

Synopsis
This review explains the mechanisms underlying choices of pharmacotherapy for hypoxic-ischemic insults of both preterm and term babies. Some pre-clinical data are strong enough so that clinical trials are now underway. Challenges remain in deciding the best combination therapies for each age and insult.
Keywords: Brain injury, hypoxic ischemic encephalopathy, prematurity, preconditioning
Mechanisms of Brain Injury: Preterm vs. Term
The two most common causes of neonatal brain injury in the United States are extreme prematurity and hypoxic ischemic encephalopathy. In the U.S., one in 8 babies is born before term (37 to 40 weeks), and 1.44% of babies (56,000 per year) are born with a birth weight of 1250 grams or less.1 These small, preterm babies are at high risk of death or neurodevelopmental impairment: approximately 20% die before hospital discharge, and 40% of survivors develop long term intellectual or physical impairment, including cerebral palsy (CP).2-4 Care of preterm infants accounts for more than half of pediatric health care dollars spent.
The brain rapidly increases in size, shape and complexity during the second and third trimesters.5 Neurodevelopmental compromise can result from an interruption of normal development, or from damage to existing tissues. Brain development during this period is vulnerable to hypoxia-ischemia, oxidant stress, inflammation, excitotoxicity and poor nutrition. These exposures can result in structural, biochemical, and cell-specific injury.6 Pre-oligodendrocytes, which emerge and mature between 24 and 32 weeks of development, are particularly susceptible to injury, and damage to these cells can result in white matter injury.7 While intracranial hemorrhage, periventricular leukomalacia, inflammatory conditions and male gender are known risk factors for poor outcomes, little is known about how to improve these outcomes.
Hypoxic-ischemic encephalopathy (HIE) is estimated to contribute significantly to 23% of the 4 million neonatal deaths that occur annually.8 In the U.S., HIE occurs in 1.5 to 2 live births per thousand, with a higher incidence in premature infants.9 Untreated, the sequelae of moderate to severe HIE includes a 60 to 65% risk of mental retardation, cerebral palsy (CP), hydrocephalus, seizures, or death. Perinatal inflammation is increasingly recognized as an important contributor to neonatal HIE and poor neurodevelopmental outcomes:10 the presence of maternal fever alone increases the risk for CP, and chorioamnionitis further increases the risks for brain injury in both preterm and term infants.11,12 Timing of infection/inflammation relative to hypoxia is critical: it can be sensitizing (increase brain injury) if it occurs acutely or after 72 hours, but may be protective if it occurs 24 hours prior to hypoxia.13 This differential response is not fully understood, but may depend on activation of fetal/neonatal Toll like receptors in brain.14,15 Understanding the complex mechanisms of brain injury is essential to devising protective strategies.
The injury cascade
Although the cellular targets of hypoxia-ischemia are different depending on age and severity of insult, the basic cascade of injury occurs in a uniform way regardless of age and continues for a prolonged period of time. Cell death occurs in two main phases: primary death from hypoxia and energy depletion, followed by reperfusion and increased free radical (FR) formation, excitotoxicity, and nitric oxide production with secondary energy failure and delayed death (Figure 1). Recently a tertiary phase has been proposed, a phase when factors can worsen outcome, predispose a newborn to further injury, or prevent repair or regeneration after an initial insult to the brain.16 Such mechanisms include persistent inflammation and epigenetic changes, which cause a blockade of oligodendrocyte maturation, impaired neurogenesis, impaired axonal growth, or altered synaptogenesis.
Figure 1.
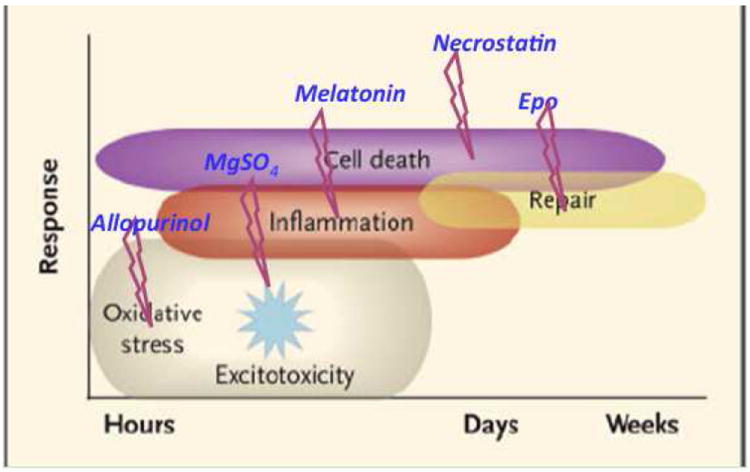
The injury cascade as it occurs over time. Potential therapeutics are inserted over the course of the cascade. See text for details on these agents.
