

Abstract
The aims of this study were: i) to analyze activation patterns of four upper limb muscles (duration of the active and non-active phase) in each lap of 200m breaststroke, ii) quantify neuromuscular fatigue, with kinematics and physiologic assessment. Surface electromyogram was collected for the biceps brachii, deltoid anterior, pectoralis major and triceps brachii of nine male swimmers performing a maximal 200m breaststroke trial. Swimming speed, SL, SR, SI decreased from the 1st to the 3rd lap. SR increased on the 4th lap (35.91 ± 2.99 stroke·min-1). Peak blood lactate was 13.02 ± 1.72 mmol·l-1 three minutes after the maximal trial. The EMG average rectified value (ARV) increased at the end of the race for all selected muscles, but the deltoid anterior and pectoralis major in the 1st lap and for biceps brachii, deltoid anterior and triceps brachii in the 4th lap. The mean frequency of the power spectral density (MNF) decreased at the 4th lap for all muscles. These findings suggest the occurrence of fatigue at the beginning of the 2nd lap in the 200m breaststroke trial, characterized by changes in kinematic parameters and selective changes in upper limb muscle action. There was a trend towards a non-linear fatigue state.
Key Points.
Fatigue in the upper limbs occurs in different way as it described by 100m swimming events.
Neuromuscular fatigue was estimated by analyzing the physiological changes (high blood lactate concentrations), biomechanical changes in the swimming stroke characteristics (decreased in swimming velocity), and by the changes in the EMG amplitude and frequency parameters at the end of the swimming bout.
The amplitude signal of EMG provided by the ARV demonstrated an increase at the end with the respect to the beginning for all muscles under study, excepted for the muscle deltoid anterior.
The mean frequency (MNF) in our study decrease at the end of the swimming in the 4th lap relative to the 1st lap for all muscles under observation, along the 200m breaststroke.
Key Words: Swimming, Kinematics, EMG, Mean frequency
Introduction
The breaststroke technique is considered one of the least economic of the four swimming styles (Barbosa et al., 2006), which can lead to early fatigue while swimming. Neuromuscular fatigue can be defined as: (i) the failure to maintain the required or expected force, accompanied by changes in muscular activity (Dimitrova et al., 2003); and (ii) the inability of skeletal muscle to generate high levels of muscular strength or maintain these levels over time (Enoka and Stuart, 1992). Additionally, the manifestations of fatigue have been associated with (Allen et al., 1995; Pagala et al., 1994): (i) a decline in muscle tension produced during and after submaximal and maximal exercise; (ii) an inability to maintain a given exercise intensity over time, reducing the speed of contraction and increased muscle relaxation time, as well as; (iii) the variation of intra and extracellular concentrations of certain metabolites and ions.
Neuromuscular fatigue can be divided into central and peripheral fatigue. Central fatigue comprises of decreases in the voluntary activation of the muscle, which is due to decreases in the number of recruited motor units and their discharge rate (González et al., 2012). Peripheral fatigue involves changes in neuromuscular transmission, muscle action potential propagation and decreases in the contractile tension of the muscle fibers (Boyas and Guevel, 2011). Peripheral fatigue during exercise is considered an impairment of the peripheral mechanisms from excitation to muscle contraction. Peripheral regulation is, therefore, related to a perturbation of calcium ion movements, an accumulation of phosphate, and/or a decrease of adenosine triphosphate stores (Boyas and Guevel, 2011).
Some studies attempted to relate the onset of fatigue with the execution of various sport techniques (Camata et al., 2011; Giangarra et al., 1993; Pink et al., 1993; Robineau et al., 2012). While swimming, neuromuscular mechanisms related to fatigue remain unclear, and the few studies which carried out research, focused mainly on the front-crawl technique (Caty et al., 2006; Figueiredo et al., 2011; Ikuta et al., 2012; Stirn et al., 2011).
Caty et al., 2006 observed a decrease in the instantaneous mean frequency in the extensor carpi ulnaris muscles (11. 4% and 8.5%, respectively) after a 4x50m high intensity front crawl. Stirn et al., 2011 reported that at the end of a 100m front crawl at a maximal effort, the mean power frequency decreased by 20-25%. Ikuta et al., 2012 suggested that the decrease in swimming velocity was related to a decrease in the activity of several muscles coordinated with each other, and that a compensating strategy was involved between the pectoralis major and other muscles during the last lap of a 4x50m front crawl test.
Important muscles activated in breaststroke swimming seem to be the biceps brachii, triceps brachii, subscapularis, latissimus dorsi, pectoralis major, supraspinatus, infraspinatus, serratus anterior, and deltoid anterior, teres minor and trapezio ) for the upper limbs (Conceição et al., 2010; Nuber et al., 1986; Ruwe et al., 1994; Yoshizawa et al., 1976). While gluteus maximus, vastus medialis, rectus femoris, biceps femoris, abductor magnus, quadriceps, gastrocnemius, tibialis anterior, abductor hallucius, abductor digiti minimi, flexor digitorum brevis are for the lower limbs (McLeod, 2010; Yoshizawa et al., 1976).
It is clear that EMG can be useful in tracking muscle fatigue for many reasons (the relationships found between sEMG features and muscle fatigue, and the possibility of recording them in almost any type of situation). Therefore, some studies have examined the relationship between muscle fatigue and EMG variables and, consequently, the possibility of using EMG models to accurately track muscle fatigue (González-Izal et al., 2012).
The linear techniques that are used to estimate muscle fatigue are based on linear regression, which relates changes in EMG parameters to changes in power loss (as a direct measurement of muscle fatigue). The non-linear techniques that are used to estimate muscle fatigue are based on neural networks to relate EMG parameters to muscle fatigue. Both linear and non-linear techniques provided good results for mapping changes in power or force loss during dynamic exercises based on sets of EMG parameters (González-Izal et al., 2012).
Apart from this, however, we have no idea of the appearance of fatigue of the upper limbs in breaststroke. Since there is a lack of studies to quantify the neuromuscular fatigue in the upper limbs, using spectral parameters, we believe that this research would have the potential to make a contribution to the limited body of knowledge in this field. The main aim for swimming coaches and researchers relies on the identification of the factors that might predict performance with higher validity and accuracy.
The aims of this study were: i) to analyze the activation patterns (duration of active and non-active phase) of the upper limbs’ four muscles during each lap for 200m breaststroke, ii) quantify neuromuscular fatigue, through kinematics and physiologic assessment. It was hypothesized that an increase in signal amplitude and a decrease in spectral parameters due to repetitive sub-maximal contractions, characterized a non-linear fatigue process. Also considered was that the fatigue process occurs differently for each of the muscles studied.
