

Abstract
Serotonin (5-HT) functions as a chemoattractant that modulates neural migration during prenatal and early postnatal development. However, its molecular mechanism remains to be elucidated. The effect of 5-HT on neural cell migration was examined using PC12 neuron-like cell line. Transwell migration assay was used to determine the effect of 5-HT on PC12 cell migration. The results demonstrated that 5-HT and nerve growth factor (NGF) induced PC12 cell migration in a dose-dependent manner. Additionally, 5-HT receptor antagonists suggest that 5-HT-induced migration was mediated by serotonin receptor 6 (5-HT6), a Gs-protein coupled receptor that elevates the intercellular cAMP level. By contrast, antagonists of serotonin receptor 3 (5-HT3) did not show any effects on PC12 cell migration. Clozapine, an inhibitor of cAMP accumulation mediated by 5-HT6, significantly reduced the effect of 5-HT on the PC12 cell migration. An inhibitor of extracellular signal-regulated kinase (ERK) also suppressed migration. These results suggest that 5-HT induces PC12 cell migration by activating cAMP/ERK signaling pathways, which is mediated by 5-HT6 receptor.
Keywords: serotonin, neural migration, serotonin receptor 6
Introduction
Neural cell migration is crucial in the formation of highly organized structures of mammalian brain. Abnormal migration in prenatal and early postnatal brain causes various types of psychiatric diseases, including mental retardation, autism, bipolar disorders and schizophrenia (1,2). Various molecules have been identified as inducers and modulators of the migration including growth/neurotrophic factors and certain types of neurotransmitters (3–5).
Serotonin (5-HT) has a critical role in neural migration, which is mainly supplied from placental sources and serotonergic projections from the dorsal raphe nuclei in the fetal brain (6). Depletion of 5-HT by injection of DL-P-chlorophenylalanine (PCPA, an inhibitor of 5-HT synthesis) during the E12-E17 stage inhibited migration and disorganized the positioning of cortical neurons (7). By contrast, cortical slices exposed to high doses of 5-HT (100–400 μM) inhibited the migration of GABergic neurons. The arrested migration was recovered by application of the 5-HT6 antagonist, SB258585 (8,9). However, the association between 5-HT dose and its effect on migration remains to be elucidated.
It is also unclear as to which serotonin receptor mediates 5-HT signal to modulate neural migration. At least 14 classes of 5-HT receptors were identified that are coupled with various types of G proteins, with the exception of 5-HT3, a ligand-gated ion channel receptor (10,11). 5-HT6 receptor coupled with Gs protein is positively linked to adenylate cyclase to increase the cAMP level. Recent studies have demonstrated that 5-HT3 and 5-HT6 are differentially expressed in migrating neurons in the cerebral cortex (9,12,13). Vitalis et al(13) identified 5-HT3 and 5-HT6 as candidates involved in the mediation of the 5-HT signal for migration of the pyramidal neurons in the cortex.
To determine the effects of 5-HT on neural cell migration, we used a PC12 neuron-like cell line that expresses 5-HT3 and 5-HT6 in experiments. 5-HT and nerve growth factor (NGF) induced PC12 cell migration via 5-HT6 but not 5-HT3, stimulating the cAMP and extracellular signal-regulated kinase (ERK) signaling pathways.
Materials and methods
Materials
5-HT, NGF, Ondansetron, SB271046, SB258585, Clozapine and PD98059 were obtained from Sigma-Aldrich (St. Louis, MO, USA). MDL7222 was obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
Cell culture
PC12 cells (RIKEN Tsukuba Institute, Tsukuba, Japan) were routinely cultured in Dulbecco’s modified Eagle’s medium (DMEM; Sigma-Aldrich) containing 10% fetal bovine serum (Life Technologies, Gaithersburg, MD, USA), 10% horse serum (Life Technologies), 50 U/ml penicillin and 50 μg/ml streptomycin (Life Technologies). The cells were incubated at 37ºC in a 5% CO2 atmosphere. For each experiment, 3×105 cells were spread onto a 60 mm dish (Becton Dickinson, Franklin Lakes, NJ, USA). Following incubation for two days, the cells were used for RNA extraction and transwell migration assay. To obtain differentiated cells, the medium was changed to NGF-supplemented medium (DMEM containing 100 ng/ml NGF, 1% horse serum, 50 U/ml penicillin and 50 μg/ml streptomycin) and incubated for one day.
