidation of the molecular mechanisms of Rac1 mediating EC migration induced by shear stress would help us to understand the pathophysiological basis of wound healing in health and diseases, and to provide new ideas for the diagnosis and prevention of cardiovascular diseases and malignant tumors.
Results
Shear stress promoted EC migration
Actually, vascular ECs in vivo are under the influence of various flow conditions. To compare the effects of different shear stress on EC migration, a strip of confluent EA.hy 926 cells were denudated and subjected to shear stress of 5.56, 10.02, and 15.27 dyn/cm2 (static culture condition as controls), respectively. The direction of the flow was from left to right (black arrow in Fig. 1A). The left-hand side of the denuded zone was denoted as “upstream” side, correspondingly, the right-hand side denoted as “downstream.” Figure 1A showed that on the upstream side, the direction of cell migrating into the denuded zone was parallel to the direction of shear stress. The area of wound healed gradually with the duration due to EC migration. The average net migration distance at the upstream edge under each shear stress was shown in Figure 1B. It can be seen from Figure 1 that the cell migration distance under relative high shear stress (15.27 dyn/cm2) increased significantly than that under the static culture condition at each duration. Especially, there was a highly significant difference at 8 h compared with static controls (P < 0.01). Under shear stress of 5.56 dyn/cm2 and 10.02 dyn/cm2, the cell migration distance increased significantly than that of static control at 4 h, 6 h, and 8 h (P < 0.05). However, there was no significant difference at initial 2 h. Correspondingly, under static condition, the average net migration distance of the cells at the upstream edge increased slowly with duration (from 23.92 ± 1.63 µm at 2 h to 38.44 ± 5.64 µm at 8 h). Remarkably, under shear stress of 5.56 dyn/cm2, 10.02 dyn/cm2, and 15.27 dyn/cm2 at 8 h, the average net migration distance of the cells at the upstream edge increased to 113.62 ± 6.06 µm, 145.31 ± 8.74 µm, and 193.13 ± 9.82 µm, respectively, compared with 38.44 ± 5.64 µm under static condition. These results suggested that EC migration was strongly depended on the magnitude of shear stress. In addition, cell migration during wound repair involves displacements both parallel and perpendicular to the direction of flow. Cell migration parallel to flow is expected to be the primary contributor to wound repair. Therefore, wound repair area ratio (as defined in Materials and Methods), in combination with migration distance, as another new parameter, was examined to confirm how flow impacts on the directionality of cell migration. The wound repair area of each group showed an increased trend with enhanced shear stress and duration (Fig. 1C), which was similar to results of migrated distance. However, repair area of groups showed a significant difference at initial 2 h compared with controls (P < 0.05). These results all demonstrated that shear stress could promote ECs migration.

Figure 1. Effects of different shear stress on EC migration. (A) The EC monolayers were kept under static conditions, or exposed to three levels of shear stress. The direction of flow was from left to right. (B and C) The net migration distance of upstream side and wound repair area increased with shear stress and duration. (n = 3, *P < 0.05, **P < 0.01 vs. static).
Shear stress-induced vascular endothelial cells migration is dependent upon upregulation of Ra1 expression
Figure 2A showed the mRNA expression of Rac1 assessed by RT-PCR under three levels of shear stress at different durations. A trend toward increasing mRNA expression of Rac1 with shear stress and duration was observed. Under low shear stress (5.56 dyn/cm2), the mRNA expression of Rac1 increased significantly only at 8 h in comparison with static controls (P < 0.05); while there are significant differences of Rac1 mRNA expression at initial 2 h under mid (10.02 dyn/cm2) and high shear stress (15.27 dyn/cm2) compared with control. It can be seen that the mRNA expression of Rac1 in EA.hy 926 cells under the different shear stress upregulated with increased duration. In addition, the mRNA expression of Rac1 also showed a time-dependent manner under shear stress, and increased significantly with duration. Correspondingly, the mRNA expression of Rac1 in static cultured EA.hy 926 cells increased slowly with duration. Furthermore, the Rac1 protein expression increased with loaded duration of laminar shear stress (under the 10.02 dyn/cm2 shear stress, from 2 h to 8 h). There were significant differences of Rac1 protein expression from 4 h to 8 h compared with static controls and 2 h. Therefore, the results indicated that shear stress-induced vascular endothelial cells migration is dependent upon upregulation of mRNA and protein expression of Rac1, which is consistent with previous results of ECs migration under different shear stress.
