

Abstract
Renal cell carcinoma (RCC) is characterized by diverse clinical manifestations, few early warning signs and a resistance to radiotherapy and chemotherapy. Although several clinical trials have investigated potential effective therapeutic strategies for RCC, the chemoresistance of RCC has not yet been overcome. An endogenous ligand for the peroxisome proliferator-activated receptor-γ (PPARγ), 15-deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2), was shown to induce apoptosis in RCC. The aim of the present study was to investigate the synergistic effects of carcinostatics on the antitumor activity of 15d-PGJ2 in the Caki-2 human RCC cell line with the MTT assay. Our results demonstrated that the topoisomerase-II inhibitor etoposide (VP-16) exhibited cytotoxic effects synergistically with 15d-PGJ2. Furthermore, the presence of the PPARγ antagonist GW9662 did not protect Caki-2 cells against 15d-PGJ2-induced cytotoxicity. Additionally, it was observed that the combined treatment of VP-16 and 15d-PGJ2 activated caspase-3 more efficiently compared to each treatment alone. Therefore, the combined treatment with 15d-PGJ2 and VP-16 exhibited synergistic antitumor activity independently of PPARγ.
Keywords: etoposide; 15-deoxy-Δ12,14-prostaglandin J2; renal cell carcinoma
Introduction
Renal cell carcinoma (RCC) accounts for ~2% of all cancer cases and is characterized by diverse clinical manifestations, few early warning signs and a resistance to radiotherapy and chemotherapy (1). Clear cell RCC accounts for the majority of RCC cases (2) and one-third of the patients present with metastases at initial diagnosis. Due to the resistance of RCC to radiotherapy and chemotherapy, the 5-year survival rate for patients with metastatic RCC is <10% (3). The responsiveness of RCC to treatment with conventional anticancer agents, such as 5-fluorouracil (5-FU) and cisplatin (CDDP), was reported to be lower compared to other types of cancer (4,5). Despite the development of various chemotherapeutic strategies, RCC remains a challenging tumor entity. A few patients were reported to exhibit complete or partial response to frequently used chemotherapeutic agents, such as gemcitabine, 5-FU, capecitabine and vinblastine (6). As RCC is known to be immunogenic, several clinical trials investigated the potency of cytokines, mainly interleukin 2 and/or interferon-α (7,8). Targeted therapies, including monoclonal antibodies and small-molecule inhibitors, have significantly modified the treatment of cancer over the last 10 years through inhibiting tyrosine kinase activity or vascular endothelial growth factor receptors (9). However, despite these novel therapies, the clinical outcome of patients with metastatic RCC remains poor (6). Thus, there is a pressing need to establish alternative therapeutic modalities against RCC.
In our previous study, we reported that the topoisomerase-I inhibitor camptothecin exhibited toxicity synergistically with a peroxisome proliferator-activated receptor-γ (PPARγ) agonist (10). 15-Deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2) is an endogenous carcinostatic agent, whose nuclear receptor is a PPARγ. PPARγ activation was shown to induce growth inhibition in human RCC cells (11). Furthermore, 15d-PGJ2 was also implicated in antiproliferation independently of PPARγ (12). The antitumor activity of 15d-PGJ2 was also found to be associated with the inhibition of topoisomerase-II (13). We previously identified novel binding sites for 15d-PGJ2 on the cell surface (14). With regard to targets for 15d-PGJ2 in the plasma membrane, molecular chaperones, glycolytic enzymes and cytoskeletal components, such as β-actin, were also identified (15). PPARγ agonists were shown to enhance 5-FU-, CDDP- or topoisomerase-II inhibitor-induced apoptosis in cancer cell types other than RCC (16–19). The aim of the present study was to evaluate the therapeutic efficacy of the combination treatment with 15d-PGJ2 and the topoisomerase-II inhibitor etoposide (VP-16) in RCC.
Materials and methods
Cell lines and cell culture
The Caki-2 human RCC cell line was obtained from Summit Pharmaceuticals International (Tokyo, Japan). The Caki-2 cells were routinely cultured in RPMI-1640 medium supplemented with 10% fetal bovine serum, 50 mg/l penicillin G and 50 mg/l streptomycin (Invitrogen, Tokyo, Japan), at 37°C in a 5% CO2 atmosphere.
