

Abstract
Birds, fish and other animals routinely use unsteady effects to save energy by alternating between phases of active propulsion and passive coasting. Here, we construct a minimal model for such behaviour that can be couched as an optimal control problem via an analogy to travelling with a rechargeable battery. An analytical solution of the optimal control problem proves that intermittent locomotion has lower energy requirements relative to steady-state strategies. Additional realistic hypotheses, such as the assumption that metabolic cost at a given power should be minimal (the fixed gear hypothesis), a nonlinear dependence of the energy storage rate on propulsion and/or a preferred average speed, allow us to generalize the model and demonstrate the flexibility of intermittent locomotion with implications for biological and artificial systems.
Keywords: intermittent locomotion, swimming, flight, optimal control
1. Introduction
The traditional approach to the mathematical study of animal locomotion is usually based on the assumption of steady state according to which organisms move with a constant speed through the surrounding environment. Yet, the actual movement of several species is characterized by bouts of activity, periodically interleaved with pauses that can last from milliseconds to minutes [1]. Indeed, recently, Gleiss et al. [2] have suggested that intermittent locomotion is an example of convergent evolution wherein animals from distant lineages have developed similar strategies to similar problems to move efficiently. For example, the unsteady locomotion of fish that alternate phases of active tail swinging with passive coasting phases saves energy relative to continuous swimming [3]. Indeed Muller et al. [4] compared the intermittent swimming performance of young and adult tropical fish and showed that the amount of time spent in the passive coasting phase increases with age, suggesting that hydrodynamical and behavioural changes can support such energetically favourable strategies of locomotion. Furthermore, intermittent swimming is not just found in fish, but is widespread across a large variety of species including jellyfish [5] and marine mammals such as dolphins and sea lions [6]. Moving beyond marine organisms, intermittent locomotion is also seen in birds and broadly classified into bounding flight and intermittent flight [7]; in the former modality, a bird periodically switches between active flapping phases and gliding phases, whereas in the second mode the passive phase corresponds to ballistic flight with folded wings. Experiments suggest that bounding flight represents an energetically efficient way of locomotion for small or young birds, whereas intermittent flight is characteristic of medium-sized birds [8–12]. In fact, the role of body lift in the ballistic or bounding phase for increasing the amount of saved energy was highlighted in [13], while a comparison of bats and swifts [14] suggests that the latter can exploit their high lift-to-drag ratio during gliding to save up to 15% of the energy compared with continuous flapping, whereas the morphology and flight style of bats do not allow such animals to efficiently use any stored potential energy and thus force them to use continuous flapping. Additionally, Kramer & McLoughlin [1] and Tobalske [15] have also suggested that intermittent locomotion allows animals to achieve secondary goals, such as fatigue recovery, manoeuvring, stability and sensory (visual) field focus and detectability.
The biological evidence showing that unsteady locomotion might in fact be the norm and steady-state locomotion is the exception begs for a move away from characterizing locomotion based on a mathematically convenient steady-state assumption. A possible guiding principle for developing new models and understanding the occurrence of intermittent locomotion is energy optimality, i.e. the assumption that animals minimize the total energy spent for locomotion [16,17]. In a pioneering example of this approach, Weihs [18] used simple energy arguments to show how alternating bouts of active tail swinging and passive coasting phases can save up to 50% of the energy used by fish compared with steady swimming. Such models have since been extended to cover a larger range of swimming speeds [19,20], and similar arguments have also been used to explain the occurrence of intermittent and bounding flight in birds [7,21,22]. The crucial ingredient of such models is the capability of storing energy, usually gravitational energy, during the active phase and then using it for propulsion in the passive phase.
On the one hand, while simple energetic formulations are capable of qualitatively describing the benefits of intermittent locomotion, they neglect the crucial dynamical aspects of locomotion, and therefore they are limited in their descriptive capability. On the other hand, dynamical models typically couple the body dynamics with the dynamics of the surrounding fluid environment and tend to be complex, amenable only to computational analysis and specifically tailored to the case at hand. For example, in [23] a model for fish-like swimming that includes a flexible body (the fish) interacting with a viscous fluid is evaluated for the efficiency of burst-and-coast swimming, while reduced order models [24,25] have been used to numerically find optimal solutions for intermittent flight and dynamic soaring in birds. The fact that similar intermittent locomotion strategies have arisen in animals that are not closely related [2] suggests a search for basic principles that might be gleaned from a general model.
Here, we propose a minimal model that incorporates the main qualitative features of the phenomenon including a dynamic formulation of the problem of locomotion with multiple allowable strategies that account for active propulsion and passive coasting, with simple constraints on the power and force that can be generated. Guided by the thought that intermittent locomotion is a strategic outcome, we couch our problem in terms of optimal control theory, which provides the natural language for such questions. Although we often refer to birds to help to build physical intuition about the formulation and the solutions, our formulation allows for a unified treatment of intermittent locomotion strategies in air and water. In particular, our minimal dynamical model for intermittent locomotion stated as an optimal control problem has solutions that describe the occurrence of unsteady locomotion and make it possible to extract the dependence of the qualitative features of unsteady motion on the system parameters. In §2, we introduce our mathematical model and use a combination of numerical and analytical methods to solve it, while providing an interpretation in terms of prior observations. In §3, we improve our initial formulation to account for additional realistic hypotheses and to show how the qualitative features of the intermittent locomotion strategy depend on the system parameters. Finally, in §4, we conclude with a discussion about the relevance of our models for natural and artificial locomotion strategies.
