

Abstract
Among Paragonimus species, P. paishuihoensis is one of the most mysterious and poorly understood species. Metacercariae are characterized by having a unique dendritically branched excretory bladder. However, the morphology of the adult worm remains unknown. To date, metacercariae of this species have been reported only in China and Thailand. In this study, we first found P. paishuihoensis metacercariae in freshwater crabs, Potamon lipkei, in Hinheub District, Vientiane, Lao PDR, with a prevalence of 77.7% and the average intensity of 10.3 (range 1-28) metacercariae per crab. The molecular data based on ITS2 and CO1 markers indicated that P. paishuihoensis from Laos and Thailand were almost completely identical and were close to members of the Paragonimus bangkokensis/Paragonimus harinasutai complex. Attempts to infect experimental animals (cats, dogs, and rats) with P. paishuihoensis were unsuccessful, suggesting that these animals might be unsuitable definitive hosts for the species. Further studies are necessary to elucidate the taxonomic status and life cycle of P. paishuihoensis.
Keywords: Paragonimus paishuihoensis, metacercaria, freshwater crab, Potamon lipkei, Lao PDR
INTRODUCTION
Among about 50 nominal species of the genus Paragonimus, Paragonimus paishuihoensis Tsao and Chung 1965 is one of the most mysterious and poorly understood species because no adult has ever been obtained [1]. The metacercariae of this species, originally found in Szechuan (Sichuan) Province, China, are unique in the genus Paragonimus because they possess a highly branched excretory bladder [2]. After experimental infection in cats, only 1 juvenile worm was obtained [2]. Subsequently, Wang et al. [3,4] found P. paishuihoensis metacercariae in freshwater crabs in Yunnan Province, China. At almost the same time, Cui et al. [5] determined nuclear ribosomal second internal transcribed spacer region (ITS2) and a partial mitochondrial cytochrome c oxidase subunit 1 gene (CO1) sequences of P. paishuihoensis and another 4 Paragonimus species in China and revealed that P. paishuihoensis was genetically close to Paragonimus bangkokensis, Paragonimus menglaensis and Paragonimus xiangshanensis. In addition to those reports from China, Rangsiruji et al. [6] discovered P. paishuihoensis metacercariae in Chanthaburi Province, Thailand, and reported genetic similarities of this species to P. bangkokensis and Paragonimus harinasutai based on analyses of ITS2 and CO1 sequences. Here, we report morphological and molecular features of P. paishuihoensis metacercariae first found in freshwater crabs, Potamon lipkei, in Lao PDR.
MATERIALS AND METHODS
Collection of metacercariae
Paragonimus metacercariae were collected from freshwater crabs, Potamon lipkei, in Hinheub district, Vientiane Province, Lao PDR, by the methods described previously [7]. Nine crabs were examined individually in detail to determine the prevalence of each Paragonimus species and their distribution in crab tissues (Table 1). P. paishuihoensis metacercariae collected from additional crabs used for experimental infection were not included in Table 1. Metacercariae were identified based on morphological characteristics. Metacercariae that have a dendritic excretory bladder were regarded as P. paishuihoensis and were used for experimental infection and for morphological and molecular analyses.
Table 1.
Prevalence and distribution of Paragonimus metacercariae in 9 crabs Potamon lipkei
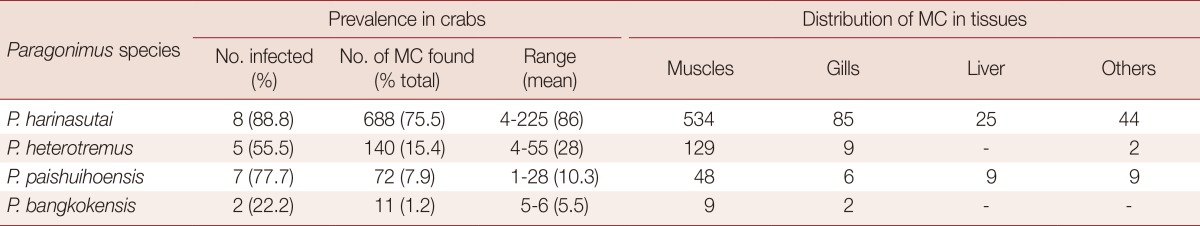
MC, Metacercariae
Experimental infection to laboratory animals
Fresh metacercariae having a dendritic excretory bladder were given to 5 cats, 1 dog, and 2 rats either orally or intraperitoneally (Table 2). Fecal examination was carried out 1 month after infection and weekly thereafter. Since none of those animals became Paragonimus egg-positive after several months, they were autopsied at the designated time (Table 2) to examine the lungs. In case of rats, the whole body was examined using pepsin digestion.
Table 2.
