

Abstract
Short sleep/dark durations are common in modern society. In a within subjects counterbalanced design, we compared the effect of 7–19 nights with a late bedtime (01:00) versus 7–19 nights with an early bedtime (22:00) on the human dim light melatonin rhythm. All ten healthy subjects experienced lights on time at 07:00 and at least 5 min of outdoor light between 07:00 and 08:00. We found that despite the morning light exposure, the melatonin rhythm following the late bedtime nights was delayed by ~0.6 h (P < 0.05), as compared to after the early bedtime nights. These results suggest that when people truncate their sleep by delaying their bedtime they also significantly phase delay their circadian rhythms and/or when people advance their bedtime their circadian rhythms advance.
Keywords: Circadian rhythm, Dark, Light, Melatonin, Night, Sleep
Modern society is chronically sleep deprived. In a recent National Sleep Foundation (NSF) poll the average nightly sleep duration was only 7 h on weeknights, and nearly a third reported regularly sleeping 6 h or less per weeknight [13]. In contrast, experimental subjects (with no work and social commitments), when free to choose their bed and wake times, sleep for approximately 9 h per night (e.g. Refs. [4,18]). Shorter sleep durations in the NSF poll were related to longer working hours [13]. This study investigated the effects of multiple days of short nocturnal dark periods on the circadian systems of people with normal sleep lengths.
Melatonin is synthesized and released from the pineal gland [11]. Melatonin secretion is suppressed by light [9], but the circadian rhythm of melatonin in dim light is a reliable marker of the phase of the circadian clock. Previous work has shown that the dim light melatonin rhythm in humans can be influenced by prior sleep times. In one study the duration of the dim light melatonin rhythm increased by 1.6 h, following 4 weeks with a very early bedtime (dark 18:00–08:00) as compared to after 1 week with a 24:00 bedtime (dark 24:00–08:00) [17]. This increase in duration was largely due to an advance in the onset of the melatonin rhythm. In a similar but shorter study, 9 long nights with an even earlier bedtime (dark 16:00–08:00) did not increase the duration of melatonin secretion, but the melatonin rhythm also advanced, this time by 2.4 h [15]. Finally, another study found that the duration of the dim light melatonin rhythm was significantly shorter in habitual short sleepers (<6 h/night) than in habitual long sleepers (>9 h/night) [1]. It remains unclear if this difference is an intrinsic characteristic of these groups or if the sleep times they habitually adopt have produced the difference in their melatonin rhythms.
Many day workers probably truncate their sleep by delaying their bedtime, but maintain a set wake time, because of 9–5 work commitments. Thus we investigated the effects of a regular early versus late bedtime with a fixed morning lights on time on the human dim light melatonin rhythm. Ten (five male, five female) healthy young adults (mean age ± SD 29.4 ± 6.5 years; BMI 23.1 ± 3.6 kg/m2) participated. They were nonsmokers, medication free, did not consume large caffeine doses (<300 mg/day) and reported no medical, psychiatric or sleep disorders. One female subject was taking an oral contraceptive. The menstrual phase of the other four female subjects during the study was not controlled, as menstrual phase does not affect the melatonin rhythm (e.g. Ref. [14]). No subject had worked night shifts in the previous 2 months, or traveled across more than two time zones in the previous month. Morningness–eveningness was assessed [8] and there were four neither, five moderate morning and one definite morning type. The self-reported mean habitual weekday sleep schedule of the subjects was 22:50 ± 0.7 h to 07:00 ± 0.8 h. These times fall within the range of weekday bed and wake times that at least 66% of people reported in the NSF poll (22:00–24:00 and 05:00–07:59, respectively [13]). The protocol was approved by the Rush University Medical Center Institutional Review Board, and all subjects gave written informed consent prior to participation.