The injury process begins with energy failure creating excitotoxicity. This is due to excessive glutamatergic activation that leads to progression of HI brain injury. Glutamate plays a key role in development, affecting progenitor cell proliferation, differentiation, migration and survival. Glutamate accumulates in the brain after HI17 from a variety of causes, including vesicular release from axons and reversal of glutamate transporters. Glutamatergic receptors include N-methyl-D-aspartate (NMDA), alpha-3-amino-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA), and kainate. Developmental differences in glutamate receptor expression contribute to the vulnerability of the immature brain (reviewed in Jensen Curr Opin Peds 2006).18 NMDA receptor activation, while important for synaptic plasticity and synaptogenesis can increase intracellular calcium, pro-apoptotic pathways via caspase-3 activation, FR formation and lipid peroxidation resulting in profound and widespread injury to the developing brain.
Oxidative stress is an important component of early injury in partnership with excitotoxicity, to the neonatal brain resulting from the excess formation of FRs (reactive oxygen species (ROS) and reactive nitrogen species (RNS)) under pathological conditions. These include superoxide anion (O2·-), hydroxyl radical (OH·), singlet oxygen (1O2·) and hydrogen peroxide (H2O2). FRs target lipids, protein and DNA, causing damage to these cellular components and initiating a cascade that results in cell death.19
These deleterious biological events trigger inflammatory processes that initially are harmful and later may be beneficial to the repair processes that occur after injury. The inflammatory response and cytokine production that accompanies infection may play a large role in cell damage and loss. Local microglia are activated early and produce pro-inflammatory cytokines such as TNF-α, IL-1β and IL-6, as well as glutamate, FRs, and nitric oxide (NO) and are the main immunocompetent cells in the immature brain. Depending on the stimulus, molecular context, and timing, these cells will acquire various phenotypes, which will be critical to the outcome of the injury.20
Cell death occurs throughout the cascade, moving from a purely necrotic type to apoptosis with a continuum of phenotypes emerging in the developing brain, the apoptosis-necrosis “continuum”. Throughout this process, mechanistic interactions between cell death and hybrid forms of cell death occur such as programmed or regulated necrosis or “necroptosis”.21 The mechanisms behind programmed necrosis in neonatal brain injury are still unraveling but are clearly regulated by the inflammatory processes, especially the TNF receptor superfamily, that are activated early in injury. There are many pharmacological agents that can impact on these injury phases (Box 1). We have chosen to highlight some of the important future therapeutics.
Box 1. Pharmacological Best Candidates for Impact on Injury phases.
| Best Candidates | |
|---|---|
| Antenatal | Postnatal |
| BH4 | Melatonin |
| Melatonin | Epo |
| nNOS inhibitors | NAC |
| Xenon | Xenon |
| Allopurinol | Allopurinol |
| Vit C and E | Vit C and E |
| Resveratrol | Resveratrol |
| NAC | Memantine |
| Topiramate | |
Targeting the injury response
Anti-excitotoxic agents
The earliest pharmacological strategies to protect the newborn brain were aimed at blocking the initial phases of injury, excitotoxicity and oxidative stress. Many of these agents failed because it is impossible to block normal developmental processes, like glutamatergic signaling, without harming the brain. Therefore, therapies aimed at blocking the NMDA receptor resulted in increased, rather than decreased cell death.22 However, some agents, like magnesium sulfate, used to stop preterm labor, appeared to have beneficial effects even though it blocked the NMDA receptor. Recent clinical trials have supported the use of antenatal magnesium.23 In 16 hospitals in Australia and New Zealand, 1062 women with fetuses younger than 30 weeks gestation were given a loading infusion of 16 mmol followed by 8 mmol MgSO4 for up to 24 h. Substantial gross motor dysfunction (3.4% vs. 6.6%; RR 0.51; 95%CI 0.29-0.91) and combined death or substantial gross motor dysfunction were significantly reduced in the MgSO4 group, although there were no significant differences in mortality or cerebral palsy in survivors. In the US, antenatal MgSO4 did not reduce the risk of the composite outcome of CP or death, much like the Australian study, but was seen to reduce moderate to severe CP without increasing the risk of death.23 The number needed to treat however was quite high at 56 (95% CI 34-164).