Results of experimental infection in laboratory animals with P. paishuihoensis metacercariae
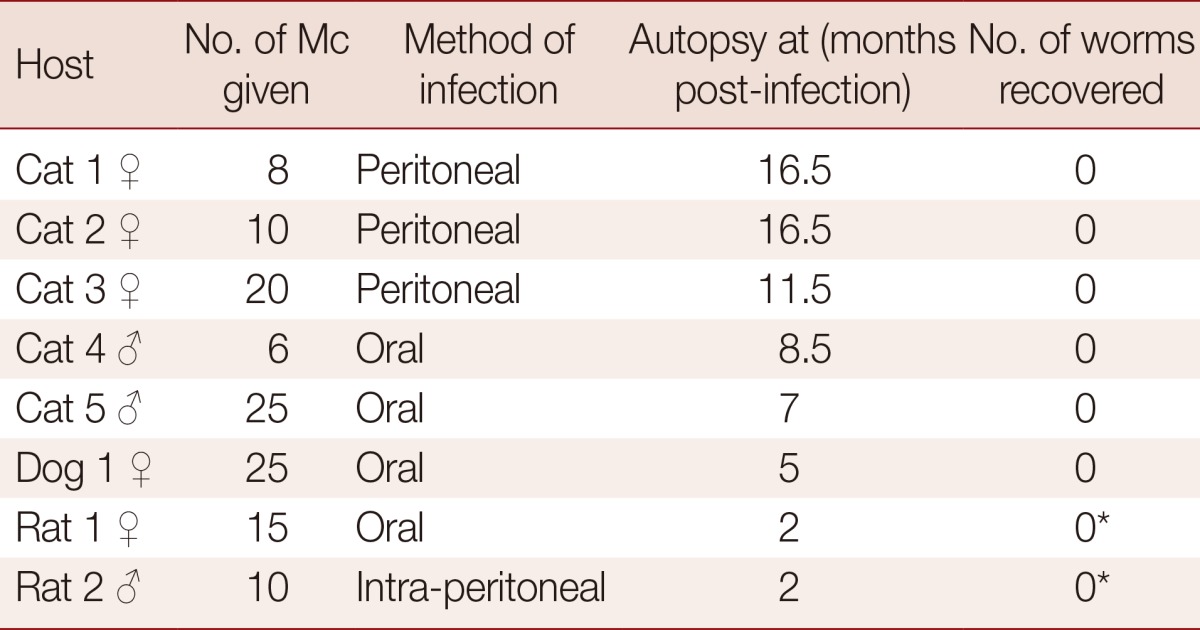
*Whole body was examined. Mc, metacercariae.
Molecular analyses
From 3 metacercariae of P. paishuihoensis (codes: MC32-MC34), we amplified and sequenced the ITS2 region using the primer pair 3S and A28 [8] and a partial CO1 gene using the primer pair of CO1-1-forward and CO1-2-reverse [9]. The methods for molecular study followed those described previously [10]. The obtained sequences were deposited in the GenBank with accession no. AB679285-AB679290. In addition, we downloaded and used further sequences of Paragonimus spp. from GenBank, including the available sequences (ITS2: AB471886 and CO1: AB471887) of P. paishuihoensis from Thailand and 1 ITS2 sequence (AB370191), which was originally registered as Paragonimus sp. from Laos and is now re-registered as P. paishuihoensis by us. Two sequence data sets, ITS2 and CO1, were used to reconstruct the phylogenetic trees. Each set was aligned using Clustal-W [11] with default options. Evolutionary analyses were conducted in MEGA5 [12]. Maximum likelihood trees were reconstructed using the best models: the Kimura 2-parameter with gamma rate heterogeneity (+G) for ITS2 tree and Hasegawa-Kishino-Yano model with invariable sites (+I) for CO1 tree.
RESULTS
Morphology of P. paishuihoensis metacercariae
When we examined Paragonimus metacercariae in the freshwater crabs, P. lipkei, caught in Hinheub District, Vientiane, Lao PDR, we found metacercariae having an excretory bladder of dendritic appearance (Fig. 1) in addition to 3 previously reported species; P. harinasutai, Paragonimus heterotremus, and P. bangkokensis [13,14]. By light microscopic observations, these metacercariae were spherical in shape with 2 cyst walls (Fig. 1A,B). The outer cyst (n=8) was 702.0±46.0×670.5±49.1 µm (range 577-725×631-755) in diameter, while the inner cyst (n=17) was 620.9±30.9×573.9±28.8 µm (range 518-622×580-681). A distinct space was present between the 2 walls. The thickness of outer and inner cyst wall was 4.9±0.3 and 5.7±0.7 µm, respectively. Two metacercariae were artificially excysted (Fig. 1C) and measured. They measured 1,502×737 and 1,436×533 µm respectively; oral suckers measured 105×118 and 117×112 µm, and ventral suckers 186×211 and 158×192 µm, respectively. A branched excretory bladder runs the length of the body. All these morphological and morphometric data of encysted and excysted metacercariae were compatible with those of P. paishuihoensis reported previously [2,3,6].