RNA extraction and RT-PCR
Total RNAs were extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), then cDNAs were synthesized with ReverTra Ace (Toyobo, Osaka, Japan) according to the manufacturer’s instructions. PCRs were performed with a denaturation step at 95ºC for 5 min, followed by 35 (Fig. 2A), 29 (Fig. 2B) or 35 cycles (Fig. 2C) of denaturation at 95ºC for 30 sec, primer annealing at 55ºC for 30 sec, and primer extension at 72ºC for 30 sec. TATA binding protein (TBP) was used as a control. The primers used were: 5-HT1A (forward) 5′-CTCTGTTGCTGGGTACTCTCATT/(reverse) 5′-AGTCTATAGGGTCGGTGATAGCC-3′, 5-HT1B 5′-3′-, 5-HT2A 5′-TGTACGTGAACCAAGTCAAAGTG-3′/ 5′-GTAGATGATGGGGTTGATGAGAG-3′, 5-HT2A 5′-ATG CTGAAAACAGAACCAACCT-3′/5′-ACATCCAGGTAAAT CCAGATCG-3′, 5-HT2B 5′-TCGTCAAGATTACGG TGGTATG-3′/5′-CACCATCTTTTCTGGTGATGAA-3′, 5-HT2C 5′-ATAGGGGGCAACATTCTTGTTAT-3′/5′-ACAGGGATAGGAACTGAAACTCC-3′, 5-HT3, 5′-GGAA GTCTCCAAGCATTCCTTAT-3′/5′-ACGTAGAACTTC ATTTCCGCATA-3′, 5-HT4 5′-CCAATATTGTGGAC CCTTTCATA-3′/5′-GACTGGCTTCTTTTCAAGCTACA, 5-HT5A 5′-AAGATTTACAAGGCTGCGAAGTT-3′/5′-ACT GATGAGCTCCGTAACAAAGA-3′, 5-HT5B 5′-CTGG ATCGCTACTGGACTATCAC-3′/5′-GTGA ATACCGTCTCA GACTCCTG-3′, 5-HT6 5′-CTGGGAATGTTCTTTGT CACCT-3′/5′-GAAGCGGAGTCTGAATCTGA GTT-3′, 5-HT7 5′-ACTTCTTCTGCAACGTCTTCATC-3′/5′-GCG GCCTTGTAAATCTGATAGTA-3′, TBP 5′-TGCTGGCGG TTTGGCTAGGTTTCTGC-3′/5′-GGTCAGAGTTTGAGAA TGGAAGAGTT-3′.
Figure 2.
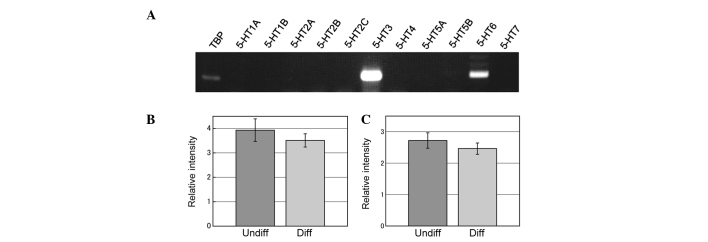
RT-PCR of serotonin (5-HT) receptors. (A) Expression of serotonin receptor 3 (5-HT3) and 5-HT6 was observed in undifferentiated PC12 cells that were used for the transwell migration assay. (B and C) Semi-quantitative RT-PCR reveals mRNA expression of 5-HT3 (B) and 5-HT6 (C) in undifferentiated (Undiff) and differentiated (Diff; 1 day after application of 100 ng/ml NGF) PC12 cells.
