

Abstract
Malaria is a potentially life-threatening protozoal parasitic disease transmitted by female Anopheles mosquitoes. Drug therapy is currently the most widely used method for the control and treatment of this disease. Several plants were found to contain substances possessing antimalarial properties. In this study, we investigated the antimalarial activity of bergenin, a sesquiterpene lactone compound derived from Rodgersia aesculifolia Batal. The results indicated that bergenin effectively inhibited Plasmodium falciparum growth in vitro (IC50, 14.1 μg/ml, with ~100% inhibition at 50 μg/ml), without apparent cytotoxicity to erythrocytes or to mammalian HeLa and HepG2 cells. Bergenin exhibited less cytotoxic activity and the selectivity index (SI) was 887 and 1,355 for HeLa and HepG2 cells, respectively. The administration of bergenin to Plasmodium berghei-infected mice for 6 days significantly inhibited the growth of the parasites. Taken together, these findings provide evidence that bergenin may be a promising novel drug for antimalarial treatment.
Keywords: malaria, bergenin, sesquiterpene lactone
Introduction
Malaria is a protozoal parasitic disease transmitted by female Anopheles mosquitoes that threatens the health of approximately one third of the world’s population and kills ~655,000 individuals in 106 malaria-endemic countries and regions worldwide annually (1). Drug therapy is currently the most widely used method for the control and treatment of this disease. Artemisinin-based combination therapies were recommended by the World Health Organization as first-line treatment for Plasmodium falciparum (P. falciparum) infection in 2006 (2,3). However, emerging resistance of the parasites to artemisinin, which is characterized by delayed parasite clearance time after treatment, has been observed in Southeast Asia. It was demonstrated that the half-life of the parasites, which normally declines after artesunate treatment, increased from a mean of 2.6 h in 2001 to 3.7 h in 2010 on the northwestern border of Thailand, compared to 5.5 h in 116 patients from Western Cambodia between 2007 and 2010 (4), where artemisinin resistance has been confirmed (5,6). Therefore, there is an urgent need for the development of novel drugs for malaria therapy, particularly in regions where resistant Plasmodium strains are present.
The use of natural products has provided a prospective strategy for identifying novel antimalarial drugs. It is estimated that >1,200 plant species from 160 families have been used to treat malaria and fever, including the widely used antimalarial drugs artemisinin and quinine derivatives (7). Since the discovery of artemisinin, the research of sesquiterpene lactone compounds has become the focus of antimalarial drug development (8,9).
Bergenin is a sesquiterpene lactone compound isolated from Rodgersia aesculifolia (R. aesculifolia) Batal, which is an effective and broad-spectrum antifungal and antiviral Chinese medicine. Bergenin was reported to possess antimicrobial properties against filamentous fungi, yeast and HIV, but not against bacteria (10–13). Bergenin also exhibits anti-inflammatory activity through the inhibition of cyclooxygenase-2 or by means of affecting the Th1- or Th2-skewed cytokine production (14,15). Bergenin also exerts an antioxidant effect by scavenging free radicals, such as •H, •OH and •CH3 (16). In addition, bergenin was reported to possess hepatoprotective, neuroprotective and gastroprotective properties (17–19). R. aesculifolia Batal containing bergenin was used to treat protozoal infection and fever in rural China; however, whether bergenin possesses antimalarial properties requires further elucidation. In this study, we evaluated the antimalarial activity of bergenin in vitro and in vivo trials.
Materials and methods
Mice and P. falciparum
Female BALB/c mice (aged 6–8 weeks) obtained from the Experimental Animal Center of The Fourth Military Medical University, Xi’an, China) were selected for the experiments. The mice were kept in a 12 h light/12 h dark cycle and were given commercial rodent solid diets and water ad libitum.
P. falciparum strain 3D7 (conserved in our laboratory) was maintained in human erythrocytes in RPMI-1640 medium supplemented with 10% human serum at 37°C in a humidified incubator with 5% CO2. The parasites were continuously cultured as previously described (20).