Another potential anti-excitotoxic agent is Xenon, a noble gas used as an anesthetic agent. It has action against the NMDA receptor and has been shown to be an effective agent against hypoxic-ischemic insult both to cortical neurons in vitro and in a number of in vivo models.24 Xenon lacks the dopamine-releasing properties that are present in other NMDA antagonists and this does not cause increased apoptotic cell death as seen with other NMDA antagonists. Perhaps the most promising aspect of Xenon pharmacology is it increases the translational efficiency of HIF-1α through an mTOR pathway25 that has resulted in its potential application in HIE for both post-injury treatment and as a preconditioner. The prolonged increase in expression of HIF 1a by xenon causes upregulation of cytoprotective proteins such as erythropoietin (Epo), VEGF and glucose transporter 1 protein.25 Indeed, when given with sevoflurane during labor to rats, it can precondition the fetal brain against a subsequent HI insult.26 When given to neonatal rats in combination with hypothermia, it improves both functional and structural outcomes, even when hypothermia is delayed, and the effect is sustained through adulthood.27
Anti-oxidants
Therapies aimed at reducing oxidative stress have proven to be efficacious both in the preterm and term injury states. Allopurinol was originally shown to be neuroprotective in postnatal day 7 rats after HI 28 but in humans was not seen to improve short or long term outcome in a small trial after birth asphyxia.29 It was postulated that the drug needed to be given before reperfusion injury set in, so trials are now underway to evaluate efficacy when given to the mother who have fetuses suspected of intrauterine hypoxia. In a randomized double blind placebo controlled multicenter study in progress, allopurinol is given IV antenatally with the primary outcome being serum brain damage markers (S100b) and oxidative stress markers (isoprostanes, etc) in umbilical cord blood; secondary outcome measures are neonatal mortality, serious composite neonatal morbidity and long-term neurological outcome.30 There is now a randomized placebo-controlled double blinded parallel group comparison study of hypothermia and allopurinol ongoing, the European ALBINO trial. Allopurinol will be given twice, 30 minutes postnatally and then 12 hours later in addition to hypothermia in moderate to severe HIE. Outcomes will be assessed at 2 years of life.
There are many antioxidants that have been investigated in both preterm and term HI injury. Scavengers such as melatonin, and vitamin E have shown promise. Lipid peroxidation inhibitors such as the lazaroids, gingko biloba and caffeic acid and free radical reducers such as ebselen and erythropoietin (Epo) have produced some amelioration of injury. Nitric oxide synthase inhibitors such as aminoguanidine, L-NAME and 7 nitroindazole and newer derivatives are still being investigated.31,32
The most promising of these agents appears to be melatonin that shows efficacy in both preterm and term injury. Melatonin has many targets along the injury cascade including oxidative stress, inflammation, apoptosis, mitochondrial failure as well as nuclear effect. The benefit lies in the lack of significant side effects in children and term neonates. A recent observational study showed that melatonin levels are deficient in preterm and term newborn infants and is now in trial daily for 7 days after premature birth to identify if it may reduce the risk of prematurity-associated brain injury (MINT; ISRCTN15119574). Another investigation is underway in the premature and full-term baby to identify optimal treatment doses (MELIP; NCT01340417 and MIND, NCT01340417), and there is a study to determine the effects of maternal supplementation on outcome in term infants (PREMELIP; ID pending). An Australian study evaluating melatonin to prevent brain injury in unborn growth restricted babies is ongoing where mothers receive melatonin during pregnancy and oxidative stress will be monitored in maternal serum, placenta and umbilical cord blood. A composite neonatal outcome will be evaluated (NCT01695070).