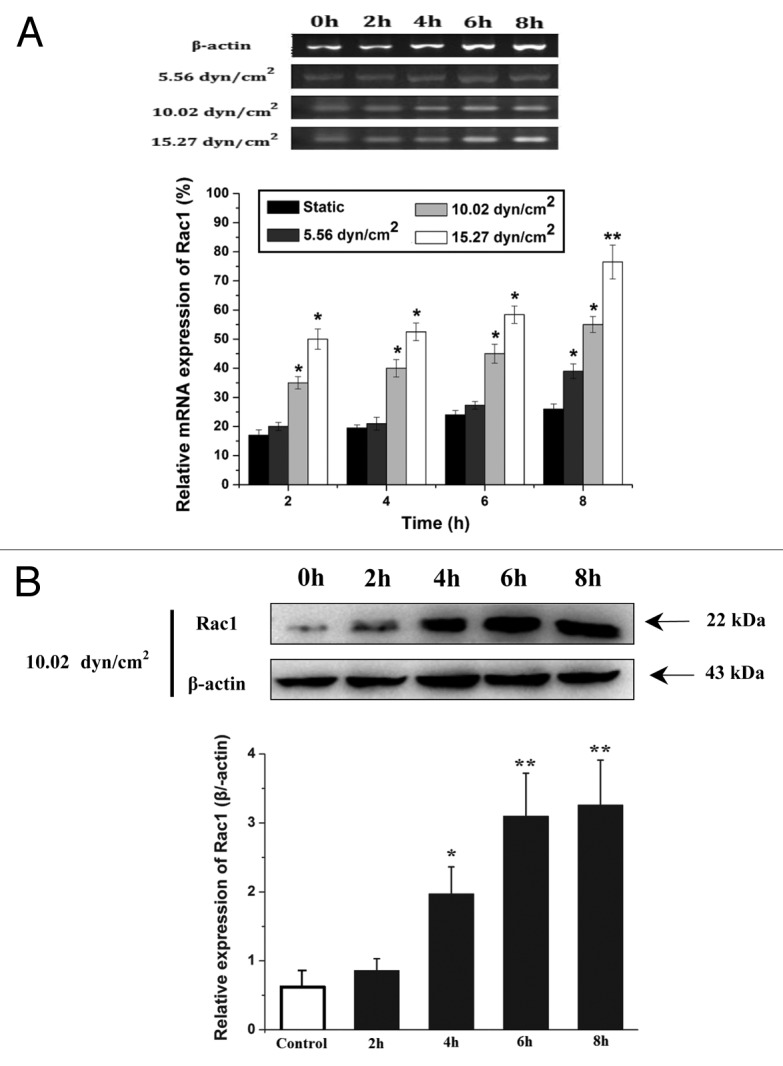
Figure 2. Effects of different shear stress on the mRNA and protein expression of Rac1 in ECs. (A) The expression of Rac1 mRNA under different shear conditions for various time were semi-quantitatively detected by RT-PCR. The electrophoresis of Rac1 mRNA expression in endothelial cells when exposed to different shear stress, β-actin was used as an endogenous control. Relative quantitative values according to images analysis by Image J. (B) Representative western blots of Rac1 expression and quantification based on image analysis of the western blot bands. The expression level of β-actin in each group was used as intrinsic controls, and relative expression of Rac1 were calculated. *P < 0.05, **P < 0.01 vs. static control.
EA.hy 926 cells migration with transfection
To further determine whether Rac1 is involved in shear stress-induced EC migration, EA.hy 926 cells transfected with respective pcDNA3-EGFP-Rac1-WT (a wild-type), pcDNA3-EGFP-Rac1-Q61L (an activated mutant), and pcDNA3-EGFP- Rac1-T17N (a dominant negative mutant) were subjected to three levels of shear stress for 8 h compared with untransfected controls. EA.hy 926 cells transfected with three plasmids were treated with or without mechanical stimulus and then fluorescence microscope was used to detect the expression of Rac1 (Fig. 3A), indicated that three plasmids had been successfully transfected into EA.hy 926 cells with transfection efficiencies were 93.2% (WT), 91% (17N), and 89.7% (Q61), respectively. The fluorescence intensities in three transfected cells were enhanced with stimulation of low shear stress (5.56 dyn/cm2) for 2 h (Fig. 3B). Figure 4B and C indicated the difference of average net migration distance and wound repair area ratio among the transfected cells and untransfected controls. Under shear stress of 5.56 dyn/cm2, both the migration distance and repair area ratio of Rac1Q61L group increased significantly at every duration, and those of Rac1T17N group decreased significantly compared with control group at 4 h, 6 h, and 8 h due to inhibition of Rac1 activities. Similarly, both the migration distance and repair area ratio of Rac1Q61L group increased significantly at 4 h, 6 h, and 8 h under shear stress of 10.02 dyn/cm2, and those of Rac1T17N group decreased significantly at 4 h, 6 h, and 8 h compared with control group. However, all groups had no significant difference at 2 h. In addition, under shear stress of 15.27 dyn/cm2, the migration distance of Rac1Q61L group increased significantly at every duration compared with control group, and Rac1T17N group decreased significantly at 4 h, 6 h, and 8 h. The repair area ratio of Rac1Q61L group increased significantly at 2 h, 4 h, and 6 h compared with control group, but there was no statistic difference at 8 h. The repair area ratio of Rac1T17N group decreased significantly at 4 h, 6 h, and 8 h. However, in Figure 4, under different shear stress, EC migration distance increased in EGFP-Rac1 Q61L transfected group, and decreased in EGFP-Rac1 T17N transfected group, but not changed in the Rac1 WT transfected group, suggesting that rac1 does not mediate the shear stress-induced EC migration by increasing the protein level, but by activated Rac1. Therefore, we speculated that the possible mechanisms through which activated Rac1 GTPase mediates the EC migration but not only total Rac1. It needs to be further examined to elucidate the exact mechanisms of Rac1 activity in regulating cell migrated behavior by shear stress.