In vitro assessment of antiplasmodial activity
For the in vitro experiment, P. falciparum was synchronized at ring stage as previously described (21), bergenin (provided by the ShaanXi Institute for Food and Drug Control, Xi’an, China) was dissolved in dimethyl sulfoxide (DMSO, Sigma-Aldrich, St. Louis, MO, USA) and dispensed as 0.2 μl with various concentrations of bergenin stock solution in a 100-μl aliquot of cell suspension with 1% parasitemia and 2% haematocrit, inoculated in a 96-well plate. The positive control was treated with 0.2 μl of DMSO and the negative control was treated with 0.2 μl of artemether stock solution (Sigma-Aldrich) also dissolved in DMSO. The parasites were cultured for 72 h to ensure they went through all the stages of the cell cycle and parasite growth was evaluated using the lactate dehydrogenase (LDH) activity assay, as described below.
Evaluation of parasite growth using the LDH activity assay
After 72 h of incubation, the plate was frozen at −70°C overnight and thawed at room temperature to hemolyze red blood cells. A 25-μl aliquot was then dispensed into a new well. To evaluate LDH activity, 120 μl of freshly made reaction mix [143 mM 3-acetyl pyridine adenine dinucleotide (APAD, Sigma-Aldrich), 286 mg/ml diaphorase (2.83 U/ml, Worthington Biochemical Corporation, Lakewood, NJ, USA), 143 mM sodium L-lactate (Bio Basic Canada Inc., Ontario, Canada), 178.75 mM Nitro Blue tetrazolium chloride (NBT, Bio Basic Canada Inc.), 0.7% Tween 20, 100 mM Tris-HCl pH 8.0) (22) was dispensed and incubated at room temperature for 30 min. The plate was shaken for 10 sec and the absorbance was monitored at 650 nm (Model 680 Microplate Reader, Bio-Rad, Hercules, CA, USA). Data were normalized to percent parasite growth using negative and positive controls using the following equation: % parasite growth = (Awell−Aneg/Apos−Aneg) × 100.
In vivo assessment of antimalarial activity
Bergenin powder was suspended in 1% sodium carboxymethyl cellulose (CMC; Sinopharm Chemical Reagent Co., Ltd., Shanghai, China) and intragastrically administered to the mice. Plasmodium berghei (conserved in our laboratory; P. berghei)-infected BALB/c mice with parasitemia levels of ~1% were randomly divided into three groups, the phosphate-buffered saline (PBS), CMC and bergenin groups. Each group included 13 mice housed in separate cages. The mice from the bergenin group were intragastrically administered 800 mg/kg/day of bergenin for 4 or 6 days (100 μl twice daily). The mice in the PBS and CMC groups, which served as control, were treated with 100 μl of PBS and CMC, respectively. Thin blood smears were performed on days 4 and 6 after drug treatment and stained with Giemsa. The parasitemia was assessed by microscopic (Olympus Corp., Tokyo, Japan) examination.
In vitro assessment of cytotoxic activity
The HepG2 and HeLa cells were conserved in our laboratory and were cultured in DMEM (high glucose) supplemented with 10% fetal bovine serum at 37°C in a humidified incubator with 5% CO2. The cells were plated in a 96-well plate with 104 cells/well and the medium was replaced the following day by a 100-μl aliquot containing 0.5 μl of bergenin stock solution of various concentrations. The positive control was treated with 0.5 μl of DMSO. The cells were cultured for 72 h and cell viability was evaluated using the MTT assay (Bio Basic Canada Inc.) (23). The absorbance of each well was read on a microplate reader at 492 nm after 30 sec of shaking.