Dietary manipulations may also prove promising in neuroprotection. Pomegranate juice is rich in polyphenols that can protect the neonatal mouse brain against a HI insult when give to mothers in their drinking water 33. Even when given after the insult to neonatal animals, there is substantial protection in hippocampus, cortex and striatum 34. Omega-3 polyunsaturated fatty acid supplementation can reduce brain damage and improve long term neurological outcomes even 5 weeks after an HI insult to rodents. The effect is best appreciated in microglia where NFkB activation and release of inflammatory mediators are inhibited, thus providing an anti-inflammatory effect as well 35.
Anti-inflammatory agents
As mentioned above, melatonin has multiple targets in the injury cascade and is a perfect candidate for manipulating inflammation. In a number of animal models, small and large, preterm and term, it has shown efficacy against excitotoxic lesions as well as hypoxia-ischemia. A recent study in rodents revealed that melatonin preserved white matter and learning disabilities after ibotenate lesions to the postnatal day 5 brain and was equally efficacious against Il1beta injections.36 In a fetal sheep model at E90, cord occlusion produced substantial white matter injury that was blocked with melatonin.37 In term brain models of hypoxia-ischemia, melatonin markedly decreased microglial activation and preserved myelination.38 The most promising study to date in piglets revealed that melatonin, in combination with hypothermia, provided substantial improvement over hypothermia in preserving brain function measured by aEEG, and reduced cell death in the thalamus, the region most affected by the asphyxia insult.39 There is now a collaborative study between Hopital Robert Debre and St Thomas Hospital (Kings college London) as a proof of concept in neuroprotection study. It will be a double blinded randomized trial of premature newborns less than 28 weeks gestational age. Babies will be randomized to placebo, low dose melatonin or high dose melatonin and outcome will be assessed by MRI and neurodevelopmental outcome at 24 months (MINT Trial ISRCTN15119574).
Growth Factors as Neuroprotectants
Many growth factors have essential roles during fetal and postnatal brain development. While the effects of some, such as brain derived neurotrophic factor (BDNF), are largely restricted to the brain, others such as Epo, vascular endothelial growth factor (VEGF), granulocyte colony-stimulating factor (GCSF) and insulin like growth factor 1 (IGF-1) have important somatic effects in addition to their role in neurodevelopment. All of the factors listed above have been evaluated as neuroprotectant therapies for adult and neonatal brain injury. At this time, Epo is the best studied for this purpose, and is the closest to clinical use. The pleiotropic nature of these growth factors makes it essential to test meticulously for safety prior to clinical use, particularly since very high doses are often required for neuroprotection, given that these large molecules do not readily cross the blood brain barrier.
Erythropoietin (Epo)
Epo and its receptor (EpoR) are expressed in the developing CNS, and are required for normal brain development.40 Acute exposure to hypoxia upregulates the expression of EpoR on oligodendrocytes and neurons, without a commensurate increase in Epo expression.41 The presence of unbound cell surface EpoR drives cells of neuronal and oligodendrocyte lineage to apoptosis, while ligand-bound EpoR activates survival signaling pathways. With Epo binding, EpoR dimerize to activate anti-apoptotic pathways via phosphorylation of JAK2, phosphorylation and activation of MAPK, ERK1/2, as well as the PI3K/Akt pathway and STAT5, which are critical in cell survival.42 Epo also stimulates production of BDNF. Epo signaling inhibits early mechanisms of brain injury by its anti-inflammatory,43,44 anti-excitotoxic,45 anti-oxidant,46,47 and anti-apoptotic effects on neurons and oligodendrocytes. Repair of brain injury is also enhanced in the presence of Epo due to its positive effects on neurogenesis and angiogenesis, which are essential for plasticity and remodeling.48,49 Epo effects are dose-dependent, with multiple doses being more effective than single doses.50,51 Epo reduces neuronal loss and learning impairment following brain injury,52 and even when initiated as late as 48-72 hours after injury, there is evidence of improved behavioral outcomes, enhanced neurogenesis, increased axonal sprouting, and reduced white matter injury in animal models of brain injury.53,54
Epo is now under investigation for both term and preterm brain injury. The anti-inflammatory, anti-excitotoxic, and anti-oxidant effects are relevant to brain injury in both age groups. The specific effects of Epo in pre-oligodendrocytes may be most relevant to the white matter injury that characterizes preterm brain injury. Treatment approaches to acute brain injury in term infants (HIE) and preterm infants (IVH) should differ from preventative strategies in preterm infants. In the former, a shorter duration of high dose Epo would be most appropriate, while for the latter, a more prolonged treatment strategy that continues during the period of oligodendrocyte vulnerability would be most likely to succeed. In addition to the specific cellular effects on neurons and oligodendrocytes, this more prolonged treatment would also decrease the availability and potential toxicity of free iron, due to the erythropoietic effects of Epo, by increasing iron utilization.