Transwell cell migration assay
PC12 cell suspension containing 2×105 cells in DMEM was applied to each upper well of the transwell chamber (Becton Dickinson), which was previously coated with type I collagen (50 μg/ml, Becton Dickinson) on both sides. In the bottom well, DMEM with or without motogen, NGF (100 ng/ml) and/or 5-HT (0.1–10 μM) were applied to allow the cells to migrate across filters (8 μm pore size). Ondansetron (1 μM, 5-HT3 antagonist), MDL7222 (1 μM, 5-HT3), SB271046 (1 μM, 5-HT6), SB258585 (1 μM, 5-HT6) and PD98059 (20 μM, ERK inhibitor) were also applied in some of the experiments. The transwell migration assay was performed at 37ºC for 5 h. After removal of the remaining cells on the top side of filters using cotton swabs, the filters were fixed with 4% PFA/PBS for 15 min. After incubation with Hoechst 33258 (Nacalai Tesque, Kyoto, Japan) at room temperature for 5 min, the cells on the bottom side of transwell inserts were washed three times with PBS and examined by fluorescence microscopy (IX83; Olympus, Tokyo, Japan). Ten images were captured randomly for one experiment and the number of nuclei in a 600×600 μm in each image was counted. Image J was used for counting as previously described (14). Relative percentages of the cell number to the average of the cell number in the control experiments (DMEM only in bottom well) were plotted on the graphs.
Statistical analysis
P-values were calculated by one- or two-way ANOVA followed by Tukey’s HSD. Data are presented as mean ± standard error.
Results
5-HT induced PC12 cells migration in a dose-dependent manner
To determine the effect of 5-HT on PC12 cell migration, the transwell migration assay was used (15,16). Cells were spread onto the upper side of the transwell insert and the number of cells migrating across the filter was counted. Addition of 5-HT to the bottom well together with DMEM significantly increased the migrating cells (Fig. 1A). 5-HT-induced migration was identified in a dose-dependent manner (Fig. 1B). An amount of 1 μM 5-HT significantly increased cell migration, while 0.1, 0.5, 5.0 and 10 μM 5-HT did not show significant changes compared with the control. NGF is known to induce PC12 cell migration (15,16), thus we added 5-HT together with NGF. NGF and 5-HT induce migration in an additive manner.
Figure 1.
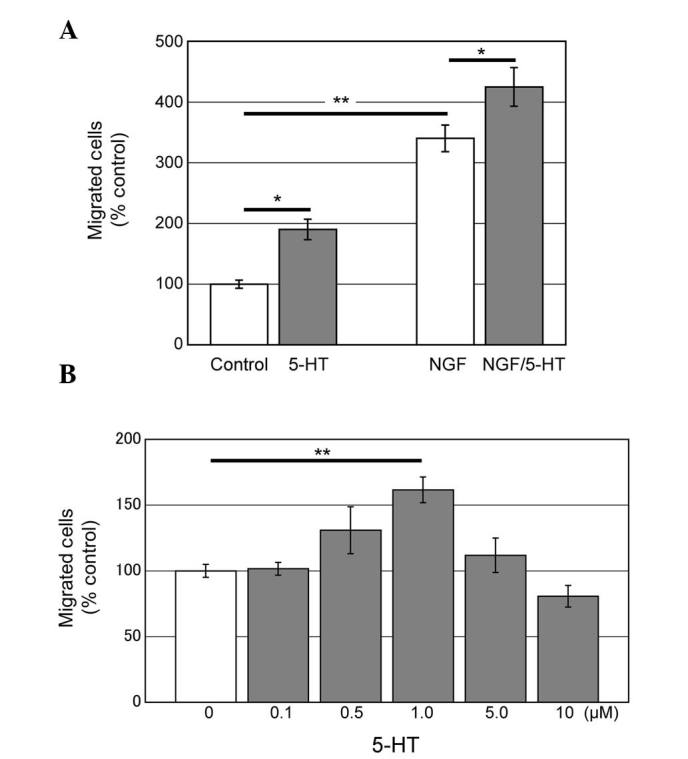
Transwell migration assay reveals serotonin (5-HT) induction of PC12 cell migration. (A) PC12 cells were spread onto the upper side of transwell inserts to allow migration through the membrane. In the bottom well, Dulbecco’s modified Eagle’s medium with or without chemical cues, serotonin (1 μM) and/or nerve growth factor (NGF) (100 ng/ml) were applied. 5-HT and NGF significantly increased the migrating cells. (B) 5-HT induced migration in a dose-dependent manner. P-values were calculated by two-way or one-way ANOVA followed by Tukey’s HSD. *P<0.05; **P<0.01. Data are presented as mean ± standard error. HSD, honestly significant difference.