Fig. 1.

Encysted form (A, B) and excysted form (C) of P. paishuihoensis metacercariae from crabs (P. lipkei) collected in Lao PDR.
Prevalence of Paragonimus metacercariae in crabs
In this study, 9 crabs were examined individually in detail to determine the prevalence of each Paragonimus species and their distributions in crab host tissues. Among 4 co-existing Paragonimus metacercariae in crabs, P. harinasutai was the most common species which was found in 8 (88.8%) crabs and accounted for 75.5% of total metacercariae found, followed by P. heterotremus, P. paishuihoensis, and P. bangkokensis (Table 1). Individual crab harbored metacercariae of 1-4 Paragonimus species. Regardless of the Paragonimus species, the majority of metacercariae were distributed in the muscles and hepatopancreas of the crab, and only a few were found in the gills or other tissues (Table 1).
Experimental infection
None of the 8 experimental animals (5 cats, 1 dog, and 2 rats), which were infected with P. paishuihoensis metacercariae, became egg-positive after infection (range of times until autopsy; 2.0-16.5 months). At autopsy, all of them were negative for Paragonimus worms (Table 2). No lesion suggestive of Paragonimus infection was found in the pleural and abdominal cavity of any experimental animal.
Molecular analyses
For each gene region, the sequences of P. paishuihoensis isolates from Lao PDR were almost completely identical with each other; only 1 and 2 nucleotide differences were observed in the ITS2 (363 bp) and CO1 (354 bp) alignments, respectively. Similarities between the Lao sequences and those from the Thai sample were 99.9% in ITS2 (in spite of an AG indel) and 96.2% in CO1 sequences. Consequently, Lao and Thai samples formed a reasonably well-supported group in trees inferred from either gene region (Fig. 2). P. paishuihoensis was shown to be close to the P. bangkokensis/P. harinasutai complex.
Fig. 2.

The Maximum Likelihood tree constructed from ITS2 (A) and CO1 (B) sequences. Bootstrap scores (percentages of 1,000 replications) are presented for each node. The sequences obtained from the samples in this study are represented in bold, while others from DNA database are shown with the GenBank accession no., species name, and country code.
DISCUSSION
The present study clearly showed that P. paishuihoensis is present in Vientiane Province, Lao PDR as the third country after China [2,3,13] and Thailand [6]. P. paishuihoensis metacercariae were often found together with 1-3 other Paragonimus species: P. harinasutai, P. heterotremus, and P. bangkokensis in a same crab, indicating that competition among these species of metacercariae may not so strict in P. lipkei. Consistent with a previous report [13], P. harinasutai was by far the commonest species in this study area (Table 1).
Until now, fully mature adult worms of P. paishuihoensis have never been found in wildlife, nor in experimentally infected animals except for 1 juvenile obtained by experimental infection in a cat in China [2]. We infected cats, a dog, and rats, which are known as susceptible definitive hosts for almost all Paragonimus spp. [1], with P. paishuihoensis metacercariae, but no worms were established themselves in these hosts. This indicates that dogs, cats, and rats are not suitable definitive hosts for P. paishuihoensis; it might have narrow host specificity to an unknown mammalian host, or require a paratenic host. If a paratenic host is required, our results suggest that it is not the rat; no juvenile worms were found when the body of each rat was completely digested. Further studies are necessary to elucidate the life cycle of P. paishuihoensis.
In molecular studies, Cui et al. [5] reported that P. paishuihoensis obtained in Yunnan Province, China, was genetically similar to P. xiangshanensis and P. bangkokensis from China. Unfortunately, those sequence data are not registered in GenBank. To date, P. paishuihoensis from Thailand is represented in GenBank by only a single ITS2 and a single CO1 sequence [6]. In the present study, molecular phylogenetic analyses revealed that P. paishuihoensis isolates from Lao PDR and Thailand were genetically almost completely identical with each other, thus forming a moderately to well-supported group in ITS2 and CO1 trees. Consistent with the previous reports [14,15], P. paishuihoensis sequences were clustered with P. harinasutai of China with a moderate bootstrap value of 76% to make a group separated from the rest of the P. bangkokensis/P. harinasutai. However, grouping with P. paishuihoensis and P. bangkokensis/P. harinasutai complex was supported by very low bootstrap values (less than 50%), especially in CO1 tree. For further investigation on taxonomic status in this group, more sampling and sequence data accumulation is necessary. Since only 1 each of ITS2 and CO1 sequence of P. harinasutai and none of other related species (P. xiangshanensis and P. bangkokensis) from China are available in the GenBank database, more sequence data of those species from China and also from other geographical locations would be useful for better understanding of their taxonomic status and phylogenetic relationship.