Methods
Subjects
Nine male swimmers (age: 22.3 ± 2.9 years; height: 1.81 ± 0.05 m; body mass: 73.60 ± 3.82 kg; mean ± SD) volunteered to participate in this study and provided written consent. They were all swimmers competing at the national level with an average personal best result for 200-m breaststroke (149.44 ± 6.59 s, corresponding respectively to 643.75 ± 53.77 FINA ranking points). All the procedures were approved by the institutional Ethics Committee and carried out according to the Helsinki Declaration.
Testing procedure
The experiments were performed in a 50m indoor swimming pool at a water temperature of 27.5 °C and 75% humidity. Subjects performed a standard warm-up of 800m front crawl, and a specific warm-up of 200m breaststroke at a medium level of effort. After a twenty minute rest subjects performed a maximal 200m breaststroke trial with a push off start.
Data acquisition: EMG data collection
Surface EMG signals from the biceps brachii (BB), deltoid anterior (DA), pectoralis major (PM) and triceps brachii (TB) on the right side of the body were collected. These muscles were selected according to their importance in breaststroke (Conceição et al., 2010; Nuber et al., 1986; Ruwe et al., 1994; Yoshizawa et al., 1976).
Bipolar surface electrodes were used (10-mm diameter discs, Plux, Lisbon, Portugal) with an inter-electrode distance of 20mm. Electrodes on the upper part of the PM were placed in the middle of the line that connects the acromion process and the manubrium (sternum) two fingers below the clavicle (Stirn et al., 2011). The electrodes on the long head of the BB, DA and TB were placed in accordance with SENIAM recommendations (Herrmens and Freriks, 1999).
The skin was shaved, rubbed with sandpaper and cleaned with alcohol so that the inter-electrode resistance did not exceed 5 KOhm (Basmajian and De Luca, 1985). The ground electrode was positioned over the cervical vertebrae. Transparent dressings with labels (Hydrofilm®, 10 cm x 12,5 cm, USA) were used to cover the electrodes to isolate them from the water (Hohmann et al., 2006). All cables were fixed to the skin by adhesive tape in several places to minimize their movement and any signal interference. Swimmers wore a full body swimming suit to further immobilize the cables (Fastskin Speedo®, Speedo Aqualab, USA).
The wireless EMG device (BioPLUX.research, Lisbon, Portugal) with 8 analog channels (12-bit), sampling rate at 1 kHz, was put in a waterproof bag (84x53x18mm) and placed inside the swimmer´s cap. Data was transmitted to the PC in real time via Bluetooth.
The EMG and video data were recorded simultaneously and were not synchronized with video recording, because the aim of this study was to understand the appearance of fatigue in relation to race parameters rather than detailed within cycle swimming kinematics.
All EMG analysis was conducted with a MATLAB routine (Mathworks, Inc.,Natick MA,USA). The process of determining the muscle activity boundaries consists of finding the neighborhood points, where the energy was 30% of muscle activation maximum peak within a stroke (Stirn et al., 2011). These were calculated by segmenting the muscle input signal energy according to the same criteria described in Stirn et al., 2011. Starting from the raw signal, DC components were removed and thereafter filtered with a fifth-order Butterworth band pass filter where the lower and upper cut-off frequencies were set to 10 and 500Hz respectively. The signal energy was then determined with a 250 ms sliding window (Stirn et al., 2011) and according to:
| (1) |
Even though the high frequencies of the input signal were filtered with a Butterworth filter, muscle energy is very noisy and presents several local maximum peaks that didn´t correspond to the muscle active window center, as shown in Figure 2. To overcome this difficulty, a strategy to determine the muscle’s “true” maximum energy peaks was devised. Each stroke taken by a swimmer produces patterns in the signal, these patterns are mainly translated by a periodicity in EMG energy, see Figure 1.
Figure 2.
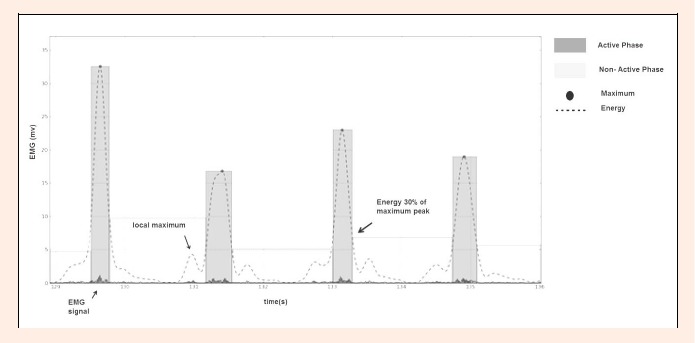
Segment of non-active phase and active phase of the EMG signal.
Figure 1.
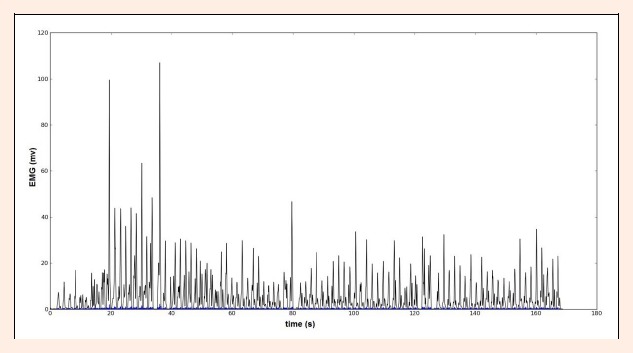
Energy envelope of a EMG signal segment
By defining the signal mean period, one can use this information to determine the maximum peak candidates with the highest and with minimal differences between two maximum candidates and the expected period.
Once the maximum candidates have been determined, the muscle activity boundaries were then selected by determining the neighborhood points where the energy is 30% of the determined maximum peaks. For each muscle activation, the active phase was defined as the part of the EMG signal for which the energy was at least 30% of the local maximum energy value, for each particular muscle activation. The raw EMG segments belonging to the active phases were extracted and used in the calculation of the active phase duration and the amplitude frequency analysis. The non-active phase was defined as the time interval between the two consecutives active phases as shown in Figure 2.
The temporal evolution of the active and non-active phases average durations during stroke were calculated for each muscle for the entire 200m. Linear regression curves were fitted to the data and the durations of the fitted curves at the beginning and completion of the swim were compared. The comparisons of the durations of the active and non-active phases of each muscle during the stroke were made to detect changes that might occur due to fatigue.
The average amplitude of EMG of each active phase was estimated using the average rectified value (ARV) of the EMG. ARV was calculated in accordance with SENIAM recommendations (Herrmens and Freriks, 1999) and plotted as a function of time. A linear regression curve was fitted to the data and the ARV values of the curve at the time of commencement and the end of the swimming bout were compared (Figure 3).
Figure 3.