As part of a within subjects design, subjects participated in a series of nights with an early bedtime (sleep/dark: 22:00–07:00, 9 h) and a late bedtime (sleep/dark: 01:00–07:00, 6 h). Six subjects completed the early bedtime nights first; one for 7 nights, one for 10 nights, two for 13 nights and two for 19 nights, followed by the same number of late bedtime nights. Four subjects participated in the late bedtime nights first; one for 7 nights, two for 10 nights and one for 13 nights, followed by the same number of early bedtime nights. There was no washout period between the early and late bedtime nights. In order to avoid excessive sleep deprivation, napping during the days with the late bedtime was permitted between 14:30 and 17:30, centered 12 h from the midpoint of the late bedtime nights. Sleep/dark at this time should not alter circadian rhythms as neither afternoon sleep/dark episodes [3] nor afternoon bright light episodes [6] phase shift circadian rhythms.
All subjects slept at home and did not have a bed partner. This study was run in Spring and Fall. Sunrise varied between 04:31 and 07:13 and sunset between 16:20 and 19:03 (central standard time). If a subject’s bedroom was not sufficiently dark (i.e. <1 lux), we covered the windows with black plastic. During their scheduled sleep/dark times at home, subjects were instructed to lie in bed in the dark regardless of whether they could sleep. To ensure compliance, subjects wore an actigraphy monitor (Acti-watch-L, Mini-Mitter, Oregon, USA) on their nondominant wrist during the entire study, except during showers and baths. Subjects called the lab voice mail (the time and date of the call were recorded) before turning out their lights at night and just after they turned on their lights each morning. They also completed a sleep log every day, noting lights off time before sleep, any awakenings during the night, and lights on time in the morning.
To monitor light exposure, all subjects wore a photosensor around their neck as a medallion (Actiwatch-L, Mini-Mitter, Oregon, USA), except when they were in bed, or during showers and baths. During sleep periods the light medallion was placed on their nightstand. The subjects were required to receive a minimum of 5 min of outdoor light between 07:00 and 08:00 every morning, in order to increase the contrast between the nighttime dark and daytime light periods. Subjects were asked not to wear sunglasses during this time, but could at any other time if they so wished.
To ensure compliance with the study protocol, subjects came to the lab every 1–3 days so that the data from the wrist actigraph, photosensor, and sleep logs could be examined in their presence. Subjects could consume up to 300 mg of caffeine before 10:00 but could not consume caffeine at any other time. Two standard drinks of alcohol per day were allowed except on the last 2 days of each condition. Subjects chose their own meal times. Non-steroidal anti-inflammatory drugs were not permitted, as they suppress melatonin [12].
On the final day in each condition, each subject participated in a 17–20 h long phase assessment which began between 14:00 and 18:00. During the phase assessments, subjects remained awake and semirecumbent in dim light (<10 lux, at the level of the subjects’ eyes, in the direction of gaze). Subjects gave a 2 ml saliva sample every 30 min using Salivettes (Sarstedt, North Carolina, USA). The samples were later radioimmunoassayed for melatonin by Pharmasan Labs (Wisconsin, USA). The sensitivity of the assay was 0.7 pg/ml, and intra- and interassay coefficient of variabilities were 12.1% and 13.2%, respectively.
Two phase markers were derived from each melatonin profile, the dim light melatonin onset (DLMO) and dim light melatonin offset (DLMOff). For each subject’s melatonin profile, a threshold was calculated as twice the mean of the first three low daytime values. The highest of the two thresholds was used for each subject. Each subject’s DLMO was the point in time (as determined with linear interpolation) when the melatonin concentration exceeded and remained above the threshold for at least 2 h. The DLMOff was the point in time when melatonin levels fell below the threshold for at least 2 h. The thresholds ranged from 1.3 to 4.8 pg/ml (mean ± SD 2.6 ± 1.1 pg/ml). We also calculated the area under the curve (AUC) [16] of each melatonin profile.