Statistical analysis
The half-maximal inhibitory concentration (IC50) of bergenin represented a 50% parasite LDH activity inhibition compared to the positive control, referred to as 100% parasite LDH activity. The half-maximal cytotoxic concentration (CC50) of bergenin represented a 50% cell growth inhibition compared to the positive control, referred to as 100% cell growth. IC50 and CC50 were estimated using GraphPad Prism 5 software (GraphPad Software Inc., La Jolla, CA, USA) (24).
Statistical analysis was performed using Statistics Toolbox of Matlab 7.1 (The MathWorks, Inc., Natick, MA, USA) and the differences among the groups were calculated using one-way ANOVA. Data are expressed as means ± SEM. P<0.05 was considered to indicate a statistically significant difference.
Results
Bergenin suppresses P. falciparum growth in vitro
Bergenin and artemisinin belong to the family of sesquiterpene lactones in structure (Fig. 1A). To demonstrate the antiplasmodial effect of bergenin, we evaluated the parasite growth after treatment with different concentrations of bergenin for 72 h with the LDH activity assay. As shown in Fig. 1B, bergenin at 10 μg/ml significantly inhibited the parasite growth compared to the DMSO control (P<0.05). The inhibitory effect of bergenin on parasite growth was dose-dependent in a range of 0.1–50 μg/ml and parasite growth was almost completely suppressed by bergenin at 50 μg/ml. The IC50 of bergenin was 14.1 μg/ml, which was in the range of moderate antiplasmodial activity (10<IC50<25 μg/ml) (25). Morphological changes in the uninfected erythrocytes were not observed, even following exposure to 50 μg/ml bergenin for 72 h (Fig. 1C). Furthermore, the group treated with an equal volume of DMSO (0.2%, v/v) exhibited no effect on parasite growth compared to normally cultured parasites (data not shown). Taken together, these results indicated that bergenin effectively suppresses parasite growth in vitro.
Figure 1.

Antiplasmodial activity of bergenin in vitro. (A) Molecular structures of artemisinin and bergenin (Ber). (B) Growth inhibition of Plasmodium falciparum (P. falciparum) by bergenin. P. falciparum samples were maintained in erythrocytes and treated with different concentrations of bergenin (0.1–50 μg/ml) for 72 h. The viability of the parasites was determined by the lactate dehydrogenase assay. Data are expressed as means ± SEM based on three independent experiments. *P<0.05 and **P<0.01 vs. dimethyl sulfoxide (DMSO) control. (C) Microscopic examination of the morphological change in P. falciparum induced by bergenin. Left panel, cell shape of red blood cells infected with P. falciparum treated with DMSO; right panel, P. falciparum-infected red blood cells treated with bergenin at 50 μg/ml for 72 h. Morphological changes in the uninfected erythrocytes were not observed during the trials, even when exposed to 50 μg/ml bergenin for 72 h. The data demonstrated that bergenin was not cytotoxic to erythrocytes at concentrations effective for inhibiting parasite growth. OD, optical density.
Bergenin is less cytotoxic to mammalian cells
To determine whether bergenin is cytotoxic to mammalian cells, we evaluated the viability of HeLa and HepG2 cells following treatment with serial concentrations of bergenin for 72 h with the MTT assay. As shown in Fig. 2, bergenin exhibited no detectable cytotoxicity against HeLa and HepG2 cells at concentrations of 1–5 mg/ml. The CC50 of bergenin was ~12.5 on HeLa cells and 19.1 mg/ml on HepG2 cells, which may be defined as non-toxic (CC50>30 μg/ml) (26).
Figure 2.