Reagents
15d-PGJ2 was obtained from Cayman Chemicals (Ann Arbor, MI; Cabru, Milan, Italy). Etoposide (VP-16) was purchased from Wako Pure Chemical Industries, Ltd. (Osaka, Japan). GW9662 was obtained from Sigma-Aldrich (St. Louis, MO, USA); and MTT was purchased from Dojindo Laboratories (Kumamoto, Japan).
Cell viability analysis
To evaluate the effects of 15d-PGJ2 and VP-16, alone or in combination, on the growth of Caki-2 cells, cell viability was determined by the MTT assay. The cells were seeded on a 96-well tissue culture plate at 10,000 cells/cm2 and incubated for 24 h prior to drug exposure. The cells were incubated with 15d-PGJ2 and VP-16 at increasing concentrations (0, 10, 20, 30, 40 and 50 μM of 15d-PGJ2; and 0, 10, 20, 30, 40, 50, 60 and 70 μM of VP-16) for 24 h. After 24 h, the cells were incubated with MTT solution (0.1 mg/ml in phosphate-buffered saline) for an additional 3 h at 37°C. The MTT solution was then aspirated off. To dissolve the formazan crystals formed in viable cells, 100 μl dimethyl sulfoxide was added to each well. Absorbance was measured at 570 nm using a spectrophotometer (iMark Microplate Reader, Bio-Rad Laboratories, Hercules, CA, USA).
Detection of chromatin condensation (fluorescence microscopy)
For nuclear staining, the cells were treated with 15d-PGJ2 and VP-16 for 24 h. Immediately following treatment, the nuclear chromatin of trypsinized cells was stained with 80 μg/ml Hoechst 33342 (Nacalai Tesque, Kyoto, Japan) for 15 min at room temperature. The cells were then observed under a brightfield fluorescent microscope (Vanox; Olympus, Tokyo, Japan) under UV excitation. The percentage of chromatin-condensed cells was determined by counting >100 cells in each experiment.
Fluorimetric assay of caspase-3 activity
Caspase-3 activity was assessed using a Caspase-3 Fluorimetric Assay kit, (Sigma-Aldrich), according to the manufacturer’s instructions. Briefly, the cells were seeded into 96-well plates at a density of 10,000 cells/cm2 and incubated with 15d-PGJ2 and VP-16 for 24 h. After exposure to the drugs for 24 h, the supernatants were aspirated and the cells were harvested with lysis buffer [50 mM HEPES (pH 7.4), 5 mM CHAPS and 5 mM DTT]. The reaction buffer, including acetyl-Asp-Glu-Val-Asp-7-amido4-methylcoumarin (Ac-DEVD-AMC), a caspase-3 specific substrate, was added to the wells and the production of AMC was sequentially detected with a CytoFluor® Plate reader (MTX Lab Systems, Vienna, VA, USA) at an excitation wavelength of 360 nm and at an emission wavelength of 460 nm. The enzyme activities were determined as initial velocities expressed as nmol AMC/min/ml and were then corrected by the quantity of protein in each well detected by bicinchoninic acid protein assays (Thermo Fisher Scientific, Waltham, MA, USA).
Statistical analysis
Data were statistically analyzed with the Student’s t-test for comparison with the control group and are expressed as means ± SEM. Data on various drugs were statistically analyzed by one-way analysis of variance, followed by Scheffe’s F test for comparison between the groups.
Results
VP-16 enhanced the antiproliferative effects of 15d-PGJ2 in Caki-2 cells
RCCs are chemoresistant to conventional anticancer agents (3), but are sensitive to the endogenous anticancer agent 15d-PGJ2 (20). It was previously confirmed that 15d-PGJ2 induced apoptosis in RCCs (10,11,20,21). Therefore, we investigated the cytotoxic effects of 15d-PGJ2 in the Caki-2 human RCC cell line with the MTT assay. Following incubation with 15d-PGJ2 for 24 h, we observed that the viability of Caki-2 cells was significantly reduced in a dose-dependent manner (from 30 to 50 μM; P<0.01; Fig. 1A). By contrast, incubation with VP-16 alone (10–70 μM) for 24 h did not affect the viability of Caki-2 cells (Fig. 1B).
Figure 1.
