

Abstract
Social animals cooperate with bonding partners to outcompete others. Predicting a competitor's supporter is likely to be beneficial, regardless of whether the supporting relationship is stable or transient, or whether the support happens immediately or later. Although humans make such predictions frequently, it is unclear to what extent animals have the cognitive abilities to recognize others’ transient bond partners and to predict others' coalitions that extend beyond the immediate present. We conducted playback experiments with wild chimpanzees to test this. About 2 h after fighting, subjects heard recordings of aggressive barks of a bystander, who was or was not a bond partner of the former opponent. Subjects looked longer and moved away more often from barks of the former opponents’ bond partners than non-bond partners. In an additional experiment, subjects moved away more from barks than socially benign calls of the same bond partner. These effects were present despite differences in genetic relatedness and considerable time delays between the two events. Chimpanzees, it appears, integrate memories of social interactions from different sources to make inferences about current interactions. This ability is crucial for connecting triadic social interactions across time, a requirement for predicting aggressive support even after a time delay.
Keywords: social memory, cognition, coalitions, playback experiment, chimpanzee, third-party knowledge
1. Introduction
Social living incurs both costs and benefits [1]. On the one hand, social animals cooperate with group members to better detect predators or exploit food [2]. On the other hand, they compete with the same individuals over food, mating partners and shelter [3,4]. This dilemma has supposedly shaped behavioural strategies and cognitive abilities of group-living animals in order to better outcompete group members when facing a conflict of interest, without losing the benefit of group living [5–7].
When facing a conflict of interest, animals are viewed as going through a ‘decision-making’ process of whether or not to escalate the conflict to overt aggression [8,9]. Besides several factors related to costs and benefits, animals are more likely to engage in aggression when they have a greater likelihood of accessing the resource after the conflict [9]. Therefore, in order to outcompete the opponent successfully, it is important to correctly predict the outcome of a conflict situation. For example, many group-living animals are aware of their relative dominance compared with an opponent's (fishes [10], birds [11] and primates [12]). Some primates, in addition, are aware of third individuals likely to intervene on behalf of another against themselves [13–15]. Information about different types of relationship, however, might be obtained in different ways. Relationships involving oneself are based on one's own social interactions and could be a result of emotional [16–18] or cognitive experience [19]. Information about others’ relationships, such as their dominance, or sexual and affiliative relationships, can be only obtained through observations of interactions between third parties.
Eavesdropping, through observing others’ social interactions, is widespread through the animal kingdom [20]. Animals can extract information from eavesdropping about others’ conspicuous relationships [10,21] as well as about less conspicuous relationships [22–24]. Different types of relationship, however, are likely to require different monitoring effort. Conspicuous relationships, such as many dominance relationships, can be determined after the observation of a single interaction, such as a single aggression, supplant [10] or a ritualized subordination signal [25]. Sexual consortships are another type of conspicuous relationship. In baboons, for example, consortships are characterized by copulation, loud vocalizations and mate guarding. Subordinate male baboons seem to keep track of others’ consortships, and are apparently aware they can resume access to the consort female within hours of the end of the consortship [26]. In contrast to conspicuous dominance and consortship relationships, there is as yet no evidence of any rapid assessment of others’ less conspicuous relationships, such as social bonds. Rather, it is assumed that determining others’ social bonds requires repeated observations of the interactions between each dyad [22]. This is because social bonds are defined by the rate at which affiliative and agonistic behaviours, such as grooming and aggression, are exchanged. In many species, the mere occurrence of grooming or absence of aggression is not indicative of social bonds because they are exchanged within most dyads in a group, but at differing frequencies. Bonded dyads engage in grooming and aggression, as do non-bonded dyads. The difference is that bonded dyads engage in higher rates of grooming and lower rates of aggression [22]. Whether observing more conspicuous or less conspicuous relationships, animals use information obtained through eavesdropping to engage with or retreat from others [10,23,24,27,28].
Of particular importance in a conflict situation is whether one's opponent is likely to gain support from a bystander, as this will reduce one's chance of accessing the resource. Support—the propensity to intervene on behalf of another—is common in social mammals [28–32] and may bring reproductive benefits [32–34]. Support often occurs in dyads that share affiliative bonds and engage in other cooperative behaviours at high rates, such as grooming and food sharing [32–37]. In social mammals, support occurs predictably between close kin [23,28,33,37].
In some species, reproductive benefits are also evident for non-kin bonding partners (e.g. humans [38], macaques [32] and horses [34]), which (at least for males) may be gained through coalitionary support [32]. In chimpanzees, the majority of bonding partners are not close kin [35,37], and most support is between distantly related individuals. In comparison with close kin bonds, keeping track of others’ distantly related bonds seems more difficult for two reasons. First, bonds with distantly related partners are less enduring and are more likely to change over time than bonds with close kin [33,35]. Because distantly related bonds can suddenly form or break, they require more frequent monitoring than close kin bonds. Second, individuals may to some extent determine others’ close kin bonds by visual identification of morphological similarities shared between kin [39–41]. However, any morphological similarities will be less pronounced between distantly related individuals, thus detection of bonds between distantly related individuals is likely to be limited to observing their social interactions over time. In chimpanzees, the presence of either a close kin or a distantly related bonding partner affects the balance of power between competitors in a conflict situation, and can increase rank and mating opportunities [35,42]. Thus, it may be beneficial for chimpanzees to keep track of others’ distantly related as well as close kin bonding partners.
Spatial associations of group members are variable over time. Thus, the balance of power can change owing to supporters arriving in or leaving from the vicinity of the conflict opponents. Although it may pay for an individual to avoid open aggression when a supporting partner of the competitor is in the vicinity, aggression may provide gains once the competitor's supporter has left. Strategies of delayed aggression, delayed support or revenge [43] appear particularly adaptive in animals with a fission–fusion social structure where subgroup composition changes regularly, such as in chimpanzees, spider monkeys, some cetaceans or hyenas [44]. In chimpanzees, where party composition changes, on average, every 20–30 min [45], power balances can change many times throughout the day. Thus, it is likely to be adaptive for chimpanzees to delay further aggression and support in order to reduce costs of aggression and increase their chance to win the aggression.