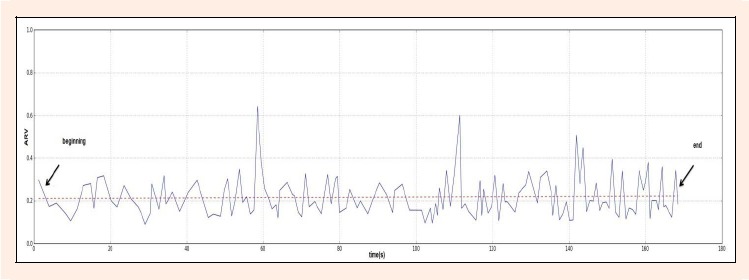
Example of a linear regression of the tríceps brachii of one swimmer, of the ARV values in the beginning and the end of the swimming bout.
For the frequency analysis, each extracted segment was zero-padded to the total length of 1s (i.e., 1,000 samples). In this way, a uniform frequency resolution (the frequency sampling interval) was used for all signal segments. The power spectral density (PSD) for each segment was estimated using the periodogram method (Proakis and Manolakis, 1996). The periodogram of a continuous signal segment x(t) of length T is defined as:
| (2) |
Even though the periodogram is a non-consistent estimate of PSD (its variance is large and does not become zero with increasing length of the signal), it was demonstrated that using more sophisticated methods for PSD estimation, does not significantly improve the estimation of power spectrum central frequency measures (the mean or the median frequency) (Farina and Merletti, 2000).
Since the differences were insignificant, we decided to use the periodogram estimate in our study. As a measure of the central tendency of PSD we used the mean frequency of the PSD (MNF). MNF is defined as the first moment of the PSD and was developed by Kwatny et al., 1970. For the continuous spectrum spanning the frequencies between 0 and fMAX is defined as:
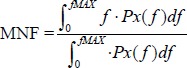 |
(3) |
The value of MNF was calculated for each segment and used as a frequency parameter of muscle fatigue.
The values of MNF belonging to each muscle were plotted as a function of time. Since all data exhibited a trend toward linear decrease with time, a linear model was fitted to all MNF data sets to the initial MNF value (the value of MNF at the time of the first stroke) and the final MNF value (the value of MNF at the time of the last stroke) and labeled as MNFbeg and MNFend, respectively. To normalize results between subjects, the final MNF values were expressed as a percentage of the initial values and labeled MNFn as:
| (4) |
The slope of the regression line was also calculated as an estimate of the rate of change over time.
Physiological data collection
Blood samples were collected from the earlobe at 3, 5, 7 minutes after the bout. Post exercise blood lactate concentrations were measured using a hand-held portable apparatus (Lactate Pro Analyser™, Arkay, Kyoto, Japan). According Pyne et al., 2001 this equipment exhibits a high accuracy compared with other lactate analyzers and high degree of versatility under a variety of testing conditions.
Kinematic data collection
The swims were videotaped simultaneously on sagittal plane with a pair of cameras providing a dual projection from both underwater (Sony Mini Dv DCR-HC42E, EUA) and above (Sony Mini Dv DCR-HC42E, EUA) the water surface. The cameras were placed stationary at 25-m of the headwall, on a lateral wall, perpendicular to the swimmer’s trajectory and 10-m away. The study comprised the kinematic analysis of stroke cycles (Ariel Performance Analysis System, Ariel Dynamics Inc., USA) at a sample rate of 50Hz. Zatsiorsky’s model with an adaptation by De Leva, 1996 was used with 8 anatomical landmarks and a trunk division in two articulated parts (Barbosa et al., 2010; Hirata and Duarte, 2007; Lafond et al., 2004; Pavol et al., 2002). To create a single image of dual projection as described previously (Barbosa et al., 2006; 2010), the independent digitalization from both cameras was reconstructed with the support of a calibration volume (16 point) and a 2D-DLT algorithm (Abdel-Aziz and Karara, 1971).
The stroke cycle was measured between the 18th m and 22th m, and identified at the end of the leg´s recovery (i.e., when the knees were in full flexion and feet on eversion).
Stroke mechanics were measured by the stroke cycle period (P, s), the stroke rate (SR = 1/P, Hz), the stroke length (SL, m) and the mean swimming speed of the full stroke (v, m·s-1). Finally, the swimming efficiency was estimated using the stroke index (SI = v SL, m2·c-1·s-1) as suggested by Costill et al., 1985. This index assumes that, at a given v, the swimmer with greater SL has the most efficient swimming technique.
Statistical procedures
The assumptions of normality of data (Kolmogorov-Smirnov test) and homogeneity of variance (Levene’s test) were both confirmed for all parameters. All results are reported as the mean values along with standard deviation values. The repeated measures ANOVA, with subsequent Tukey’s test as post hoc analysis where applicable, were used for multiple comparisons between the laps (p < 0.05).
Results
Kinematics data
Figure 4 presents the mean and SD of the kinematics parameters (Swimming Speed, SL, SR, SI) in the 4th lap of the 200m breaststroke. Swimming speed decreased from 1.38 ± 0.09 m·s-1 in the 1st lap to 1. 14 ± 0.08 m·s-1 in the 4th lap. Significant variations were observed on swimming speed between the 1st lap and 2nd lap, and between the 1st lap and 3rd lap [F = 21.27, p < 0.05].
Figure 4.
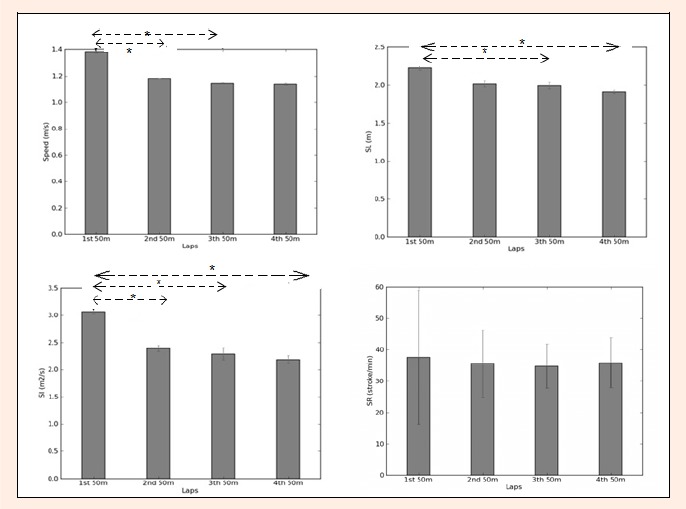
Mean and SD of Swimming Speed (m·s-1); Stroke length (SL) m; Stroke index (SI) m2·c-1·s-1 and Stroke rate (SR) stroke min-1 in the four 50m lap of a 200-m breaststroke. * p< 0.05.
SL decreased from 2.23 ± 0.18 m in the 1st lap to 1.92 ± 0.15 m in 4th lap. Significant variations were observed on the SL between the 1st lap and 3rd lap, and between the 1st lap and 4th lap [F = 4.41, p < 0.05].
