

Abstract
Protein arginine methyltransferases (PRMTs) are a family of enzymes that can methylate arginine residues on histones and other proteins. PRMTs play a crucial role in influencing various cellular functions, including cellular development and tumorigenesis. Arginine methylation by PRMTs is found on both nuclear and cytoplasmic proteins. Recently, there is increasing evidence regarding post-translational modifications of non-histone proteins by PRMTs, illustrating the previously unknown importance of PRMTs in the regulation of various cellular functions by post-translational modifications. In this review, we present the recent developments in the regulation of non-histone proteins by PRMTs.
Keywords: arginine, protein arginine methyltransferases, post-translational modification
The Protein Arginine Methyltransferase Family
Ten mammalian PRMTs have been identified in humans so far.1 The PRMTs vary in length, but all contain a conserved core region that includes a methyltransferase (MTase) domain, a β-barrel, and a dimerization arm. PRMTs are AdoMet-dependent methyltransferases, which means they catalyze highly specific methyl group transfers from the ubiquitous cofactor S-adenosyl-l-methionine (AdoMet) to a multitude of biological targets in the cell. Besides PRMTs, there are many other types of AdoMet-dependent methyltransferases as well. While the MTase binding domain is highly conserved in other AdoMet-dependent methyltransferases, the 7 β-barrel domains are quite unique to the PRMT family.2 As shown in Figure 1, PRMTs have 1–2 highly conserved MTase domains, which include motif I (VLD/EVGXGXG), post I (V/IXG/AXD/E), motif II (F/I/VDI/L/K), motif III (LR/KXXG), and THW loop.3

Figure 1. The structure of PRMTs (adapted from ref. 1). All PRMTs universally have 1–2 methyltransferase (MTase) domains. Some have other particular domains. For example, PRMT2 has a SRC homology 3 (SH3) domain. PRMT3 and 9 have zinc finger domains. In addition, PRMT9 also has an F-box domain. PRMT10 has a tetratricopeptide repeat 2 (TPR2) domain. PRMT8 has a unique N-terminal myristoylation (myr) motif.
Although PRMT family members share many common features, they also have their own unique features and different distribution patterns. As summarized in Table 1, PRMT1 is the predominant PRMT in mammalian cells, and performs over 80% of PRMT activity in cell. It was the first human PRMT isolated, cloned, purified, and characterized regarding its substrates and its pattern of methylation,4 and it is found throughout the cell. The sequence is highly conserved in all eukaryotes. PRMT1 is the smallest in the PRMT family, containing 361 amino acids. PRMT1 in humans has at least 3 major transcript variants.4 The only differences in the polypeptide sequences of these isoforms are alternative translation initiation sites at the N termini. It has been suggested that there may be differences in substrate preference between the 3 isoforms.4 PRMT2 is primarily situated within the nucleus (Table 1) and contains an N-terminal SH3 domain. It contains 433 amino acids. The putative AdoMet binding site cap is located about 104 amino acids upstream of the YFGSY sequence.4 PRMT3 predominantly exists in the cytoplasm and nucleus (Table 1) and contains a C2H2 zinc-finger domain at its N terminus.5 PRMT3 has 531 amino acids. Its N-terminal portion, between residues 230 and 367, has the most highly conserved sequences; the C-terminal half is less conserved, except residues 466 and 479.6 Co-activator-associated methyltransferase CARM1/PRMT4 mainly exists in the nucleus as a transcriptional co-activator.7 The core domain starts from the conserved Y149FQFY153 motif.4 It contains 608 amino acids. PRMT5 was originally named Janus kinase-binding protein 1 (JBP1), located in both the nucleus and cytoplasm,8 but predominately in the cytoplasm. In humans, PRMT5 has 637 amino acids. The sequences near the beginning and end of the conserved domain are SYLQY and FSWFP, respectively.4 PRMT6 has a very similar structure to PRMT1. It is the second smallest member in PRMTs family, containing 375 amino acids. PRMT6 is exclusively located in the nucleus.9,10 Its well-conserved sequence includes the YYECY motif.4 Another member of PRMTs family is PRMT7, a 692-amino acids-long protein. PRMT7 is very unique, because it contains 2 MTase domains that are located at the N and C termini: the N-terminal MTase domain is highly conserved.11,12 PRMT7 is found both in cytoplasm and nucleus. PRMT8 is a plasma membrane-associated protein mainly expressed in brain tissue.13 The structure is extremely similar to PRMT1 (over 80%), but has a unique N-terminal myristoylation motif that helps PRMT8 localize on the plasma membrane. This unique N-terminal domain of PRMT8 makes its methylation ability 10-fold lower than PRMT1, suggesting that the unique N-terminal domain of PRMT8 can inhibit its catalytic activity. PRMT8 automethylates at R58 and R73 in the N-terminal region.14 So far, only PRMT6 (at R35 site) and PRMT8 are known to have automethylation capability.15 PRMT8, the third smallest member in the PRMT family, has 394 amino acids. PRMT9, also called F-box only protein 11 (FBXO11), has been found throughout the cell, including the nucleus and the cytoplasm.16 It has a unique F-box domain on the N terminus and zinc finger domain on the C terminus. It is the largest in the PRMT family, containing 845 amino acids. The last PRMT family member currently known is PRMT10. It is a putative protein that also has 2 MTase motifs and a TPR2 (tetratricopeptide repeats) motif on its N terminus. It is located exclusively in the cytoplasm and has 843 amino acids, making it the second largest protein among PRMT family members.
Table 1. Distribution of PRMTs.
| Name | Primary localization | Alternate localization | Tissue | Enzyme classification |
|---|---|---|---|---|
| PRMT1 | Nucleus | Cytoplasm | Adrenal, Brain, Cerebellum, Ciliary body, Colon, Esophagus, Frontal cortex, Frontal lobe, Heart, Immature Dendritic cell, Kidney, Liver, Lung, Mature Dendritic cell, Ovary, Pancreas, Placenta, Prostate, Retina, Stomach, Testis, Thyroid, Urinary bladder | Type I |
| PRMT2 | Nucleus | Serum, Testis | Type I | |
| PRMT3 | Cytoplasm | Nucleus, Ribosome | Adrenal, Heart, Liver, Ovary | Type I |
| PRMT4 | Nucleus | Adrenal, Cerebellum, Ciliary body, Colon, Esophagus, Frontal cortex, Frontal lobe, Gall bladder, Kidney, Liver, Lung, Ovary, Pancreas, Placenta, Prostate, Stomach, Testis, Thyroid, Urinary bladder | Type I | |
| PRMT5 | Cytoplasm | Nucleus | Adrenal, Bile, Brain, Cerebellum, Ciliary body, Colon, Esophagus, Frontal cortex, Frontal lobe, Gall bladder, Heart, Immature Dendritic cell, Kidney, Liver, Lung, Optic nerve, Ovary, Pancreas, Prostate, Retina, Stomach, Testis, Urinary bladder, Urine | Type II |
| PRMT6 | Nucleus | Adrenal, Liver, Ovary | Type I | |
| PRMT7 | Cytoplasm | Nucleus | Brain, Lung, Testis | Type II and III |
| PRMT8 | Plasma Membrane | Brain | Type I | |
| PRMT9 | Nucleus | Cytoplasm | Liver | Type II |
| PRMT10 | Cytoplasm | Esophagus, Ovary, Stomach | N/A |
Besides the classification above, PRMTs can be further classified as type I–IV according to the methylarginine products. As shown in Figure 2, type I enzymes catalyze the formation of ω- NG- monomethylarginine (ωMMA) and asymmetric ω-NG, NG- dimethylarginines (ω-aDMA); Type-II enzymes catalyze the formation of ωMMA and symmetric ω-NG, NG- dimethylarginines (ω-sDMA); type-III enzymes catalyze the formation of ωMMA only,3 and type IV enzymes uniquely catalyze the formation of δ-NG-MMA.17 Type I–III exist in mammalian cells; type IV has only been described in yeast and possibly in plants due to its high sequence similarities.17 PRMT1, 2, 3, 4, 6, and 8 display type-I activity;18 PRMT5, 7 and 9 display type-II activity; in addition to type-II activity, PRMT7 also displays type-III activity. The activity of PRMT10 has not yet been characterized.3