In this study, we investigated whether chimpanzees could link two separate aggressive events involving two different opponents by the quality of the relationship between the two opponents, even if the opponents' relationships were less enduring and even if the events were separated by prolonged time periods, with interference from other social events. To this end, we used a playback experiment designed to simulate support against subjects, but with a time delay between a naturally occurring aggressive event and the simulated support (see the electronic supplementary material, figure S1). About 2 h after an unreconciled aggressive interaction between a subject and an opponent, we simulated the arrival of an aggressively motivated third individual in the vicinity of the subject by broadcasting his/her aggressive barks. Using a within-subjects design, this ‘newly arriving’ individual was a bond partner—either a close kin or a distantly related bond partner—of the former opponent in the test condition, and a non-bonded individual with respect to the former opponent in the control condition. We were interested in whether subjects were able to recognize the arrival of the third individual as a socially relevant event, not just because of the aggressive motivation it displayed, but also because of the social bond with the former opponent. If so, we expected subjects to show more attention and more aversive behaviour to newly arriving bond partners than non-bonded individuals of former opponents, despite the fact that both types of individuals were aggressively motivated. Specifically, we expected longer looking durations in the direction of the call provider (i.e. playback speaker) and more subsequent avoidance of the location. In addition, if subjects keep track of others’ bonded relationships, whether between close kin or distantly related partners, we expected that subjects should show more aversive behaviour to barks from a former opponent's bond than non-bond partner, whether or not they were closely related.
An alternative hypothesis that would have a similar expected behavioural reaction is that subjects associate the arrival of a call provider with the simultaneous arrival of the former opponent and react to the playback just because they expect the former opponent to arrive too—a reasonable assumption, because bonded individuals often travel together [46] (cf. [15]). To test this alternative hypothesis, we conducted a second playback experiment designed to simulate the arrival of the former opponent's bond partner, first with a socially benign call, and second, about an hour later, with an aggressive bark to simulate aggressive motivation typical of a support situation. Similar to the first playback experiment, we predicted that subjects would act aversively after hearing an aggressively motivated bond partner of the former opponent. In addition, however, we expected that, if subjects’ aversive responses would be merely owing to anticipating the presence of the former opponent, then subjects should react with aversion, regardless of the call provider's motivation (i.e. whether they produced a socially benign or aggressive call). If, however, subjects’ responses were not only based on assessments about party composition but included assessment of the call provider's motivation or intentions (i.e. the likelihood of call providers directing aggression towards subjects), then they should react weakly to playbacks of socially benign calls (bond partner present, but not aggressive) and aversively to playbacks of subsequent barks.
2. Material and methods
(a). Subjects and behavioural observations
Chimpanzee subjects were wild-living, habituated chimpanzees in the Sonso community in the Budongo forest, Uganda [47] from February 2008 to July 2010. Experiments were conducted from October 2008 to July 2010. Out of a total of 77 chimpanzees, we selected 16 subjects: nine adult females, three adult (more than 15 year) males and four subadult males (10–15 years). The first 16 individuals more than 10 years old observed in an aggression interaction that fulfilled the criteria for conducting playback experiments were selected as subjects.
R.M.W., C.C. and seven field assistants collected behavioural data in the form of 1 h focal samples [48] and ‘all occurrence’ data [48] of specific behaviours (aggression, grooming, resting in close proximity, support, food-sharing and pant–grunt vocalizations). Human observers followed individual chimpanzees in different parties, maintaining contact by using Motorola GP340 handheld radios via a transmitter station. Social interaction rates were calculated from focal animal data, where all social interactions with the focal animal were noted (see the electronic supplementary material, text S1). When in different subgroups, the identity of individuals that produced vocalizations and the reaction of focal animals were broadcasted over the radio. We calculated dominance hierarchies using pant–grunt vocalizations [25], a unidirectional indicator of dominance relationships (see the electronic supplementary material, text S2 and table S1). To describe the quality of social relationships, we calculated a ‘composite relationship index’ (CRI) [18,49] over quarter-year periods for all possible dyads across all three ‘participants’ (i.e. subjects, opponents and call providers). The CRI contrasts socio-positive and -negative behaviours in relation to average behaviour rates. A positive CRI indicated a more socio-positive relationship and a negative CRI showed the opposite (see the electronic supplementary material, text S3). The CRI was calculated from focal follows and ‘all occurrence’ data.
(i). Determining social bonds
‘Bond partners’ were defined as dyads having a net socio-positive relationship lasting at least six months (at least two consecutive blocks of three months). This can occur through either a mutual socio-positive relationship (CRI > 0) during the annual quarter of the experiment and the preceding quarter, or a large mutual socio-positive relationship (CRI > 10) during one of the quarters and a socio-neutral or positive relationship (CRI ≥ 0) during the other quarter [18,49].
(ii). Determining kinship
All five close kin bond dyads were mother–offspring pairs (r = 0.5), as determined from behavioural observations, parentage analyses using 19 autosomal microsatellites and the sharing of the same mitochondrial DNA haplotype (see the electronic supplementary material, table S2). It is difficult to determine kinship relations other than parent–offspring solely from genetic data. We therefore used parentage assignments and mtDNA and Y-chromosome haplotype sharing information to construct partial pedigrees, from which we calculated the maximum possible relatedness value of dyads, incorporating several simplifying assumptions based on known aspects of chimpanzee demography. Dyads with a relatedness value of 0 ≤ r ≤ 0.1875 (see the electronic supplementary material, text S4) were defined as distantly related dyads, similar to previous studies [18,49]. Distantly related dyads included non-kin and possible distant kin dyads [37]. We used the r-values as a proxy for possible perceptual similarities.