Translation to clinical trials
Epo does not cross the placenta, so prenatal treatment is not an option. It is FDA approved and has a robust safety profile in neonates, with over 3000 neonates randomized to placebo controlled trials testing its erythropoietic effects.55 Doses required for neuroprotection are higher than those used for prevention and treatment of anemia, as only a small fraction of circulating Epo crosses the blood brain barrier. In animal models of neonatal brain injury, Epo doses of 1000-5000 U/kg/dose result in sustained neuroprotection, improving both short and long-term structure and function.56 Phase I/II trials have been done to establish safety and translational pharmacokinetics of Epo in preterm57,58 and term neonates.59 These studies suggest that 1000 U/kg/dose provides an area under the curve (AUC) most similar to a neuroprotective dose of 5000 U/kg in rodents (Table 1). The optimal dose and duration of treatment is likely to differ for treatment of HIE as compared to preventing or treating brain injury in preterm infants, and is not yet known. Note that the pharmacokinetics in preterm and term asphyxiated infants are different, with a longer half life noted at higher doses in HIE infants. Phase II and III studies are now underway for neuroprotection of both extreme prematurity and HIE in term infants (Box 2). In the U.S., a multicenter randomized controlled trial of preterm Epo neuroprotection is beginning (PENUT trial, NCT01378273). This study will use 1000 U/kg × 6 doses followed by 400 U/kg three times a week until 33 weeks of gestation. A Swiss trial has used 3000 U/kg for 3 doses in the first weeks of life. Enrollment is complete for this trial, with follow up underway. The BRITE study, comparing Darbepoetin, Epo and placebo, is showing improved outcomes in preterm neonates receiving either Darbepoetin or Epo. Erythropoietic agents (Epo and Darbepoetin) are also being studied in combination with hypothermia for the treatment of HIE in term infants. Pilot studies have shown safety and early signs of benefit, and larger studies are planned or ongoing in the U.S., France (NCT01732146), and China.
TABLE 1.
Epo Pharmacokinetics
| Epo Dose | AUC | Cmax | T1/2 | |
|---|---|---|---|---|
| P7 Rodents s.c | 5000 U/kg | 117,677 | 6,224 | 8.4 |
| P7 Rodents i.p. | 5000 U/kg | 140,331 | 10,015 | 6.7 |
|
| ||||
| Preterm infants < 1000 gm | 500 U/kg | 31,412 ± 2,780 | 8078 ± 538 | 5.4 ± 0.6 |
| 1000 U/kg | 81,498 ± 7,067 | 14,017 ± 1,293 | 7.1 ± 0.7 | |
| 2500 U/kg | 317,881±22,941 | 46,467 ± 2,987 | 8.7 ± 1.4 | |
|
| ||||
| Term HIE infants | 500 U/kg | 50,306 ± 6,7426 | 7046 ± 814 | 7.2 ± 1.9 |
| 1000 U/kg | 131,054 ± 17 083 | 13,780 ± 2,674 | 15.0 ± 4.5 | |
| 2500 U/kg | 328,002 ± 61 945 | 33,316 ± 7,377 | 18.7 ± 4.7 | |
Box 2. Clinical Trials of Neuroprotective agents.