Fig. 1 and Table 1 show a phase delay in the melatonin rhythm following the late bedtime nights, as compared with after the early bedtime nights. The DLMO and DLMOff occurred significantly later in time (paired t-test, t[9] = 5.60, P < 0.001; t[9] = 2.69, P = 0.025, respectively). On average the DLMO occurred 0.6 ± 0.3 h later (shift ranged from an advance of 0.2 h to a delay of 1.0 h) and a delay occurred in nine of the ten subjects. On average the DLMOff occurred 0.5 ± 0.6 h later (shift ranged from an advance of 0.2 h to a delay of 1.4 h) and a delay occurred in seven of the ten subjects. The AUC and DLMO to DLMOff duration were not significantly different between conditions (Table 1, P > 0.05). Thus, a week of the short and long nights was not sufficient to produce a change in duration of the dim light melatonin rhythm, whereas changes in duration have been observed following 4 weeks of long nights [17], and in habitual short versus long sleepers [1].
Fig. 1.

Mean dim light salivary melatonin profiles following a series of nights with the early bedtime (22:00, light line) and late bedtime (01:00, dark line). Error bars represent SEs. The horizontal line indicates the average threshold (2.6 pg/ml). The mean melatonin profiles were constructed by referencing each individual subject’s data to the time of their DLMO following the early bedtime nights.
Table 1.
Parameters of the dim light melatonin rhythm after the late bedtime and early bedtime nights
| Early bedtime
|
Late bedtime
|
|||
|---|---|---|---|---|
| Mean | SD (h) | Mean | SD (h) | |
| DLMO* | 20:08 | 1.1 | 20:46 | 1.3 |
| DLMOff* | 06:49 | 1.1 | 07:18 | 1.4 |
| DLMO–DLMOff duration | 10.7 | 0.9 | 10.5 | 1.3 |
| AUC | 251.2 | 144.4 | 235.1 | 142.9 |
Significant change between the early bedtime and late bedtime nights (P < 0.05).
Although our sample size was small, we did examine the data for trends due to various factors. There was no significant sex difference in the shift of the DLMO or DLMOff (both P > 0.10). No significant correlations existed between the shift in the DLMO and the number of nights in each condition, nor between the shift in the DLMOff and the number of nights (both P > 0.10). This suggests that most of the shift in the DLMO and DLMOff occurred in the first week of each condition. Similarly, there was no significant correlation between morningness–eveningness and the shift in the DLMO and DLMOff (both P > 0.10), suggesting that the results are not affected by circadian type. There was also no difference in the shift of the DLMO and DLMOff between subjects who participated in the late bedtime nights first versus early bedtime nights first (both P > 0.10). There was no difference between subjects who participated in the Fall (n = 6) and those who participated in Spring (n = 4) (both P > 0.10). Finally, there was no significant difference in the shift of the DLMO nor DLMOff between those subjects who wore sunglasses during the study (n = 2) and those that did not (n = 8, P > 0.10).
Our results indicate that despite a fixed lights on time and morning outdoor light exposure a later bedtime will significantly delay, or an earlier bedtime will significantly advance, the circadian clock. Advances in the DLMO with early bedtimes have been reported previously, but these were extremely early bedtimes that people would rarely freely adopt (16:00 [15], 18:00 [17]).
The melatonin rhythm may have been delayed after the late bedtime nights because of the indoor light that subjects received between 22:00 and 01:00 every night. The melatonin rhythm may have been advanced after the early bedtime nights because of the dark that subjects were in between 22:00 and 01:00 every night. Analysis of the light data from the medallion photosensor, corrected for sunglasses use, supports this (Fig. 2). The series of late bedtime nights was associated with significantly more dim evening light exposure (~40 lux), while morning light exposure between 07:00 and 09:00 was not significantly different between conditions. While previous work has found that light of typical indoor intensity (~180 lux) can phase shift the circadian system, in that case the room light was surrounded by very dim light (<15 lux) and was thus the most intense light experienced each day [2]. The results from our study suggest that even when humans experience a typical 24 h light/dark cycle, including bright outdoor morning light, the human circadian system may still remain sensitive to dim indoor evening room light.
Fig. 2.