(b). Playback experiment
(i). Selection of playback stimuli
Most primate calls are individually distinctive and playback experiments have shown that primates rapidly recognize the identity of callers across a range of call types, from soft to loud calls [22]. Barks and socially benign calls used as playback stimuli were recorded opportunistically from known individuals using Sennheiser MKH416 and MKH418 microphones, and Marantz PMD 660 digital recorders. Digital sound files were saved in .wav format and used within 24 months from the time of the recording. After transferring the calls to a laptop computer, we used Cool Edit (Syntrillum, Phoenix, AZ) to screen for calls of high quality and free of undesired background noise. Playback stimuli consisted of a natural series of two consecutive barks that were produced during intra-group aggression. Bark series were selected to be similar in length and amplitude. All stimuli were calibrated to match the natural amplitude for each call type. Sound pressures at a distance of 1 m were mean ± s.d. = 91.1 dB ± 4.8 for barks. Calls were stored on an Apple iPod and broadcasted from a Nagra DSM speaker placed in a specially modified backpack for non-obstructed sound presentation.
(ii). Design of playback experiment 1
The experiment followed a within-group design, with each subject exposed to the same playback stimulus in two separate trials: test and control (see the electronic supplementary material, figure S1). Playback experiments (see the electronic supplementary material, table S3a) were carried out after subjects had either attacked or been attacked by a community member, the ‘opponent’. Attacks were defined as interactions that contained threats, displays or contact aggression. After such events, one observer followed the subject's opponent, and another located and followed the individual whose calls were about to be used as a playback stimulus (the ‘call provider’). Communication between observers was maintained using handheld radios, as described above (§2a).
Playbacks were conducted after the subject had separated from the opponent without any further interaction between them, the call provider had not been present during the aggression and was located more than 200 m away from the subject at the time of the playback (so that the call provider would not be able to hear the broadcast of his/her own call), and the subject was in a static position (resting, feeding). If these conditions were met, the speaker was hidden about 25 m from the subject in the general direction of the call provider's current location. The speaker was positioned at an angle of 90–180° from the subject's head orientation and about 90° (± 45°) from the subject's previous travel trajectory. The experimenter filmed the subject before (more than 10 s), during and after (more than 60 s) the playback presentation using a Panasonic NV-GS 330 DV camera. A second observer watched the subject from a different angle for additional observations.
We broadcast a brief series of two aggressive barks from the concealed speaker. Subjects heard the exact same stimuli in both the test and the control condition. The identity of the former opponents, however, changed between test and control conditions (see the electronic supplementary material, figure S1). In the test condition, the former opponent was a bond partner of the call provider, whereas in the control condition there was no bond between former opponent and call provider. A key condition in our experiment was that there was no apparent continuity between the original aggressive interaction and the subsequent playback experiment.
After each trial, the subject was followed for 60 min under a focal animal sampling regime. To avoid habituation to any of the playback stimuli, we conducted experiments below the frequency of naturally occurring barks, and none of the subjects was tested on consecutive days. Across trials, former opponents were matched for sex, age and relative dominance rank, whereas the order of presentation was counterbalanced across subjects (see the electronic supplementary material, text S5 and table S4). In addition, we tested whether factors relating to the experimental set-up, the preceding aggression or the relative dominance relationships within the triad [50] affected the results (see the electronic supplementary material, text S5, and tables S4 and S5). All subjects and variables are presented in the electronic supplementary material, table S3a.
(iii). Design of playback experiment 2
We conducted a second playback experiment that generally followed the same design as experiment 1 with the difference that each subject heard two independent playbacks following the same aggression. About 90 min after the aggression, subjects first heard a short series of one to four socially benign calls (four experiments with travel hoos and one experiment with food grunts; see the electronic supplementary material, table S3b) by the former opponent's bond partner. About 1 h after the socially benign call playback, subjects heard two aggressive barks from the same individual.
(c). Data analysis
(i). Variable extraction
Using Adobe Premier Pro video software, two observers, R.M.W. and C.C., extracted the response variables from the videos using a frame-by-frame method (25 frames per second). We calculated the total duration of all looks to the speaker by the subject during the first 60 s after the playback. Using video and focal observations, we determined, in addition, whether the first move of the subject was towards the speaker (moving to the speaker with a deviation of 0 ± 45°), parallel to the speaker (90 ± 45°) or away from the speaker (180 ± 45°). We used both variables in the statistical models and tests. Furthermore, we used the focal data of the subject during the first hour after the playback to see whether or not the subject joined the same party with the former opponent or the call provider, and whether the subject approached or retreated in response to their presence.
(ii). Hypotheses testing
In order to test our central hypothesis, we conducted three analyses with data from experiment 1 and one analysis with data from experiment 2. First, we investigated the general influence of the quality of the social relationship between the ‘participants’ (i.e. the subject, opponent and the call provider) on subjects’ responses to the playback. We used a multivariate general linear model (GLM) and included two response variables: the duration subjects looked to the speaker in the first minute and the direction of subjects’ first move towards, parallel or away from the speaker after the playback. The CRI value of each of the three pairs of participants involved in the triad and the subject's identity were included as predictor variables.
Second, to examine whether chimpanzees recognize affiliative relationships that fluctuate in strength over time as well as permanent close kin bonds, we conducted a within-subjects comparison in which we treated subjects tested with close kin bond or distantly related bond recognition separately.
In a third analysis, we investigated whether subjects met the call provider or former opponent in the hour following an experiment. We compared test and control trials in a linear mixed model (LMM), with subject identity as a random factor.
Finally, to exclude the possible explanation that subjects simply associated the arrival of the call provider with the simultaneous arrival of the former opponent, we ran another multivariate GLM using the results of experiment 2, including the same two response variables: the duration subjects looked to the speaker in the first minute and the direction of subjects’ first move towards, parallel or away from the speaker after the playback. We used the type of vocalization broadcast as a playback stimulus (socially benign call or bark) and the subjects’ identity as predictor variables. All tests were conducted using SPSS v. 20.