-
Phase 1 and 2
Xenon and cooling therapy in babies at high risk of brain injury following poor condition at birth: Randomised pilot study. Bristol UK
-
Phase 2 and 3
Safety and efficacy of topiramate in neonates with HIE treated with hypothermia (NeoNATI) Florence Italy. NCT01241019
Neonatal erythropoietin and therapeutic hypothermia short-term outcome study (NEAT O Study) UCSF, multicenter, Thrasher funded
Darbepoetin administration in newborns undergoing cooling for encephalopathy (DANCE Study) Utah, multicenter, Thrasher funded. NCT01471015
-
Phase 3 Trials
A multicenter RCT of therapeutic hypothermia plus magnesium sulphate (MgSO4) versus therapeutic hypothermia plus placebo in the management of term and near term babies with HIE. Turkey NCT01646619
Optimizing cooling strategies at < 6 hours of age for neonatal HIE An NICHD funded project. NCT01192776
Phase III study of efficacy of high dose erythropoietin to prevent HIE sequelea in term newborn Paris, France. NCT01732146
Evaluation of systemic hypothermia initiated after 6 hours of age in infants ≥36 weeks gestation with HIE: A bayesian evaluation. An NICHD funded project. NCT00614744
Preterm Epo Neuroprotection (PENUT Trial). A multi-center, randomized placebo-controlled phase III 940-subject trial of erythropoietin for the neuroprotection of extremely low gestational age neonates An NINDS funded project. NCT01378273
Melatonin as a novel neuroprotectant in Preterm Infants Trial Study (MINT) UK trial funded by Medical Research Council. ISRCTN15119574
Brain derived neurotrophic factor (BDNF)
BDNF is an important growth factor during fetal brain formation, particularly in the hippocampus, cerebral cortex, basal forebrain and cerebellum. It is also active in adult neurogenesis. BDNF binds primarily to TrkB, a receptor tyrosine kinase, and activates MAP kinase and CAMKII which regulate CREB and synapsin transcription. Neuroprotection from glutamate toxicity is mediated through PI3-K and the Ras/MAPK signaling pathways, and involves an increase in bcl-2 proteins.60 Both exercise and caffeine increase BDNF secretion, thereby increasing recognition memory and neurogenesis.
Vascular endothelial growth factor (VEGF) is a growth factor, stimulated by HIF-1, that stimulates vasculogenesis and angiogenesis, essential processes needed for brain development and brain repair. In addition, VEGF exhibits specific neurotrophic and neuroprotective effects in adult and neonatal models of hypoxic ischemic brain injury.61 These effects also involve activation of Akt and ERKs. Tight regulation of this factor is needed because both over and under-expression can contribute to disease.
Granulocyte colony-stimulating factor (GCSF) is a hematopoietic glycoprotein that stimulates the clonal maturation of neutrophil progenitors and increases many functional activities of mature neutrophils. In addition to these hematopoietic effects, G-CSF and its receptor are expressed on neuronal cells in a variety of brain regions.62 GCSF has demonstrated neuroprotection in several models of brain injury, and is well tolerated at high doses. G-CSF has been reported to have anti-apoptotic, anti-inflammatory, anti-excitotoxic, and neurotrophic properties with demonstrated improved long term outcomes.63 It is currently being evaluated in a Phase II clinical trial for adult ischemic stroke (NCT00132470).64
Insulin like growth factor 1 (IGF-1)
IGF-1 has an important role in normal brain development, promoting neuronal growth, cellular proliferation and differentiation in vitro, and when injected directly into brain or when given intranasally.65 It has also been found to have neuroprotective effects, improving long-term function after hypoxic ischemic brain injury. However, its clinical application to neurological disorders is limited by its large molecular size, poor central uptake, and mitogenic potential.
Drug Delivery
When considering drugs for neuroprotection, drug delivery is an important consideration. For example, if a drug crosses the placenta and can be tolerated by the mother, prenatal treatments are an option, such as with xenon and melatonin. The ability to give the drug intravenously is important, given that many critically ill neonates cannot take oral medications. Some medications, such as melatonin, are presently only available as oral formulations. Inhalational agents such as xenon face the issue that if a significant oxygen requirement exists, the ability to deliver neuroprotective concentrations (30 to 50%) may be limited.
Small neuroactive peptides may have a therapeutic advantage over larger molecules such as Epo or GCSF, which are relatively large glycoproteins that do not cross the blood brain barrier well. Modified neuropeptides and mimetic peptides have been designed to overcome these barriers, and are in various stages of testing.