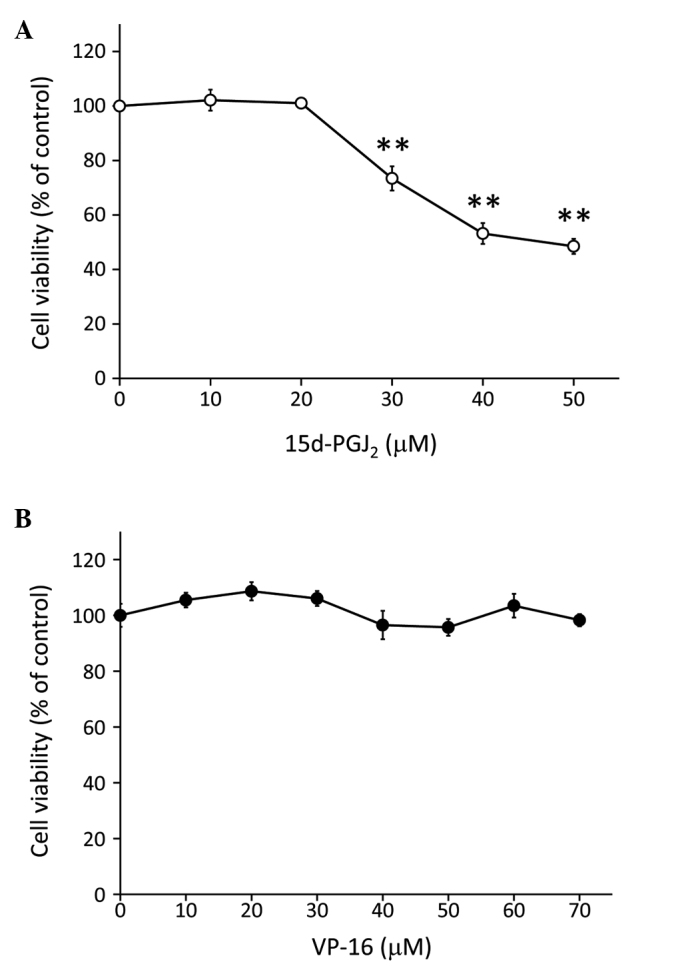
Treatment with 15d-PGJ2 inhibited the proliferation of Caki-2 cells, but treatment with VP-16 alone did not affect cell viability. The cells were assayed for viability using the MTT assay after treatment for 24 h with increasing doses of (A) 15d-PGJ2 (0, 10, 20, 30, 40 and 50 μM) and (B) VP-16 (0, 10, 20, 30, 40, 50, 60 and 70 μM). **P<0.01 vs. control cells. Data are expressed as means ± standard error of the mean of three independent experiments. 15d-PGJ2, 15-deoxy-Δ12,14-prostaglandin J2; VP-16, etoposide.
To investigate the synergistic cytotoxicity of VP-16 and 15d-PGJ2, we exposed Caki-2 cells to VP-16 and 15d-PGJ2 combination treatment (Fig. 2). The Caki-2 cells were co-treated with VP-16 (10–70 μM) and 15d-PGJ2 (20 μM) simultaneously and we observed that cell viability was significantly lower compared to that of cells treated with 15d-PGJ2 alone (Fig. 2A). Furthermore, the phase contrast microscopy analysis indicated that the combination of VP-16 and 15d-PGJ2 induced more prominent morphological changes compared to 15d-PGJ2 alone (Fig. 2B). Caki-2 cell cultures treated with either VP-16 or 15d-PGJ2 exhibited marginally broadened cells, whereas cells treated with VP-16 and 15d-PGJ2 exhibited significant atrophy. As previously reported, topoisomerase-II activity is inhibited by 15d-PGJ2 (13). Thus, the synergistic antitumor activity of 15d-PGJ2 and VP-16 may be mediated via the topoisomerase-II inhibition pathway.
Figure 2.