3. Results
(a). Playback experiment 1: aggression from bond versus non-bond partner
We conducted a total of 32 playback experiments in 16 matched pairs (males n = 7, females n = 9), of which half simulated that a close kin bond partner (n = 8) and the other half a distantly related bond partner (n = 8) barking at the subject in the test condition. The order of presentation of the conditions was counterbalanced between subjects (subjects heard test first: n = 9, subjects heard control first: n = 7). Latencies between the aggressive interaction and the playback ranged from 30 to 300 min (see the electronic supplementary material, table S3a; test: mean ± s.d. = 125 ± 60 min, range: 30–300 min; control: 90 ± 50 min, range: 30–180 min) and did not predict variation in responses (the electronic supplementary material, table S5a).
(i). Influence of the quality of social relationships
When investigating the general influence of the quality of the social relationship between the ‘participants’ (i.e. the subject, opponent and the call provider) on subjects’ responses to the playback, a GLM revealed that variation in subjects’ responses was significantly explained by the CRI values between the former opponent and the call provider explaining 65% of variance. Subjects looked for longer to the speaker (β = 0.324, t = 3.66, p = 0.003; figure 1) and were more likely to move away from the speaker (β = 0.019, t = 3.692, p = 0.003) the higher the CRI values were between the former opponent and the call provider (table 1). The subjects’ own CRI values with the call provider or opponent had no effect across conditions (table 1 and figure 1).
Figure 1.
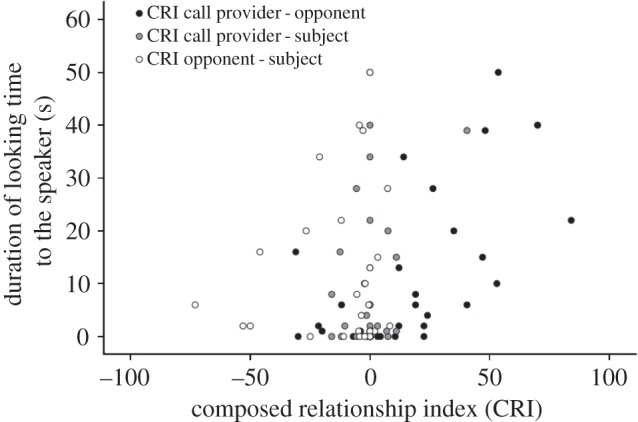
Subjects' total duration of looking to the speaker during the first minute after the playback depending on the composite relationship index (CRI) between the three dyads of the triadic interaction.
Table 1.
The influence of social relationship strength between subjects, opponents and call providers on subjects’ responses (figure 2). Results of a multivariate GLM using two response variables: (i) duration of subjects’ looking time to the speaker in the first 60 s post playback of the bark stimulus, and (ii) direction of the first move subjects took following the playback, whether towards, parallel or away from the playback speaker. Social relationship strength is calculated using the composite relationship index (CRI). Dyads with strong, compared with weak, relationships have high, compared with low, rates of cooperative behaviours. Although we excluded cases in which subjects were bond partners with their former opponents or the call providers, the relationship quality with the subject could still vary by subjects having a neutral or negative relationship with opponents or call providers. Bold: p < 0.05.
| predictor | Pillai value | F | d.f. | p |
|---|---|---|---|---|
| relationship strength: subject–opponent | 0.108 | 0.73 | 1 | 0.505 |
| relationship strength: subject–call provider | 0.031 | 0.20 | 1 | 0.826 |
| relationship strength: opponent–call provider | 0.650 | 11.2 | 1 | 0.002 |
| subject identity | 0.987 | 0.84 | 15 | 0.675 |
(ii). The role of kin relations and the stability of social relations
We tested whether subjects responded to distantly related bonds as well as close kin bonds of potential coalition partners. Results showed that subjects looked significantly longer to the speaker, and were more likely to move away from it, if confronted with aggressive barks of an opponent's close kin bond partner than of an opponent's non-bond partner (figure 2; Wilcoxon exact tests: looking duration to speaker during first minute: T+ = 3, ties = 0, n = 8, p = 0.039; moving away from speaker: T+ = 0, ties = 1, n = 8, p = 0.016). We found the same effect if we compared the distantly related bond partner condition with the non-bonded partner condition (figure 2; Wilcoxon exact tests: looking duration: T+ = 2, ties = 0, n = 8, p = 0.023; moving away: T+ = 0, ties = 2, n = 8, p = 0.031). Indeed, when opponent and call provider were not bond partners, subjects were more likely to move towards the speaker or parallel to the speaker than to move away from it (figure 2).
Figure 2.

Subjects’ responses depending on the social relationship between call provider and the former opponent. (a) Subjects’ responses (n = 8) to call providers when they are either distantly related bond partners or non-bonded individuals to the former opponent. (b) Subjects’ responses (n = 8) to call providers when they are either close kin bond partners or non-bonded individuals to the former opponent. Two different behavioural responses are shown: (i) duration of looking to the speaker during the first 60 s, and (ii) direction of first move after the playback experiment. Box plot represents the second and third quartile around the median, and error bar represents the standard deviation; dots represent outliers.
In this study of the relationships between the former opponent and playback provider, the seven distantly related bonded relationships lasted for 0.5–1.5 years (mean = 1 year) of the study period (2.25 year), whereas the four close kin bonded relationships lasted the whole length of the study period. In the former, bonds could have extended either before or after the study period, whereas in the latter, bonds could have extended both before and after the study period.
(iii). Longer-term effects
In a third analysis, we investigated whether subjects met the call provider or former opponent in the hour following an experiment. When comparing test and control trials in an LMM, subjects were significantly less likely to meet the call provider if the call provider was a bond partner of the former opponent than if he was not (figure 3; F32 = 8.23, p = 0.007; estimate β = 0.47, t = 2.87, p = 0.007). Interestingly, subjects did not show any difference in their likelihood to meet up with the former opponent across conditions (figure 3; F32 = 0.23, p = 0.635; estimate β = 0.08, t = 0.48, p = 0.635). This is consistent with the rationale that subjects expected similar (socio-negative) behaviour from the opponent across conditions given that the context remained the same in both conditions (meeting after unreconciled aggression). By contrast, the results suggest that subjects anticipated different behaviour from the call provider across conditions, in line with the differing relationships between call providers and former opponents (that is, whether they were bond partners or not).