2. A minimal model for intermittent locomotion
Intermittent locomotion can be broadly defined as a strategy where animals periodically alternate between phases of active propulsion and phases of passive coasting. Such a strategy is viable only if animals are capable of storing part of the energy spent during the active phase in a ‘reservoir’ from which energy can be drawn during the passive phase. A minimal model for intermittent locomotion then needs to have at least the following three ingredients: (i) a dynamical model of the body in its environment, (ii) a (bounded) amount of energy that can be used for active propulsion, and (iii) a mechanism to store a fraction of the actuation energy to be used later during the passive phase.
A mechanical caricature of a system embedding all these elements is a car with a rechargeable battery, shown in figure 1. When the car moves, it is propelled on a horizontal track either by an engine (active phase) or by a battery that has been previously charged by the engine itself (passive phase). Here, the battery plays the role of ‘energy reservoir’ that is usually played by the gravitational potential energy in fish and birds (but see also [21] for a discussion about the importance of kinetic energy as an energy reservoir in birds).
Figure 1.
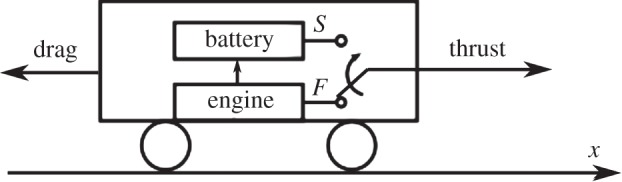
Schematic of a minimal model for intermittent locomotion. During the active phase, the engine provides the thrust and, at the same time, charges a battery. The charge stored in the battery can then be used to provide thrust when the engine is turned off, during the passive phase. The dimensionless dynamics of such a system is described by equations (2.5)–(2.7).
The position of the car is governed by the dynamical equation
| 2.1 |
where x is the location of the centre of mass along the track, F is the thrust provided by the engine, m is the car mass, c is the viscous friction coefficient and S is the potential energy stored in the battery. For simplicity, here we have assumed the drag force to be proportional to the velocity 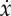 , but the introduction of a quadratic drag changes our results only quantitatively, not qualitatively, as shown in appendix A.
, but the introduction of a quadratic drag changes our results only quantitatively, not qualitatively, as shown in appendix A.
In this minimal setting, we start with the mechanical energy as a proxy for the total energy e(t) required for locomotion, writing
| 2.2 |
although as we increase the complexity of the model, we also account for a metabolic contribution later, in §3.
Finally, we assume that the charge in the battery is governed by the dynamical equation
| 2.3 |
where β represents the fraction of the total actuation energy that is transformed into stored potential energy and γ is the natural discharge rate. We note that, if F(t) is piecewise constant, a plausible solution based on our numerical results discussed in §2a is S(t)=(F(t)β−γ)x(t) and the dynamical equation (2.1) reads
| 2.4 |
In the following, we use this equation for the dynamics of the body (car), keeping in mind that the results are self-consistent only if the thrust F is piecewise constant.
Assuming that there is an upper bound on the thrust 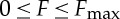 , as is to be expected in any biological situation, we find that the problem has a characteristic length scale
, as is to be expected in any biological situation, we find that the problem has a characteristic length scale 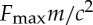 and a typical time scale m/c. Then we can write the dimensionless equations of motion as, with a little abuse of notation,
and a typical time scale m/c. Then we can write the dimensionless equations of motion as, with a little abuse of notation,
| 2.5 |
| 2.6 |
| 2.7 |
| 2.8 |
| 2.9 |
| 2.10 |
Now, we consider the problem of wanting the car to travel from the initial position x(0)=0 to a desired location xf. There are an infinite number of strategies to achieve this goal, ranging from one extreme of using continuous thrust to varying degrees of intermittent locomotion where active phases of duration ta are periodically interleaved by coasting phases of duration tg with possibly different duty cycles ta/(ta+tg)=ta/tf. We think of the non-trivial strategy associated with intermittent locomotion as an optimal one aimed at minimizing the total energy [7,21], although our approach demands the solution of a dynamical problem rather than energy minimization. To characterize this optimal strategy, we neglect the transients associated with the initial and final part of the trajectory, focus on a single period of on–off locomotion, and define the performance index
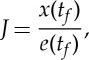 |
2.11 |
which is the inverse of the specific energy, i.e. of the energy required per unit length of travel. Our optimal control problem then states that the best strategy requires maximizing the performance index J subject to the dynamical constraints (2.5)–(2.8), the starting conditions x(0)=S(0)=e(0)=0 and the periodic boundary conditions S(0)=S(tf)=0 and 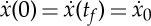 that are necessary to make it possible to arrive at the best combination of active propulsion and passive coasting.
that are necessary to make it possible to arrive at the best combination of active propulsion and passive coasting.
(a). Numerical results
Analytical solutions to optimal control problems such as (2.5)–(2.10) are, in general, very difficult to obtain; therefore, we first use numerical tools to gain some intuition. There exist several open-source and commercial numerical packages for finding solutions to optimal control problems, for example GPOPS [26] and PSOPT [27], which convert a dynamical optimal control problem to a standard finite-dimensional static optimization problem that is easier to solve. While implementation details such as the choice of time discretization or finite difference scheme differ among different packages, if the time grid is sufficiently fine the approximation error introduced is negligible, except in pathological cases. After converting the original optimal control problem to a standard finite-dimensional optimization problem where the dynamical equations and the input and state constraints on a generic variable s(t) become nonlinear constraints in the optimization variables si=s(ti), i=0,1,…,N, we use nonlinear optimization techniques, such as interior point methods or sequential quadratic programming, to solve the resulting finite-dimensional optimization problem [28]. We emphasize that most nonlinear optimization routines converge to local minima, and therefore one usually needs to use different initial guesses to find the global minimum. Here, we use the GPOPS package that exploits pseudo-spectral methods to achieve fast convergence and the sequential quadratic programming solver SNOPT [29] as our nonlinear optimization routine, available freely at www.gpops.org.