SR decreased from 37.58± 4.90 stroke min-1 in the 1st lap to 34. 80±2.83 stroke min-1 in 3rd lap and then increased in the 4th lap (35.91 ± 2.99 stroke·min-1). No significant variations were observed between laps [F = 0.92, p > 0. 05]. Lastly, the SI was higher in the 1st lap, and decreasing from the 1st (3.07 ± 0.25 m2·s-1) to the last lap (2.19 ± 0.29 m2·s-1). Significant variations were observed on the SI between the 1st lap and 2nd lap, between the 1st lap and 3rd lap and between the 1st lap and 4th lap, [F = 16.94, p < 0.05].
Physiology
The means and standard deviations in blood lactate concentration (La) measured after the 200 m breaststroke are shown in the Figure 5. The highest value (13.02 ± 1.72 mmol·l-1) was observed at the 3rd minute after the trial. Significant variations were observed in La between the 1st minute and 3rd minute, and between the 1st minute and the 5th minute [F = 2. 47, p < 0.05].
Figure 5.
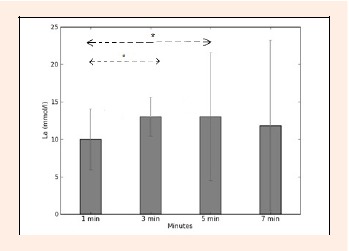
Blood lactate concentration after swim 1, 3, 5 and 7 min. * p < 0.05.
Neuromuscular
The duration of the active phase (Table 1) was, for the BB muscle, higher in the beginning phase compared to the end phase of the swim for all four laps. The DA muscle presented a higher value in the beginning phase in the 1st lap and 3rd lap, but exhibited a higher value in the end phase for the 2nd and 4th lap. The PM muscle demonstrated a higher duration of active phase at the end for all laps with the exception of the 3rd lap (PMbeg: 0.69 ± 0.62 s and PMend: 0.55 ± 0.33 s) where the active phase was higher at the beginning than the end of the session. The TB muscle showed a higher active phase at the beginning in the 1st and 3rd lap, and higher values in the end in the 2nd and 4th lap.
Table 1.
Mean (±SD) duration of active phase for active phasebeg and active phaseend four all the muscles (BB- bíceps brachii; DA- deltoid anterior; PM- pectoralis major and TB- triceps brachii), over the four laps of the 200 m breaststroke maximal effort.
| 1st lap | 2nd lap | 3rd Lap | 4th Lap | |||||
|---|---|---|---|---|---|---|---|---|
| Laps | Active phasebeg (s) |
Active phaseend (s) |
Active phasebeg (s) |
Active phaseend (s) |
Active phasebeg (s) |
Active phaseend (s) |
Active phasebeg (s) |
Active phaseend (s) |
| BB | .99 (.78) | .98 (.90) | .73 (.26) | .58 (.14) | .68 (.27) | .56 (.20) | .70 (.24) | .63 (.10) |
| DA | .78 (.30) | .64 (.17) | .63 (.14) | .72 (.14) | .87 (.67) | .58 (.25) | .82 (.38) | .93 (.58) |
| PM | .61 (.18) | .71 (.32) | .64 (.17) | .88 (.48) | .69 (.62) | .55 (.33) | .62 (.35) | .79 (.63) |
| TB | .71 (.21) | .69 (.25) | .54 (.14) | .58 (.14) | .54 (.22) | .52 (.15) | .84 (.74) | .85 (.76) |
The long duration of active phase was shown in BB at the beginning of the swim in the 1st lap (0.99 ± 0.78 s) and the shorter for the TB at the end of swimming the 3rd lap (0.52 ± 0. 15 s). Moreover, for all the muscles, the active phase showed the same trend in the 3rd lap. The duration was higher in the beginning compared to the end of the swim. The differences between beginning and the end for each muscle were not statistically significant for p < 0.05.
The non-active phase in Table 2, showed an increase in the end of the test for all the muscles in the 3rd lap and a decrease in the 4th lap. The muscles BB, DA and PM demonstrated the same behavior in the relative duration of the non-active phase for each lap of the 200m breaststroke, that is, in the 1st and 3rd lap they had a longer duration at the end compared to the beginning of the swim, unlike in the 2nd and 4th lap where the higher duration was observed in the beginning of the swim.
Table 2.
Mean (±SD) duration of non-active phase for non- active phasebeg and non- active phaseend for all the muscles (BB- biceps brachii; DA- deltoid anterior; PM- pectoralis major and TB- triceps brachii), over the four laps of the 200 m breaststroke maximal effort.
| 1st lap | 2nd lap | 3rd Lap | 4th Lap | |||||
|---|---|---|---|---|---|---|---|---|
| Laps | Non-Active phasebeg (s) |
Non-Active phaseend (s) |
Non-Active phasebeg (s) |
Non-Active phaseend (s) |
Non-Active phasebeg (s) |
Non-Active phaseend (s) |
Non-Active phasebeg (s) |
Non-Active phaseend (s) |
| BB | .76 (.24) | .80 (.24) | .81 (.21) | .72 (.22) | .77 (.20) | .81 (.28) | .79 (.26) | .78 (.17) |
| DA | .66 (.16) | .71 (.14) | .79 (.15) | .69 (.17) | .70 (.16) | .75 (.19) | .70 (.24) | .68 (.26) |
| PM | .60 (.18) | .60 (.13) | .70 (.24) | .68 (.32) | .63 (.21) | .65 (.17) | .72 (.20) | .68 (.16) |
| TB | .63 (.27) | .62 (.18) | .55 (.17) | .67 (.26) | .59 (.17) | .64 (.29) | .61 (.18) | .53 (.22) |
The TB showed a different behavior compared to the other muscles studied, which presented a higher duration in the non-active phase in the beginning compared to the end of the swimming the 1st and 4th lap, and in the end for the 2nd and 3rd laps, moreover for all the muscles under study the non-active phase showed the same behavior in the 3rd lap, where the duration was higher in the beginning which also happened in the active phase. The differences between the beginning and the end for each muscle were not statistically significant for p < 0.05.
The relative duration of activation (RAF) in Figure 6, decreases at the beginning phase in the 1st and 3rd lap for the DA and PM, whereas for the 2nd and 4th lap it was higher in the end for both the DA and PM. The BB demonstrated a RAF higher in the end phase in the 1st and 2nd lap, and the reverse in the 3rd and 4th lap. The TB decreased at the beginning phase over the first three laps, in contrast to the 4th lap where the value was higher in the end. The differences between the beginning and the end for each muscle were not statistically significant for p < 0.05.
Figure 6.