Figure 3.

Subjects’ responses within the first hour after hearing the bark playback. Panels show how often 16 subjects met (a) the call provider (where ‘met’ means the two were >200 m apart during the playback but now join the same subgroup and see each other), or (b) the former opponent, when the two were either bonded or not bonded.
(b). Playback experiment 2: socially benign versus aggressive vocalizations
We conducted a total of 10 playback trials with five subjects (males n = 1, females n = 4), first broadcasting a series of socially benign calls (average latency after the aggression ± s.d. = 93 ± 58 min), followed, after about an hour, by two aggressive barks (average latency after the socially benign call ± s.d. = 69 ± 36 min) from the same individual (see the electronic supplementary material, table S3b). Because both playbacks followed the same aggressive event, we neither control for the preceding aggression nor for the experimental set-up.
Results of the GLM showed that the vocalization type used as a playback stimulus had a significant effect on the first move of the subject, explaining about 80% of the variance (F = 16.0, d.f. = 1, p = 0.016), with subjects more likely to move away from the speaker after the barks compared with the socially benign calls (estimate β = 0.8, t = 4.0, p = 0.016). By contrast, looking duration did not vary between playbacks of barks and socially benign calls (F = 1.21, d.f. = 1, p = 0.333). Although subjects always heard the socially benign calls first, they were significantly more likely to move away from the speaker when subsequently hearing a series of barks from the same individual. This was in spite of the time delay between the initial aggression and playback always being longer for the barks than for the socially benign calls.
4. Discussion
In these experiments, we have shown that about 2 h after an unreconciled aggressive event, chimpanzees expressed aversive behaviour when hearing the simulated arrival of another, aggressively motivated group member, provided this individual shared a close social bond with the former opponent. Because we ensured that the call providers were not present during the initial aggression, subjects had to base their responses on their general knowledge of others’ social relationships. Equally relevant, we were able to rule out that subjects’ responses were simply driven by their own relationship to the call provider or by the acoustic properties of the playback stimuli, because the same bark recording was played in both the test and the control conditions.
In the second playback experiment, we showed that the arrival of the former opponent's bond partner triggered the subjects’ attention whether subjects heard a benign or an aggressive call. An aversive reaction of moving away from the speaker was only observed after subsequently hearing the same call providers’ bark vocalizations. As with the first playback experiment, this suggests that subjects were neither averse to the call provider per se, nor to their presence after fighting with their bond partner. Taking the two playback experiments together, subjects showed aversion only when the call provider was a bond partner of their former opponent and was aggressively motivated. This suggests that subjects remembered the aggression from hours earlier, linked it to their stored knowledge of their previous opponents’ third-party relationships and to the relevance of the current bark, and inferred whether it might or might not be directed towards themselves [13,15]. Subjects’ responses to the playbacks are consistent with responses expected in a situation of delayed vocal support.
Our results provide experimental evidence that chimpanzees are aware of the possible impact others’ behaviour has on them, and can attain this awareness by remembering and linking specific social interactions with different individuals across time. This confirms observational results from a chimpanzee community in Taï, where social bonding partners were able to reconcile conflicts between their bonding partner and his former opponent [51]. Although the underlying mechanisms remain unclear, such as whether explicit or implicit memory is involved [20,52], these memories apparently contain details about the type of social interaction, the participants’ identity and social relations, and, most impressively, the triadic implications of an interaction (in this case, an extended expectation of delayed aggression from individuals bonded with the former opponents). This was in spite of experiencing and witnessing multiple interactions in the intervening period (see electronic supplementary material, text S1), where such interference may have accelerated memory decay [53]. While we have not tested the upper limit of chimpanzees’ social memory, these results extend the operational window through which the specific details of chimpanzees’ social world remain salient. In addition, these results suggest that chimpanzees are familiar enough with triadic behavioural mechanisms, such as delayed support, to assess whether or not they are the target of aggression and that the possibility of third-party aggression remains salient over hours. Whether this is also the case for other delayed third-party interventions, such as reconciliation or revenge, remains to be tested.
We have shown that chimpanzees are aware of bonded relationships between others. There is good evidence that primates and other mammals are generally able to keep track of others’ affiliative close kin relationships (e.g. hyenas [13] and monkeys [15,21,24]; for review see [22]), even though, unlike with dominance, they are less conspicuous relationships. This is particularly interesting in view of cognitive abilities. Detecting less conspicuous affiliative bonds seems to be a complex problem, because there is no single or specific behaviour that allows one to distinguish between a closely bonded pair and a non-bonded pair [22]. Alternatively, being the recipient of a coalitionary attack may contain such strong negative emotional valence that victims may remember it across time. If this was the case, then one might expect that victims remembered the aggression towards themselves rather than the bonding behaviour between others [52]. There are two points that weaken this argument. First, coalitionary attacks do not always occur simultaneously but can be separated in time [15,54]. Second, apparent coalitionary attacks are not always conducted by bond partners, but sometimes occur opportunistically, by two non-bonded ‘enemies’ of the victim [55]. Thus, only monitoring patterns of aggression is unlikely to be a reliable system for assessing potential attacks towards oneself.