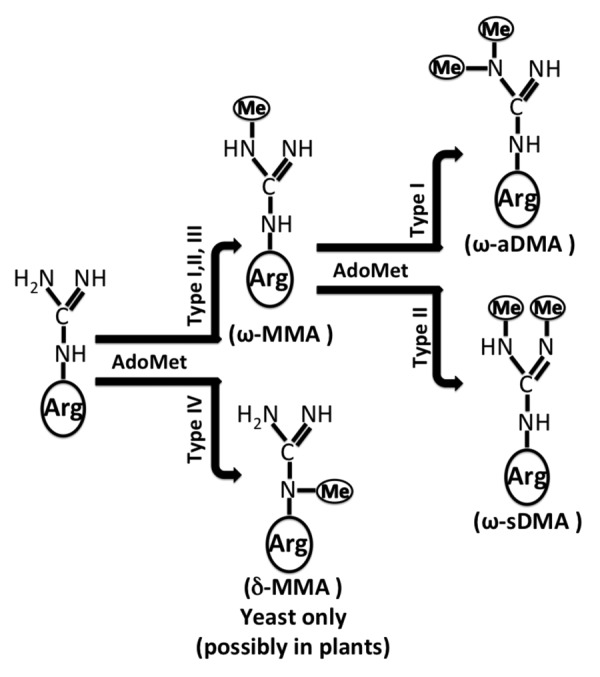
Figure 2. Methylation of arginine residues (adapted from ref. 17). Three types of methylated arginine residue products have been proven in mammalian cells. They are ωMMA, ω-aDMA, and ω-sDMA. ωMMA refers to a single methyl group placed on the terminal nitrogen atom. Its production is catalyzed by type I, II, or III PRMTs. ω-aDMA refers to 2 methyl groups placed on the same terminal nitrogen atom. It is catalyzed by type I PRMTs. ω-sDMA refers to 2 methyl groups placed on each of the terminal nitrogens. It is catalyzed by type II PRMTs. δ-MMA so far only has been described in yeast (possibly in plants), and refers to a single methyl group placed on the internal guanidino nitrogen atom. Abbreviations: ω-aDMA, asymmetric ω- NG, NG- dimethylarginines; ωMMA, ω - NG- monomethylarginine; ω-sDMA, symmetric ω - NG, NG- dimethylarginines; δ-MMA, δ-monomethylarginine
Methylation of Histone Proteins by PRMTs
PRMTs were initially found to directly methylate various histone proteins (Table 2) to regulate different cellular functions. For example, post-translational modification of histones is known to occur on the tail region and contributes to the histone code. PRMT1 and 2 are known to methylate histone H4R3;19,20 histone H3 is methylated by PRMT4 on R2, R17, and R26.21 PRMT5 methylates histone H2AR3, H3R8, and H4R3, negatively regulating the expression of tumor-suppressor genes and thus acting as an oncogene. PRMT5 protein levels are increased in leukemia and lymphoma cells.22 The overexpressed PRMT5 elevates symmetric methylation of H3R8 and H4R3, located in the promoter regions of target genes (such as Retinoblastoma protein [RB1] [p105], Retinoblastoma-like 1 [RBL1] [p107], and Retinoblastoma-like 2 [RBL2] [p130]), in turn silencing the RB family of tumor suppressor genes.23 In addition, PRMT5, in association with methylosome protein 50 (Mep50), methylates cytosolic histone H2A (H2AR3me2) to repress differentiation genes in ES cells.24 Most recently, H3R2 was shown to be symmetrically dimethylated by PRMT5. H3R2me2 binds to WD repeat domain 5 (WDR5) and helps to recruit WDR5 to euchromatin for transcriptional activation in P493–6 human B cells and human promyelocytic leukemia cell line HL60.25 Symmetric dimethylation of H3R2 is a newly identified histone mark that supports euchromatin maintenance. Additionally, PRMT5 negatively regulates some tumor suppressors, including ST7 and NM23, whose decreased expression correlates with H3R8 methylation in vitro.22 Moreover, PRMT5 protein levels are high in transformed chronic lymphocytic leukemia (B-CLL) cell lines.
Table 2. Specific examples of arginine methylations on histone and non-histone proteins.
| Family member | Known histone substrate | Known non-histone protein substrates | Year published |
|---|---|---|---|
| PRMT1 | H4R319 | hnRNP A1 (R194)81 | 1994 |
| BTG1 (Non specific sites)82 | 1996 | ||
| TIS2 (Non specific sites)82 | 1996 | ||
| IFNα/β(interferon-α receptors (IFNAR1)- intracytoplasmic domain (IC) domain)83 | 1997 | ||
| ILF3 (COOH-terminal region)84 | 2000 | ||
| SPT5 (R681, 696, and 698)34 | 2003 | ||
| Scaffold attachment factor A (SAF-A) (from amino acid 778 to 793)85 | 2004 | ||
| 53BP1(GAR motif between amino acids 1319–1480)39 | 2005 | ||
| MRE11 (GAR domain)37 | 2005 | ||
| FMRP (R544 and 546)86 | 2006 | ||
| Sam68 (proline motif P3, R280–339)87 | 2007 | ||
| SLM (GAR)87 | 2007 | ||
| ERα (R260)44 | 2008 | ||
| RUNX1 (R206 and R210)88 | 2008 | ||
| TAF15 (Arg-Gly-Gly (RGG) repeats)89 | 2009 | ||
| BCR (R198)40 | 2010 | ||
| CF Im59 (C termini) and CF Im68 (C termini and GAR motif)72 | 2010 | ||
| Ash2L (R296)64 | 2011 | ||
| PRMT2 | H4R319,20 | ERα (AF-1, DNA binding domain and hormone binding domain)90 | 2002 |
| Glutathione transferase (GAR motif)91 | 2009 | ||
| PRMT3 | N/A | FMRP (R544 and R546)86 | 2006 |
| rpS2 (GAR motif)92 | 2007 | ||
| PRMT4/CARM1 | H3R221 H3R1721 H3R2621 |
PABP1 (R455 and 460)93 | 2002 |
| (CBP)/p300 (R2142)94 | 2005 | ||
| FMRP (R544 and R546)86 | 2006 | ||
| Sox9 (high-mobility group (HMG) domain)95 | 2009 | ||
| CA150 (proline-, glycine-, methionine-rich (PGM) motif)96 | 2009 | ||
| SmB (PGM motif)96 | 2009 | ||
| U1C (PGM motif)96 | 2009 | ||
| SF3b4 (PGM motif)96 | 2009 | ||
| PRMT5 | H2AR322 | MBP (R107)97 | 1971 |
| H3R225 | LSm4 (No specific sites)98 | 2000 | |
| H3R822 | Sm D1 (GAR motif)98 | 2000 | |
| H4R322 | Sm D3 (GAR motif)98 | 2000 | |
| EBNA-2 (R325–376)99 | 2003 | ||
| SPT5 (R698)34 | 2003 | ||
| EBNA-1 (R325–376)69 | 2006 | ||
| RBL2 (H3R8 and H4R3)23 | 2008 | ||
| p53 (R333, 335, 337)40 | 2008 | ||
| CBP-1 (R234)41 | 2009 | ||
| CF Im68 (GAR motif)72 | 2010 | ||
| Ash2L (R296)64 | 2011 | ||
| PDCD4(R110)61 | 2011 | ||
| HoxA (R140)71 | 2012 | ||
| NFkB (R30)49 | 2013 | ||
| PRMT6 | H2AR299 | GAR motif100 | 2002 |
| H3R210 | PRMT6100 | 2002 | |
| HIV Tat (from amino acid 49 to 63)101 | 2005 | ||
| HMGA1a (R57 and 59)102 | 2006 | ||
| DNA polymerase β (R83 and 152)42 | 2006 | ||
| PRMT6 (R35)15 | 2013 | ||
| PRMT7 | H2AR3103 H3R225 |
GAR motif11 | 2004 |
| H4R3103 | |||
| PRMT8 | H2A (No specific sites)14 H4 (No specific sites)14 |
GAR motif14 | 2002 |
| Myelin basic protein (MBP) (No specific sites)14 | 2002 | ||
| PRMT8 (R58and 73)14 | 2007 | ||
| Ewing sarcoma (EWS) (arginine-glycine-glycine repeats (RGG))104 | 2008 | ||
| PRMT9/FBXO11 | N/A | P-SmD1 (No specific sites)105 | 2006 |
| PRMT10 | N/A | N/A |
Note: Exact R residues are noted in Table 2 only if it was clearly stated in the original report.
PRMT6 has been shown to methylate histones H2AR29 and H3R2, decreasing transcription of homeobox (Hox) genes and myelocytomatosis oncogene-dependent genes.9,26 Additionally, knockdown of PRMT6 leads to delay of cell cycle progression and defect of G2 checkpoint in human osteosarcoma cell line U2OS. Overexpression of PRMT6 leads to increased H3R2 dimethylation at the transcriptional start sites of p21 and p27 promoters and decreased expression of p21 and p27.27
PRMT7 plays a role in male germ-line imprinted gene methylation through interaction with testis-specific factor CCCTC-binding factor (zinc finger protein)-like (CTCFL) and H4R3me2s accumulation.28 Like PRMT5, PRMT7 symmetrically dimethylates H3R2. H3R2me2 enhances binding of WDR5, correlating with changes in chromatin markers and expression of genes belonging to fundamental pathways of transcription regulation and protein post-translational modification.25 PRMT7 also dimethylates H2AR3 and H4R3 in the promoter region of polymerase (DNA directed) δ1, catalytic subunit (POLD1), negatively regulating cellular response to DNA damage.29 PRMT8 is able to methylate H2A (no specific site) and H4 (no specific site) via N-terminal myristoylation.14 No histone substrates for PRMT3, 9, or 10 have been found so far.
Methylation of Non-Histone Proteins By PRMTs
Since arginine methylation of non-histone proteins is an emerging area for which few reviews are available,30,31 we will mainly focus on recent discoveries of arginine methylations of non-histone proteins and their important roles in various cellular functions.