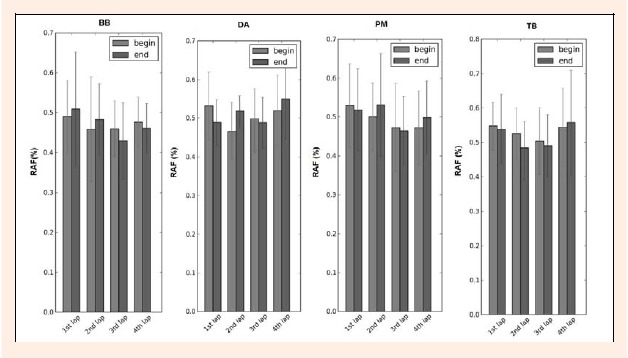
Comparison of the mean relative duration of activation value with SD for RAFbeg (gray bars) and RAFend (black bars) four all the muscles (BB- bíceps brachii; DA- deltoid anterior; PM- pectoralis major and TB- triceps brachii), over the four laps of the 200 m breaststroke maximal effort
Figure 7 demonstrated that the average rectified value increases at the end phase compared to the beginning phase for all muscles comparing the 1st lap with the 4th lap, except for the DA (DAbeg:0.42 ± 0.13; DAend: 0.42 ± 0.10) that demonstrated higher values in the beginning phase. The BB and TB muscles have similarities throughout the laps, that is, in the first three laps (1st, 2nd and 3rd laps) the ARV was higher in the end compared to the beginning and in the 4th lap the reverse was observed. The PM showed similarities in the 1st and 3rd lap which had higher values in the beginning, and in the 2nd and 4th lap had higher values at the end.
Figure 7.
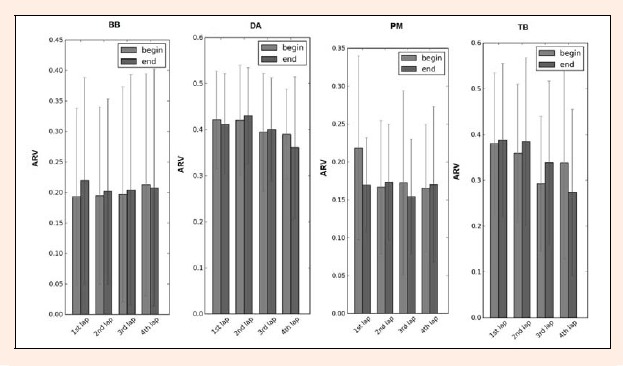
Mean ARV value with SD for ARVbeg (gray bars) and ARVend (black bars) four all the muscles (BB- bíceps brachii; DA- deltoid anterior; PM- pectoralis major and TB- triceps brachii), over the four laps of the 200 m breaststroke maximal effort.
In Figure 8, the mean MNF value for the BB increased in all the laps at the end except the 3rd lap (BBend: 82.47 ± 16.20 Hz BBbeg: 86.15 ± 11.72 Hz). DA decreases in all the laps at the beginning except in the 3rd lap (DAend: 77.38 ± 13.68 Hz, DAbeg: 74.54 ± 15. 52 Hz). The PM decreases at the beginning in the 1st lap (PMbeg: 89.65 ± 23.61 Hz; PMend: 89.53 ± 17.74 Hz) and 3rd lap (PMbeg: 79.37 ± 10.58 Hz; PMend: 76.16 ± 16.92 Hz), and increases at the beginning in the 2nd lap (PMbeg: 82.13 ± 15.33 Hz; PMend: 86.11 ± 18.12 Hz) and 4th lap (PMbeg: 74. 33 ± 14.61 Hz; PMend: 75.18 ± 13.92 Hz). TB decreases at the beginning in the 1st lap (TBbeg: 78.17 ± 10.83 Hz; TBend: 74.79 ± 16.37 Hz) and in the 4th lap (TBbeg: 72.45 ± 11.83 Hz; TBend: 65.72 ± 12. 94 Hz), whereas in the 2nd (TBend: 73.72 ± 13.68 Hz; TBbeg: 72.41 ± 12.65 Hz) and 3rd lap (TBend: 71.32 ± 11.26 Hz; TBbeg: 68.78 ± 11.03 Hz) we can observe an increase at the end phase compared to the beginning. Moreover, the MNF decreases at the end of swimming the 4th lap relative to the 1st lap for all muscles under observation for the 200m breaststroke. Note that the differences between the beginning and the end for each muscle were not statistically significant for p < 0.05.
Figure 8.
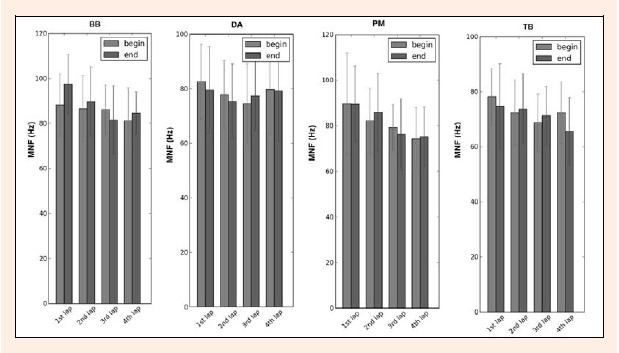
Mean MNF value with SD for MNFbeg (black bars) and MNFend (gray bars) four all the muscles (BB- bíceps brachii; DA- deltoid anterior; PM- pectoralis major and TB- triceps brachii) and swimming bouts, over the four laps of the 200 m breaststroke maximal effort.
Discussion
The aims of this study were: i) to analyze four muscle activation patterns (duration of active and non-active phases) of the upper limbs during each lap for the 200m breaststroke, ii) quantify neuromuscular fatigue, through kinematics and physiologic assessment. The main findings were that some indicators of neuromuscular fatigue begin to appear shortly into the 2nd lap, but are more evident in the 3rd lap (i.e amplitudes increase and spectral variables decrease) in a non-linear fashion.
Kinematics
The average decrease in the swimming speed from the 1st lap to the 4th lap shows a much greater decline in speed compared to the results of similar studies (Takagi et al., 2004; Thompson et al., 2000; 2004). In breaststroke, an increase in swimming speed is associated with an increase in SR, but has a decline more in SL relative to the other swimming strokes (Thompson et al., 2004), supporting the changes in the swimming technique under the 200m breaststroke (Takagi et al., 2004). In this study SL suffered a decrease in the 1st lap to the 4th lap, whereas the SR decreased from the 1st lap to the 3rd lap and then increased in the 4th lap.
As swimming speed decreased, different SR and SL combinations were observed, giving the best performance in the face of the fatigue task constraints, and decreasing the swimmer’s ability to maintain a constant SL. Increasing SR in the last 50m lap is due to the inability to generate sufficient extra effort to overcome high drag. The decrease in the swimming speed SL, SI and the increase of SR in the last lap suggested a trend for the inability to generate sufficient extra effort needed to overcome high drag throughout the test (Craig and Pendergast, 1979; Chatard et al., 2003; Thompson et al., 2004). This change in the stroke kinematics due to a decrease in the power output can be a consequence of peripherical fatigue phenomena (Ikuta et al., 2012; Stirn et al., 2011).