While the ability to keep track of kin bonds is evident in several mammal species, the recognition of others’ distantly related social bond partners has not yet been documented (cf. [26]). Our results showed that subjects’ reactions were similar whether the simulated delayed support was between close kin or distantly related bond partners, showing that, at least in the context of support, chimpanzees treat others’ distantly related bonds as being similar to close kin bonds. Keeping track of others’ bonded relationships with distantly related partners seems to be an even more complex problem than bonds between close kin. First, the number of possible distantly related bonding partners exceeds multiple times the number of close kin pairs [37]. Second, distantly related bonds can be more transient (see the electronic supplementary material, table S2) and exhibit greater quality changes over time [33,35] than close kin bonds. Taking these two cognitive tasks together, the increase in the number of potential bonding partners and the relative transience of distantly related bonded relationships compared with close kin relationships, we suggest that for chimpanzees, keeping track of others’ less conspicuous bonded relationships is likely to require more frequent monitoring effort of their social environment than in species that predominantly rely on close kin bonds. The monitoring task is further complicated, however, by the fission–fusion nature of the chimpanzee's social structure, where not everybody can be monitored all the time [44,56].
One relevant question is how much of the monitoring of others’ social relationships that we have observed can be ascertained by a rule based on a single parameter and how much requires more complex cognition, such as integration of stored information from different sources [46,50,57]. We checked whether single rules might be driving subjects’ responses to the playbacks in two ways. First, although there are no predictable rank relationships between bond partners in chimpanzees [31], we determined that subjects’ response to the playback stimuli were not driven by simple rules of relative rank. We controlled for this within the experimental design such that relative ranks did not vary between conditions within subjects (see the electronic supplementary material, table S3a). Second, in the second playback experiment (see the electronic supplementary material, table S3b), we demonstrated, as in a previous study [15], that our results were not purely based on the simple rule of spatial proximity patterns of ‘who travels with whom’, but included an awareness of the consequences of others’ motivations in a triadic context.
Our results extend the literature in four ways. First, chimpanzees appear to link specific hours-old memories of their own interactions to their general knowledge of others’ interactions to make inferences about a current social situation. Although it is likely that an emotional component may be aiding memory [58], emotion is likely to have a less direct influence in such a triadic interaction than a dyadic interaction. Second, triadic social interactions are operational over hours, adding significantly to the complexity of social life, and probably the cognition required to resolve social problems. Third, chimpanzees simultaneously keep track not only of others’ close kin relationships, but also of others’ more transient, distantly related bonded relationships. Fourth, predictions of whether a bystander will act as friend or foe are not always based on one's own relationship with the bystander, but also on the bystander's relationships to one's other recent interaction partners.
It is often stated that successful coordination of coalitions to outcompete conspecifics may be among the most cognitively demanding tasks faced by animals [22,36,59]. Chimpanzees apparently retain access to many past social interactions, both those they have engaged in and those they have observed. They then assess the relevance of these memories to any current interaction, even those with different individuals, even with individuals they have not recently interacted with, and even when relevance judgements may not relate to a current emotional state. Evidence that triadic interventions can be delayed over time implies that social events that happen hours apart, and even involving different individuals, may nonetheless be connected in an animals’ mind. Whether this capacity has evolved in species where group members are not always visible to each other, such as for animals living in fission–fusion social groups [44,56], or is more widespread, remains to be tested. Nonetheless, these results have considerable implications when examining animal behaviour and social cognition. Observed social interactions may not always be based on recent social information but may extend to social information from the past.
Acknowledgements
We thank S. Adue, J. Alyo, M. Gideon and J. Okuti for their support, C. Rowney for laboratory work, V. Reynolds, L. Vigilant and Z. Zommers for providing faecal samples, D. Cheney and J. Call for very insightful comments on the manuscript, Budongo Conservation Field Station, and the Ugandan Authorities (UWA, UNCST) for permission to conduct the study. Genetic analyses were funded by the Max Planck Society.
Funding statement
The study was supported by the Leverhulme Trust (Research Leadership Award), the British Academy and the Leakey Foundation. The data of this study are available on request. We acknowledge the Royal Zoological Society of Scotland for providing core funding to the Budongo Conservation Field Station.
References
- 1.Alexander RD. 1974. The evolution of social behaviour. Annu. Rev. Ecol. Syst. 5, 325–383 (doi:10.1146/annurev.es.05.110174.001545) [Google Scholar]