RNA processing and transcriptional elongation
RNA binding proteins (RBPs) often contain glycine–arginine-rich (GAR) motifs and are considered major substrates for PRMTs. Methylation may either negatively or positively influence RNA–protein interactions by preventing formation of hydrogen bonds by steric hindrance and causing arginine residues to become more hydrophobic.32 It has been reported that arginine methylation by PRMT5 helps small nuclear ribonucleoprotein-associated protein B/B' (Sm protein B/B'), like Sm4 (LSm4), small ribonucleoprotein particle protein SmD1 and 3 (SmD1 and 3) assemble into mature small nuclear ribonucleoproteins (snRNPs).33 All 25 occurrences of the triplet GAR in these 4 proteins are symmetrically dimethylated, with the exception of 3 arginines in LSm4 and one in SmD3 for which no data are available. Further use of methylated peptides or arginine methylation inhibitors can block the survival motor neuron (SMN) and Sm interactions so that Sm proteins cannot translocate into the nucleus to facilitate snRNP assembly.33 Furthermore, PRMT5 methylates transcription elongation factor SPT5 at R698, which decreases SPT5 association with RNA polymerase II, suggesting that PRMT5 might be involved in regulating transcriptional elongation.34
Cell cycle and DNA repair
During mitosis, the 68-kDa Src-associated substrate in mitosis (Sam68) was bound and methylated by PRMT1 on the proline motif P3 at amino acids 280–339, which promoted the Sam68 nuclear localization and facilitated the export of unspliced HIV RNA into the cytoplasm. In PRMT1-deficient embryonic stem cells, Sam68 was localized to the cytoplasm. Additionally, deletion of the PRMT1 methylation sites and the use of methylase inhibitors resulted in Sam68 accumulation in the cytoplasm.35 PRMT1 is also required for proper cell cycle progression, such as G2/M and G1/S checkpoint activations.36 Meiotic recombination 11 (MRE11) contains a nuclease domain which is active in homologous recombination repair (HRR). In addition to the nuclease domain, MRE11 harbors a GAR motif on amino acids 554–680 that can be methylated by PRMT1. The data show cells containing hypomethylated MRE11 displayed inefficient DNA damage repair that was significantly rescued by induction of the MRE11-RAD50-NBS1 complex (MRN).37 Tumor suppressor 53-binding protein 1 (53BP1) is another critical protein involved in DNA repair due to the high affinity with both single- and double-stranded DNA.38 PRMT1 methylates the GAR motif of 53BP1 on amino acids 1319–1480 and helps 53BP1 localize to DNA damage sites in fibroblast cells.39 Transfected 53BP1 with a mutated GAR motif would no longer function as a methyl-acceptor.39 Additionally, PRMT1-deficient mouse embryonic fibroblasts (MEFs) are hypersensitive to the DNA damaging agent etoposide and fail to recruit homologous recombination RAD51 recombinase to DNA damage foci in vitro.36
PRMT5 also plays important roles in cell cycle and DNA repair. PRMT5 knockout triggers cell cycle arrest in G1 by inducing p53 expression upon recognition of DNA damage, eventually leading to p53-dependent apoptosis.40 PRMT5 has been shown to affect DNA repair by methylating p53 at selected arginine residues R333, R335, and R337 in the p53 oligomerization domain, increasing the response to DNA repair.40 A study demonstrated that in C. elegans PRMT5 downregulated DNA damage induced apoptosis by methylating CBP-1 at R234.41 When DNA damage-induced apoptosis is negatively regulated, the risk of genomic instability and tumor progression is augmented.
In addition to PRMT1 and 5, PRMT6 has also been shown to methylate Polβ on R83 and R152, thus enhancing DNA binding ability and stimulating DNA polymerase activity, eventually triggering base excision repair.42
Regulation of signaling downstream of receptors
Recent studies showed that PRMTs could mediate signal transduction downstream of the B-cell antigen receptor (BCR)43 and estrogen receptor α (ERα).44 PRMT1 methylated the Igα subunit of the BCR at R198, negatively regulating the phosphoinositide-3 kinase (PI3K) pathways and promoting B cell differentiation.43 Moreover, PRMT1 can methylate ERα at R260 in its DNA-binding domain. This is required for mediating the extranuclear function of the receptor by triggering its interaction with the p85 subunit of PI3K and Rous sarcoma oncogene (Src) and recruitment of focal adhesion kinase (FAK).44
Regulation of transcription factors (e.g., nuclear factor κB)
PRMTs can also regulate the function of transcription factors. As we mentioned above, p53 is regulated by PRMT5 through direct arginine methylation.40 There are several studies showing that PRMT1, 2, and 4 can regulate the activity of the ubiquitous inducible transcription factor, nuclear factor κB (NFκB). NFκB plays an important role in the transcriptional regulation of genes involved in inflammation, cell survival, and tumorigenesis. Hottiger’s group showed that PRMT4 is a novel transcriptional coactivator of NFκB and functions as a promoter-specific regulator of NFκB recruitment to chromatin. PRMT4 acts in a gene-specific manner, mainly by enhancing NFκB recruitment to cognate sites. Moreover, PRMT4 synergistically coactivates NFκB-mediated transactivation in concert with the transcriptional coactivators, p300/CREB-binding protein, and the p160 family of steroid receptor coactivators. These results suggest that the cooperative action between protein arginine methyltransferase PRMT4 and protein lysine acetyltransferases p300 regulates NFκB-dependent gene activation.45
In another report, the same research group further demonstrated that PRMT1 synergistically coactivates NFκB-dependent gene expression at the promoters of macrophage inflammatory protein 2 and human immunodeficiency virus 1 long-terminal repeat in concert with the transcriptional coactivators p300/CREB binding protein, PRMT4, and poly (ADP-ribose) polymerase 1(PARP1).46,47
Ganesh et al. reported that PRMT2 inhibits NFκB-dependent transcription and promotes apoptosis. PRMT2 exerted this effect by blocking nuclear export of the inhibitor of NFκB (IκBα) through a leptomycin-sensitive pathway, increasing nuclear IκBα and decreasing NFκB DNA binding mediated by the highly conserved S-adenosylmethionine-binding domain of PRMT2.48
Although the above reports suggest the correlation between NFκB and PRMT1, 2, and 4, none of the reports had shown the regulation of NFκB is through direct arginine methylation. Recently, our lab discovered that PRMT5 dimethylates the subunit p65 at R30 to activate NFκB.49 Complex post-translational modifications of the p65 subunit of NFκB are a major aspect of the extremely flexible regulation of NFκB activity. Although phosphorylation, acetylation, ubiquitilation, and lysine methylation of NFκB have been well described,46,50-55 direct arginine methylation on NFκB has not previously been found. We discovered that expression of the R30A mutant of p65 substantially decreased the ability of NFκB to bind to κB elements and to drive gene expression. A model in which dimethyl R30 is placed into the crystal structure of p65 predicts new Van der Waals contacts that stabilize intraprotein interactions and indirectly increase the affinity of p65 for DNA.49 PRMT5 was the only arginine methyltransferase that co-precipitated with p65, and its overexpression increased NFκB activity, while PRMT5 knockdown had the opposite effect. Microarray analysis revealed that most of the NFκB-inducible genes downregulated by the R30A mutation are similarly downregulated by knocking down PRMT5. Many cytokine and chemokine genes are among these, and conditioned media from cells expressing the R30A mutant of p65 had much less NFκB-inducing activity than media from cells expressing the wild-type protein. A model in Figure 3 summarizes this novel finding. Furthermore, PRMT5 is frequently found overexpressed in many types of cancer, often to a striking degree, indicating that high levels of this enzyme may promote tumorigenesis, at least in part by facilitating NFκB-induced gene expression.
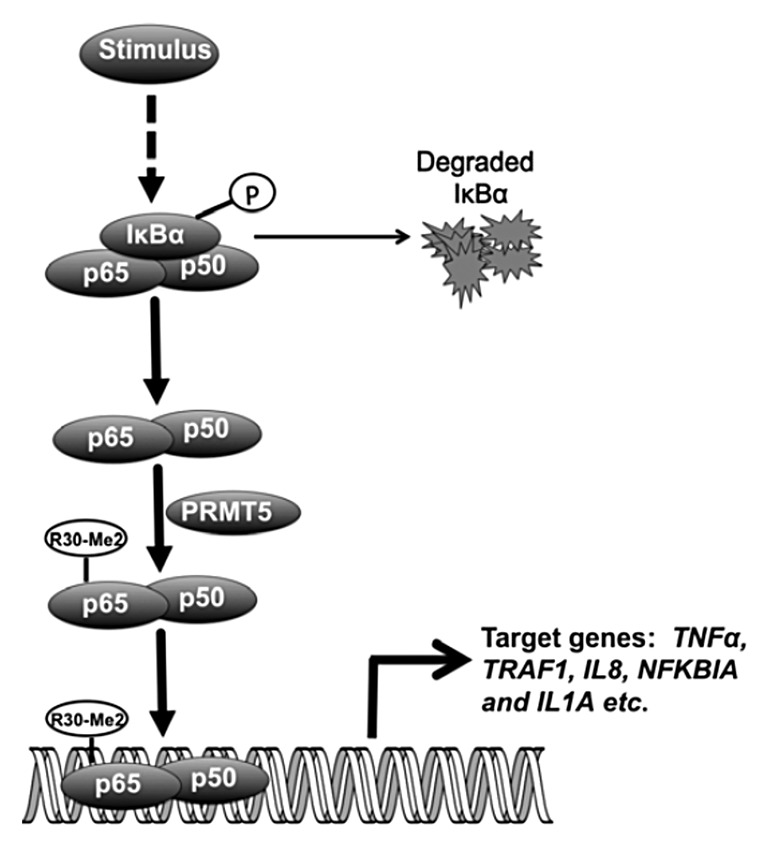
Figure 3. A model of regulation of NFκB by PRMT5 (adopted from ref. 49). In addition to previously known regulatory pathways, NFκB is regulated by PRMT5-mediated methylation of p65 on R30, which affects the expression of many NFκB-induced genes.