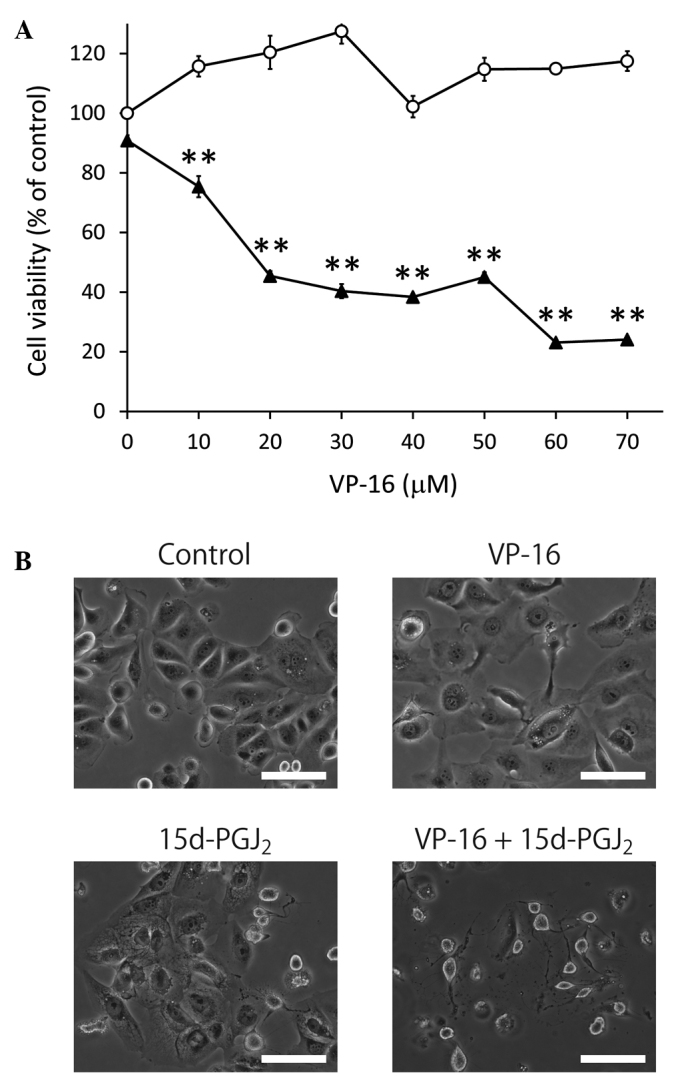
Treatment with VP-16 enhanced the antiproliferative effect of 15d-PGJ2 in Caki-2 cells. (A) The cells were assayed for viability using MTT following treatment with VP-16 (0, 10, 20, 30, 40, 50, 60 and 70 μM) (open circles) and combination treatment with VP-16 and 15d-PGJ2 (20 μM) (closed triangles) for 24 h. The results are expressed as the means ± SEM of three independent experiments. **P<0.01, vs. control cells. (B) The combination of 15d-PGJ2 and VP-16 induced morphological changes in Caki-2 cells. The cells were treated with 15d-PGJ2 alone (20 μM), VP-16 alone (70 μM) and the combination of the two. Caki-2 cells were then examined by phase contrast microscopy following 24 h of incubation. Scale bar, 100 μm. 15d-PGJ2, 15-deoxy-Δ12,14-prostaglandin J2; VP-16, etoposide.
15d-PGJ2 enhanced VP-16-induced apoptosis via the activation of caspase-3
To elucidate whether the inhibition of cell proliferation induced by the combined treatment of VP-16 and 15d-PGJ2 is associated with apoptosis, we assessed nuclear chromatin condensation in Caki-2 cells treated with VP-16 (70 μM) and/or 15d-PGJ2 (20 μM) (Fig. 3A). Treatment with either VP-16 or 15d-PGJ2 exhibited a tendency to increase chromatin condensation, whereas a combination of the two was found to strongly induce chromatin condensation (P<0.05). We then assessed caspase-3 activity in Caki-2 cells treated with VP-16 (70 μM) and/or 15d-PGJ2 (20 μM) (Fig. 3B). Cells treated with either VP-16 or 15d-PGJ2 exhibited a tendency to activate caspase-3, whereas the combination of the two significantly induced caspase-3 activation (P<0.05). These results suggested that 15d-PGJ2 enhanced VP-16-induced apoptosis via the activation of caspase-3. Topoisomerase-II inhibitor-induced apoptosis was shown to be mediated by caspase-3 (22). Thus, the synergistic inhibition of topoisomerase-II by the combination of 15d-PGJ2 and VP-16 may induce caspase-3 activation.
Figure 3.