5-HT6 receptor mediated 5-HT-induced migration
We examined which serotonin receptor mediates PC12 cell migration induced by 5-HT. Previously, it was reported that 5-HT3 enhanced neurite outgrowth induced by NGF in PC12 cells (17). However, little is known regarding the expression and molecular function of other 5-HT receptors in PC12 cells.
mRNA expression of 5-HT receptors was examined by RT-PCR and 5-HT6 and 5-HT3 were found to be expressed (Fig. 2A) in undifferentiated and differentiated (1 day after application of 100 ng/ml NGF) PC12 cells (Fig. 2B and C).
Using antagonists against 5-HT3 and 5-HT6 receptors, we investigated which 5-HT receptor mediates the migration induced by 5-HT. Addition of Ondansetron (5-HT3 antagonist, 1 μM) or MDL7222 (5-HT3, 1 μM) with 5-HT did not reveal any significant changes (Fig. 3A) while antagonists of 5-HT6, SB258585 (1 μM) or SB271046 (1 μM) inhibited the migration induced by 5-HT (Fig. 3A). The data indicate that 5-HT6 but not 5-HT3 mediates serotonin-induced PC12 cell migration.
Figure 3.
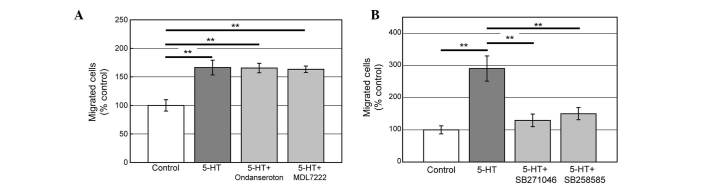
Effects of serotonin receptor antagonists. (A) Serotonin receptor 3 (5-HT3) antagonists, Ondansetron (1 μM) or MDL7222 (1 μM) did not inhibit migration. (B) 5-HT6 antagonists, SB271046 (1 μM) or SB258585 (1 μM) significantly reduced PC12 cell migration induced by 5-HT. Data were statistically analyzed by one-way ANOVA followed by Tukey’s HSD. **P<0.01. Data are presented as mean ± standard error.
Involvement of cAMP and ERK in 5-HT6-mediated signaling pathways
5-HT6 receptor is known to be coupled with Gs protein which mediates the accumulation of cAMP (18). To examine whether cAMP pathway is involved in 5-HT6 signaling in order to induce migration, Clozapine, which was reported to inhibit cAMP accumulation mediated by 5-HT6, was used (19). Clozapine (1 μM) significantly reduced the effect of 5-HT on migration (Fig. 4A). Increasing of cAMP activates ERK through protein kinase A-Rap1 (20). Thus, we examined whether the inhibition of ERK activation affects 5-HT-induced PC12 cell migration. Application of ERK inhibitor, PD98059 (20 μM), significantly suppressed the cell migration induced by 5-HT (Fig. 4B). These data indicate that cAMP and ERK are involved in the 5-HT6-mediated signaling pathways in order to induce PC12 cell migration.
Figure 4.
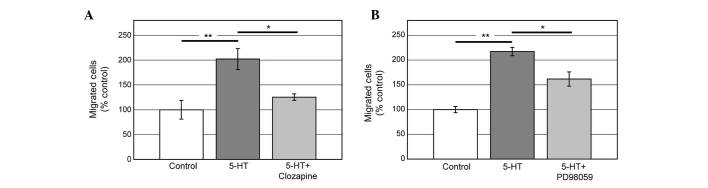
Effect of Clozapine (A) and PD98059 (B) on serotonin (5-HT)-induced PC12 cell migration. Clozapine (1 μM) and PD98059 (20 μM) significantly reduced migrating cells induced by 5-HT. Data were statistically analyzed by one-way ANOVA followed by Tukey’s HSD. *P<0.05; **P<0.01. Data are presented as mean ± standard error.