PRMTs as Therapeutic Targets for Diseases
Cancer
PRMTs play an important role in carcinogenesis. For example, dysregulation of the PRMT1 and PRMT6 genes promote growth of bladder and lung cancer cells, and abrogation of the expression of these genes significantly suppresses growth of bladder and lung cancer cells.56 This phenomenon appears to be attributable to increased serum aDMA levels during carcinogenesis.56 Knockdown of the expression of PRMT1 and PRMT6 by siRNA transfection can suppress breast cancer cell proliferation.56 The methylation status of H4R3 that is methylated by PRMT1 is highly relevant to tumor grade and helps to predict cancer recurrence.3 Besides PRMT1 and 6, PRMT4 has also shown to be overexpressed in human grade-III breast tumors, colorectal cancer, and prostrate adenocarcinomas; knockdown of PRMT4 inhibits proliferation of breast, colorectal, and prostate cancer cell lines.57
The above examples, such as PRMT1, 4 and 6 are all from type-I PRMTs. PRMT5, a typical enzyme of type-II PRMTs, has been shown to promote cell growth and transformation by different mechanisms. Aggarwal et al. suggested that the nuclear cyclin D1/CDK4 kinase regulates cullin4 (CUL4) expression and triggers neoplastic growth via activation of PRMT5.58 Gu et al. suggested that PRMT5 is essential for growth of lung adenocarcinoma A549 cells. Knockdown of PRMT5 in A549 cells suppresses cell growth and tumor xenografts in nude mice partially through downregulation of the fibroblast growth factor receptor (FGFR) signaling pathway.59 Oncomine data also suggest that among colon cancer patients, most have elevated PRMT5 expression levels, strongly indicating that PRMT5 promotes colon tumor growth. In fact, PRMT5 is overexpressed in a variety of cancers, such as colon, ovary, kidney, lung, bladder, liver, pancreas, breast, prostate, cervix, and skin, with especially high expression in colon cancer.49 PRMT5 was also called JAK-binding protein 1, because it interacts with JAK. It has been shown that myeloproliferative leukemia neoplasm patients have the JAK2 mutant JAK2V617F, which binds to PRMT5 more strongly than wild-type JAK2, leading to the neoplastic phenotype.60 Furthermore, PRMT5 can accelerate tumor growth by methylating R110 on tumor suppressor programmed cell death 4 (PDCD4) in an orthotopic model of breast cancer,61 leading to the utility of PDCD4/PRMT5 as both a prognostic biomarker and a potential target for chemotherapy.61 It has also been shown that PRMT5 is involved in tumor formation by upregulating G1 cyclins/cyclin-dependent kinases and the PI3K/AKT signaling cascade in lung cancer cell lines A549 and H1299.62 In tumor cells, E2F-1 is methylated on R111 and R113 by PRMT5, reducing its stability. Analysis of a subgroup of colorectal cancer cells indicates that high levels of PRMT5 frequently coincide with low levels of E2F-1 and reflect a poor clinical outcome.63 In addition, PRMT5 has also been shown to methylate Ash2L (absent, small, or homeotic)-like (Drosophila) at R296, which is associated with transformation of human tumors.64
Cardiovascular and pulmonary disease
Type-I PRMTs catalyze ω-MMA and ω-aDMA synthesis. Studies showed that elevated ω-aDMA plasma levels are associated with endothelial dysfunction-related cardiovascular and pulmonary diseases, including coronary heart disease (CHD)65 and pulmonary hypertension.66 Dimethylarginine dimethylaminohydrolase-1 and -2 (DDAH) regulate endothelial nitric oxide (NO) production through interactions with ω-aDMA that may increase susceptibility to cardiovascular diseases.67
Other diseases
In addition to the diseases mentioned above, PRMTs are also found to be involved in virus-related diseases. For example, PRMT4 increases trans-activator X (Tax) transactivation of the human T-cell lymphotropic virus type 1 (HTLV-1) long-terminal repeat (LTR) through direct binding to Tax. This binding promotes methylation of histone H3 (R2, R17, and R26).68 PRMT5 is also known to methylate EBNA1, an Epstein-Barr virus protein in a stretch of multiple regions from R325 to R376, helping the localization of EBNA1 and consequently inducing the formation of EBNA1 rings around the nucleoli.69 In addition to PRMT4 and 5, PRMT6 has been shown to decrease RNA pol II transcription of viral genes by negatively regulating the trans-activator of transcription (Tat) expression. Overexpression of PRMT6 reduced Tat transactivation of HIV-1 production and viral replication.70
In addition to the above examples regarding virus-related diseases, PRMTs also have been shown to be involved in the endothelial cell (EC) inflammatory response. One report suggested that PRMT5-induced methylation of HOXA9 (Homeobox protein Hox-A9) on R140 is required for EC-leukocyte adhesion molecule (ELAM) expression during the EC inflammatory response.71
Moreover, PRMT5, together with its cofactor WD45, methylates the mammalian cleavage factor I (CF I [m]) subunit of CF I(m)68 at its GAR motif, stimulating methylosome subunit pICln (pICln, also named CLNS1A). pICln induces an outwardly rectifying, nucleotide-sensitive chloride current when expressed in Xenopus oocytes.72 pICln prevents the interaction of Sm proteins with the SMN protein, inhibiting snRNP biogenesis.73 pICln may act as a PRMT5 co-substrate when bound to Sm proteins, by stimulating Sm protein methylation at its RG repeats.73
Perspectives and Future Directions
There are several known PRMT inhibitors, such as S‐adenosyl homocysteine (AdoHcy), which compete with products of the methyltransferase reaction to bind the active site of the enzyme without specificity for the PRMT isotype.74 Recently, several small-molecule PRMT inhibitors have been synthesized. C21, a chloroacetamidine-bearing histone H4 tail analog, acts as an irreversible PRMT1 inhibitor.75 AMI-1 (protein arginine N-methyltransferases inhibitor 1) specifically inhibits arginine, but not lysine, methyltransferase activity in vitro and does not compete for the AdoMet binding site.76 Also, an inhibitor named compound 2 was reported to be able to inhibit co-activator-associated PRMT1 with an IC50 of 30 nM.77 Recently, 2 potent and selective inhibitors of PRMT4 were reported. One was methylgene compound 7a,78 the other was Bristol-Myers Squibb compound 7f.79 Both were pyrazole-based inhibitors, and functioned at nanomolar concentrations, displaying >100-fold selectivity for the PRMT4 over other related enzymes. The structure of PRMTs and existing PRMT inhibitors provide a guide for the synthesis of new inhibitors.
As we discussed in this review, PRMTs play a crucial role in various biological processes, including RNA processing, DNA repair, signal transduction, and transcriptional regulation. To fully understand these processes, several progressions are required. First, the site of histone methylation needs to be further assessed. PRMTs are extremely promising therapeutic targets for various types of cancer. Discovery of additional methylation sites may lead to the development of more specific inhibitors in the near future. Second, due to the wide range of non-histone substrates compared with histone substrates and the cellular functions mediated by these non-histone substrates, it is important to understand the interaction mechanisms between PRMTs and non-histone substrates. Third, PRMT5 is notable in that it was the first enzyme known to synthesize sDMA on arginine residues in proteins8 and is known to have important roles in transcriptional modulation, such as the dimethylation of R30 of the p65 subunit of NFκB, leading to its activation. In the future, chemical inhibition of PRMT5 methyltransferase activity could be used to affect function of PRMT5 and its factors. Recently, the crystal structure of the human PRMT5:MEP50 complex has been reported.80 The structure of the surprising hetero-octametric complex reveals the close interaction between the 7-bladed β-propeller MEP50 and the N-terminal domain of PRMT5, and it delineates the structural elements of substrate recognition. This information would shed light on the synthesis of new inhibitors of PRMT5, and facilitate the development of novel treatments for human diseases.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Ms Lisa King at the Department of Pharmacology and Toxicology at Indiana University School of Medicine for her generous and professional help with revising this review. The research is supported by grants 23-862-07TL (to TL) and 036433730102 (to TL).