Physiology
The blood lactate concentration levels after the test reached values that would be expected after a 200m breaststroke race and are in agreement with the results in other studies in which similar tests were performed with swimmers of the same or similar competitive level (e.g., Capelli et al., 1998; Lomax and Castle, 2011). These results suggest that the swimmers performed the swimming test very close to their maximum effort level. The average blood lactate concentration measured after 3 minutes was 13.2 ± 1.72 mmol·l-1, representing the highest values. Increases of lactate concentration are responsible for fatigue by changes in the intracellular pH. As a result, muscle fiber conduction velocity decreases and changes the shape of the motor unit action potential waveform (Cifrek et al., 2009).
Neuromuscular
The amplitude of EMG signals is influenced by the number of active motor units (Moritani et al., 1986), their discharge rates, and the shape and propagation velocity of the intracellular action potentials (Dimitrova and Dimitrov, 2002).
The amplitude signal of EMG provided by the ARV demonstrated an increase and maintenance at the end phase compared to the beginning for BB and PM, comparing the 1st lap with the 4th lap, and a small decrease for DA and TB that demonstrated higher values in the beginning phase compared to the end phase. The swimmers, during the end of the swim, increase the period in the recovery phase, decreasing the amplitude of the EMG signal for DA and TB. These muscles are responsible for the movement of antepulsion of the arms up to the point where it is stretched in front.
EMG amplitude has been observed to increase during submaximal dynamic exercise (Tesch et al., 1990) and to decrease during exercises at maximal levels of voluntary contraction (Komi and Tesch, 1979). An increase of the EMG amplitude during swimming has already been observed in previous studies (Rouard and Clarys, 1995; Rouard et al., 1997; Stirn et al., 2011; Wakayoshi et al., 1994). Ikuta et al., 2012 noted that the mean amplitude value of the PM was significantly higher for the 4th lap than for the 2nd and 3rd laps in 4x50 m front crawl. The authors noted that the increase activity of the PM may have been a compensatory strategy to maintain swimming speed during the test swim. Moreover, Nuber et al., 1986 verified that the PM continued to activate through recovery in the breaststroke.
So, it seems that a decrease in the active phase duration and the increase in the non-active phase on the 3rd lap, with all the muscles monitored, reveal that in this lap muscles start to display the onset of neuromuscular fatigue indicating that this is a milestone of the 200m breaststroke race.
The MNF decreases at the 4th lap in comparison to the 1st lap for all muscles. Dynamic fatiguing tasks have been found to show a decrease in the mean power frequency (Tesch et al., 1990). A number of authors reported similar data in several other human muscles (Basmajian and De Luca, 1985; De Luca, 1984). The same is also apparent for swimming, or at least for front-crawl (Figueiredo et al., 2011; Stirn et al., 2011). The reduction of muscle fiber conduction velocity is one of the causes of signal power spectrum shift toward lower frequencies, and also of the increase in the EMG signal amplitude because of a spatial low-pass filtering effect of tissue as a volume conductor (De Luca, 1984).
However, during a fatiguing exercise, two oppositing effects might occur (Petrofsky and Lind, 1980): (i) a decrease in the mean frequency as discussed above and; (ii) increase in intramuscular temperature due to the exercise, which causes an increase in the mean frequency. So it is possible that during some types of exercise, the two effects compensate each other, and the decrements found in the mean frequency can be small or non-significant.
Associations
A decrease in swimming speed, SL, SI and an increase in the SR in the last lap, can be associated to the higher La after the test, to a non-significant increase of the amplitude and a non-significant decrease in the spectral variables. These suggest that the presence of fatigue in the upper limbs occurs in a different way for the different muscles studied, by a non-linear process. However, more investigation in order to better understand this phenomenon is required in the near future.
As blood flow is restricted, at a certain level of contraction, by intramuscular pressure, muscle becomes ischemic. Myoelectric manifestations of muscle fatigue might be affected by this event (Merletti et al., 1984). However, these are evidences for land-based exercises, and to the best of our knowledge none of this data has been collected during swimming.
The shape of the ARV during the four laps represent the increased activation patterns in the 2nd and 3th lap in accordance with the normal adaptation and the tactic to use the remaining capacity in the final lap to achieve the best result. The higher activation by DA and TB showed that these muscles play a significant role in this event (Yoshizawa et al., 1976; Conceição et al., 2010).
With this research we can provide quick practical information for swimmers and coaches to adjust training methods.
Conclusion
This study showed a trend for a neuromuscular fatigue state in a non-linear fashion, with the 2nd and 3rd lap being the critical moments of a 200m breaststroke race. Decreases in swimming speed are related to SL, SI, and increases of the SR in the last lap, increases in the blood lactate concentration after the test, and a non-significant increase of the amplitude and non-significant decrease in the spectral parameters.
Both the technical and physiologic training in breaststroke swimmers should take into account the onset of fatigue in a non-linear fashion, including specific training to prevent sharp falls in the 2nd and 3rd laps of competitive racing.
Acknowledgements
The authors acknowledge the support of all swimmers and their coaches that participated in this study.
Biographies

Ana CONCEIÇÃO
Employment
Assistant Professor at the Sports Science Institute of Rio Maior, Rio Maior, Portugal.
Degree
PhD
Research interests
Swimming biomechanics and muscle mechanics.
E-mail: anaconceicao@esdrm.ipsantarem.pt

António José SILVA
Employment
Full Professor at the Sport Sciences Department of the University of Trás-os-Montes and Alto Douro, UTAD - Portugal.
Degree
PhD
Research interests
The physiological and biomechanical indicators of energy cost during physical activities, namely in swimming.
E-mail: ajsilva@utad.pt

Tiago BARBOSA
Employment
Professor at the Nanyang Technological University, and Member of the Research Centre in Sports, Health and Human Development, Portugal.
Degree
PhD
Research interests
The biomechanical and physiological determinant factors of swimming performance.
E-mail: tiago.barbosa@nie.edu.sg

István KARSAI
Employment
Assistant Professor at the University of Pécs, Pécs, Hungary.
Degree
PhD
Research interests
Swimming biomechanics and muscle mechanics.
E-mail: karsai@gamma.ttk.pte.hu

Hugo LOURO
Employment
Professor at the Sports Science Institute of Rio Maior, Rio Maior, Portugal.
Degree
PhD
Research interests
Methodological observation in swimming.