Discussion
5-HT induced PC12 cell migration in a dose-dependent manner
In the present study, we have demonstrated that 5-HT induced PC12 cell migration in a transwell migration assay in a dose-dependent manner (Fig. 1A). A number of studies have indicated that 5-HT affects proliferation of neural cells (7,21–23) as well as neurite outgrowth (17,24–26), however, its effect on neural migration remains to be clarified. Findings of previous studies suggest 5-HT affects cortical neuron migration during prenatal development (7–9,13). Depletion of 5-HT by injection of DL-P-chlorophenylalanine (PCPA, an inhibitor of 5-HT synthesis) during the E12-E17 stage arrested migration and disorganized the positioning of cortical neurons (7), suggesting a positive effect of 5-HT on migration. By contrast, cortical slices exposed to high doses of 5-HT (100–400 μM) arrested the migration of GABergic neurons (8,9). Although findings of those studies reported negative effects of 5-HT, the concentration of 5-HT was considerably higher than that of another study focusing on the prenatal cortex (100–200 fmol/mg) (7). The effect of 5-HT on neural migration therefore remains to be elucidated. A positive effect of 5-HT at the concentration of 1 μM was observed, which is similar to the in vivo results obtained in that study.
5-HT induced migration via 5-HT6 receptor-cAMP pathway independently from NGF-ERK
Blocking of 5-HT stimulation by 5-HT6 antagonists, SB271046 and SB258585 (Fig. 3B) indicate that 5-HT requires 5-HT6 receptor but not 5-HT3 to induce PC12 cell migration (Fig. 3A). 5-HT6 coupled with Gs-protein is known to elevate the cAMP level that stimulates ERK via protein kinase A (19,26). Inhibition of cAMP accumulation by Clozapine (19) and ERK by PD98059 (27) reduced the 5-HT effect on the migration (Fig. 4).
Previous studies suggest that NGF showed a positive effect on PC12 cell migration by activating ERK signaling (15,16). EGF and cAMP pathways activate ERK independently in order to promote PC12 cell differentiation (5,28,29). Similarly, it is possible that the 5-HT6-cAMP activates an independent pathway from the NGF-ERK signal to yield an additive effect on PC12 cell migration (Fig. 1A).
Insensitivity of 5-HT3 in undifferentiated PC12 cells
Antagonists of 5-HT3 did not show any effects on PC12 cell migration in our experiment (Fig. 3A) although 5-HT was expressed (Fig. 2A), suggesting independence of 5-HT3 signaling pathway from cAMP or ERK. Another possibility is that 5-HT3 is not sensitive to a 5-HT induction in the experiments of this study. Homma et al(17) suggest a difference of 5-HT sensitivity between differentiated (pretreated with NGF for 3 days) and undifferentiated PC12 cells to enhance neurite outgrowth mediated by 5-HT3. In their experiment, 50 μM 5-HT were required to enhance neurite outgrowth of undifferentiated PC12 cells whereas 5 μM of 5-HT were sufficient for differentiated cells. In our experiment, undifferentiated PC12 cells were treated with 1 μM 5-HT together with 5-HT3 antagonists (Fig. 3A). Higher concentrations of 5-HT may be required to activate the 5-HT3-dependent pathway in order to modulate PC12 cell migration.
5-HT3 and 5-HT6 are proposed as candidates to modulate neural migration (30). Although we observed an effect of 5-HT6 but not 5-HT3 on PC12 migration, it is possible 5-HT3 also modulated the PC12 cell migration in different conditions, such as in the presence of NGF. Transwell migration assay with PC12 cells is useful in the study of molecular mechanisms of the neural migration induced by 5-HT or by NGF in vitro.
Acknowledgements
This study was supported by JSPS KAKENHI Grant no. 23500427. We thank Kimie Iwasaki for her help.