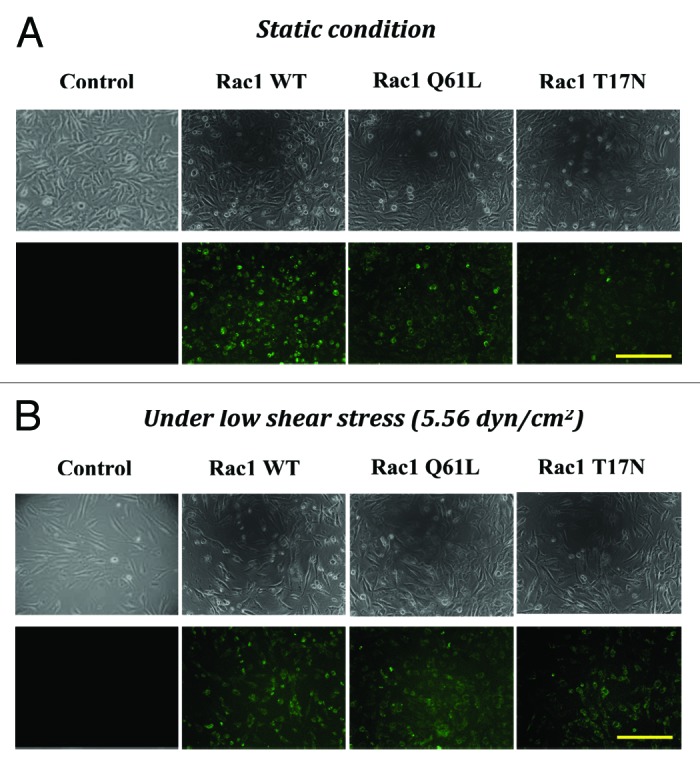
Figure 3. The microphotograph and fluorescent images of EA.hy cell with or without transfecting. The untransfected cells and those transfected with EGFP-Rac1 WT, EGFP-Rac1 T17N, and EGFP-Rac1 Q61L were kept under static condition (A) or subjected to low shear stress (5.56 dyn/cm2) for 2 h (B) (100 × ). Fluorescence microscope showed the expression of Rac1 among EA.hy 926 cells transfected with EGFP-Rac1 WT, EGFP-Rac1 T17N, and EGFP-Rac1 Q61L. Scale bar = 100 μm.

Figure 4. Effects of different shear stress on the migration of transfected ECs. EA.hy926 cells were transfected with pcDNA3-EGFP-Rac1-WT, pcDNA3-EGFP-Rac1-T17N, and pcDNA3-EGFP- Rac1-Q61L, and cell migration under static and flow conditions were monitored for 8 h. The direction of flow is from left to right. (A) The distributed images of EC migration in control and each experimental group at 0 h, 4 h, and 8 h, under three levels of shear stress. (B and C) Compared the difference of cell migration distance (B) and wound repair area (C) among Rac1WT, Rac1T17N, Rac1Q61L, and control group under low, mid, and high shear stress at 2, 4, 6, and 8 h. (n = 3, *P < 0.05 vs. control).
Compared with control group under each level of shear stress, cells transfected with pcDNA3-EGFP-Rac1-Q61L promoted cell migration, and cells transfected with pcDNA3-EGFP-Rac1-T17N inhibited cell migration. These results further demonstrated the prominent role of Rac1 in cell migration, which is regulated by the degree of shear stress directly.