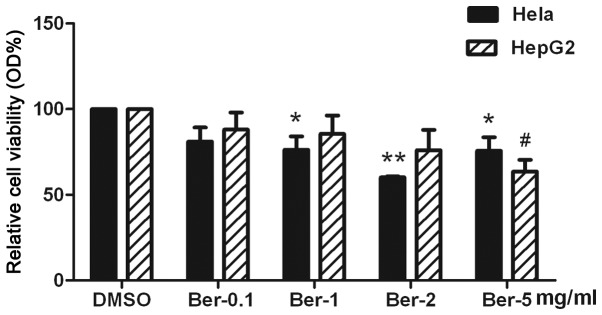
Cytotoxic activity of bergenin on HeLa and HepG2 cells. The cells were treated with different concentrations of bergenin (0.1–5 mg/ml) for 72 h. The cytotoxic activity was assessed with the MTT assay. Data are expressed as means ± SEM and the results represent three independent experiments. *P<0.05 and **P<0.01 vs. dimethyl sulfoxide (DMSO) control of HeLa cells. #P<0.05 vs. DMSO control of HepG2 cells. OD, optical density.
Bergenin inhibits parasite growth in vivo
To determine whether bergenin suppresses plasmodium growth in vivo, we treated P. berghei-infected mice with bergenin by oral administration at 800 mg/kg/day for 6 days. The mouse parasitemia was assessed on days 4 and 6 during the trial. As shown in Fig. 3, there was no significant difference in parasitemia among the PBS, CMC and bergenin groups on day 4. However, on day 6, the parasitemia in bergenin-treated mice decreased from 35 to 27%, with a statistically significant difference compared to the PBS or CMC control groups (P<0.01). These data indicated that bergenin suppresses the growth of P. berghei in vivo.
Figure 3.
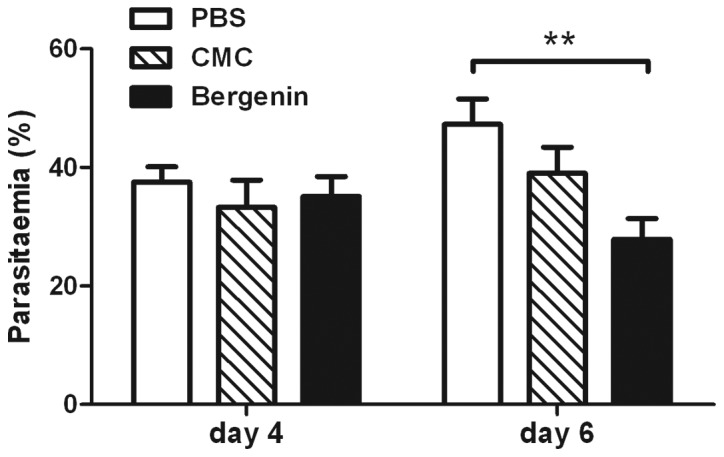
Growth inhibition of Plasmodium berghei (P. berghei) by bergenin in mice. P. berghei-infected mice were treated with phosphate-buffered saline (PBS), carboxymethyl cellulose (CMC), or bergenin for 4 and 6 days, followed by parasitemia assessment. Data are expressed as means ± SEM, generated from three independent experiments. **P<0.01.
The body appearance, ingestion and defecation of bergenin-treated mice appeared to be normal during the trials, with no detectable differences between the bergenin-treated, CMC or PBS control group mice, which indicated that bergenin exhibited good tolerability even at considerably high dosage (e.g., 800 mg/kg/day).
Discussion
Plants have been used as the basis of traditional medicine throughout history and they continue to serve as sources of numerous pharmaceuticals widely used today (27). Plant-derived antimalarial drugs, such as quinine, artemisinin, atovaquone and azithromycin, are currently the primary choice for malaria treatment. Although several plant-derived compounds cannot be used directly as therapeutic agents due to their low bioavailability or poor solubility, their chemical structure may be further improved, in terms of increased efficacy or target specificity, in order to develop new antimalarial drugs (28). Bergenin was isolated from R. aesculifolia Batal, which is one of the most common plants in China and has been reported to possess multiple pharmacological properties, including antifungal, anti-inflammatory and antioxidant (29). As bergenin was used in folk medicine to treat protozoal infection and fever, we hypothesized that it may also exhibit antimalarial activity.