- 2.Clutton-Brock T. 2009. Cooperation with non-kin in animal societies. Nature 462, 51–57 (doi:10.1038/nature08366) [DOI] [PubMed] [Google Scholar]
- 3.Sterck EHM, Watts DP, van Schaik CP. 1997. The evolution of female social relationships in nonhuman primates. Behav. Ecol. Sociobiol. 41, 291–309 (doi:10.1007/s002650050390) [Google Scholar]
- 4.Clutton-Brock TH, et al. 2006. Intrasexual competition and sexual selection in cooperative mammals. Nature 444, 1065–1068 (doi:10.1038/nature05386) [DOI] [PubMed] [Google Scholar]
- 5.Jolly A. 1966. Lemur social behaviour and primate intelligence. Science 153, 501–506 (doi:10.1126/science.153.3735.501) [DOI] [PubMed] [Google Scholar]
- 6.Dunbar RIM, Schultz S. 2007. Evolution in the social brain. Science 317, 1344–1347 (doi:10.1126/science.1145463) [DOI] [PubMed] [Google Scholar]
- 7.Cheney DL. 2011. Extend and limits of cooperation in animals. Proc. Natl Acad. Sci. USA 108, 10 902–10 909 (doi:10.1073/pnas.1100291108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.de Waal FBM. 2000. Primates: a natural heritage of conflict resolution. Science 289, 586–590 (doi:10.1126/science.289.5479.586) [DOI] [PubMed] [Google Scholar]
- 9.Wittig RM, Boesch C. 2003. ‘Decision-making’ in conflicts of wild chimpanzees (Pan troglodytes): an extension to the relational model. Behav. Ecol. Sociobiol. 54, 491–504 (doi:10.1007/s00265-003-0654-8) [Google Scholar]
- 10.Oliveira RF, McGregor PK, Latruffe C. 1998. Know thine enemy: fighting fish gather information from observing conspecific interaction. Proc. R. Soc. Lond. B 265, 1045–1049 (doi:10.1098/rspb.1998.0397) [Google Scholar]
- 11.Paz-y-Miño G, Bond AB, Kamil AC, Balda RP. 2004. Pinyon jays use transitive inference to predict social dominance. Nature 430, 778–781 (doi:10.1038/nature02723) [DOI] [PubMed] [Google Scholar]
- 12.Silk JB. 1999. Male bonnet macaques use information about third-party rank relationships to recruit allies. Anim. Behav. 58, 45–51 (doi:10.1006/anbe.1999.1129) [DOI] [PubMed] [Google Scholar]
- 13.Engh AL, Hoffmeier RR, Cheney DL, Seyfarth RM. 2006. Who, me? Can baboons infer the target of vocalizations? Anim. Behav. 71, 381–387 (doi:10.1016/j.anbehav.2005.05.009) [Google Scholar]
- 14.Judge PG, Mullen SH. 2005. Quadratic postconflict affiliation among bystanders in a hamadryas baboon group. Anim. Behav. 69, 1345–1355 (doi:10.1016/j.anbehav.2004.08.016) [Google Scholar]
- 15.Wittig RM, Crockford C, Seyfarth RM, Cheney DL. 2007. Vocal alliances in chacma baboons. Behav. Ecol. Sociobiol. 61, 899–909 (doi:10.1007/s00265-006-0319-5) [Google Scholar]
- 16.De Waal FBM. 2000. Attitudinal reciprocity in food sharing among brown capuchin monkeys. Anim. Behav. 60, 253–261 (doi:10.1006/anbe.2000.1471) [DOI] [PubMed] [Google Scholar]
- 17.Schino G, Aureli F. 2009. Reciprocal altruism in primates: partner choice, cognition, and emotions. Adv. Stud. Behav. 39, 45–69 (doi:10.1016/s0065-3454(09)39002-6) [Google Scholar]
- 18.Crockford C, Wittig RM, Langergraber KE, Ziegler TE, Zuberbühler K, Deschner T. 2013. Urinary oxytocin and social bonding in related and unrelated wild chimpanzees. Proc. R. Soc. B 280 (doi:10.1098/rspb.2012.2765) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cheney DL, Moscovice LR, Heesenc M, Mundry R, Seyfarth RM. 2010. Contingent cooperation between wild female baboons. Proc. Natl Acad. Sci. USA 107, 9562–9566 (doi:10.1073/pnas.1001862107) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Shettleworth S. 2010. Cognition, evolution and behaviour. Oxford, UK: Oxford University Press [Google Scholar]
- 21.Bergman TJ, Beehner JC, Cheney DL, Seyfarth RM. 2003. Hierarchical classification by rank and kinship in baboons. Science 302, 1234–1236 (doi:10.1126/science.1087513) [DOI] [PubMed] [Google Scholar]
- 22.Cheney DL, Seyfarth RM. 2007. Baboon metaphysics: the evolution of a social mind. Chicago, IL: Chicago University Press [Google Scholar]
- 23.Engh AL, Siebert ER, Greenberg DA, Holekamp KE. 2005. Patterns of alliance formation and post-conflict aggression indicate spotted hyenas recognize third-party relationships. Anim. Behav. 69, 209–217 (doi:10.1016/j.anbehav.2004.04.013) [Google Scholar]
- 24.Wittig RM, Crockford C, Wikberg E, Seyfarth RM, Cheney DL. 2007. Kin-mediated reconciliation substitutes for direct reconciliation in female baboons. Proc. R. Soc. B 274, 1109–1115 (doi:10.1098/rspb.2006.0203) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wittig RM, Boesch C. 2003. Food competition and linear dominance hierarchy among female chimpanzees of the Taï National Park. Int. J. Primatol. 24, 847–867 (doi:10.1023/A:1024632923180) [Google Scholar]
- 26.Crockford C, Wittig RM, Seyfarth RM, Cheney DL. 2007. Baboons eavesdrop to deduce mating opportunities. Anim. Behav. 73, 885–890 (doi:10.1016/j.anbehav.2006.10.016) [Google Scholar]
- 27.Peak TM, Terry AMR, McGregor PK, Dabelsteen T. 2002. Do great tits assesses rivals by combining direct experience with information gathered by eavesdropping? Proc. R. Soc. Lond. B 269, 1925–1929 (doi:10.1098/rspb.2002.2112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Connor RC, Heithaus MR, Barre LM. 2001. Complex social structure, alliance stability, and mating access in a bottlenose dolphin ‘super-alliance’. Proc. R. Soc. Lond. B 268, 263–267 (doi:10.1098/rspb.2000.1357) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Harcourt AH, de Waal FBM. 1992. Coalitions and alliances in humans and other animals. Oxford, UK: Oxford University Press [Google Scholar]
- 30.Feh C. 1999. Alliances and reproductive success in Camargue stallions. Anim. Behav. 57, 705–713 (doi:10.1006/anbe.1998.1009) [DOI] [PubMed] [Google Scholar]
- 31.Duffy KG, Wrangham RW, Silk JB. 2007. Male chimpanzees exchange political support for mating opportunities. Curr. Biol. 17, R586–R587 (doi:10.1016/j.cub.2007.06.001) [DOI] [PubMed] [Google Scholar]