In figure 2, we report two examples of optimal trajectories obtained by setting β=0.75, γ=0.25 and 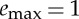 and starting from different initial guesses for the optimization variables. We note that, although the thrust is piecewise constant in both cases, with different optimal values for the engine force F*, the travelled distance per unit energy J=3, and thus our formulation for the optimal control problem does not admit a unique solution. We note that the introduction of a quadratic drag does not affect the qualitative behaviour of the optimal trajectories, as shown in appendix A. Before we describe how additional assumptions allow us to obtain unique minima, we first use analytical approaches to understand the minimal model described above.
and starting from different initial guesses for the optimization variables. We note that, although the thrust is piecewise constant in both cases, with different optimal values for the engine force F*, the travelled distance per unit energy J=3, and thus our formulation for the optimal control problem does not admit a unique solution. We note that the introduction of a quadratic drag does not affect the qualitative behaviour of the optimal trajectories, as shown in appendix A. Before we describe how additional assumptions allow us to obtain unique minima, we first use analytical approaches to understand the minimal model described above.
Figure 2.

Optimal trajectories showing (i) displacement x, (ii) energy spent e (solid) and stored S (dashed), (iii) velocity 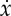 and (iv) thrust force F. These trajectories have been obtained by maximizing (2.11) under the constraints (2.5)–(2.10) with β=0.75, γ=0.25 and
and (iv) thrust force F. These trajectories have been obtained by maximizing (2.11) under the constraints (2.5)–(2.10) with β=0.75, γ=0.25 and 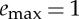 . The solution shown in (a) is obtained for F*=0.5 and the total distance x(tf)=0.5 is reached in tf=1.5 using a total energy e(tf)=0.17. The performance index J=3 and the control duty cycle ta/tf=67%. The solution reported in (b) is obtained for F*=1 and the total distance x(tf)=0.4 is reached in tf=1.2 using a total energy e(tf)=0.13. The performance index is still J=3 and the control duty cycle ta/tf=33%. (Online version in colour.)
. The solution shown in (a) is obtained for F*=0.5 and the total distance x(tf)=0.5 is reached in tf=1.5 using a total energy e(tf)=0.17. The performance index J=3 and the control duty cycle ta/tf=67%. The solution reported in (b) is obtained for F*=1 and the total distance x(tf)=0.4 is reached in tf=1.2 using a total energy e(tf)=0.13. The performance index is still J=3 and the control duty cycle ta/tf=33%. (Online version in colour.)
(b). Model analysis
Here, we first exploit an energetic formulation of the problem to characterize the dependence of the performance index J on the model parameters (β,γ). While this analysis leads to some general global insights, it does not provide detailed information about the behaviour of optimal trajectories. However, as we will see, an analytical solution of the optimal control problem (2.5)–(2.11) does allow us to describe a family of optimal solutions from which we can select one that satisfies additional conditions that are biologically realistic.
The numerical results presented in §2a suggest that the optimal thrust is piecewise constant,
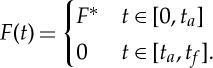 |
2.12 |
This allows us to determine the value of the performance index (2.11) by performing a phase plane analysis, i.e. by looking at the behaviour of e and S as a function of the position x instead of the temporal variable t. Indeed, the linearity in  of equations (2.6) and (2.7) implies that the battery charge S(t) and the mechanical energy e(t) are linear functions of the travelled distance x(t), as illustrated in figure 3. Therefore, we can write the maximum value for the potential energy, achieved at the end of the active phase, as
of equations (2.6) and (2.7) implies that the battery charge S(t) and the mechanical energy e(t) are linear functions of the travelled distance x(t), as illustrated in figure 3. Therefore, we can write the maximum value for the potential energy, achieved at the end of the active phase, as
| 2.13 |
so that the total travelled distance reads
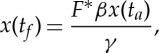 |
2.14 |
while the energy spent to cover the distance is
| 2.15 |
By comparing (2.14) and (2.15), we obtain
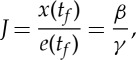 |
2.16 |
that is, the optimal performance index x(tf)/e(tf) is independent of the details of the dynamics, but depends on the balance between the efficiency of conversion between thrust and potential energy β and the passive discharge rate γ, allowing us to understand the reason for the degenerate nature of the optimal strategy obtained numerically.
Figure 3.
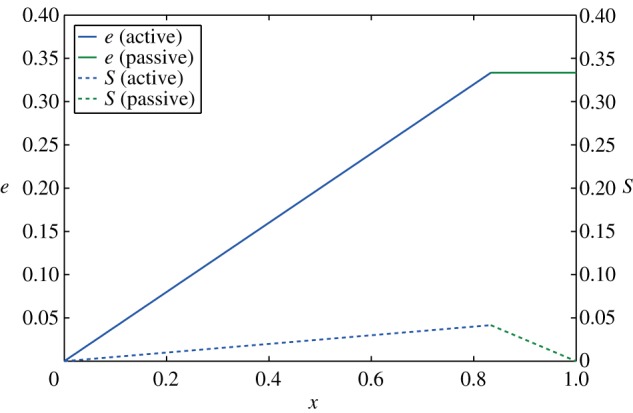
Mechanical energy e (solid) and stored potential energy S (dashed) as a function of the body position x, as obtained by setting β=0.75 and γ=0.25 in (2.6) and (2.7). (Online version in colour.)