Through parasite LDH activity assay and morphologic examination, we demonstrated that bergenin was able to kill cultured P. falciparum effectively with a considerably low dosage, whereas it exerted no toxic effect on normal erythrocytes and other mammalian cells, such as HepG2 or HeLa cells. Furthermore, the parasitemia in P. berghei-infected mice was significantly decreased following treatment with bergenin for 6 days. These data indicated that bergenin possesses antimalarial properties. However, compared to its high potency in vitro, bergenin exhibited limited antimalarial effectiveness in vivo, which may be explained as follows: i) the water solubility of bergenin is poor, which may significantly reduce the bioavailability of orally administered bergenin and poor water solubility may hinder drug development in ~40% of lead candidates (30); ii) it is well-known that there is currently no suitable small animal model for P. falciparum research; therefore, although we assessed the anti-P. falciparum activity in vitro and anti-P. berghei activity in vivo, the possibility that P. berghei exhibits a different response to bergenin compared to P. falciparum cannot be excluded; and iii) chemical modification may be necessary to enhance the bioavailability of bergenin. For example, the bioavailability of the bergenin-phospholipid complex was significantly increased to 439% compared to that of bergenin (31). In addition, bergenin pentaacetate was shown to exhibit considerable antibacterial activity, while bergenin was inactive (11). Those findings indicate that changing the physicochemical properties of bergenin to enhance its antimalariral activity may be feasible.
In addition, we assessed the possible cytotoxic effect of bergenin on normal erythrocytes and two other mammalian types of cells (HeLa and HepG2 cells). According to the bergenin maximal solubility in DMSO and the maximal tolerance of the cells for DMSO [the cells will die when DMSO reaches 1% (v/v)], the maximal dosage of bergenin was set to 5 mg/ml in our trials. The calculated CC50 of bergenin was >10 mg/ml (HeLa cells, 12.5 mg/ml and HepG2 cells, 19.1 mg/ml). Although the antiplasmodial IC50 of bergenin was found to be >10 μg/ml (14.1 μg/ml), the selectivity index (SI), defined as SI = CC50/IC50, was considerably high (HeLa cells, 887; HepG2 cells, 1,355), due to its good tolerability, which suggests that bergenin may be a promising novel antimalarial compound (SI>60) (32).
In summary, this study demonstrated the antimalarial activity of bergenin, a sesquiterpene lactone compound isolated from R. aesculifolia Batal, against P. falciparum in vitro and P. berghei in vivo. However, further investigation is required to establish bergenin as a novel lead compound for antimalarial drug research and development.
Acknowledgements
We would like to thank chief physician Zuoxun Shen (ShaanXi Institute for Food and Drug Control) for donating the bergenin. This study was supported by grants from the National Natural Science Foundation of China (nos. 81000744 and 30901250).
References
- 1.World Health Organization. World Malaria Report. 2011 http://www.who.int/malaria/world_malaria_report_2011/en/
- 2.Ashley EA, White NJ. Artemisinin-based combinations. Curr Opin Infect Dis. 2005;18:531–536. doi: 10.1097/01.qco.0000186848.46417.6c. [DOI] [PubMed] [Google Scholar]
- 3.World Health Organization. Guidelines for the treatment of malaria. http://www.who.int/malaria/docs/TreatmentGuidelines2006.pdf.