Discussion
Cell migration is an essential process for normal development and homeostasis that can also contribute to important pathologies. It is well established that shear stress can modulate EC morphology and cytoskeleton organization, while the relations of these changes to lamellipodial protrusion, cell migration, and intracellular signaling remain to be established.8 The process of EC migration is the cell directional motility in response to extracellular cues. EC migration is a very complex process, which is affected by many factors, including chemical factors and mechanical factors. Generally, these factors can promote cell movement and trigger intracellular signals that result in spatially and temporally coordinated reorganization of cytoskeletal and development of transient and definitive extracellular matrix contacts.23-25
Mechanical factors play an important role in EC migration. Previous studies demonstrated that fluid shear stress could regulate EC structure and function through mechano-chemical transduction.6-9 It is found that laminar flow had more important roles in promoting EC migration than disturbed flow.26,27 Most experiments in previous research were performed under a certain shear stress. However, vascular ECs in vivo are under the influence of various flow conditions. Our results suggested that the migration distance of EA.hy 926 cells increased with shear stress and duration, and the flow could regulate the direction and speed of migration. The reasons why flow could promote cell migration were discussed. To name a few, laminar flow promotes the diffusion of intracellular chemical substances in the injured area;28,29 laminar flow can stimulate the activation of intracellular signal pathway;26,29 and laminar flow can promote physical migration of cells in the direction of flow.26
So far as we know, Rho is one of the main factors that are involved in EC directional migration under shear stress. Although the exact pathway to activate small Rho GTPases is not well understood, it is widely recognized that shear stress-induced integrin-extracellular matrix signaling pathway was the main pathway of its activation. Rac1 is one member of small GTPases of Rho family is a cycle between an active GTP-bound form and an inactive GDP-bound form, thereby functioning as molecular switches to turn on/off the downstream signal transduction processes.30 Cell migration will be inhibited as Rac1 is inhibited.31 Therefore, Rac1 is a key molecule of intracellular signal transduction in the process of migration, which responds to diverse extracellular cues. In this study, our results showed that Rac1 was shear stress-dependent expression of regulating molecules in EC migration.
Our experiments also showed that, compared with control group under shear condition, the high degree activation of Rac1 by Rac1Q61L transfection promoted a great degree of cell migration, while the negative mutant of Rac1 (Rac1T17N) inhibited cell migration. Both Tzima32 and Wojciak-Stothard33 did high-quality research on the effects of Rac1 in EC migration, showed that Rac1 actived at 5 min and reach to peak at 30 min, and subsequently decreased at 1 h (still higher than static controls) under shear conditions. The main difference is that we applied three levels of shear stress, while 12 dyn/cm2 and 3 dyn/cm2 were applied in other research.32,33 Our results indicated that the level of Rac1 activation played an important role in shear stress-induced cell migration. Hu et al.8 reported that bovine aortic endothelial cells (BAECs) transfected with active mutant Rac1V12 increased lamellipodia in all directions, but inhibited the polarization of lamellipodial protrusion for migration. The different migration results between Hu et al.’s and ours may result from the different active mutant of Rac1 and magnitudes of shear stress (12 dyn/cm2 in Hu et al.’s). Different active mutant of Rac1 may cause a different increased degree of Rac1 activity. The overactivation of Rac1 induced excessive lamellipodial formation, which was associated with non-polarized adhesions that are too strong for the cell to migrate.8 Meanwhile, our results showed that shear stress could increase the mRNA expression of Rac1. Three levels of shear stress were applied in this study, which could cause different mRNA expression of Rac1 in comparison with the shear stress of 12 dyn/cm2. Consequently, Rac1 involved in signaling events play a significant role in shear stress-induced EC migration, which is expected to explore the mechanism of shear stress-induced EC migration in the future research. However, it should be noted that only total Rac1 expression was concentrated in this study, the activity of Rac1 GTPase in laminar shear stress-induced vascular endothelial cell migration should be addressed. Moreover, knockdown of Rac1 would help us to further understand how shear-stress affects the small GTPase Rac1, which is expected to be done in our near-future study.
In this study, our results confirmed that Rac1 mediated different shear stress-induced EC migration under three levels of shear stress. These findings conduce to elucidate the molecular mechanisms of EC migration induced by shear stress, which is expected to understand the pathophysiological basis of wound healing in health and disease.