In order to obtain the detailed structure of optimal trajectories, we now analyse the active and the gliding phase separately and characterize them using just four quantities: control duty cycle ta/tf, total period tf, thrust amplitude F* during the active phase and the initial velocity at the beginning of each cycle  .
.
During the active phase, i.e. for t∈[0,ta], we can analytically integrate (2.5) to obtain
| 2.17 |
and
| 2.18 |
where we have defined Γ=F*(1−β)+γ. The boundary conditions S(0)=S(tf)=0 imply that the charge in the battery must be always positive and this, together with (2.6), induces a lower bound on the minimum thrust during the active phase,
| 2.19 |
Similarly, during the gliding phase with t∈[ta,tf], we can analytically integrate (2.5)–(2.7) by setting F=0 and considering x(ta), e(ta) and S(ta) as new initial conditions, thus obtaining
| 2.20 |
| 2.21 |
| 2.22 |
| 2.23 |
We now use our explicit knowledge of the evolution of the state variables to convert the optimal control problem into a static optimization problem with  as unknowns and the two periodic boundary conditions
as unknowns and the two periodic boundary conditions 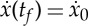 and S(tf)=0 as nonlinear constraints. By solving this system of equations, we then expect to find a two-parameter family of solutions satisfying the constraints. To derive analytical expressions for the quantities of interest, we first write the boundary condition
and S(tf)=0 as nonlinear constraints. By solving this system of equations, we then expect to find a two-parameter family of solutions satisfying the constraints. To derive analytical expressions for the quantities of interest, we first write the boundary condition 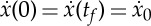 as
as
| 2.24 |
and use the expression to obtain the total period tf as a function of the active phase duration ta, the thrust F* and the initial velocity 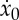 as
as
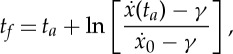 |
2.25 |
where x(ta) is given by (2.17) evaluated for t=ta. Similarly, by exploiting (2.23), we can rewrite the boundary condition S(0)=S(tf)=0 as
| 2.26 |
and express tf as a function of only ta, F* and 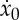 as
as
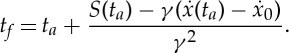 |
2.27 |
Finally, by imposing a consistency condition between (2.25) and (2.27), we derive an equation involving only ta, 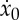 and F*,
and F*,
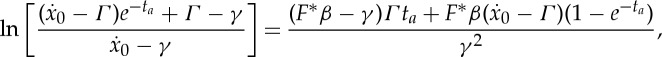 |
2.28 |
that can be numerically solved to find 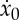 as a function of ta and F*. We note that equation (2.28), together with (2.25), allows us to build a family of admissible trajectories parametrized by ta and F*, each one having an associated range
as a function of ta and F*. We note that equation (2.28), together with (2.25), allows us to build a family of admissible trajectories parametrized by ta and F*, each one having an associated range
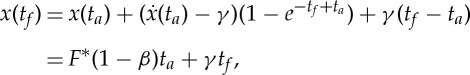 |
2.29 |
where we have used (2.24) and (2.17) to simplify the expressions above.
3. Adding additional realism
Our analysis of the minimal model in the previous section shows that there is no unique intermittent locomotion strategy that maximizes the performance index. Although this is a degenerate case from a mathematical point of view, such a degeneracy might actually be beneficial for animals because they can exploit precisely this flexibility to achieve secondary goals, as suggested by Kramer & McLaughlin [1] and Tobalske [15]. Here, we discuss additional hypotheses, of both a kinematic and dynamic nature, that have been suggested as being associated with intermittent locomotion, reframing them in the context of generalizing our minimal model. We show that a combination of a dynamical hypothesis along with a kinematic constraint allows us to recover a unique solution for the optimal control problem, and we discuss how this solution depends on the model parameters and is amenable to biological interpretation.
(a). Dynamical hypothesis I: fixed-gear hypothesis
Our minimal model used mechanical energy e(t) as a proxy for measuring the total energy spent during the propelled phase, completely neglecting the metabolic contribution. However, Alexander [17] and Tobalske [15] have highlighted the role of this additional contribution to the total energy budget for locomotion. Indeed, some authors have proposed a variant of this called the ‘fixed-gear hypothesis’, according to which the metabolic efficiency is a non-monotone function of the generated thrust [7], with the assumption that there is a sweet spot where the metabolic power reaches a minimum. In the context of our model, this corresponds to replacing equation (2.7) with
| 3.1 |
where we have assumed for simplicity that the metabolic cost is quadratic in F and reaches a minimum at 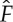 . We point out that it is only the presence of a (global) minimum at
. We point out that it is only the presence of a (global) minimum at  for F∈[0,1] that matters; the exact form of the metabolic contribution as a function of F is not relevant. Using the analogy between S and the gravitational potential energy, the quantity Fβ−γ corresponding to the ascending angle of flight and thus equation (3.1) characterizes the existence of an optimal ascending angle for which the lift-to-drag ratio is maximal.
for F∈[0,1] that matters; the exact form of the metabolic contribution as a function of F is not relevant. Using the analogy between S and the gravitational potential energy, the quantity Fβ−γ corresponding to the ascending angle of flight and thus equation (3.1) characterizes the existence of an optimal ascending angle for which the lift-to-drag ratio is maximal.
If 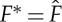 during the active phase, then the power spent on locomotion is minimized, and thus the optimal strategy corresponds to a choice of this value for the thrust. In our simplified setting, the additional term in (3.1) vanishes and the analysis in §2b allows us to find the optimal values for the period tf, the duty cycle ta/tf and the initial velocity
during the active phase, then the power spent on locomotion is minimized, and thus the optimal strategy corresponds to a choice of this value for the thrust. In our simplified setting, the additional term in (3.1) vanishes and the analysis in §2b allows us to find the optimal values for the period tf, the duty cycle ta/tf and the initial velocity 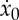 .