E-mail: hlouro@esdrm.ipsantarem.pt
References
- Abdel-Aziz Y., Karara H. (1971) Direct linear transformation: from comparator coordinates into object coordinates in close range photogrammetry. Proceedings of the Symposium on Close-Range Photogrammetry. Illinois: Church Falls: 1-18 [Google Scholar]
- Allen D.G., L.annergren J., Westerblad H. (1995) Muscle cell function during prolonged activity: cellular mechanisms of fatigue. Experimental Physiology 80, 497-527 [DOI] [PubMed] [Google Scholar]
- Barbosa T.M., Fernandes R., Keskinen K.L., Colaço P., Cardoso C., Silva J., Vilas-Boas J.P. (2006) Evaluation of the energy expenditure in competitive swimming strokes. International Journal Sports Medicine 27, 894-899 [DOI] [PubMed] [Google Scholar]
- Barbosa T.M., Silva A.J., Reis A., Costa M., Garrido N., Policarpo F., Reis V. (2010) Kinematical changes in swimming front crawl and breaststroke with the Aqua trainer snorkel. European Journal of Applied Physiology 109, 1155-1162 [DOI] [PubMed] [Google Scholar]
- Basmajian J.V., De Luca C.J. (1985) Muscles Alive: Their Function Revealed by Electromyography. Fifth edition Williams and Wilkins, Baltimore [Google Scholar]
- Boyas S., Guevel A. (2011) Neuromuscular fatigue in healthy muscle: underlying factors and adaptation mechanisms. Annals of Physical Rehabilitation Medicine 54, 88-108 [DOI] [PubMed] [Google Scholar]
- Cabri J., Gamboa H. (2009) The use of electromyography in physiotherapy- application in Hydrotherapy. In:Book of Abstracts of European Workshop on Movement Science, Lisbon, Portugal. Eds:Araújo D., Cabri J., Barreiros J.37 [Google Scholar]
- Camata T.V., Altimari L.R., Bortolotti H., Dantas J.L., Fontes E.B., Smirmaul B.P., Okano A.H., Chacon-Mikahil M.P., Moraes A.C. (2011) Electromyography activity and rate of muscle fatigue of the quadriceps femoris during cycling exercise in the severe domain. Journal of Strength & Condition Research 25, 2537-2543 [DOI] [PubMed] [Google Scholar]
- Capelli C., Pendergast D.R., Termin B. (1998) Energetics of swimming at maximal speeds in humans. European Journal of Applied Physiology & Occupational Physiology 78, 385-393 [DOI] [PubMed] [Google Scholar]
- Caty V., Rouard A., Hintzy Y., Aujoannet Y., Molinari M., Knaflitz M. (2006) Time- frequency parameters of wrist muscles EMG after an exhaustive freestyle test. Portuguese Journal of Sport Science 6, 28-30 [Google Scholar]
- Chatard J.C., Wilson B. (2003) Drafting distance in swimming. Medicine and Science Sports Exercise 35, 1176-1181 [DOI] [PubMed] [Google Scholar]
- Cifrek M., Medved V., Tonkovic S., Ostojic S. (2009) Surface EMG based muscle fatigue evaluation in biomechanics. Clinical Biomechamincs 24, 327-340 [DOI] [PubMed] [Google Scholar]
- Conceição A., Gamboa H., Palma S., Araújo T., Nunes N., Marinho D., Costa A., Silva A., Louro H. (2010) Comparison between the standard average muscle activation with the use of snorkel and without snorkel in breaststroke technique. In:Abstract Book of XITH International Symposium Biomechanics and Medicine in Swimming, Oslo. 46-47 [Google Scholar]
- Costill D., Kovaleski J., Porter D., Fielding R., King D. (1985) Energy expenditure during front crawl swimming: predicting success in middle-distance events. International Journal Sports Medicine 6, 266-270 [DOI] [PubMed] [Google Scholar]
- Craig A., Pendergast D.R. (1979) Relationships of stroke rate, distance per stroke, and velocity in competitive swimming. Medicine & Science in Sports & Exercise 11, 278-283 [PubMed] [Google Scholar]
- De Leva P. (1996) Adjustments to Zatsiorsky-Seluyanov’s segment inertia parameters. Journal of Biomechanics 29, 1223-1230 [DOI] [PubMed] [Google Scholar]
- De Luca C.J. (1984) Myoelectrical manifestation of localized muscular fatigue. Critical Reviews in Biomedical Engineering 11, 251-279 [PubMed] [Google Scholar]
- Dimitrova N.A., Dimitrov G.V. (2002) Amplitude-related characteristics of motor unit and Mwave potentials during fatigue. A simulation study using literature data on intracellular potential changes found in vitro. Journal of Electromyography and Kinesiology 12, 339-349 [DOI] [PubMed] [Google Scholar]
- Dimitrova N.A., Dimitrov G.V. (2003) Interpretation of EMG changes with fatigue: facts, pitfalls, and fallacies. Journal of Electromyography and Kinesiology 13, 13-36 [DOI] [PubMed] [Google Scholar]
- Enoka R., Stuart D. (1992) Neurobiology of muscle fatigue. Journal of Applied Physiology 72, 1631-1648 [DOI] [PubMed] [Google Scholar]
- Farina D., Merletti R. (2000) Comparison of algorithms for estimation of EMG variables during voluntary isometric contractions. Journal of Electromyography and Kinesiology 10, 337-349 [DOI] [PubMed] [Google Scholar]
- Figueiredo P., Pereira S., Gonçalves P., Vilas-Boas J.P.and Fernandes R. (2011) Muscular fatigue during 200m front crawl. Portuguese Journal of Sport Science 11(Suppl.2), 735-738 [Google Scholar]
- Ikuta J., Matsuda Y., Yamada Y., Kida N., Oda S., Moritani T. (2012) Relationship between decreased swimming velocity and muscle activity during 200-m front crawl. European Journal of Applied Physiology 112(9), 3417-3429 [DOI] [PubMed] [Google Scholar]
- Giangarra C.E, Conroy B., Jobe F.W., Pink M., Perry J. (1993) Electromyography and cinematographic analysis of elbow function in tennis players using single and double handed backhand strokes. American Journal of Sports Medicine 21, 394-399 [DOI] [PubMed] [Google Scholar]
- González-Izal M., Malanda A., Gorostiaga E., Izquierdo M. (2012) Electromyographic models to assess muscle fatigue. Journal of Electromyography and Kinesiology 22, 501-512 [DOI] [PubMed] [Google Scholar]
- Herrmens H.J., Freriks B. (1999) European recommendations for surface electromyography, results of the SENIAM project (CDrom). Roessingh Research and Development, Enschede [Google Scholar]
- Hirata R.P., Duarte M. (2007) Effect of relative knee position on internal mechanical loading during squatting. Brazilian Journal of Physical Therapy 11(2), 121-125 [Google Scholar]