Glossary
Abbreviations:
- ADMA
asymmetric dimethylarginines
- AdoHcy
S-adenosyl homocysteine
- AdoMet
S-adenosyl methionine
- AMI-1
protein arginine N-methyltransferase inhibitor 1
- AshL
absent, small, or homeotic-like (Drosophila)
- B-CLL
B-cell chronic lymphocytic leukemia
- BCR
B-cell antigen receptor
- CARM1
coactivator-associated arginine methyltransferase
- CBP
CREB binding protein
- CDK
cyclin-dependent kinase
- CFI
cleavage factor I
- CHD
coronary heart disease
- CIP
CDK interacting protein
- CREB
cAMP response element-binding protein
- CTCFL
CCCTC-binding factor (zinc finger protein)-like
- CUL
cullin
- DDAH
dimethylarginine dimethylaminohydrolase
- DNA
deoxyribonucleic acid
- EBNA
Epstein-Barr virus nuclear antigen
- EC
endothelial cell
- ELAM
EC-leukocyte adhesion molecules
- ER
estrogen receptor
- ES
embryonic stem cell
- FAK
focal adhesion kinase
- FBXO
F-box only protein
- FGFR
fibroblast growth factor receptor
- 53BP1
53-binding protein1
- G
glycine
- GAR
glycine arginine rich
- HIV
human immunodeficiency virus
- Hox
homeobox protein
- HRR
homologous recombination repair
- HTLV
human T cell lymphotropic virus
- IκBα
nuclear factor of κ light polypeptide gene enhancer in B-cells inhibitor, α
- IC50
half maximal inhibitory concentration
- JBP
Janus kinase-binding protein
- K
Lysine
- KIP
kinase inhibitory protein
- LSm
like Sm
- LTR
long terminal repeat
- MEFs
mouse embryonic fibroblast cells
- Mep
methylosome protein
- MMA
monomethylarginine
- MMP
matrix metalloproteinase
- Mre
meiotic recombination
- MRN
MRE11-RAD50-NBS1 complex
- MTase
methyltransferase
- NBS
Nijmegen breakage syndrome
- NFκB
nuclear factor-κB
- NM
nonmetastatic protein
- NO
nitric oxide
- PARP
Poly ADP-ribose polymerase
- PDCD
programmed cell death protein
- p53
tumor suppressor 53
- PI3K
phosphoinositide 3-kinase
- POLD
DNA polymerase delta catalytic subunit
- PRMTs
protein arginine methyltransferases
- R
arginine
- Rb
retinoblastoma protein
- RBL
retinoblastoma-like
- RBPs
RNA binding proteins
- RNA
ribonucleic acid
- Sam
Scr-associated substrate in mitosis
- sDMA
symmetric dimethyarginines
- Sm
small nuclear ribonucleoprotein-associated protein
- SMN
survival of motor neuron
- snRNPs
small nuclear ribonucleoproteins
- Src
Rous sarcoma oncogene
- ST
suppressor of tumorigenicity protein
- Tat
trans-activator of transcription
- Tax
trans-activator X
- TPR
tetratricopeptide repeats
- TSP
thrombospondin
- TNFα
tumor necrosis factor α
- WDR5
WD repeat domain 5
Footnotes
Previously published online: www.landesbioscience.com/journals/cc/article/27353
References
- 1.Bedford MT. Arginine methylation at a glance. J Cell Sci. 2007;120:4243–6. doi: 10.1242/jcs.019885. [DOI] [PubMed] [Google Scholar]
- 2.Cheng X, Collins RE, Zhang X. Structural and sequence motifs of protein (histone) methylation enzymes. Annu Rev Biophys Biomol Struct. 2005;34:267–94. doi: 10.1146/annurev.biophys.34.040204.144452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Yang Y, Bedford MT. Protein arginine methyltransferases and cancer. Nat Rev Cancer. 2013;13:37–50. doi: 10.1038/nrc3409. [DOI] [PubMed] [Google Scholar]
- 4.Krause CD, Yang ZH, Kim YS, Lee JH, Cook JR, Pestka S. Protein arginine methyltransferases: evolution and assessment of their pharmacological and therapeutic potential. Pharmacol Ther. 2007;113:50–87. doi: 10.1016/j.pharmthera.2006.06.007. [DOI] [PubMed] [Google Scholar]
- 5.Tang J, Gary JD, Clarke S, Herschman HR. PRMT 3, a type I protein arginine N-methyltransferase that differs from PRMT1 in its oligomerization, subcellular localization, substrate specificity, and regulation. J Biol Chem. 1998;273:16935–45. doi: 10.1074/jbc.273.27.16935. [DOI] [PubMed] [Google Scholar]
- 6.Zhang X, Zhou L, Cheng X. Crystal structure of the conserved core of protein arginine methyltransferase PRMT3. EMBO J. 2000;19:3509–19. doi: 10.1093/emboj/19.14.3509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hong H, Kao C, Jeng MH, Eble JN, Koch MO, Gardner TA, Zhang S, Li L, Pan CX, Hu Z, et al. Aberrant expression of CARM1, a transcriptional coactivator of androgen receptor, in the development of prostate carcinoma and androgen-independent status. Cancer. 2004;101:83–9. doi: 10.1002/cncr.20327. [DOI] [PubMed] [Google Scholar]
- 8.Branscombe TL, Frankel A, Lee JH, Cook JR, Yang Z, Pestka S, Clarke S. PRMT5 (Janus kinase-binding protein 1) catalyzes the formation of symmetric dimethylarginine residues in proteins. J Biol Chem. 2001;276:32971–6. doi: 10.1074/jbc.M105412200. [DOI] [PubMed] [Google Scholar]
- 9.Waldmann T, Izzo A, Kamieniarz K, Richter F, Vogler C, Sarg B, Lindner H, Young NL, Mittler G, Garcia BA, et al. Methylation of H2AR29 is a novel repressive PRMT6 target. Epigenetics Chromatin. 2011;4:11. doi: 10.1186/1756-8935-4-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Guccione E, Bassi C, Casadio F, Martinato F, Cesaroni M, Schuchlautz H, Lüscher B, Amati B. Methylation of histone H3R2 by PRMT6 and H3K4 by an MLL complex are mutually exclusive. Nature. 2007;449:933–7. doi: 10.1038/nature06166. [DOI] [PubMed] [Google Scholar]
- 11.Miranda TB, Miranda M, Frankel A, Clarke S. PRMT7 is a member of the protein arginine methyltransferase family with a distinct substrate specificity. J Biol Chem. 2004;279:22902–7. doi: 10.1074/jbc.M312904200. [DOI] [PubMed] [Google Scholar]
- 12.Lee JH, Cook JR, Yang ZH, Mirochnitchenko O, Gunderson SI, Felix AM, Herth N, Hoffmann R, Pestka S. PRMT7, a new protein arginine methyltransferase that synthesizes symmetric dimethylarginine. J Biol Chem. 2005;280:3656–64. doi: 10.1074/jbc.M405295200. [DOI] [PubMed] [Google Scholar]
- 13.Lee J, Sayegh J, Daniel J, Clarke S, Bedford MT. PRMT8, a new membrane-bound tissue-specific member of the protein arginine methyltransferase family. J Biol Chem. 2005;280:32890–6. doi: 10.1074/jbc.M506944200. [DOI] [PubMed] [Google Scholar]
- 14.Sayegh J, Webb K, Cheng D, Bedford MT, Clarke SG. Regulation of protein arginine methyltransferase 8 (PRMT8) activity by its N-terminal domain. J Biol Chem. 2007;282:36444–53. doi: 10.1074/jbc.M704650200. [DOI] [PubMed] [Google Scholar]
- 15.Singhroy DN, Mesplède T, Sabbah A, Quashie PK, Falgueyret JP, Wainberg MA. Automethylation of protein arginine methyltransferase 6 (PRMT6) regulates its stability and its anti-HIV-1 activity. Retrovirology. 2013;10:73. doi: 10.1186/1742-4690-10-73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wolf SS. The protein arginine methyltransferase family: an update about function, new perspectives and the physiological role in humans. Cell Mol Life Sci. 2009;66:2109–21. doi: 10.1007/s00018-009-0010-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Fisk JC, Read LK. Protein arginine methylation in parasitic protozoa. Eukaryot Cell. 2011;10:1013–22. doi: 10.1128/EC.05103-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gary JD, Clarke S. RNA and protein interactions modulated by protein arginine methylation. Prog Nucleic Acid Res Mol Biol. 1998;61:65–131. doi: 10.1016/s0079-6603(08)60825-9. [DOI] [PubMed] [Google Scholar]
- 19.Strahl BD, Briggs SD, Brame CJ, Caldwell JA, Koh SS, Ma H, Cook RG, Shabanowitz J, Hunt DF, Stallcup MR, et al. Methylation of histone H4 at arginine 3 occurs in vivo and is mediated by the nuclear receptor coactivator PRMT1. Curr Biol. 2001;11:996–1000. doi: 10.1016/S0960-9822(01)00294-9. [DOI] [PubMed] [Google Scholar]
- 20.Lakowski TM, Frankel A. Kinetic analysis of human protein arginine N-methyltransferase 2: formation of monomethyl- and asymmetric dimethyl-arginine residues on histone H4. Biochem J. 2009;421:253–61. doi: 10.1042/BJ20090268. [DOI] [PubMed] [Google Scholar]