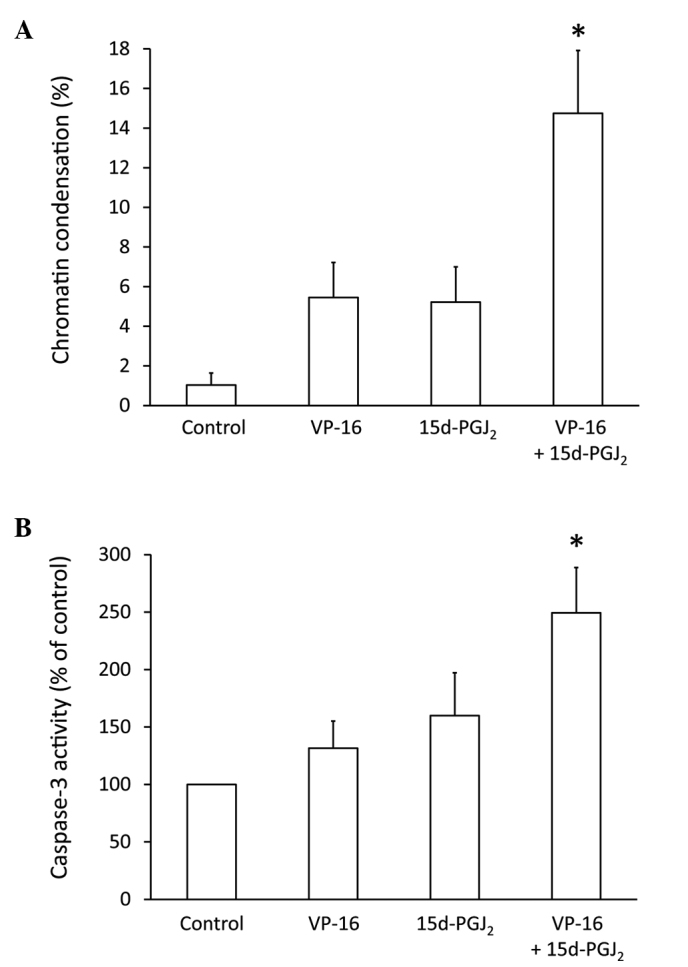
Caki-2 cells were assayed for (A) nuclear chromatin condensation and (B) caspase-3 activity following treatment for 24 h with 15d-PGJ2 alone (20 μM), VP-16 alone (70 μM) and the combination of the two. Treatment with 15d-PGJ2 enhanced the VP-16-induced Caki-2 cell apoptosis via the activation of caspase-3. The results are expressed as the means ± SEM of three independent experiments. *P<0.05 vs. control cells. 15d-PGJ2, 15-deoxy-Δ12,14-prostaglandin J2; VP-16, etoposide.
15d-PGJ2 enhanced the antitumor activity of VP-16 independently of PPARγ
It was previously reported that 15d-PGJ2 treatment inhibits cell proliferation in several types of cancer cells via PPARγ (23–26). In addition, topoisomerase-II inhibitors enhance the cytotoxicity of 15d-PGJ2 in RCC. However, whether the inhibition of topoisomerase-II and the activation of PPARγ result in synergistic toxicity, has not been fully elucidated. To determine whether 15d-PGJ2 enhanced the antitumor activity of VP-16 via PPARγ activation, Caki-2 cells were co-treated with VP-16 (70 μM), 15d-PGJ2 (20 μM) and the PPARγ inhibitor, GW9662 (0.1–50 μM) (Fig. 4). Our results demonstrated that the synergistic cytotoxic effects of VP-16 and 15d-PGJ2 combination treatment were not decreased by PPARγ inhibition, suggesting that 15d-PGJ2 enhanced the antitumor activity of VP-16 independently of PPARγ. 15d-PGJ2 was also reported to induce apoptosis via the activation of caspase-3 independently of PPARγ (26).
Figure 4.

The synergistic cytotoxicity of 15d-PGJ2 and VP-16 on the proliferation of Caki-2 cells was not associated with PPARγ. The cells were assayed for viability using the MTT assay following treatment with VP-16 alone (70 μM) (open circles) and with the combination of VP-16 (70 μM), 15d-PGJ2 (20 μM) and GW9662 (0, 0.1, 0.5, 1, 5, 10 and 50 μM) (closed triangles) for 24 h. The results are expressed as means ± standard error of the mean of three independent experiments. 15d-PGJ2, 15-deoxy-Δ12,14-prostaglandin J2; VP-16, etoposide.
Discussion
In the present study, we demonstrated that the topoisomerase-II inhibitor, VP-16, enhanced the cytotoxicity of 15d-PGJ2 in RCC. Moreover, the PPARγ antagonist, GW9662, did not protect Caki-2 cells against 15d-PGJ2-induced cytotoxicity. These findings suggested that 15d-PGJ2 exhibited synergistic antitumor activity with VP-16 independently of the PPARγ pathway.