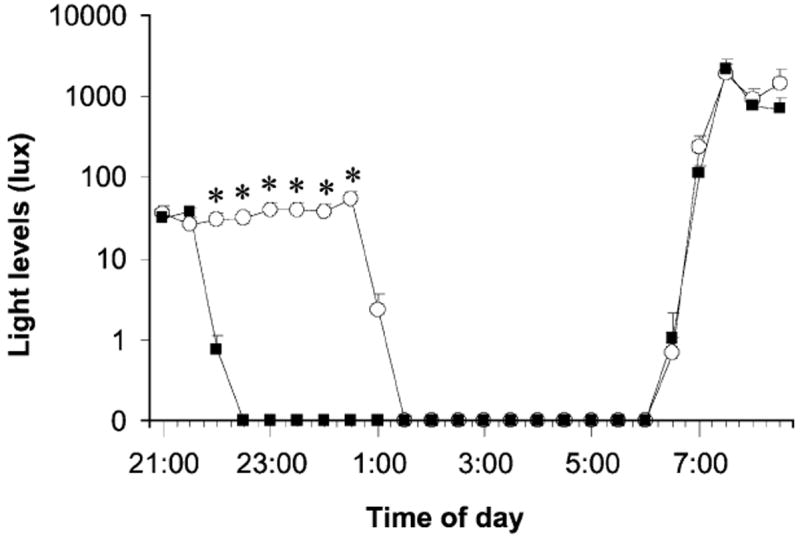
Mean light levels from 21:00 to 09:00 during the series of nights with the early bedtime (22:00, filled squares) and late bedtime (01:00, open circles), averaged into 30 min bins. Data are corrected for sunglasses use. Error bars represent SEs. Asterisks indicate times when the light intensity was significantly different between the early and late bedtime nights (corrected paired t-tests, all P < 0.005).
It is also possible that changes in nonphotic zeitgebers between conditions, such as activity, posture, food intake and the timing of sleep per se, may have led to the phase change in the melatonin rhythm. However, as such nonphotic zeitgebers are typically much less powerful than light (e.g. Refs. [5,7]), we believe that the difference between the dim evening light and the dark is the most likely cause of the phase shift. Another explanation of our results may be due to the subjects’ cumulative sleep debt that certainly developed during the late bedtime nights (as no subject napped every day). Recent animal work has suggested that sleep deprivation may reduce the phase shifting effects of bright light [10]. If the same is true of humans, then morning bright light may have had less of a phase advancing effect following the late bedtime nights than following the early bedtime nights. If so, this could explain why the melatonin rhythm was delayed after the late bedtime nights.
In conclusion, regardless of the mechanism our results suggest that when people shorten their sleep by going to bed later they significantly phase delay their circadian rhythms, or that when they go to bed earlier they phase advance their circadian rhythms. These effects occur even with a fixed lights on (wake) time and subsequent outdoor morning light. Future studies are needed to determine whether short nights delay circadian rhythms when compared to normal bedtimes, and if more morning light would reduce the phase change observed between the early and late nights.
Acknowledgments
This work was supported by a NIH grant to C. Eastman (NINDS R01 NS35695). We thank Stephanie Crowley, Cliff Gazda, Clara Lee, Barbara Trzop, and Christine Tseng for their assistance with data collection. We thank Cindy Hiltz for her assistance with the light analysis.