Materials and Methods
Cell culture
EA.hy 926 cells (purchased from the Blood Research Institute) are hybridoma cell lines between human umbilical vein endothelial cells (HUVECs) and the epithelioma A549 cells, and retain most of the features of HUVECs, including the expression of endothelial adhesion molecules and human factor VIII, related Ag. EA.hy 926 cells were cultured in RPMI1640 medium (Invitrogen Company) supplemented with 10% fetal bovine serum (FBS, Hangzhou Sijiqing Biological Engineering Materials Co., Ltd.), 20 mmol/L HEPES (Sigma), 2% NaHCO3 and 2% HAT (Sigma). The cells were maintained in a 5% CO2 incubator at 37 °C.
Shear stress experiment
The flow chamber was designed and developed by our laboratory.34 The flow pattern in the experimental area of the flow chamber was laminar, two-dimensional, and fully established. The steady flow chamber is provided by a peristaltic pump (MASTER FLEX 07524-40 Cole Parmer). Three levels of shear stress were applied in this study to investigate the effects of different shear stress on EC migration, which were a relatively “low” shear stress of 5.56 dyn/cm2 that denoted shear stress levels within disturbed flow regions, a relatively “mid” shear stress of 10.02 dyn/cm2 reflecting within the physiological range in human major arteries and a relatively “high” shear stress of 15.27 dyn/cm2 that was typical for undisturbed flow zones within large arteries. Therefore, all experiments were performed at the flow rate of 140, 240, and 350 ml/min, corresponding to shear stress of 5.56, 10.02, and 15.27 dyn/cm2, respectively.
Scratch wound migration assay
Cell migration was measured using a monolayer scratch injury assay as described previously.35,36 Briefly, the EA.hy 926 cells were plated onto glass slides with confluence, and then cells were washed and incubated with serum-free media overnight. At same time, hydroxyurea (Sigma) was added to a final concentration of 5 mmol/L to further inhibit cell proliferation.37 A uniform straight scratch was performed in the cell monolayer using a plastic Cell Scrapper (Corning). Then, cell monolayers were washed gently, marked (for reference), and photographed using an inverted microscope (Olympus IX-71). The glass slides with wounded monolayers perpendicular to laminar shear flow direction were placed into flow chamber. Images of the wounds under both static and flow conditions were acquired at 2, 4, 6, and 8 h, respectively.
Migration distance and wound repair area analysis
EA.hy 926 cell migration during wound repair involves in displacements of both parallel and orthogonal to the direction of flow. Cell migration parallel to the flow direction is expected to be the primary contributor to wound repair. Therefore, the repair area ratio combined with migration distance was examined to determine whether flow impacts on the directionality of cell migration in this study. The analysis method of migrated distance and wound repair area was described in our previous study.27 Briefly, cell migration during the wound repair was analyzed using the image analysis software (Image Pro Plus 6.0, Media Cybernetics). The acquired images were converted from pixels to micrometers with the use of a calibration image. For each experiment, 10 cells were randomly chosen along each edge of the wound, and cell migration distance at the end of each recording period was calculated by Dn = Ln - Ln where Dn and Ln are the net cell migration distance and the cell position at the metering point n (h), respectively. L0 is the original position.
Using Image Pro Plus software, the outline of wound could be traced and the area could be tabulated.38,39 The wound repair area ratio at the end of each recording period was calculated by
where
was the wound repair area ratio, An was the area of the polygon that defines the wound’s outline at the metering point n (h), and A0 was the origin wound repair area.
RT-PCR
The expression of Rac1 mRNA under laminar flow for various durations was semi-quantitatively detected by reverse transcription–polymerase chain reaction (RT-PCR). Total RNA was isolated from the cells using the TRIzol reagent (Invitrogen Company) and RT-PCR was performed by using the TaKaRa One Step RNA PCR Kit (AMV) (TaKaRa, Dalian) according to the manufacturer’s instructions. To obtain the relative quantitative values for gene expression, β-actin was used as the endogenous control. Sequences of the primers used for RT-PCR listed in Table 1 were designed by Dalian Biotechnology. The PCR products were visualized by electrophoresis on a 1% agarose gel in 1 × TRIS-acetate-EDTA (TAE) buffer after staining with 0.5 μg·ml−1 ethidium bromide.