- 4.Phyo AP, Nkhoma S, Stepniewska K, et al. Emergence of artemisinin-resistant malaria on the western border of Thailand: a longitudinal study. Lancet. 2012;379:1960–1966. doi: 10.1016/S0140-6736(12)60484-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Noedl H, Se Y, Schaecher K, et al. Artemisinin Resistance in Cambodia 1 (ARC1) Study Consortium. Evidence of artemisinin-resistant malaria in western Cambodia. N Engl J Med. 2008;359:2619–2620. doi: 10.1056/NEJMc0805011. [DOI] [PubMed] [Google Scholar]
- 6.Dondorp AM, Nosten F, Yi P, et al. Artemisinin resistance in Plasmodium falciparum malaria. N Engl J Med. 2009;361:455–467. doi: 10.1056/NEJMoa0808859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Willcox ML, Bodeker G. Traditional herbal medicines for malaria. BMJ. 2004;329:1156–1159. doi: 10.1136/bmj.329.7475.1156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Saxena S, Pant N, Jain DC, Bhakuni RS. Antimalarial agents from plant sources. Curr Sci. 2003;85:1314–1329. [Google Scholar]
- 9.Kaur K, Jain M, Kaur T, Jain R. Antimalarials from nature. Bioorg Med Chem. 2009;17:3229–3256. doi: 10.1016/j.bmc.2009.02.050. [DOI] [PubMed] [Google Scholar]
- 10.Raj MK, Duraipandiyan V, Agustin P, Ignacimuthu S. Antimicrobial activity of bergenin isolated from Peltophorum pterocarpum DC. flowers. Asian Pac J Trop Biomed. 2012;2:S901–S904. [Google Scholar]
- 11.Nazir N, Koul S, Qurishi MA, Najar MH, Zargar MI. Evaluation of antioxidant and antimicrobial activities of bergenin and its derivatives obtained by chemoenzymatic synthesis. Eur J Med Chem. 2011;46:2415–2420. doi: 10.1016/j.ejmech.2011.03.025. [DOI] [PubMed] [Google Scholar]
- 12.Da Silva SL, Oliveira VG, Yano T, Nunomura RCS. Antimicrobial activity of bergenin from Endopleura uchi(Huber) Cuatrec. Acta Amazonica. 2009;39:187–192. [Google Scholar]
- 13.Piacente S, Pizza C, De Tommasi N, Mahmood N. Constituents of Ardisia japonica and their in vitro anti-HIV activity. J Nat Prod. 1996;59:565–569. doi: 10.1021/np960074h. [DOI] [PubMed] [Google Scholar]
- 14.Nunomura RCS, Oliveira VG, Da Silva SL, Nunomura SM. Characterization of bergenin in Endopleura uchi bark and its anti-inflammatory activity. J Brazil Chem Soc. 2009;20:1060–1064. [Google Scholar]
- 15.Nazir N, Koul S, Qurishi MA, Taneja SC, Ahmad SF, Bani S, Qazi GN. Immunomodulatory effect of bergenin and norbergenin against adjuvant-induced arthritis - a flow cytometric study. J Ethnopharmacol. 2007;112:401–405. doi: 10.1016/j.jep.2007.02.023. [DOI] [PubMed] [Google Scholar]
- 16.De Abreu HA, Aparecida Dos S, Lago I, Souza GP, Pilo-Veloso D, Duarte HA, de C Alcantara AF. Antioxidant activity of (+)-bergenin: a phytoconstituent isolated from the bark of Sacoglottis uchi Huber (Humireaceae) Org Biomol Chem. 2008;6:2713–2718. doi: 10.1039/b804385j. [DOI] [PubMed] [Google Scholar]
- 17.Lim HK, Kim HS, Choi HS, Oh S, Choi J. Hepatoprotective effects of bergenin, a major constituent of Mallotus japonicus, on carbon tetrachloride-intoxicated rats. J Ethnopharmacol. 2000;72:469–474. doi: 10.1016/s0378-8741(00)00260-9. [DOI] [PubMed] [Google Scholar]
- 18.Takahashi H, Kosaka M, Watanabe Y, Nakade K, Fukuyama Y. Synthesis and neuroprotective activity of bergenin derivatives with antioxidant activity. Bioorg Med Chem. 2003;11:1781–1788. doi: 10.1016/s0968-0896(02)00666-1. [DOI] [PubMed] [Google Scholar]