References
- 1.Aeschbach D, Sher L, Postolache TT, Matthews JR, Jackson MA, Wehr TA. A longer biological night in long sleepers than in short sleepers. J Clin Endocrinol Metab. 2003;88:26–30. doi: 10.1210/jc.2002-020827. [DOI] [PubMed] [Google Scholar]
- 2.Boivin DB, Duffy JF, Kronauer RE, Czeisler CA. Dose-response relationships for resetting of human circadian clock by light. Nature. 1996;379:540–542. doi: 10.1038/379540a0. [DOI] [PubMed] [Google Scholar]
- 3.Buxton OM, L’Hermite-Baleriaux M, Turek FW, Van Cauter E. Daytime naps in darkness phase shift the human circadian rhythms of melatonin and thyrotropin secretion. Am J Physiol. 2000;278:R373–R382. doi: 10.1152/ajpregu.2000.278.2.R373. [DOI] [PubMed] [Google Scholar]
- 4.Campbell SS, Zulley J. Ultradian components of human sleep/wake patterns during disentrainment. Exp Brain Res. 1985;12:234–255. [Google Scholar]
- 5.Danilenko KV, Cajochen C, Wirz-Justice A. Is sleep per se a zeitgeber in humans? J Biol Rhythms. 2003;18:170–178. doi: 10.1177/0748730403251732. [DOI] [PubMed] [Google Scholar]
- 6.Dumont M, Carrier J. Daytime sleep propensity after moderate circadian phase shifts induced with bright light exposure. Sleep. 1997;20:11–17. doi: 10.1093/sleep/20.1.11. [DOI] [PubMed] [Google Scholar]
- 7.Eastman CI, Hoese EK, Youngstedt SD, Liu L. Phase-shifting human circadian rhythms with exercise during the night shift. Physiol Behav. 1995;58:1287–1291. doi: 10.1016/0031-9384(95)02031-4. [DOI] [PubMed] [Google Scholar]
- 8.Horne JA, Ostberg O. Self-assessment questionnaire to determine morningness-eveningness in human circadian rhythms. Int J Chronobiol. 1976;4:97–110. [PubMed] [Google Scholar]
- 9.Lewy AJ, Wehr TA, Goodwin FK, Newsome DA, Markey SP. Light suppresses melatonin secretion in humans. Science. 1980;210:1267–1269. doi: 10.1126/science.7434030. [DOI] [PubMed] [Google Scholar]
- 10.Mistlberger RE, Landry GL, Marchant EG. Sleep deprivation can attenuate light-induced phase shifts of circadian rhythms in hamsters. Neurosci Lett. 1997;238:5–8. doi: 10.1016/s0304-3940(97)00815-x. [DOI] [PubMed] [Google Scholar]
- 11.Moore RY. The innervation of the mammalian pineal gland. Prog Reprod Biol. 1978;4:1–29. [Google Scholar]
- 12.Murphy PJ, Myers BL, Badia P. Nonsteroidal anti-inflammatory drugs alter body temperature and suppress melatonin in humans. Physiol Behav. 1996;59:133–139. doi: 10.1016/0031-9384(95)02036-5. [DOI] [PubMed] [Google Scholar]
- 13.National Sleep Foundation, Less fun, less sleep, more work: an American portrait. A National Sleep Foundation Poll. 2001 http://www.sleepfoundation.org/publications/2001poll.html.
- 14.Parry BL, Berga SL, Mostofi N, Klauber MR, Resnick A. Plasma melatonin circadian rhythms during the menstrual cycle and after light therapy in premenstrual dysphoric disorder and normal control subjects. J Biol Rhythms. 1997;12:47–64. doi: 10.1177/074873049701200107. [DOI] [PubMed] [Google Scholar]
- 15.Rajaratnam SMW, Dijk DJ, Middleton B, Stone BM, Arendt J. Melatonin phase-shifts human circadian rhythms with no evidence of changes in the duration of endogenous melatonin secretion or the 24-hour production of reproductive hormones. J Clin Endocrinol Metab. 2003;88:4303–4309. doi: 10.1210/jc.2003-030460. [DOI] [PubMed] [Google Scholar]
- 16.Salas SL, Hille E. Calculus: One and Several Variables with Analytic Geometry. Wiley; New York: 1982. pp. 366–372. [Google Scholar]
- 17.Wehr TA, Moul DE, Barbato G, Giesen HA, Seidel JA, Barker C, Bender C. Conservation of photoperiod-responsive mechanisms in humans. Am J Physiol. 1993;265:R846–R857. doi: 10.1152/ajpregu.1993.265.4.R846. [DOI] [PubMed] [Google Scholar]
- 18.Wever RA. Properties of human sleep-wake cycles: parameters of internally synchronized free-running rhythms. Sleep. 1984;7:27–51. doi: 10.1093/sleep/7.1.27. [DOI] [PubMed] [Google Scholar]