- Hohmann A., Kirsten R., Kruger T. (2006) EMG-Model of the backstroke start technique. In: X International Symposium of Biomechanics and Medicine in Swimming. Eds:Vilas Boas J.P., Alves F., Marques A.Portuguese Journal of Sport Science 6, 38-39 [Google Scholar]
- Ikai M., Ishii K., Miyashita M. (1964) An electromyographic study of swimming. Journal of Physical Education 7, 47-54 [Google Scholar]
- Komi P.V., Tesch P. (1979) EMG frequency spectrum, muscle structure, and fatigue during dynamic contractions in man. European Journal of Applied Physiology and Occupational Physiology 42, 41-50 [DOI] [PubMed] [Google Scholar]
- Kwatny E., Thomas D.H., Kwatny H.G. (1970) An application of signal processing techniques to the study of myoelectric signals. IEEE Transactions Biomedical. Engineering 17, 303-313 [DOI] [PubMed] [Google Scholar]
- Lafdond D., Duarte M., Prince F. (2004) Comparison of three methods to estimate the center of mass during balance assessment. Journal of Biomechanics, 37, 1421-1426 [DOI] [PubMed] [Google Scholar]
- Lewillie L. (1967) Analyse télemétrique de l’electromyogramme du nageur. Societé Médicine Belge Education Physique Sport 20, 174-177 (In French) [Google Scholar]
- Lewillie L. (1973) Muscular activity in swimming. Biomechanics III. Karger, Basel: 440-445 [Google Scholar]
- Lomax M., Castle S. (2011) Inspiratory muscle fatigue significantly affects breathing frequency, stroke rate, and stroke length during 200-m front-crawl swimming. Journal Strength Condition Research 25, 2691-2695 [DOI] [PubMed] [Google Scholar]
- McLeod I. (2010) Swimming Anatomy. Human Kinetics, USA [Google Scholar]
- Merletti R., Sabbahi M.A., DeLuca C.J. (1984) Median frequency of the myoelectric signal effects of muscle ischemia and cooling. European Journal of Applied Physiology and Occupational Physiology 52, 258-265 [DOI] [PubMed] [Google Scholar]
- Moritani T., Muro M., Nagata A. (1986) Intramuscular and surface electromyogram changes during muscle fatigue. Journal of Applied Physiology 60, 1179-1185 [DOI] [PubMed] [Google Scholar]
- Nuber G.W., Jobe F.W., Perry J., Moynes D.R., Antonelli D. (1986) Fine wire electromyography analysis of muscles of the shoulder during swimming. American Journal of Sports Medicine 14, 7-11 [DOI] [PubMed] [Google Scholar]
- Pagala M., Ravindran K., Amaladevi B., Namba T., Grob D. (1994) Potassium and caffeine contractures of mouse muscles before and after fatiguing stimulation. Muscle and Nerve Journal 17, 852-859 [DOI] [PubMed] [Google Scholar]
- Pavol M., Owings T., Grabiner M. (2002) Body Segment inertial parameter estimation for the general population of older adults, Journal of Biomechanics, 35, 707-712 [DOI] [PubMed] [Google Scholar]
- Petrofsky J.S., Lind A.R. (1980) The influence of temperature on the amplitude and frequency components of the EMG during brief and sustained isometric contractions. European Journal of Applied Physiology and Occupational Physiology 44, 189-200 [DOI] [PubMed] [Google Scholar]
- Pink M., Jobe F.W., Perry J., Browne A., Scovazzo M.L., Kerrigan J. (1993) The painful shoulder during the butterfly stroke. An electromyography and cinematographic analysis of twelve muscles. Clinical Orthopaedics and Related Research 288, 48-59 [PubMed] [Google Scholar]
- Proakis J.G., Manolakis D.G. (1996) Digital Signal Processing. Upper Saddle River, NJ:Prentice Hall [Google Scholar]
- Pyne D.B., Lee H., Swanwick K.M. (2001) Monitoring the lactate threshold in world ranked swimmers. Medicine & Science in Sports & Exercise 33, 291-297 [DOI] [PubMed] [Google Scholar]
- Robineau J., Jouaux T., Lacroix M., Babault N. (2012) Neuromuscular fatigue induced by a 90-minute soccer game modeling. Journal of Strength & Condition Research 26, 555-262 [DOI] [PubMed] [Google Scholar]
- Rouard A., Clarys J. (1995) Co-contraction in the elbow and shoulder muscles during rapid cyclic movements in an aquatic environment. Journal of Electromyography and Kinesiology 5, 177-183 [DOI] [PubMed] [Google Scholar]
- Rouard A.H., Billat R.P., Deschodt V., Clarys J.P. (1997) Muscular activations during repetitions of sculling movements up to exhaustion in swimming. Archives of Physiology and Biochemistry 105, 655-662 [DOI] [PubMed] [Google Scholar]
- Ruwe P.A., Pink M., Jobe F.W., Perry J., Scovazzo M.L. (1994) The normal and the painful shoulders during the breaststroke. Electromyography and cinematographic analysis of twelve muscles. American Journal of Sports Medicine 22, 789-796 [DOI] [PubMed] [Google Scholar]
- Stirn I., Jarm T., Kapus V., Strojnik V. (2011) Evaluation of muscle fatigue during 100-m front crawl. European Journal of Applied Physiology 111, 101-113 [DOI] [PubMed] [Google Scholar]
- Takagi H., Sugimoto S., Nishijima N., Wilson B. (2004) Differences in stroke phases, arm-leg coordination and velocity fluctuation due to event, gender and performance level in breaststroke. Journal of Sport Biomechanics 3, 15-27 [DOI] [PubMed] [Google Scholar]
- Tesch P.A., Dudley G.A., Duvoisin M.R., Hather B.M., Harris R.T. (1990) Force and EMG signal patterns during repeated bouts of concentric or eccentric muscle actions. Acta Physiologica Scandinavica 138, 263-271 [DOI] [PubMed] [Google Scholar]
- Thompson K.G., Haljand R., MacLaren D.P. (2000) An analysis of selected kinematic variables in national and elite male and female 100-m and 200-m breaststroke swimmers. Journal of Sports Science 18, 421-431 [DOI] [PubMed] [Google Scholar]
- Thompson K.G., Haljand R., Lindley M. (2004) A comparison of Selected Kinematic variables between races in national to elite male 200m breaststroke swimmers. Journal of Swimming Research 16, 6-10 [Google Scholar]
- Wakayoshi K., Moritani T., Mutoh Y., Miyashita M. (1994) Electromyography evidence of selective muscle fatigue during competitive swimming. In:Medicine and Sports Science. Eds:Miyashita M., Mutoh Y., Richardson A.B.39, 16-23 [Google Scholar]
- Yoshizawa M., Tokuyama H., Okamoto T., Kumamoto M. (1976) Electromyographic study of the breaststroke. Biomechanics V-B. International series on Biomechanics. Baltimore, University Park Press; Komi P.V.1B, 222-279 [Google Scholar]