References
- 1.Valiente M, Marín O. Neuronal migration mechanisms in development and disease. Curr Opin Neurobiol. 2010;20:68–78. doi: 10.1016/j.conb.2009.12.003. [DOI] [PubMed] [Google Scholar]
- 2.Brennand KJ, Simone A, Tran N, Gage FH. Modeling psychiatric disorders at the cellular and network levels. Mol Psychiatry. 2012;17:1239–1253. doi: 10.1038/mp.2012.20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Oliveira SL, Pillat MM, Cheffer A, et al. Functions of neurotrophins and growth factors in neurogenesis and brain repair. Cytometry A. 2013;83:76–89. doi: 10.1002/cyto.a.22161. [DOI] [PubMed] [Google Scholar]
- 4.Wang DD, Kriegstein AR. Defining the role of GABA in cortical development. J Physiol. 2009;587:1873–1879. doi: 10.1113/jphysiol.2008.167635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hernandez A, Kimball B, Romanchuk G, Mulholland MW. Pituitary adenylate cyclase-activating peptide stimulates neurite growth in PC12 cells. Peptides. 1995;16:927–932. doi: 10.1016/0196-9781(95)00059-s. [DOI] [PubMed] [Google Scholar]
- 6.Bonnin A, Goeden N, Chen K, et al. A transient placental source of serotonin for the fetal forebrain. Nature. 2011;472:347–350. doi: 10.1038/nature09972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Vitalis T, Cases O, Passemard S, et al. Embryonic depletion of serotonin affects cortical development. Eur J Neurosci. 2007;26:331–344. doi: 10.1111/j.1460-9568.2007.05661.x. [DOI] [PubMed] [Google Scholar]
- 8.Riccio O, Potter G, Walzer C, et al. Excess of serotonin affects embryonic interneuron migration through activation of the serotonin receptor 6. Mol Psychiatry. 2009;14:280–290. doi: 10.1038/mp.2008.89. [DOI] [PubMed] [Google Scholar]
- 9.Riccio O, Jacobshagen M, Golding B, et al. Excess of serotonin affects neocortical pyramidal neuron migration. Transl Psychiatry. 2011;1:e47. doi: 10.1038/tp.2011.49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Millan MJ, Marin P, Bockaert J, et al. Signaling at G-protein-coupled serotonin receptors: recent advances and future research directions. Trends Pharmacol Sci. 2008;29:454–464. doi: 10.1016/j.tips.2008.06.007. [DOI] [PubMed] [Google Scholar]
- 11.Hannon J, Hoyer D. Molecular biology of 5-HT receptors. Behav Brain Res. 2008;195:198–213. doi: 10.1016/j.bbr.2008.03.020. [DOI] [PubMed] [Google Scholar]
- 12.Lee S, Hjerling-Leffler J, Zagha E, et al. The largest group of superficial neocortical GABAergic interneurons expresses ionotropic serotonin receptors. J Neurosci. 2010;30:16796–16808. doi: 10.1523/JNEUROSCI.1869-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Vitalis T, Ansorge MS, Dayer AG. Serotonin homeostasis and serotonin receptors as actors of cortical construction: special attention to the 5-HT3Aand 5-HT6receptor subtypes. Front Cell Neurosci. 2013;7:93. doi: 10.3389/fncel.2013.00093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hassoun AT, Erdélyi F, Szabó G, Davis MI. A rapid screening method for population-specific neuronal motogens, substrates and associated signaling pathways. J Neurosci Methods. 2007;166:178–194. doi: 10.1016/j.jneumeth.2007.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ho W, Uniyal S, Meakin SO, et al. A differential role of extracellular signal-regulated kinase in stimulated PC12 pheochromocytoma cell movement. Exp Cell Res. 2001;263:254–264. doi: 10.1006/excr.2000.5112. [DOI] [PubMed] [Google Scholar]