Table 1. Sequence of primers used for RT-PCR analysis.
| Gene name | Sense primer | Anti-sense primer |
|---|---|---|
| β-actin | 5’-CCAAGGCCAAC CGCGAGAAGA TGAC -3’ | 5’-AGGGTACATG GTGGTGCCGC CAGAC -3’ |
| Rac1 | 5’-GGGGATCCCA GGCCATCAAG TGTGTGGTGG-3’ | 5’-GGAATTCTTA CAACAGCAGC AGGCATTTTC TCTTCC-3’ |
Western blot analysis
EA.hy926 cells were treated with mechanical stimulus (10.02 dyn/cm2) for 2 h, and then proteins were extracted according to protein extraction kit (PMSF methods, Beyotime Biotech). Protein samples were separated on precast SDS-PAGE (10% resolving gel with 5% stacking) and semi-dry transferred to the polyvinylidene fluoride membranes (PVDF, Millipore). Membranes were blocked for 2 h at room temperature with blocking buffer containing 5% non-fat dry milk powder in Tris buffered saline containing 0.1% Tween-20 (TBS-T), and probed with 1:100 diluted primary antibodies (Rac 1 [C-1]: sc-95) specific for the target protein, and β-actin was used as the endogenous control. Membranes were incubated at 4 °C overnight, followed by incubation for 2 h at room temperature with a 1:8000 diluted HRP-linked secondary antibodies goat anti-rabbit IgG (Zhongshan Goldenbridge Biotechnology Co., Ltd.). Immunoreactive proteins were visualized using the enhanced chemiluminescence (ECL, Pierce) following the manufacturer’s instructions.
Liposome plasmid DNA transfection and immunofluorescence
Plasmids were extracted by using plasmid mini preparation kit (Omega) according to manufacturer’s instructions. Plasmids encoding the wild-type (WT), an activated mutant (Q61L), and a dominant negative mutant (T17N) were transfected into EA.hy 926 cells, respectively. To facilitate the detection of individual cells harboring the exogenously introduced GTPases, Rac1 was fused to the C-terminal of the enhanced GFP (EGFP).40 Plasmids encoding the GFP-tagged proteins were transfected into EA.hy 926 cells at 80% confluence using the lipofectamine method.8 EA.hy 926 cells transfected with three plasmids were treated with or without mechanical stimulus for various duration and then fluorescence microscope was used to detect the expression of Rac1.
Data analysis
All experiments repeated three times (n = 3). Means, together with standard deviations, were calculated for the experimental and control groups. One-Way ANOVA with Duncan’s new multiple range method was performed by using data analysis software of SPSS12.0 (SPSS, Inc.). The results were expressed as means ± SD. The P values were calculated to test the level of statistical significance. The differences were considered significant at P < 0.05 and highly significant at P < 0.01.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
This study was supported in part by grant from National Natural Science Foundation of China (No.11372203 and No.11172189), Specialized Research Fund for the Doctoral Program of Higher Education (20120181120058), and China Medical Board (82-412).
Footnotes
Previously published online: www.landesbioscience.com/journals/celladhesion/article/27171
References
- 1.Lauffenburger DA, Horwitz AF. Cell migration: a physically integrated molecular process. Cell. 1996;84:359–69. doi: 10.1016/S0092-8674(00)81280-5. [DOI] [PubMed] [Google Scholar]
- 2.Ridley AJ, Allen WE, Peppelenbosch M, Jones GE. Rho family proteins and cell migration. Biochem Soc Symp. 1999;65:111–23. [PubMed] [Google Scholar]
- 3.Sheetz MP, Felsenfeld D, Galbraith CG, Choquet D. Cell migration as a five-step cycle. Biochem Soc Symp. 1999;65:233–43. [PubMed] [Google Scholar]
- 4.Bao G, Suresh S. Cell and molecular mechanics of biological materials. Nat Mater. 2003;2:715–25. doi: 10.1038/nmat1001. [DOI] [PubMed] [Google Scholar]
- 5.Barakat AI, Davies PF. Mechanisms of shear stress transmission and transduction in endothelial cells. Chest. 1998;114(Suppl):58S–63S. doi: 10.1378/chest.114.1_Supplement.58S. [DOI] [PubMed] [Google Scholar]