- 21.Schurter BT, Koh SS, Chen D, Bunick GJ, Harp JM, Hanson BL, Henschen-Edman A, Mackay DR, Stallcup MR, Aswad DW. Methylation of histone H3 by coactivator-associated arginine methyltransferase 1. Biochemistry. 2001;40:5747–56. doi: 10.1021/bi002631b. [DOI] [PubMed] [Google Scholar]
- 22.Pal S, Vishwanath SN, Erdjument-Bromage H, Tempst P, Sif S. Human SWI/SNFassociated PRMT5 methylates histone H3 arginine 8 and negatively regulates expression of ST7 and NM23 tumor suppressor genes. Mol Cell Biol. 2004;24:9630–45. doi: 10.1128/MCB.24.21.9630-9645.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wang L, Pal S, Sif S. Protein arginine methyltransferase 5 suppresses the transcription of the RB family of tumor suppressors in leukemia and lymphoma cells. Mol Cell Biol. 2008;28:6262–77. doi: 10.1128/MCB.00923-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Tee WW, Pardo M, Theunissen TW, Yu L, Choudhary JS, Hajkova P, Surani MA. Prmt5 is essential for early mouse development and acts in the cytoplasm to maintain ES cell pluripotency. Genes Dev. 2010;24:2772–7. doi: 10.1101/gad.606110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Migliori V, Müller J, Phalke S, Low D, Bezzi M, Mok WC, Sahu SK, Gunaratne J, Capasso P, Bassi C, et al. Symmetric dimethylation of H3R2 is a newly identified histone mark that supports euchromatin maintenance. Nat Struct Mol Biol. 2012;19:136–44. doi: 10.1038/nsmb.2209. [DOI] [PubMed] [Google Scholar]
- 26.Hyllus D, Stein C, Schnabel K, Schiltz E, Imhof A, Dou Y, Hsieh J, Bauer UM. PRMT6-mediated methylation of R2 in histone H3 antagonizes H3 K4 trimethylation. Genes Dev. 2007;21:3369–80. doi: 10.1101/gad.447007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kleinschmidt MA, de Graaf P, van Teeffelen HA, Timmers HT. Cell cycle regulation by the PRMT6 arginine methyltransferase through repression of cyclin-dependent kinase inhibitors. PLoS One. 2012;7:e41446. doi: 10.1371/journal.pone.0041446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jelinic P, Stehle JC, Shaw P. The testis-specific factor CTCFL cooperates with the protein methyltransferase PRMT7 in H19 imprinting control region methylation. PLoS Biol. 2006;4:e355. doi: 10.1371/journal.pbio.0040355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Karkhanis V, Wang L, Tae S, Hu YJ, Imbalzano AN, Sif S. Protein arginine methyltransferase 7 regulates cellular response to DNA damage by methylating promoter histones H2A and H4 of the polymerase δ catalytic subunit gene, POLD1. J Biol Chem. 2012;287:29801–14. doi: 10.1074/jbc.M112.378281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bedford MT, Clarke SG. Protein arginine methylation in mammals: who, what, and why. Mol Cell. 2009;33:1–13. doi: 10.1016/j.molcel.2008.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lee YH, Stallcup MR. Minireview: protein arginine methylation of nonhistone proteins in transcriptional regulation. Mol Endocrinol. 2009;23:425–33. doi: 10.1210/me.2008-0380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pahlich S, Zakaryan RP, Gehring H. Protein arginine methylation: Cellular functions and methods of analysis. Biochim Biophys Acta. 2006;1764:1890–903. doi: 10.1016/j.bbapap.2006.08.008. [DOI] [PubMed] [Google Scholar]
- 33.Brahms H, Meheus L, de Brabandere V, Fischer U, Lührmann R. Symmetrical dimethylation of arginine residues in spliceosomal Sm protein B/B’ and the Sm-like protein LSm4, and their interaction with the SMN protein. RNA. 2001;7:1531–42. doi: 10.1017/S135583820101442X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kwak YT, Guo J, Prajapati S, Park KJ, Surabhi RM, Miller B, Gehrig P, Gaynor RB. Methylation of SPT5 regulates its interaction with RNA polymerase II and transcriptional elongation properties. Mol Cell. 2003;11:1055–66. doi: 10.1016/S1097-2765(03)00101-1. [DOI] [PubMed] [Google Scholar]
- 35.Côté J, Boisvert FM, Boulanger MC, Bedford MT, Richard S. Sam68 RNA binding protein is an in vivo substrate for protein arginine N-methyltransferase 1. Mol Biol Cell. 2003;14:274–87. doi: 10.1091/mbc.E02-08-0484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Yu Z, Chen T, Hébert J, Li E, Richard S. A mouse PRMT1 null allele defines an essential role for arginine methylation in genome maintenance and cell proliferation. Mol Cell Biol. 2009;29:2982–96. doi: 10.1128/MCB.00042-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Boisvert FM, Déry U, Masson JY, Richard S. Arginine methylation of MRE11 by PRMT1 is required for DNA damage checkpoint control. Genes Dev. 2005;19:671–6. doi: 10.1101/gad.1279805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Iwabuchi K, Basu BP, Kysela B, Kurihara T, Shibata M, Guan D, Cao Y, Hamada T, Imamura K, Jeggo PA, et al. Potential role for 53BP1 in DNA end-joining repair through direct interaction with DNA. J Biol Chem. 2003;278:36487–95. doi: 10.1074/jbc.M304066200. [DOI] [PubMed] [Google Scholar]
- 39.Boisvert FM, Rhie A, Richard S, Doherty AJ. The GAR motif of 53BP1 is arginine methylated by PRMT1 and is necessary for 53BP1 DNA binding activity. Cell Cycle. 2005;4:1834–41. doi: 10.4161/cc.4.12.2250. [DOI] [PubMed] [Google Scholar]
- 40.Jansson M, Durant ST, Cho EC, Sheahan S, Edelmann M, Kessler B, La Thangue NB. Arginine methylation regulates the p53 response. Nat Cell Biol. 2008;10:1431–9. doi: 10.1038/ncb1802. [DOI] [PubMed] [Google Scholar]
- 41.Yang M, Sun J, Sun X, Shen Q, Gao Z, Yang C. Caenorhabditis elegans protein arginine methyltransferase PRMT-5 negatively regulates DNA damage-induced apoptosis. PLoS Genet. 2009;5:e1000514. doi: 10.1371/journal.pgen.1000514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.El-Andaloussi N, Valovka T, Toueille M, Steinacher R, Focke F, Gehrig P, Covic M, Hassa PO, Schär P, Hübscher U, et al. Arginine methylation regulates DNA polymerase beta. Mol Cell. 2006;22:51–62. doi: 10.1016/j.molcel.2006.02.013. [DOI] [PubMed] [Google Scholar]
- 43.Infantino S, Benz B, Waldmann T, Jung M, Schneider R, Reth M. Arginine methylation of the B cell antigen receptor promotes differentiation. J Exp Med. 2010;207:711–9. doi: 10.1084/jem.20091303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Le Romancer M, Treilleux I, Leconte N, Robin-Lespinasse Y, Sentis S, Bouchekioua-Bouzaghou K, Goddard S, Gobert-Gosse S, Corbo L. Regulation of estrogen rapid signaling through arginine methylation by PRMT1. Mol Cell. 2008;31:212–21. doi: 10.1016/j.molcel.2008.05.025. [DOI] [PubMed] [Google Scholar]
- 45.Covic M, Hassa PO, Saccani S, Buerki C, Meier NI, Lombardi C, Imhof R, Bedford MT, Natoli G, Hottiger MO. Arginine methyltransferase CARM1 is a promoter-specific regulator of NFkappaB-dependent gene expression. EMBO J. 2005;24:85–96. doi: 10.1038/sj.emboj.7600500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lu T, Jackson MW, Singhi AD, Kandel ES, Yang M, Zhang Y, Gudkov AV, Stark GR. Validation-based insertional mutagenesis identifies lysine demethylase FBXL11 as a negative regulator of NFkappaB. Proc Natl Acad Sci U S A. 2009;106:16339–44. doi: 10.1073/pnas.0908560106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hassa PO, Covic M, Bedford MT, Hottiger MO. Protein arginine methyltransferase 1 coactivates NFkappaB-dependent gene expression synergistically with CARM1 and PARP1. J Mol Biol. 2008;377:668–78. doi: 10.1016/j.jmb.2008.01.044. [DOI] [PubMed] [Google Scholar]
- 48.Ganesh L, Yoshimoto T, Moorthy NC, Akahata W, Boehm M, Nabel EG, Nabel GJ. Protein methyltransferase 2 inhibits NFkappaB function and promotes apoptosis. Mol Cell Biol. 2006;26:3864–74. doi: 10.1128/MCB.26.10.3864-3874.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wei H, Wang B, Miyagi M, She Y, Gopalan B, Huang DB, Ghosh G, Stark GR, Lu T. PRMT5 dimethylates R30 of the p65 subunit to activate NFκB. Proc Natl Acad Sci U S A. 2013;110:13516–21. doi: 10.1073/pnas.1311784110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Perkins ND. Post-translational modifications regulating the activity and function of the nuclear factor kappa B pathway. Oncogene. 2006;25:6717–30. doi: 10.1038/sj.onc.1209937. [DOI] [PubMed] [Google Scholar]