- 16.Yoshizawa M, Kawauchi T, Sone M, et al. Involvement of a Rac activator, P-Rex1, in neurotrophin-derived signaling and neuronal migration. J Neurosci. 2005;25:4406–4419. doi: 10.1523/JNEUROSCI.4955-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Homma K, Kitamura Y, Ogawa H, Oka K. Serotonin induces the increase in intracellular Ca2+that enhances neurite outgrowth in PC12 cells via activation of 5-HT3receptors and voltage-gated calcium channels. J Neurosci Res. 2006;84:316–325. doi: 10.1002/jnr.20894. [DOI] [PubMed] [Google Scholar]
- 18.Choi YH, Kang H, Lee WK, et al. An inhibitory compound against the interaction between Gαsand the third intracellular loop region of serotonin receptor subtype 6 (5-HT6) disrupts the signaling pathway of 5-HT6. Exp Mol Med. 2007;39:335–342. doi: 10.1038/emm.2007.37. [DOI] [PubMed] [Google Scholar]
- 19.Kohen R, Metcalf MA, Khan N, et al. Cloning, characterization, and chromosomal localization of a human 5-HT6serotonin receptor. J Neurochem. 1996;66:47–56. doi: 10.1046/j.1471-4159.1996.66010047.x. [DOI] [PubMed] [Google Scholar]
- 20.Vossler MR, Yao H, York RD, et al. cAMP activates MAP kinase and Elk-1 through a B-Raf- and Rap1-dependent pathway. Cell. 1997;89:73–82. doi: 10.1016/s0092-8674(00)80184-1. [DOI] [PubMed] [Google Scholar]
- 21.Côté F, Fligny C, Bayard E, et al. Maternal serotonin is crucial for murine embryonic development. Proc Natl Acad Sci USA. 2007;104:329–334. doi: 10.1073/pnas.0606722104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Narboux-Nême N, Angenard G, Mosienko V, et al. Postnatal growth defects in mice with constitutive depletion of central serotonin. ACS Chem Neurosci. 2013;4:171–181. doi: 10.1021/cn300165x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cheng A, Scott AL, Ladenheim B, et al. Monoamine oxidases regulate telencephalic neural progenitors in late embryonic and early postnatal development. J Neurosci. 2010;30:10752–10762. doi: 10.1523/JNEUROSCI.2037-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lieske V, Bennett-Clarke CA, Rhoades RW. Effects of serotonin on neurite outgrowth from thalamic neurons in vitro. Neuroscience. 1999;90:967–974. doi: 10.1016/s0306-4522(98)00501-6. [DOI] [PubMed] [Google Scholar]
- 25.Lotto B, Upton L, Price DJ, Gaspar P. Serotonin receptor activation enhances neurite outgrowth of thalamic neurones in rodents. Neurosci Lett. 1999;269:87–90. doi: 10.1016/s0304-3940(99)00422-x. [DOI] [PubMed] [Google Scholar]
- 26.Zachor DA, Moore JF, Brezausek C, et al. Cocaine inhibits NGF-induced PC12 cells differentiation through D1-type dopamine receptors. Brain Res. 2000;869:85–97. doi: 10.1016/s0006-8993(00)02355-6. [DOI] [PubMed] [Google Scholar]
- 27.Whelchel A, Evans J, Posada J. Inhibition of ERK activation attenuates endothelin-stimulated airway smooth muscle cell proliferation. Am J Respir Cell Mol Biol. 1997;16:589–596. doi: 10.1165/ajrcmb.16.5.9160841. [DOI] [PubMed] [Google Scholar]
- 28.Vaudry D, Stork PJ, Lazarovici P, Eiden LE. Signaling pathways for PC12 cell differentiation: making the right connections. Science. 2002;296:1648–1649. doi: 10.1126/science.1071552. [DOI] [PubMed] [Google Scholar]
- 29.Lazarovici P, Jiang H, Fink D., Jr The 38-amino-acid form of pituitary adenylate cyclase-activating polypeptide induces neurite outgrowth in PC12 cells that is dependent on protein kinase C and extracellular signal-regulated kinase but not on protein kinase A, nerve growth factor receptor tyrosine kinase, p21rasG protein, and pp60c-srccytoplasmic tyrosine kinase. Mol Pharmacol. 1998;54:547–558. doi: 10.1124/mol.54.3.547. [DOI] [PubMed] [Google Scholar]
- 30.Engel M, Smidt MP, van Hooft JA. The serotonin 5-HT3receptor: a novel neurodevelopmental target. Front Cell Neurosci. 2013;7:76. doi: 10.3389/fncel.2013.00076. [DOI] [PMC free article] [PubMed] [Google Scholar]