.
(b). Dynamical hypothesis II: nonlinear charge dynamics
However, experimental data reported by Tobalske et al. [11] challenge the validity of the fixed-gear hypothesis as a universal mechanism for choosing a unique value of the thrust during the active flapping phase in birds. An alternative, and somewhat complementary, approach to the fixed-gear hypothesis takes into account the nonlinear behaviour of the lift and drag coefficients on the value of the thrust F* and corresponds to assuming that β depends on the thrust amplitude F*. Therefore, we can replace equation (2.6) with
| 3.2 |
Once again, the exact shape of the function β(F) is not very relevant, and the optimal trajectory corresponds to the choice
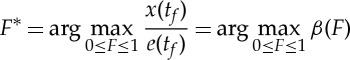 |
3.3 |
because x(tf)/e(tf)=β(F)/γ in analogy to (2.16).
(c). Kinematic constraint on average velocity
Both the fixed-gear and the nonlinear battery hypotheses imply that there is an optimal value of the thrust F* for which the performance index is maximized. Assuming that the thrust amplitude during the active phase is determined according to either of these hypotheses, we still have flexibility in choosing the duration ta of the active phase or, alternatively, the actuation duty cycle ta/tf. This remaining degree of freedom can be exploited to modulate the average velocity x(tf)/tf, as shown in figure 4, where we note that a given average velocity can be sustained by following one of two strategies: small accelerations for long times (small F* but large ta) or large acceleration for short times (large F* but small ta). We can quantify this behaviour by dividing equation (2.29) by tf to obtain
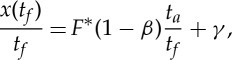 |
3.4 |
making the relationship between the average speed and actuation duty cycle ta/tf explicit. Thus, with one of the dynamical hypotheses above and this kinematic hypothesis, we find a unique optimal strategy for intermittent locomotion that maximizes the performance index while also hewing to maintain metabolic constraints and an optimal average velocity.
Figure 4.
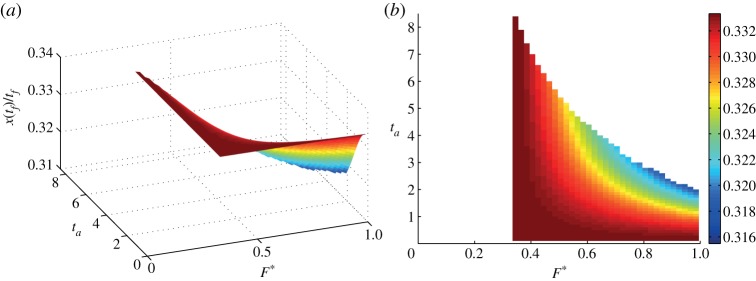
(a) Average velocity x(tf)/tf as a function of the active phase duration ta and the thrust amplitude F*. Data obtained with β=0.75, γ=0.25 and 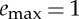 . (b) Top view showing that there is a minimal thrust γ/β below which no solution exists, whereas the curved boundary corresponds to the energy constraint (2.10). The colour bar on the right refers to the average velocity. (Online version in colour.)
. (b) Top view showing that there is a minimal thrust γ/β below which no solution exists, whereas the curved boundary corresponds to the energy constraint (2.10). The colour bar on the right refers to the average velocity. (Online version in colour.)
We note that, for our choice of parameters used in figure 4 (β=0.75, γ=0.25, 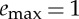 ), the average velocity can be varied by only about 5% because the large value of β does not allow the net force F*(1−β) to accelerate the body during the active phase and, at the same time, the small value for
), the average velocity can be varied by only about 5% because the large value of β does not allow the net force F*(1−β) to accelerate the body during the active phase and, at the same time, the small value for 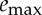 implies that the active phase itself does not last for a long time. Different choices for the parameters can allow for variations of 20–30% in the admissible average velocity.
implies that the active phase itself does not last for a long time. Different choices for the parameters can allow for variations of 20–30% in the admissible average velocity.
(d). Parameter sensitivity analysis
In the light of the unique solution obtained using the ideas discussed in the previous section, we now ask how the structure of the optimal trajectory is influenced by different choices for the system parameters, namely β and γ. Alternatively, we fix β(F)=αF so that F*=1 and study how the family of solutions varies as a function of the active phase duration ta.
In figure 5a, we report the performance index J=x(tf)/e(tf), the average and initial velocities x(tf)/tf and 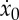 and the optimal duty cycle ta/tf as a function of α and for 0≤ta≤10. We note that, in agreement with equation (2.16), the performance index J scales linearly with α and does not depend on ta. Moreover, the duty cycle ta/tf is a monotonically decreasing function of α, consistent with the intuition that the benefits of intermittent locomotion become more evident as the animal improves its ability to store energy during the active phase. On the other hand, as α becomes vanishingly small (or better to its lower bound γ/F*; see (2.19)), the duty cycle tends to unity and the optimal strategy reduces to steady-state locomotion.
and the optimal duty cycle ta/tf as a function of α and for 0≤ta≤10. We note that, in agreement with equation (2.16), the performance index J scales linearly with α and does not depend on ta. Moreover, the duty cycle ta/tf is a monotonically decreasing function of α, consistent with the intuition that the benefits of intermittent locomotion become more evident as the animal improves its ability to store energy during the active phase. On the other hand, as α becomes vanishingly small (or better to its lower bound γ/F*; see (2.19)), the duty cycle tends to unity and the optimal strategy reduces to steady-state locomotion.