RCCs are chemoresistant to conventional anticancer agents (3), but are sensitive to the endogenous anticancer agent 15d-PGJ2 (20). Several previous studies confirmed that 15d-PGJ2 induced apoptosis in RCCs (11,20,21). Additionally, the responsiveness of RCC cells to treatment with 5-FU and CDDP was found to be lower compared to that of other types of cancer cells (4,5), whereas cancer cells other than RCC cells were found to be sensitive to conventional anticancer agents when co-treated with 15d-PGJ2 (27).
Topoisomerase-II inhibitors enhance the cytotoxicity of 15d-PGJ2 in RCC. However, whether the inhibition of topoisomerase-II and the activation of PPARγ synergistically produce toxicity, has not been fully elucidated. The PPARγ antagonist, GW9662, did not affect the responsiveness of RCC to the combined treatment with 15d-PGJ2 and VP-16, suggesting that 15d-PGJ2 exhibited synergistic antitumor activity with VP-16 independently of PPARγ. Topoisomerase-II was shown to be inhibited by 15d-PGJ2 (13). Thus, 15d-PGJ2 may exhibit synergistic antitumor activity with VP-16 via the inhibition of topoisomerase-II. Topoisomerase-II introduces double-strand breaks in DNA, which may subsequently be converted into chromosomal damage following chromatin condensation (28). In this study, increased chromatin condensation was observed following 15d-PGJ2 and VP-16 combination treatment for RCC. However, chromatin condensation was not significantly increased following treatment with 15d-PGJ2 alone. Furthermore, 15d-PGJ2 treatment induced marked morphological changes in Caki-2 cells, whereas treatment with VP-16 alone did not affect cell morphology. Cytoskeletal proteins are responsible for maintaining cell morphology. The effects of 15d-PGJ2 on the organization of the actin cytoskeleton were shown to be mediated by a direct covalent modification of proteins through electrophilic cyclopentenone binding (15,29). It has been hypothesized that VP-16 and 15d-PGJ2 induce chromatin condensation and morphological changes via the inhibition of topoisomerase-II and the disruption of the actin cytoskeleton, respectively.
It was reported that 15d-PGJ2 may induce apoptosis via the activation of caspase-3 independently of PPARγ (26). Topoisomerase-II inhibitor-induced apoptosis is also mediated by caspase-3 (30). In the present study, 15d-PGJ2 and VP-16 increased caspase-3 activity, both individually and synergistically.
In conclusion, we demonstrated that 15d-PGJ2 and VP-16 synergistically inhibited the proliferation of RCC independently of the PPARγ pathway. Furthermore, 15d-PGJ2 enhanced VP-16-induced apoptosis. We hypothesized that 15d-PGJ2 induced changes in cell morphology independent of the PPARγ pathway and that VP-16 induced chromatin condensation via topoisomerase-II inhibition; thus, the combination of 15d-PGJ2 and VP-16 exerted synergistic anticancer effects involving caspase-3 activation. Our results suggested that the 15d-PGJ2 and VP-16 combination treatment may be a novel chemotherapeutic option for the treatment of RCC.
Acknowledgements
This study was supported by a Grants-in-Aid for Scientific Research from the Japan Society for the Promotion of Science (grant no. 25860072). The authors would like to thank Tsutomu Minami, Daichi Yamaki, Yoshiya Kobayasi, Akihiro Yamada and Tomonori Nakagawa from the Hyogo Prefectural Kobe High School for supporting this study.