- 6.Chappell DC, Varner SE, Nerem RM, Medford RM, Alexander RW. Oscillatory shear stress stimulates adhesion molecule expression in cultured human endothelium. Circ Res. 1998;82:532–9. doi: 10.1161/01.RES.82.5.532. [DOI] [PubMed] [Google Scholar]
- 7.Chien S, Li S, Shyy YJ. Effects of mechanical forces on signal transduction and gene expression in endothelial cells. Hypertension. 1998;31:162–9. doi: 10.1161/01.HYP.31.1.162. [DOI] [PubMed] [Google Scholar]
- 8.Hu YL, Li S, Miao H, Tsou TC, del Pozo MA, Chien S. Roles of microtubule dynamics and small GTPase Rac in endothelial cell migration and lamellipodium formation under flow. J Vasc Res. 2002;39:465–76. doi: 10.1159/000067202. [DOI] [PubMed] [Google Scholar]
- 9.Traub O, Berk BC. Laminar shear stress: mechanisms by which endothelial cells transduce an atheroprotective force. Arterioscler Thromb Vasc Biol. 1998;18:677–85. doi: 10.1161/01.ATV.18.5.677. [DOI] [PubMed] [Google Scholar]
- 10.Lamalice L, Le Boeuf F, Huot J. Endothelial cell migration during angiogenesis. Circ Res. 2007;100:782–94. doi: 10.1161/01.RES.0000259593.07661.1e. [DOI] [PubMed] [Google Scholar]
- 11.Li S, Huang NF, Hsu S. Mechanotransduction in endothelial cell migration. J Cell Biochem. 2005;96:1110–26. doi: 10.1002/jcb.20614. a. [DOI] [PubMed] [Google Scholar]
- 12.Li Y-SJ, Haga JH, Chien S. Molecular basis of the effects of shear stress on vascular endothelial cells. J Biomech. 2005;38:1949–71. doi: 10.1016/j.jbiomech.2004.09.030. b. [DOI] [PubMed] [Google Scholar]
- 13.Resnick N, Yahav H, Schubert S, Wolfovitz E, Shay A. Signalling pathways in vascular endothelium activated by shear stress: relevance to atherosclerosis. Curr Opin Lipidol. 2000;11:167–77. doi: 10.1097/00041433-200004000-00010. [DOI] [PubMed] [Google Scholar]
- 14.Wennerberg K, Rossman KL, Der CJ. The Ras superfamily at a glance. J Cell Sci. 2005;118:843–6. doi: 10.1242/jcs.01660. [DOI] [PubMed] [Google Scholar]
- 15.Kraynov VS, Chamberlain C, Bokoch GM, Schwartz MA, Slabaugh S, Hahn KM. Localized Rac activation dynamics visualized in living cells. Science. 2000;290:333–7. doi: 10.1126/science.290.5490.333. [DOI] [PubMed] [Google Scholar]
- 16.Raftopoulou M, Hall A. Cell migration: Rho GTPases lead the way. Dev Biol. 2004;265:23–32. doi: 10.1016/j.ydbio.2003.06.003. [DOI] [PubMed] [Google Scholar]
- 17.Tzima E. Role of small GTPases in endothelial cytoskeletal dynamics and the shear stress response. Circ Res. 2006;98:176–85. doi: 10.1161/01.RES.0000200162.94463.d7. [DOI] [PubMed] [Google Scholar]
- 18.Katoh H, Hiramoto K, Negishi M. Activation of Rac1 by RhoG regulates cell migration. J Cell Sci. 2006;119:56–65. doi: 10.1242/jcs.02720. [DOI] [PubMed] [Google Scholar]
- 19.Small JV, Stradal T, Vignal E, Rottner K. The lamellipodium: where motility begins. Trends Cell Biol. 2002;12:112–20. doi: 10.1016/S0962-8924(01)02237-1. [DOI] [PubMed] [Google Scholar]
- 20.Urbich C, Walter DH, Zeiher AM, Dimmeler S. Laminar shear stress upregulates integrin expression: role in endothelial cell adhesion and apoptosis. Circ Res. 2000;87:683–9. doi: 10.1161/01.RES.87.8.683. [DOI] [PubMed] [Google Scholar]
- 21.Lai Y, Liu X, Wu J, Zeng Y, Yue A, He X. [Rac1 mediates the migration of endothelial cells induced by IL-8] Sheng Wu Yi Xue Gong Cheng Xue Za Zhi. 2008;25:357–62. [in Chinese] [PubMed] [Google Scholar]
- 22.Zeng Y, Sun HR, Yu C, Lai Y, Liu XJ, Wu J, Chen HQ, Liu XH. CXCR1 and CXCR2 are novel mechano-sensors mediating laminar shear stress-induced endothelial cell migration. Cytokine. 2011;53:42–51. doi: 10.1016/j.cyto.2010.09.007. [DOI] [PubMed] [Google Scholar]
- 23.Cascone I, Audero E, Giraudo E, Napione L, Maniero F, Philips MR, Collard JG, Serini G, Bussolino F. Tie-2-dependent activation of RhoA and Rac1 participates in endothelial cell motility triggered by angiopoietin-1. Blood. 2003;102:2482–90. doi: 10.1182/blood-2003-03-0670. [DOI] [PubMed] [Google Scholar]