In conclusion, we demonstrated the presence of P. paishuihoensis metacercariae in Lao PDR, suggesting a wider distribution of this species in Indochina than previously suspected. Since the definitive hosts of P. paishuihoensis have not yet been identified, further studies are required to clarify the taxonomic status and life cycle of this species.
References
- 1.Blair D, Xu ZB, Agatsuma T. Paragonimiasis and the genus Paragonimus. Adv Parasitol. 1999;42:113–222. doi: 10.1016/s0065-308x(08)60149-9. [DOI] [PubMed] [Google Scholar]
- 2.Tsao WC, Chung HL. Preliminary observations on metacercariae cysts, excysted metacercariae and adolescent worm of a new species of lung fluke, Paragonimus paishuihoensis sp. nov. discovered in P'enghsien in Szechuan Province. Acta Parasitol. 1965;2:252–256. [Google Scholar]
- 3.Wang WL, Lei L, Zhou BJ, Zhang X, Wang ML, Xia DG. Investigation and observation on Paragonimus paishuihoensis in Mengla County, Yunnan Province. Endemic Dis Bull. 2002;17:62–65. [Google Scholar]
- 4.Wang WL, Shen YZ, Lei L, Zhou BJ, Song JL, Liu AW, Han D. Observations on the metacercariae of Paragonimus paishuihoensis by transmission electron microscopy. Chinese J Zoonosis. 2005;21:150–152. [Google Scholar]
- 5.Cui AL, Chang ZS, Chen MG, Blair D, Chen SH, Zhang YN, Feng Z. Taxonomic status in DNA sequences of five species of genus Paragonimus. Zhongguo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi. 2003;21:27–30. [PubMed] [Google Scholar]
- 6.Rangsiruji A, Binchai S, Qian BZ, Morishimia Y, Sugiyama H. A new form of Paragonimus metacercariae isolated from freshwater crabs: possible discovery of P. paishuihoensis in Thailand; Proceedings of Joint International Tropical Medicine Meeting; 2008. p. 137. [Google Scholar]
- 7.Habe S, Lai KPF, Agatsuma T, Ow Y, Kawashima K. Crab hosts for Paragonimus westermani (Kerbert, 1878) in Malaysia. Jpn J Trop Med Hyg. 1993;21:137–142. [Google Scholar]
- 8.Bowles J, Blair D, McManus DP. A molecular phylogeny of the human schistosomes. Mol Phylogenet Evol. 1995;4:103–109. doi: 10.1006/mpev.1995.1011. [DOI] [PubMed] [Google Scholar]
- 9.Thaenkham U, Waikagul J. Molecular phylogenetic relationship of Paragonimus pseudoheterotremus. Southeast Asian J Trop Med Public Health. 2008;39:217–221. [PubMed] [Google Scholar]
- 10.Doanh PN, Shinohara A, Horii Y, Habe S, Nawa Y, The DT, Le NT. Morphological and molecular identification of two Paragonimus spp., of which metacercariae concurrently found in a land crab, Potamiscus tannanti, collected in Yenbai Province, Vietnam. Parasitol Res. 2007;100:1075–1082. doi: 10.1007/s00436-006-0411-9. [DOI] [PubMed] [Google Scholar]
- 11.Thompson JD, Higgins DG, Gibson TJ. CLUSTAL-W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22:4673–4680. doi: 10.1093/nar/22.22.4673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28:2731–2739. doi: 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Odermatt P, Habe S, Manichanh S, Tran DS, Duong V, Zhang W, Phommathet K, Nakamura S, Barennes H, Strobel M, Dreyfuss G. Paragonimiasis and its intermediate hosts in a transmission focus in Lao People's Democratic Republic. Acta Trop. 2007;103:108–115. doi: 10.1016/j.actatropica.2007.05.015. [DOI] [PubMed] [Google Scholar]
- 14.Ngoc Doanh P, Shinohara A, Horii Y, Yahiro S, Habe S, Vannavong N, Strobel M, Nakamura S, Nawa Y. Morphological differences and molecular similarities between Paragonimus bangkokensis and P. harinasutai. Parasitol Res. 2009;105:429–439. doi: 10.1007/s00436-009-1402-4. [DOI] [PubMed] [Google Scholar]
- 15.Doanh PN, Hien HV, Nonaka N, Horii Y, Nawa Y. Co-existence of Paragonimus harinasutai and Paragonimus bangkokensis in Central Viet Nam with special emphasis on their close phylogenetic relationship. Parasitol Int. 2012;61:399–404. doi: 10.1016/j.parint.2012.01.011. [DOI] [PubMed] [Google Scholar]