References
- 1.Motzer RJ, Bander NH, Nanus DM. Renal-cell carcinoma. N Engl J Med. 1996;335:865–875. doi: 10.1056/NEJM199609193351207. [DOI] [PubMed] [Google Scholar]
- 2.Costa LJ, Drabkin HA. Renal cell carcinoma: new developments in molecular biology and potential for targeted therapies. Oncologist. 2007;12:1404–1415. doi: 10.1634/theoncologist.12-12-1404. [DOI] [PubMed] [Google Scholar]
- 3.Mickisch GH. Gene therapy on renal-cell carcinoma: magic bullet or tragic insanity? World J Urol. 1995;13:178–185. doi: 10.1007/BF00184876. [DOI] [PubMed] [Google Scholar]
- 4.Chen XL, Cao LQ, She MR, Wang Q, Huang XH, Fu XH. Gli-1 siRNA induced apoptosis in Huh7 cells. World J Gastroenterol. 2008;14:582–589. doi: 10.3748/wjg.14.582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Jin KL, Park JY, Noh EJ, et al. The effect of combined treatment with cisplatin and histone deacetylase inhibitors on HeLa cells. J Gynecol Oncol. 2010;21:262–268. doi: 10.3802/jgo.2010.21.4.262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Motzer RJ, Russo P. Systemic therapy for renal cell carcinoma. J Urol. 2000;163:408–417. [PubMed] [Google Scholar]
- 7.Aso Y, Tazaki H, Umeda T, Marumo K. Treatment of renal cell carcinoma with systemic administration of intermediate doses of recombinant human interleukin-2 alone. Recombinant Human Interleukin-2 (S-6820) Research Group on Renal Cell Carcinoma. Prog Clin Biol Res. 1989;303:681–688. [PubMed] [Google Scholar]
- 8.Hayashi T, Miyagawa Y, Tsujimura A, Nonomura N, Minami M, Okuyama A. A case of renal cell carcinoma with multiple lung metastases refractory to interferon-α showing complete remission by interleukin-2 monotherapy. Int J Urol. 2006;13:805–808. doi: 10.1111/j.1442-2042.2006.01408.x. [DOI] [PubMed] [Google Scholar]
- 9.Logan JE, Rampersaud EN, Sonn GA, et al. Systemic therapy for metastatic renal cell carcinoma: a review and update. Rev Urol. 2012;14:65–78. [PMC free article] [PubMed] [Google Scholar]
- 10.Yamamoto Y, Fujita M, Koma H, et al. 15-Deoxy-Δ12,14-prostaglandin J2enhanced the anti-tumor activity of camptothecin against renal cell carcinoma independently of topoisomerase-II and PPARγ pathways. Biochem Biophys Res Commun. 2011;410:563–567. doi: 10.1016/j.bbrc.2011.06.026. [DOI] [PubMed] [Google Scholar]
- 11.Inoue K, Kawahito Y, Tsubouchi Y, et al. Expression of peroxisome proliferator-activated receptor gamma in renal cell carcinoma and growth inhibition by its agonists. Biochem Biophys Res Commun. 2001;287:727–732. doi: 10.1006/bbrc.2001.5640. [DOI] [PubMed] [Google Scholar]
- 12.Ward JE, Gould H, Harris T, Bonacci JV, Stewart AG. PPARγ ligands, 15-deoxy-Δ12,14-prostaglandin J2and rosiglitazone regulate human cultured airway smooth muscle proliferation through different mechanisms. Br J Pharmacol. 2004;141:517–525. doi: 10.1038/sj.bjp.0705630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Suzuki K, Uyeda M. Inhibitory properties of antitumor prostaglandins against topoisomerases. Biosci Biotechnol Biochem. 2002;66:1706–1712. doi: 10.1271/bbb.66.1706. [DOI] [PubMed] [Google Scholar]
- 14.Yagami T, Ueda K, Asakura K, et al. Novel binding sites of 15-deoxy-Δ12,14-prostaglandin J2in plasma membranes from primary rat cortical neurons. Exp Cell Res. 2003;291:212–227. doi: 10.1016/s0014-4827(03)00369-0. [DOI] [PubMed] [Google Scholar]