Figure 5.

Dependence of (i) performance index J=x(tf)/e(tf), (ii) average velocity x(tf)/tf, (iii) duty cycle ta/tf and (iv) initial velocity 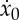 on (a) the fraction α of actuation energy that is stored as potential energy (γ=0.25) and (b) the natural discharge rate γ (β=0.75). Data obtained by solving (2.28) with F*=1 and 0≤ta≤10.
on (a) the fraction α of actuation energy that is stored as potential energy (γ=0.25) and (b) the natural discharge rate γ (β=0.75). Data obtained by solving (2.28) with F*=1 and 0≤ta≤10.
In figure 5b, we report the dependence of the same quantities on the natural discharge rate γ. We note that, as indicated by equation (2.16), variations in γ play a reciprocal effect with respect to variations in α. Once again, the performance index J is independent of ta, but is inversely proportional to γ. Larger values of γ correspond to steeper descent angles during the coasting phase, pushing the optimal strategy towards steady-state motion because now the system is not capable of efficiently using the stored energy during the passive phase.
This suggests that we can understand the bifurcation between continuous and intermittent locomotion by focusing on the transition regime, where the duty cycle can be written as δ=ta/tf=1−ε with ε≪1. By dividing both sides of (2.25) by tf and rearranging the terms, we find that
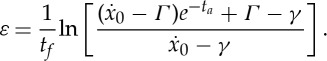 |
3.5 |
In the limit ta≪1, we can expand the exponential and the logarithmic terms in (3.5) to the first order and write
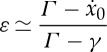 |
3.6 |
or, alternatively,
| 3.7 |
We can use this expression to derive 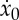 when the value of the force F or the duty cycle δ has been determined in accordance with, for example, one of the dynamical conditions and the kinematic constraint discussed previously in this section. Similarly, we can expand both sides of (2.28) to the first order to obtain
when the value of the force F or the duty cycle δ has been determined in accordance with, for example, one of the dynamical conditions and the kinematic constraint discussed previously in this section. Similarly, we can expand both sides of (2.28) to the first order to obtain
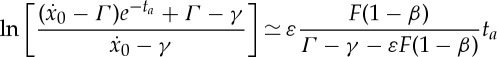 |
3.8 |
and
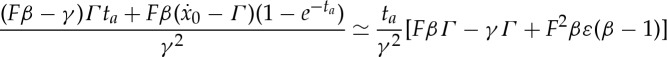 |
3.9 |
and, by equating the resulting expressions, we can obtain an analytical estimate for the duty cycle δ as
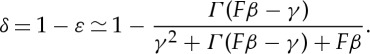 |
3.10 |
Although this expression is valid only in the limit ta≪1, it shows all the qualitative features depicted in figure 5. In particular, as the force F approaches its lower bound γ/β, the duty cycle approaches unity because the vehicle is not capable of storing energy in the reservoir S. On the other hand, as the force becomes very large, the duty cycle becomes vanishingly small and the locomotory strategy becomes a succession of short powerful thrust pulses interleaved with long passive phases. Similarly, as γ goes to zero (i.e. the passive phase becomes very efficient in using the stored energy), the duty cycle converges towards (Γ+1)−1, i.e. it is inversely proportional to F and β as shown in figure 5a. Finally, for a fixed value of the other parameters, the duty cycle is expected to increase as γ increases.
4. Discussion
The fact that intermittent locomotion has arisen multiple times in animal lineages suggests that it is an evolutionarily convergent solution in animal locomotion owing to similar constraints, such as maximizing efficiency in a system with finite energy and bounds on the power output. Our minimal one-dimensional model couches these facts in an optimal control setting and accounts for the basic constraints and a simple dynamical law that together determine locomotory strategy. Our analytical and numerical solutions of the optimal control problem show that unsteady locomotion represents an effective way for saving a conspicuous amount of energy. For example, if we compare the trajectory shown in figure 2b with the corresponding continuous locomotion strategy having the same average velocity and metabolic cost (3.1), we find that the performance index of the latter (J=0.58) is about five times lower than the one achieved with unsteady motion (J=3). The introduction of a quadratic drag instead of the linear function used in the analysis does not qualitatively alter the behaviour of optimal trajectories; indeed, we find that unsteady strategies are still energetically favourable, as discussed in appendix A. However, our model does not yield a single optimum and instead yields a two-parameter family of optimal control solutions. We interpret this flexibility in terms of the freedom to achieve secondary goals during intermittent locomotion, associated with different kinematic and dynamic hypotheses that constrain average velocity and metabolic activity. The dynamical behaviour of our model depends only on two non-dimensional parameters, the fractional rate of storing energy during the active phase β and the efficiency of using such energy during the passive phase γ. It thus suggests that a wide range of natural biological locomotory behaviours might lie in a low-dimensional phase space that might be usefully exploited by organisms.
In the context of biologically inspired locomotion, engineering solutions often operate under the implicit assumption that nature has optimized the performance of organisms with respect to the environment where they live in. For example, in [30], the authors discuss how flapping-wing propulsion observed in biology can be used as inspiration for designing new water and aerial vehicles, whereas in [31] the author proposes intermittent flight as an efficient strategy for small-scale autonomous aerial vehicles. Additionally, methods of controlling such autonomous vehicles using biologically inspired sensors and control strategies have been proposed in [32] and [33], but, in all these cases, the parameters that optimize various performance indices are rarely known. Our minimal model can characterize unsteady control strategies to improve performance, and the small number of parameters yields explicit suggestions for how to implement this in terms of the rate of charging and expending energy and the duty cycle.