- 51.Lu T, Jackson MW, Wang B, Yang M, Chance MR, Miyagi M, Gudkov AV, Stark GR. Regulation of NFkappaB by NSD1/FBXL11-dependent reversible lysine methylation of p65. Proc Natl Acad Sci U S A. 2010;107:46–51. doi: 10.1073/pnas.0912493107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lu T, Yang M, Huang DB, Wei H, Ozer GH, Ghosh G, Stark GR. Role of lysine methylation of NFκB in differential gene regulation. Proc Natl Acad Sci U S A. 2013;110:13510–5. doi: 10.1073/pnas.1311770110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Yang XD, Huang B, Li M, Lamb A, Kelleher NL, Chen LF. Negative regulation of NFkappaB action by Set9-mediated lysine methylation of the RelA subunit. EMBO J. 2009;28:1055–66. doi: 10.1038/emboj.2009.55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ea CK, Baltimore D. Regulation of NFkappaB activity through lysine monomethylation of p65. Proc Natl Acad Sci U S A. 2009;106:18972–7. doi: 10.1073/pnas.0910439106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Levy D, Kuo AJ, Chang Y, Schaefer U, Kitson C, Cheung P, Espejo A, Zee BM, Liu CL, Tangsombatvisit S, et al. Lysine methylation of the NFκB subunit RelA by SETD6 couples activity of the histone methyltransferase GLP at chromatin to tonic repression of NFκB signaling. Nat Immunol. 2011;12:29–36. doi: 10.1038/ni.1968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Yoshimatsu M, Toyokawa G, Hayami S, Unoki M, Tsunoda T, Field HI, Kelly JD, Neal DE, Maehara Y, Ponder BA, et al. Dysregulation of PRMT1 and PRMT6, Type I arginine methyltransferases, is involved in various types of human cancers. Int J Cancer. 2011;128:562–73. doi: 10.1002/ijc.25366. [DOI] [PubMed] [Google Scholar]
- 57.Frietze S, Lupien M, Silver PA, Brown M. CARM1 regulates estrogen-stimulated breast cancer growth through up-regulation of E2F1. Cancer Res. 2008;68:301–6. doi: 10.1158/0008-5472.CAN-07-1983. [DOI] [PubMed] [Google Scholar]
- 58.Aggarwal P, Vaites LP, Kim JK, Mellert H, Gurung B, Nakagawa H, Herlyn M, Hua X, Rustgi AK, McMahon SB, et al. Nuclear cyclin D1/CDK4 kinase regulates CUL4 expression and triggers neoplastic growth via activation of the PRMT5 methyltransferase. Cancer Cell. 2010;18:329–40. doi: 10.1016/j.ccr.2010.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Gu Z, Gao S, Zhang F, Wang Z, Ma W, Davis RE, Wang Z. Protein arginine methyltransferase 5 is essential for growth of lung cancer cells. Biochem J. 2012;446:235–41. doi: 10.1042/BJ20120768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Liu F, Zhao X, Perna F, Wang L, Koppikar P, Abdel-Wahab O, Harr MW, Levine RL, Xu H, Tefferi A, et al. JAK2V617F-mediated phosphorylation of PRMT5 downregulates its methyltransferase activity and promotes myeloproliferation. Cancer Cell. 2011;19:283–94. doi: 10.1016/j.ccr.2010.12.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Powers MA, Fay MM, Factor RE, Welm AL, Ullman KS. Protein arginine methyltransferase 5 accelerates tumor growth by arginine methylation of the tumor suppressor programmed cell death 4. Cancer Res. 2011;71:5579–87. doi: 10.1158/0008-5472.CAN-11-0458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Wei TY, Juan CC, Hisa JY, Su LJ, Lee YC, Chou HY, Chen JM, Wu YC, Chiu SC, Hsu CP, et al. Protein arginine methyltransferase 5 is a potential oncoprotein that upregulates G1 cyclins/cyclin-dependent kinases and the phosphoinositide 3-kinase/AKT signaling cascade. Cancer Sci. 2012;103:1640–50. doi: 10.1111/j.1349-7006.2012.02367.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Cho EC, Zheng S, Munro S, Liu G, Carr SM, Moehlenbrink J, Lu YC, Stimson L, Khan O, Konietzny R, et al. Arginine methylation controls growth regulation by E2F-1. EMBO J. 2012;31:1785–97. doi: 10.1038/emboj.2012.17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Butler JS, Zurita-Lopez CI, Clarke SG, Bedford MT, Dent SY. Protein-arginine methyltransferase 1 (PRMT1) methylates Ash2L, a shared component of mammalian histone H3K4 methyltransferase complexes. J Biol Chem. 2011;286:12234–44. doi: 10.1074/jbc.M110.202416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Pope AJ, Karuppiah K, Cardounel AJ. Role of the PRMT-DDAH-ADMA axis in the regulation of endothelial nitric oxide production. Pharmacol Res. 2009;60:461–5. doi: 10.1016/j.phrs.2009.07.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Landburg PP, Teerlink T, van Beers EJ, Muskiet FA, Kappers-Klunne MC, van Esser JW, Mac Gillavry MR, Biemond BJ, Brandjes DP, Duits AJ, et al. CURAMA study group Association of asymmetric dimethylarginine with sickle cell disease-related pulmonary hypertension. Haematologica. 2008;93:1410–2. doi: 10.3324/haematol.12928. [DOI] [PubMed] [Google Scholar]
- 67.Valkonen VP, Tuomainen TP, Laaksonen R. DDAH gene and cardiovascular risk. Vasc Med. 2005;10(Suppl 1):S45–8. doi: 10.1177/1358836X0501000107. [DOI] [PubMed] [Google Scholar]
- 68.Jeong SJ, Lu H, Cho WK, Park HU, Pise-Masison C, Brady JN. Coactivator-associated arginine methyltransferase 1 enhances transcriptional activity of the human T-cell lymphotropic virus type 1 long terminal repeat through direct interaction with Tax. J Virol. 2006;80:10036–44. doi: 10.1128/JVI.00186-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Shire K, Kapoor P, Jiang K, Hing MN, Sivachandran N, Nguyen T, Frappier L. Regulation of the EBNA1 Epstein-Barr virus protein by serine phosphorylation and arginine methylation. J Virol. 2006;80:5261–72. doi: 10.1128/JVI.02682-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Xie B, Invernizzi CF, Richard S, Wainberg MA. Arginine methylation of the human immunodeficiency virus type 1 Tat protein by PRMT6 negatively affects Tat Interactions with both cyclin T1 and the Tat transactivation region. J Virol. 2007;81:4226–34. doi: 10.1128/JVI.01888-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Bandyopadhyay S, Harris DP, Adams GN, Lause GE, McHugh A, Tillmaand EG, Money A, Willard B, Fox PL, Dicorleto PE. HOXA9 methylation by PRMT5 is essential for endothelial cell expression of leukocyte adhesion molecules. Mol Cell Biol. 2012;32:1202–13. doi: 10.1128/MCB.05977-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Martin G, Ostareck-Lederer A, Chari A, Neuenkirchen N, Dettwiler S, Blank D, Rüegsegger U, Fischer U, Keller W. Arginine methylation in subunits of mammalian pre-mRNA cleavage factor I. RNA. 2010;16:1646–59. doi: 10.1261/rna.2164210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Pesiridis GS, Diamond E, Van Duyne GD. Role of pICLn in methylation of Sm proteins by PRMT5. J Biol Chem. 2009;284:21347–59. doi: 10.1074/jbc.M109.015578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kuhn P, Xu W. Protein arginine methyltransferases: nuclear receptor coregulators and beyond. Prog Mol Biol Transl Sci. 2009;87:299–342. doi: 10.1016/S1877-1173(09)87009-9. [DOI] [PubMed] [Google Scholar]
- 75.Obianyo O, Causey CP, Jones JE, Thompson PR. Activity-based protein profiling of protein arginine methyltransferase 1. ACS Chem Biol. 2011;6:1127–35. doi: 10.1021/cb2001473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Cheng D, Yadav N, King RW, Swanson MS, Weinstein EJ, Bedford MT. Small molecule regulators of protein arginine methyltransferases. J Biol Chem. 2004;279:23892–9. doi: 10.1074/jbc.M401853200. [DOI] [PubMed] [Google Scholar]
- 77.Arrowsmith CH, Bountra C, Fish PV, Lee K, Schapira M. Epigenetic protein families: a new frontier for drug discovery. Nat Rev Drug Discov. 2012;11:384–400. doi: 10.1038/nrd3674. [DOI] [PubMed] [Google Scholar]