- 15.Yamamoto Y, Takase K, Kishino J, et al. Proteomic identification of protein targets for 15-deoxy-Δ12,14-prostaglandin J2in neuronal plasma membrane. PLoS One. 2011;6:e17552. doi: 10.1371/journal.pone.0017552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhang YQ, Tang XQ, Sun L, et al. Rosiglitazone enhances fluorouracil-induced apoptosis of HT-29 cells by activating peroxisome proliferator-activated receptor γ. World J Gastroenterol. 2007;13:1534–1540. doi: 10.3748/wjg.v13.i10.1534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hamaguchi N, Hamada H, Miyoshi S, et al. In vitro and in vivo therapeutic efficacy of the PPARγ agonist troglitazone in combination with cisplatin against malignant pleural mesothelioma cell growth. Cancer Sci. 2010;101:1955–1964. doi: 10.1111/j.1349-7006.2010.01632.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tikoo K, Kumar P, Gupta J. Rosiglitazone synergizes anticancer activity of cisplatin and reduces its nephrotoxicity in 7,12-dimethyl benz{a}anthracene (DMBA) induced breast cancer rats. BMC Cancer. 2009;9:107. doi: 10.1186/1471-2407-9-107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kanbe E, Abe A, Towatari M, Kawabe T, Saito H, Emi N. DR1-like element in human topoisomerase IIα gene involved in enhancement of etoposide-induced apoptosis by PPARγ ligand. Exp Hematol. 2003;31:300–308. doi: 10.1016/s0301-472x(03)00003-1. [DOI] [PubMed] [Google Scholar]
- 20.Yuan J, Takahashi A, Masumori N, et al. Ligands for peroxisome proliferator-activated receptor gamma have potent antitumor effect against human renal cell carcinoma. Urology. 2005;65:594–599. doi: 10.1016/j.urology.2004.10.019. [DOI] [PubMed] [Google Scholar]
- 21.Yoshimura R, Matsuyama M, Hase T, et al. The effect of peroxisome proliferator-activated receptor-γ ligand on urological cancer cells. Int J Mol Med. 2003;12:861–865. [PubMed] [Google Scholar]
- 22.Khelifa T, Beck WT. Merbarone, a catalytic inhibitor of DNA topoisomerase II, induces apoptosis in CEM cells through activation of ICE/CED-3-like protease. Mol Pharmacol. 1999;55:548–556. [PubMed] [Google Scholar]
- 23.Brockman JA, Gupta RA, Dubois RN. Activation of PPARγ leads to inhibition of anchorage-independent growth of human colorectal cancer cells. Gastroenterology. 1998;115:1049–1055. doi: 10.1016/s0016-5085(98)70072-1. [DOI] [PubMed] [Google Scholar]
- 24.Chen YX, Zhong XY, Qin YF, Bing W, He LZ. 15d-PGJ2inhibits cell growth and induces apoptosis of MCG-803 human gastric cancer cell line. World J Gastroenterol. 2003;9:2149–2153. doi: 10.3748/wjg.v9.i10.2149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Clay CE, Atsumi GI, High KP, Chilton FH. Early de novo gene expression is required for 15-deoxy-Δ12,14-prostaglandin J2-induced apoptosis in breast cancer cells. J Biol Chem. 2001;276:47131–47135. doi: 10.1074/jbc.C100339200. [DOI] [PubMed] [Google Scholar]
- 26.Li L, Tao J, Davaille J, et al. 15-deoxy-Δ12,14-prostaglandin J2induces apoptosis of human hepatic myofibroblasts. A pathway involving oxidative stress independently of peroxisome-proliferator-activated receptors. J Biol Chem. 2001;276:38152–38158. doi: 10.1074/jbc.M101980200. [DOI] [PubMed] [Google Scholar]
- 27.McClay EF, Winski PJ, Jones JA, Jennerette J, III, Gattoni-Celli S. Δ12-Prostaglandin-J2is cytotoxic in human malignancies and synergizes with both cisplatin and radiation. Cancer Res. 1996;56:3866–3869. [PubMed] [Google Scholar]
- 28.Huang X, Okafuji M, Traganos F, Luther E, Holden E, Darzynkiewicz Z. Assessment of histone H2AX phosphorylation induced by DNA topoisomerase I and II inhibitors topotecan and mitoxantrone and by the DNA cross-linking agent cisplatin. Cytometry A. 2004;58:99–110. doi: 10.1002/cyto.a.20018. [DOI] [PubMed] [Google Scholar]
- 29.Aldini G, Carini M, Vistoli G, et al. Identification of actin as a 15-deoxy-Δ12,14-prostaglandin J2target in neuroblastoma cells: mass spectrometric, computational, and functional approaches to investigate the effect on cytoskeletal derangement. Biochemistry. 2007;46:2707–2718. doi: 10.1021/bi0618565. [DOI] [PubMed] [Google Scholar]
- 30.Mizutani H, Tada-Oikawa S, Hiraku Y, Oikawa S, Kojima M, Kawanishi S. Mechanism of apoptosis induced by a new topoisomerase inhibitor through the generation of hydrogen peroxide. J Biol Chem. 2002;277:30684–30689. doi: 10.1074/jbc.M204353200. [DOI] [PubMed] [Google Scholar]