Of course, our one-dimensional model is a simplification and thus leaves out many effects. A natural generalization of our model to describe motion in two and three dimensions can be potentially used to model and explore phugoid motion and other unsteady flight modes, although the additional (coupled) degrees of freedom render the optimal control problem nonlinear and thus prevent any analytical insight; nevertheless, these variations constitute new avenues of exploration.
Acknowledgements
We thank M. Kovac for discussions on the implications of our strategy for robotic systems.
Appendix A. The role of quadratic drag in intermittent locomotion
In our discussion in the main paper, we assumed the drag in (2.1) to be linear for simplicity. At first sight, one might expect that the introduction of a quadratic drag law that is more realistic at high velocities (and Reynolds numbers characteristic of many locomotory situations) will alter the results reported in the main body of this paper. However, if we replace equation (2.1) with
| A1 |
and solve the optimal control problem
| A2 |
| A3 |
| A4 |
| A5 |
| A6 |
| A7 |
using the same numerical method discussed in §2a, we obtain unsteady strategies as optimal solutions and note the appearance of bang–bang solutions. For example, in figure 6, we report the solution obtained by setting the parameters as in figure 2a in the main text. Thus, although there are quantitative differences relative to the linear drag setting, the qualitative behaviour of the optimal strategies remains the same, so that the qualitative results associated with our minimal model in the main text persist.
Figure 6.
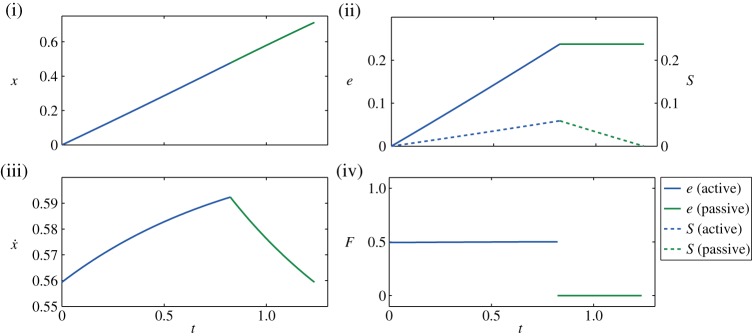
Optimal trajectory showing (i) displacement x, (ii) energy spent e (solid) and stored S (dashed), (iii) velocity 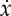 and (iv) thrust force F. This trajectory has been obtained by maximizing (2.11) under the constraints (A2)–(A7) with β=0.75, γ=0.25 and
and (iv) thrust force F. This trajectory has been obtained by maximizing (2.11) under the constraints (A2)–(A7) with β=0.75, γ=0.25 and 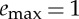 . The solution for F*=0.5 and the total distance x(tf)=0.71 is reached in tf=1.2 using a total energy e(tf)=0.237. The performance index J=3 and the control duty cycle ta/tf=67%. (Online version in colour.)
. The solution for F*=0.5 and the total distance x(tf)=0.71 is reached in tf=1.2 using a total energy e(tf)=0.237. The performance index J=3 and the control duty cycle ta/tf=67%. (Online version in colour.)
Funding statement
Harvard-Kavli Institute for Nano Bio Science and Technology, the Wyss Institute for Bioinspired engineering and the NSF Robobees project for partial financial support.
References
- 1.Kramer D, McLaughlin R. 2001. The behavioral ecology of intermittent locomotion. Am. Zool. 41, 137–153 (doi:10.1093/icb/41.2.137) [Google Scholar]
- 2.Gleiss A, et al. 2011. Convergent evolution in locomotory patterns of flying and swimming animals. Nat. Commun. 2, 352 (doi:10.1038/ncomms1350) [DOI] [PubMed] [Google Scholar]
- 3.Wu T. 2011. Fish swimming and bird/insect flight. Annu. Rev. Fluid Mech. 43, 25–58 (doi:10.1146/annurev-fluid-122109-160648) [Google Scholar]
- 4.Muller U, Stamhuis E, Videler J. 2000. Hydrodynamics of unsteady fish swimming and the effects of body size: comparing the flow fields of fish larvae and adults. J. Exp. Biol. 203, 193–206 [DOI] [PubMed] [Google Scholar]
- 5.Daniel T. 1985. Cost of locomotion: unsteady medusan swimming. J. Exp. Biol. 119, 149–164 [Google Scholar]
- 6.Williams T. 2001. Intermittent swimming by mammals: a strategy for increasing energetic efficiency during diving. Am. Zool. 41, 166–176 (doi:10.1093/icb/41.2.166) [Google Scholar]
- 7.Rayner J. 1985. Bounding and undulating flight in birds. J. Theor. Biol. 117, 47–77 (doi:10.1016/S0022-5193(85)80164-8) [Google Scholar]
- 8.Tobalske B, Dial K. 1994. Neuromuscular control and kinematics of intermittent flight in budgerigars (Melopsittacus undulatus). J. Exp. Biol. 187, 1–18 [DOI] [PubMed] [Google Scholar]
- 9.Tobalske B. 1995. Neuromuscular control and kinematics of intermittent flight in the European starling (Sturnus vulgaris). J. Exp. Biol. 198, 1259–1273 [DOI] [PubMed] [Google Scholar]
- 10.Tobalske B, Peacock W, Dial K. 1999. Kinematics of flap-bounding flight in the zebra finch over a wide range of speeds. J. Exp. Biol. 202, 1725–1739 [DOI] [PubMed] [Google Scholar]