- 32.Schülke O, Bhagavatula J, Vigilant L, Ostner J. 2010. Social bonds enhance reproductive success in male macaques. Curr. Biol. 20, 2207–2210 (doi:10.1016/j.cub.2010.10.058) [DOI] [PubMed] [Google Scholar]
- 33.Silk JB, et al. 2009. The benefits of social capital: close social bonds among female baboons enhance offspring survival. Proc. R. Soc. B 276, 3099–3104 (doi:10.1098/rspb.2009.0681) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cameron EZ, Setsaas TH, Linklater WL. 2009. Social bonds between unrelated females increase reproductive success in feral horses. Proc. Natl Acad. Sci. USA 106, 13 850–13 853 (doi:10.1073/pnas.0900639106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Mitani JC. 2009. Male chimpanzees form enduring and equitable social bonds. Anim. Behav. 77, 633–640 (doi:10.1016/j.anbehav.2008.11.021) [Google Scholar]
- 36.Gilby IC, et al. 2013. Fitness benefits of coalitionary aggression in male chimpanzees. Behav. Ecol. Sociobiol. 67, 373–381 (doi:10.1007/s00265-012-1457-6) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Langergraber KE, Mitani JC, Vigilant L. 2007. The limited impact of kinship on cooperation in wild chimpanzees. Proc. Natl Acad. Sci. USA 104, 7786–7790 (doi:10.1073/pnas.0611449104) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Manson JH, Wrangham RW. 1991. Intergroup aggression in chimpanzees and humans. Curr. Anthropol. 32, 369–390 (doi:10.1086/203974) [Google Scholar]
- 39.Barrett L, Henzi P, Rendall D. 2007. Social brains, simple minds: does social complexity really require cognitive complexity? Phil. Trans. R. Soc. B 362, 561–575 (doi:10.1098/rstb.2006.1995) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kazem AJM, Widdig A. 2013. Visual phenotype matching: cues to paternity are present in rhesus macaque faces. PLoS ONE 8, e55846 (doi:10.1371/journal.pone.0055846) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pfefferle D, Ruiz-Lambides AV, Widdig A. 2014. Female rhesus macaques discriminate unfamiliar paternal sisters in playback experiments: support for acoustic phenotype matching. Proc. R. Soc. B 281, 20131628 (doi:10.1098/rspb.2013.1628) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Nishida T. 1983. Alpha status and agonistic alliance in wild chimpanzees (Pan troglodytes schweinfurthii). Primates 24, 318–336 (doi:10.1007/BF02381978) [Google Scholar]
- 43.de Waal FBM, Luttrell LM. 1988. Mechanisms of social reciprocity in three primate species: symmetrical relationship characteristics or cognition? Ethol. Sociobiol. 9, 101–118 (doi:10.1016/0162-3095(88)90016-7) [Google Scholar]
- 44.Aureli F, et al. 2008. Fission–fusion dynamics. Curr. Anthropol. 49, 627–654 (doi:10.1086/586708) [Google Scholar]
- 45.Lehmann J, Boesch C. 2004. To fission or to fusion: effects of community size on wild chimpanzee (Pan troglodytes verus) social organisation. Behav. Ecol. Sociobiol. 56, 207–216 (doi:10.1007/s00265-004-0781-x) [Google Scholar]
- 46.Schusterman RJ, Reichmuth C, Kastak C, Kastak D. 2003. Equivalence classification as an approach to social knowledge: from sea lions to simians. In Animal social complexity (eds Waal FBM, Tyack PL.), pp. 179–206 Cambridge, MA: Harvard University Press [Google Scholar]
- 47.Reynolds V. 2005. Chimpanzees of the Budongo forest. Oxford, UK: Oxford University Press [Google Scholar]
- 48.Altman J. 1974. Observational study of behaviour: sampling methods. Behaviour 49, 227–267 (doi:10.1163/156853974X00534) [DOI] [PubMed] [Google Scholar]
- 49.Crockford C, Wittig RM, Mundry R, Zuberbühler K. 2012. Wild chimpanzees inform ignorant group members of danger. Curr. Biol. 22, 142–146 (doi:10.1016/j.cub.2011.11.053) [DOI] [PubMed] [Google Scholar]
- 50.Range F, Noë R. 2005. Can simple rules account for the pattern of triadic interactions in juvenile and adult female sooty mangabeys? Anim. Behav. 69, 445–452 (doi:10.1016/j.anbehav.2004.02.025) [Google Scholar]
- 51.Wittig RM, Boesch C. 2010. Receiving post-conflict affiliation from the enemy's friend reconciles former opponents. PLoS ONE 5, e13995 (doi:10.1371/journal.pone.0013995) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Manning JR, Polyn SM, Baltuch GH, Litt B, Kahana MJ. 2010. Oscillatory patterns in temporal lobe reveal context reinstatement during memory search. Proc. Natl Acad. Sci. USA 108, 12 893–12 897 (doi:10.1073/pnas.1015174108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Stevens JR, Hauser MD. 2004. Why be nice? Psychological constraints on the evolution of cooperation. Trends Cogn. Sci. 8, 60–65 (doi:10.1016/j.tics.2003.12.003) [DOI] [PubMed] [Google Scholar]
- 54.Cheney DL, Seyfarth RM. 1986. The recognition of social alliances by vervet monkeys. Anim. Behav. 34, 1722–1731 (doi:10.1016/S0003-3472(86)80259-7) [Google Scholar]
- 55.de Waal FBM, van Hoof JARAM. 1981. Side-directed communication and agonistic interactions in chimpanzees. Behaviour 77, 164–198 (doi:10.1163/156853981X00211) [Google Scholar]
- 56.Amici F, Aureli F, Call J. 2008. Fission–fusion dynamics, behavioral flexibility, and inhibitory control in primates. Curr. Biol. 18, 1415–1419 (doi:10.1016/j.cub.2008.08.020) [DOI] [PubMed] [Google Scholar]
- 57.Turesson HK, Ghazanfar AA. 2011. Statistical learning of social signals and its implications for the social brain hypothesis. Interact. Stud. 12, 397–417 (doi:10.1075/is.12.3.02tur) [Google Scholar]
- 58.Ross HE, Young LJ. 2009. Oxytocin and the neural mechanisms regulating social cognition and affiliative behavior. Front. Neuroendocrinol. 30, 534–547 (doi:10.1016/j.yfrne.2009.05.004) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Barrett L, Henzi P, Dunbar RIM. 2003. Primate cognition: from ‘what now?’ to ‘what if?’ Trends Cogn. Sci. 7, 494–497 (doi:10.1016/j.tics.2003.09.005) [DOI] [PubMed] [Google Scholar]