- 24.Chung CY, Funamoto S, Firtel RA. Signaling pathways controlling cell polarity and chemotaxis. Trends Biochem Sci. 2001;26:557–66. doi: 10.1016/S0968-0004(01)01934-X. [DOI] [PubMed] [Google Scholar]
- 25.Nobes CD, Hall A. Rho GTPases control polarity, protrusion, and adhesion during cell movement. J Cell Biol. 1999;144:1235–44. doi: 10.1083/jcb.144.6.1235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hsu PP, Li S, Li YS, Usami S, Ratcliffe A, Wang X, Chien S. Effects of flow patterns on endothelial cell migration into a zone of mechanical denudation. Biochem Biophys Res Commun. 2001;285:751–9. doi: 10.1006/bbrc.2001.5221. [DOI] [PubMed] [Google Scholar]
- 27.Li S. Analysis of endothelial cell migration under flow. Methods Mol Biol. 2005;294:107–21. doi: 10.1385/1-59259-860-9:107. [DOI] [PubMed] [Google Scholar]
- 28.Coomber BL. Centrosome reorientation in regenerating endothelial monolayers requires bFGF. J Cell Biochem. 1993;52:289–96. doi: 10.1002/jcb.240520305. [DOI] [PubMed] [Google Scholar]
- 29.Shiu Y-T, Li S, Marganski WA, Usami S, Schwartz MA, Wang Y-L, Dembo M, Chien S. Rho mediates the shear-enhancement of endothelial cell migration and traction force generation. Biophys J. 2004;86:2558–65. doi: 10.1016/S0006-3495(04)74311-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hall A, Nobes CD. Rho GTPases: molecular switches that control the organization and dynamics of the actin cytoskeleton. Philos Trans R Soc Lond B Biol Sci. 2000;355:965–70. doi: 10.1098/rstb.2000.0632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ridley AJ. Rho GTPases and cell migration. J Cell Sci. 2001;114:2713–22. doi: 10.1242/jcs.114.15.2713. [DOI] [PubMed] [Google Scholar]
- 32.Tzima E, Del Pozo MA, Kiosses WB, Mohamed SA, Li S, Chien S, Schwartz MA. Activation of Rac1 by shear stress in endothelial cells mediates both cytoskeletal reorganization and effects on gene expression. EMBO J. 2002;21:6791–800. doi: 10.1093/emboj/cdf688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wojciak-Stothard B, Ridley AJ. Shear stress-induced endothelial cell polarization is mediated by Rho and Rac but not Cdc42 or PI 3-kinases. J Cell Biol. 2003;161:429–39. doi: 10.1083/jcb.200210135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Liu X, Chen H. [Simulation on the hydrodynamics of a flow chamber system] Sheng Wu Yi Xue Gong Cheng Xue Za Zhi. 1999;16:441–4. [in Chinese] [PubMed] [Google Scholar]
- 35.Fan WH, Pech M, Karnovsky MJ. Connective tissue growth factor (CTGF) stimulates vascular smooth muscle cell growth and migration in vitro. Eur J Cell Biol. 2000;79:915–23. doi: 10.1078/0171-9335-00122. [DOI] [PubMed] [Google Scholar]
- 36.Liu X, Luo F, Pan K, Wu W, Chen H. High glucose upregulates connective tissue growth factor expression in human vascular smooth muscle cells. BMC Cell Biol. 2007;8:1. doi: 10.1186/1471-2121-8-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zeng Y, Liu XH, Shen Y, Lai Y, Liu XJ. Laminar shear stress promotes endothelial cell migration and inhibits cell apoptosis in the presence of hydroxyurea. Cell Mol Biol (Noisy-le-grand) 2011;57(Suppl):OL1550–7. [PubMed] [Google Scholar]
- 38.Albuquerque ML, Waters CM, Savla U, Schnaper HW, Flozak AS. Shear stress enhances human endothelial cell wound closure in vitro. Am J Physiol Heart Circ Physiol. 2000;279:H293–302. doi: 10.1152/ajpheart.2000.279.1.H293. [DOI] [PubMed] [Google Scholar]
- 39.Savla U, Waters CM. Mechanical strain inhibits repair of airway epithelium in vitro. Am J Physiol. 1998;274:L883–92. doi: 10.1152/ajplung.1998.274.6.L883. [DOI] [PubMed] [Google Scholar]
- 40.del Pozo MA, Vicente-Manzanares M, Tejedor R, Serrador JM, Sánchez-Madrid F. Rho GTPases control migration and polarization of adhesion molecules and cytoskeletal ERM components in T lymphocytes. Eur J Immunol. 1999;29:3609–20. doi: 10.1002/(SICI)1521-4141(199911)29:11<3609::AID-IMMU3609>3.0.CO;2-S. [DOI] [PubMed] [Google Scholar]