- 19.Goel RK, Maiti RN, Manickam M, Ray AB. Antiulcer activity of naturally occurring pyrano-coumarin and isocoumarins and their effect on prostanoid synthesis using human colonic mucosa. Indian J Exp Biol. 1997;35:1080–1083. [PubMed] [Google Scholar]
- 20.Trager W, Jensen JB. Human malaria parasites in continuous culture. Science. 1976;193:673–675. doi: 10.1126/science.781840. [DOI] [PubMed] [Google Scholar]
- 21.Arnot DE, Ronander E, Bengtsson DC. The progression of the intra-erythrocytic cell cycle of Plasmodium falciparum and the role of the centriolar plaques in asynchronous mitotic division during schizogony. Int J Parasitol. 2011;41:71–80. doi: 10.1016/j.ijpara.2010.07.012. [DOI] [PubMed] [Google Scholar]
- 22.Gamo FJ, Sanz LM, Vidal J, et al. Thousands of chemical starting points for antimalarial lead identification. Nature. 2010;465:305–310. doi: 10.1038/nature09107. [DOI] [PubMed] [Google Scholar]
- 23.Lazaro JE, Gay F. Plasmodium falciparum: in vitro cytotoxicity testing using MTT. J Biomol Screen. 1998;3:49–53. [Google Scholar]
- 24.Saha S, Verma R. Inhibitory potential of traditional herbs on alpha-amylase activity. Pharm Biol. 2012;50:326–331. doi: 10.3109/13880209.2011.608075. [DOI] [PubMed] [Google Scholar]
- 25.Mbatchi SF, Mbatchi B, Banzouzi JT, Bansimba T, Nsonde Ntandou GF, Ouamba JM, Berry A, Benoit-Vical F. In vitro antiplasmodial activity of 18 plants used in Congo Brazzaville traditional medicine. J Ethnopharmacol. 2006;104:168–174. doi: 10.1016/j.jep.2005.08.068. [DOI] [PubMed] [Google Scholar]
- 26.Malebo HM, Tanja W, Cal M, Swaleh SA, Omolo MO, Hassanali A, Sequin U, Hamburger M, Brun R, Ndiege IO. Antiplasmodial, anti-trypanosomal, anti-leishmanial and cytotoxicity activity of selected Tanzanian medicinal plants. Tanzan J Health Res. 2009;11:226–234. doi: 10.4314/thrb.v11i4.50194. [DOI] [PubMed] [Google Scholar]
- 27.Newman DJ, Cragg GM. Natural products as sources of new drugs over the last 25 years. J Nat Prod. 2007;70:461–477. doi: 10.1021/np068054v. [DOI] [PubMed] [Google Scholar]
- 28.Ginsburg H, Deharo E. A call for using natural compounds in the development of new antimalarial treatments - an introduction. Malar J. 2011;10(Suppl 1):S1. doi: 10.1186/1475-2875-10-S1-S1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Patel DK, Patel K, Kumar R, Gadewar M, Tahilyani V. Pharmacological and analytical aspects of bergenin: a concise report. Asian Pac J Trop Dis. 2012;2:163–167. doi: 10.1016/S2221-1691(12)60239-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Merisko-Liversidge EM, Liversidge GG. Drug nanoparticles: formulating poorly water-soluble compounds. Toxicol Pathol. 2008;36:43–48. doi: 10.1177/0192623307310946. [DOI] [PubMed] [Google Scholar]
- 31.Qin X, Yang Y, Fan TT, Gong T, Zhang XN, Huang Y. Preparation, characterization and in vivo evaluation of bergenin-phospholipid complex. Acta Pharmacol Sin. 2010;31:127–136. doi: 10.1038/aps.2009.171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Upadhyay HC, Sisodia BS, Verma RK, Darokar MP, Srivastava SK. Antiplasmodial potential of extracts from two species of genus Blumea. Pharm Biol. 2013;51:1326–1330. doi: 10.3109/13880209.2013.790453. [DOI] [PubMed] [Google Scholar]