- 11.Tobalske B, Puccinelli L, Sheridan D. 2005. Contractile activity of the pectoralis in the zebra finch according to mode and velocity of flap-bounding flight. J. Exp. Biol. 208, 2895–2901 (doi:10.1242/jeb.01734) [DOI] [PubMed] [Google Scholar]
- 12.Yoda K, Kohno H, Naito Y. 2004. Development of flight performance in the brown booby. Proc. R. Soc. Lond. B 271, 240 (doi:10.1098/rsbl.2003.0157) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tobalske B, Hearn J, Warrick D. 2009. Aerodynamics of intermittent bounds in flying birds. Exp. Fluids 46, 963–973 (doi:10.1007/s00348-009-0614-9) [Google Scholar]
- 14.Muijres FT, Henningsson P, Stuiver M, Hedenström A. 2012. Aerodynamic flight performance in flap-gliding birds and bats. J. Theor. Biol. 306, 120–128 (doi:10.1016/j.jtbi.2012.04.014) [DOI] [PubMed] [Google Scholar]
- 15.Tobalske B. 2007. Biomechanics of bird flight. J. Exp. Biol. 210, 3135–3146 (doi:10.1242/jeb.000273) [DOI] [PubMed] [Google Scholar]
- 16.Alexander RM. 1989. Optimization and gaits in the locomotion of vertebrates. Physiol. Rev. 69, 1199–1227 [DOI] [PubMed] [Google Scholar]
- 17.Alexander RM. 2003. Principles of animal locomotion. Princeton, NJ: Princeton University Press [Google Scholar]
- 18.Weihs D. 1974. Energetic advantages of burst swimming of fish. J. Theor. Biol. 48, 215–229 (doi:10.1016/0022-5193(74)90192-1) [DOI] [PubMed] [Google Scholar]
- 19.Videler JJ, Weihs D. 1982. Energetic advantages of burst-and-coast swimming of fish at high speeds. J. Exp. Biol. 97, 169–178 [DOI] [PubMed] [Google Scholar]
- 20.Stöcker S, Weihs D. 2001. Optimization of energetic advantages of burst swimming of fish. Math. Methods Appl. Sci. 24, 1387–1400 (doi:10.1002/mma.187) [Google Scholar]
- 21.Rayner J, Viscardi P, Ward S, Speakman J. 2001. Aerodynamics and energetics of intermittent flight in birds. Am. Zool. 41, 188–204 (doi:10.1093/icb/41.2.188) [Google Scholar]
- 22.Tobalske B. 2010. Hovering and intermittent flight in birds. Bioinspir. Biomim. 5, 045004 (doi:10.1088/1748-3182/5/4/045004) [DOI] [PubMed] [Google Scholar]
- 23.Chung M-H. 2009. On burst-and-coast swimming performance in fish-like locomotion. Bioinspir. Biomim. 4, 036001 (doi:10.1088/1748-3182/4/3/036001) [DOI] [PubMed] [Google Scholar]
- 24.Sachs G, Lenz J. 2011. New modeling approach for bounding flight in birds. Math. Biosci. 234, 75–83 (doi:10.1016/j.mbs.2011.08.005) [DOI] [PubMed] [Google Scholar]
- 25.Zhao Y. 2004. Optimal patterns of glider dynamic soaring. Opt. Control Appl. Methods 25, 67–89 (doi:10.1002/oca.739) [Google Scholar]
- 26.Rao A, Benson D, Darby C, Patterson M, Francolin C, Sanders I, Huntington G. 2010. Algorithm 902: GPOPS, a MATLAB software for solving multiple-phase optimal control problems using the Gauss pseudospectral method. ACM Trans. Math. Softw. 37, 22:1–22:39 (doi:10.1145/1731022.1731032) [Google Scholar]
- 27.Becerra VM. 2010. Solving complex optimal control problems at no cost with PSOPT. In Proc. 2010 IEEE Int. Symp. on Computer-Aided Control System Design (CACSD), Yokohama, Japan, 8–10 September 2010, pp. 1391–1396 Piscataway, NJ: IEEE [Google Scholar]
- 28.Luenberger DG, Ye Y. 2008. Linear and nonlinear programming, 3rd edn. New York, NY: Springer [Google Scholar]
- 29.Gill P, Murray W, Saunders M. 2005. SNOPT: an SQP algorithm for large-scale constrained optimization. SIAM Rev. 47, 99–131 (doi:10.1137/S0036144504446096) [Google Scholar]
- 30.Rozhdestvensky K, Ryzhov V. 2003. Aerohydrodynamics of flapping-wing propulsors. Prog. Aerospace Sci. 39, 585–633 (doi:10.1016/S0376-0421(03)00077-0) [Google Scholar]
- 31.Keating HA. 2002. A literature review on bounding flight in birds with applications to micro uninhabited air vehicles. DSTO Aeronautical and Maritime Research Laboratory Report DSTO-GD-0320. Fishermans Bend, Victoria, Australia: DSTO Aeronautical and Maritime Research Laboratory. See http://www.dsto.defence.gov.au/publications/2432/DSTO-GD-0320.pdf.
- 32.Schenato L. 2003. Analysis and control of flapping flight: from biological to robotic insects. PhD thesis, University of California, Berkeley, CA, USA
- 33.Epstein M, Waydo S, Fuller S, Dickson W, Straw A, Dickinson M, Murray R. 2007. Biologically inspired feedback design for Drosophila flight. In Proc. American Control Conf. (ACC '07), New York City, NY, 11–13 July 2007, pp. 3395–3401 Troy, NY: American Automatic Control Council [Google Scholar]