- 78.Allan M, Manku S, Therrien E, Nguyen N, Styhler S, Robert MF, Goulet AC, Petschner AJ, Rahil G, Robert Macleod A, et al. N-Benzyl-1-heteroaryl-3-(trifluoromethyl)-1H-pyrazole-5-carboxamides as inhibitors of co-activator associated arginine methyltransferase 1 (CARM1) Bioorg Med Chem Lett. 2009;19:1218–23. doi: 10.1016/j.bmcl.2008.12.075. [DOI] [PubMed] [Google Scholar]
- 79.Huynh T, Chen Z, Pang S, Geng J, Bandiera T, Bindi S, Vianello P, Roletto F, Thieffine S, Galvani A, et al. Optimization of pyrazole inhibitors of Coactivator Associated Arginine Methyltransferase 1 (CARM1) Bioorg Med Chem Lett. 2009;19:2924–7. doi: 10.1016/j.bmcl.2009.04.075. [DOI] [PubMed] [Google Scholar]
- 80.Antonysamy S, Bonday Z, Campbell RM, Doyle B, Druzina Z, Gheyi T, Han B, Jungheim LN, Qian Y, Rauch C, et al. Crystal structure of the human PRMT5:MEP50 complex. Proc Natl Acad Sci U S A. 2012;109:17960–5. doi: 10.1073/pnas.1209814109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Rajpurohit R, Lee SO, Park JO, Paik WK, Kim S. Enzymatic methylation of recombinant heterogeneous nuclear RNP protein A1. Dual substrate specificity for S-adenosylmethionine:histone-arginine N-methyltransferase. J Biol Chem. 1994;269:1075–82. [PubMed] [Google Scholar]
- 82.Lin WJ, Gary JD, Yang MC, Clarke S, Herschman HR. The mammalian immediate-early TIS21 protein and the leukemia-associated BTG1 protein interact with a protein-arginine N-methyltransferase. J Biol Chem. 1996;271:15034–44. doi: 10.1074/jbc.271.25.15034. [DOI] [PubMed] [Google Scholar]
- 83.Abramovich C, Yakobson B, Chebath J, Revel M. A protein-arginine methyltransferase binds to the intracytoplasmic domain of the IFNAR1 chain in the type I interferon receptor. EMBO J. 1997;16:260–6. doi: 10.1093/emboj/16.2.260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Tang J, Kao PN, Herschman HR. Protein-arginine methyltransferase I, the predominant protein-arginine methyltransferase in cells, interacts with and is regulated by interleukin enhancer-binding factor 3. J Biol Chem. 2000;275:19866–76. doi: 10.1074/jbc.M000023200. [DOI] [PubMed] [Google Scholar]
- 85.Herrmann F, Bossert M, Schwander A, Akgün E, Fackelmayer FO. Arginine methylation of scaffold attachment factor A by heterogeneous nuclear ribonucleoprotein particle-associated PRMT1. J Biol Chem. 2004;279:48774–9. doi: 10.1074/jbc.M407332200. [DOI] [PubMed] [Google Scholar]
- 86.Dolzhanskaya N, Merz G, Denman RB. Alternative splicing modulates protein arginine methyltransferase-dependent methylation of fragile X syndrome mental retardation protein. Biochemistry. 2006;45:10385–93. doi: 10.1021/bi0525019. [DOI] [PubMed] [Google Scholar]
- 87.Rho J, Choi S, Jung CR, Im DS. Arginine methylation of Sam68 and SLM proteins negatively regulates their poly(U) RNA binding activity. Arch Biochem Biophys. 2007;466:49–57. doi: 10.1016/j.abb.2007.07.017. [DOI] [PubMed] [Google Scholar]
- 88.Zhao X, Jankovic V, Gural A, Huang G, Pardanani A, Menendez S, Zhang J, Dunne R, Xiao A, Erdjument-Bromage H, et al. Methylation of RUNX1 by PRMT1 abrogates SIN3A binding and potentiates its transcriptional activity. Genes Dev. 2008;22:640–53. doi: 10.1101/gad.1632608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Jobert L, Argentini M, Tora L. PRMT1 mediated methylation of TAF15 is required for its positive gene regulatory function. Exp Cell Res. 2009;315:1273–86. doi: 10.1016/j.yexcr.2008.12.008. [DOI] [PubMed] [Google Scholar]
- 90.Qi C, Chang J, Zhu Y, Yeldandi AV, Rao SM, Zhu YJ. Identification of protein arginine methyltransferase 2 as a coactivator for estrogen receptor alpha. J Biol Chem. 2002;277:28624–30. doi: 10.1074/jbc.M201053200. [DOI] [PubMed] [Google Scholar]
- 91.Buss H, Dörrie A, Schmitz ML, Frank R, Livingstone M, Resch K, Kracht M. Phosphorylation of serine 468 by GSK-3beta negatively regulates basal p65 NFkappaB activity. J Biol Chem. 2004;279:49571–4. doi: 10.1074/jbc.C400442200. [DOI] [PubMed] [Google Scholar]
- 92.Swiercz R, Cheng D, Kim D, Bedford MT. Ribosomal protein rpS2 is hypomethylated in PRMT3-deficient mice. J Biol Chem. 2007;282:16917–23. doi: 10.1074/jbc.M609778200. [DOI] [PubMed] [Google Scholar]
- 93.Lee J, Bedford MT. PABP1 identified as an arginine methyltransferase substrate using high-density protein arrays. EMBO Rep. 2002;3:268–73. doi: 10.1093/embo-reports/kvf052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Lee YH, Coonrod SA, Kraus WL, Jelinek MA, Stallcup MR. Regulation of coactivator complex assembly and function by protein arginine methylation and demethylimination. Proc Natl Acad Sci U S A. 2005;102:3611–6. doi: 10.1073/pnas.0407159102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Ito T, Yadav N, Lee J, Furumatsu T, Yamashita S, Yoshida K, Taniguchi N, Hashimoto M, Tsuchiya M, Ozaki T, et al. Arginine methyltransferase CARM1/PRMT4 regulates endochondral ossification. BMC Dev Biol. 2009;9:47. doi: 10.1186/1471-213X-9-47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Cheng D, Côté J, Shaaban S, Bedford MT. The arginine methyltransferase CARM1 regulates the coupling of transcription and mRNA processing. Mol Cell. 2007;25:71–83. doi: 10.1016/j.molcel.2006.11.019. [DOI] [PubMed] [Google Scholar]
- 97.Baldwin GS, Carnegie PR. Specific enzymic methylation of an arginine in the experimental allergic encephalomyelitis protein from human myelin. Science. 1971;171:579–81. doi: 10.1126/science.171.3971.579. [DOI] [PubMed] [Google Scholar]
- 98.Brahms H, Raymackers J, Union A, de Keyser F, Meheus L, Lührmann R. The C-terminal RG dipeptide repeats of the spliceosomal Sm proteins D1 and D3 contain symmetrical dimethylarginines, which form a major B-cell epitope for anti-Sm autoantibodies. J Biol Chem. 2000;275:17122–9. doi: 10.1074/jbc.M000300200. [DOI] [PubMed] [Google Scholar]
- 99.Barth S, Liss M, Voss MD, Dobner T, Fischer U, Meister G, Grässer FA. Epstein-Barr virus nuclear antigen 2 binds via its methylated arginine-glycine repeat to the survival motor neuron protein. J Virol. 2003;77:5008–13. doi: 10.1128/JVI.77.8.5008-5013.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Frankel A, Yadav N, Lee J, Branscombe TL, Clarke S, Bedford MT. The novel human protein arginine N-methyltransferase PRMT6 is a nuclear enzyme displaying unique substrate specificity. J Biol Chem. 2002;277:3537–43. doi: 10.1074/jbc.M108786200. [DOI] [PubMed] [Google Scholar]
- 101.Boulanger MC, Liang C, Russell RS, Lin R, Bedford MT, Wainberg MA, Richard S. Methylation of Tat by PRMT6 regulates human immunodeficiency virus type 1 gene expression. J Virol. 2005;79:124–31. doi: 10.1128/JVI.79.1.124-131.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Sgarra R, Lee J, Tessari MA, Altamura S, Spolaore B, Giancotti V, Bedford MT, Manfioletti G. The AT-hook of the chromatin architectural transcription factor high mobility group A1a is arginine-methylated by protein arginine methyltransferase 6. J Biol Chem. 2006;281:3764–72. doi: 10.1074/jbc.M510231200. [DOI] [PubMed] [Google Scholar]
- 103.Karkhanis V, Wang L, Tae S, Hu YJ, Imbalzano AN, Sif S. Protein arginine methyltransferase 7 regulates cellular response to DNA damage by methylating promoter histones H2A and H4 of the polymerase δ catalytic subunit gene, POLD1. J Biol Chem. 2012;287:29801–14. doi: 10.1074/jbc.M112.378281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Kim JD, Kako K, Kakiuchi M, Park GG, Fukamizu A. EWS is a substrate of type I protein arginine methyltransferase, PRMT8. Int J Mol Med. 2008;22:309–15. [PubMed] [Google Scholar]
- 105.Cook JR, Lee JH, Yang ZH, Krause CD, Herth N, Hoffmann R, Pestka S. FBXO11/PRMT9, a new protein arginine methyltransferase, symmetrically dimethylates arginine residues. Biochem Biophys Res Commun. 2006;342:472–81. doi: 10.1016/j.bbrc.2006.01.167. [DOI] [PubMed] [Google Scholar]