Another relatively novel approach is to use alternative delivery methods, targeting the inflammatory system. Polyamidoamine dendrimers have been shown to localize in activated microglia and astrocytes in the brain of newborn rabbits with CP, but not healthy controls.66 This nanotechnology approach has been used to deliver dendrimer-bound N-acetyl-l-cysteine (NAC) to the target inflammatory cells to suppresses neuroinflammation with excellent results, using much lower concentrations than are needed with systemic dosing.
Combination Therapies
As more information is learned about mechanisms of brain injury and neuroprotection, combination therapies may be applied. As mentioned above, some preclinical combination studies are already underway, for example, hypothermia + xenon, and hypothermia + Epo for the treatment of term HIE.
Gaps in knowledge
There remain gaps in knowledge (Box 3). All of these must be considered in the context of the developing neonate, as immune function, cell populations, and specific vulnerabilities and response to injury changes over time. As we become more knowledgeable in these areas, new approaches to neuroprotection may become apparent.
Box 3. Gaps in knowledge.
-
Cytokine response
-
Innate immunity differs in newborns compared to adults
Th1 preponderance in newborns
Th2 in adults
When does it change?
How is this affected by prematurity?
How does this affect brain injury/repair?
-
-
Microglial response
-
3 activation states of CNS microglia
M1- classical activation (tissue defense, proinflammatory)
M2 - alternative activation (repair, anti-inflammatory, fibrosis, matrix reconstruction)
M3 - acquired deactivation (immunosuppression, phagocytosis of apoptotic cells)
How are these states regulated?
Can we harness this response for healing?
Does it differ for preterm vs. term infants?
-
-
Preconditioning
Hypoxia
-
Inflammation
What are the molecular mechanisms?
Can these mechanisms be harnessed to improve outcomes?
Key Points.
There are many ways to achieve “neuroprotection”: preconditioning, salvaging, repair
Hypothermia is now standard of care for term HIE so studies to investigate additional therapies will be add-on to that treatment
Strategies that target multiple mechanisms and consider age appropriate mechanisms will be most fruitful
Footnotes
Disclosure statement: Neither author has anything to disclose
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Contributor Information
Sandra E. Juul, Email: sjuul@uw.edu, University of Washington, Department of Pediatrics, 1959 NE Pacific St, Box 356320, Seattle, Washington 98195, Telephone: (206) 221-6814; Fax: (206) 543-8926.
Donna M. Ferriero, Email: ferrierod@neuropeds.ucsf.edu, Neonatal Brain Disorders Laboratory, University of California, San Francisco, 675 Nelson Rising Lane, Room 494, Box 0663, San Francisco, California 94143, Phone: (415) 502-7319, Fax: (415) 486-2297.
References
- 1.Hamilton BE, Hoyert DL, Martin JA, Strobino DM, Guyer B. Annual summary of vital statistics: 2010-2011. Pediatrics. 2013;131:548–58. doi: 10.1542/peds.2012-3769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.O’Shea TM, Allred EN, Dammann O, et al. The ELGAN study of the brain and related disorders in extremely low gestational age newborns. Early human development. 2009;85:719–25. doi: 10.1016/j.earlhumdev.2009.08.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Stoll BJ, Hansen NI, Bell EF, et al. Neonatal outcomes of extremely preterm infants from the NICHD Neonatal Research Network. Pediatrics. 2010;126:443–56. doi: 10.1542/peds.2009-2959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gargus RA, Vohr BR, Tyson JE, et al. Unimpaired outcomes for extremely low birth weight infants at 18 to 22 months. Pediatrics. 2009;124:112–21. doi: 10.1542/peds.2008-2742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lodygensky GA, Vasung L, Sizonenko SV, Huppi PS. Neuroimaging of cortical development and brain connectivity in human newborns and animal models. J Anat. 2010;217:418–28. doi: 10.1111/j.1469-7580.2010.01280.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Volpe JJ. Brain injury in premature infants: a complex amalgam of destructive and developmental disturbances. Lancet Neurol. 2009;8:110–24. doi: 10.1016/S1474-4422(08)70294-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Back SA, Riddle A, McClure MM. Maturation-dependent vulnerability of perinatal white matter in premature birth. Stroke. 2007;38:724–30. doi: 10.1161/01.STR.0000254729.27386.05. [DOI] [PubMed] [Google Scholar]
- 8.Black RE, Cousens S, Johnson HL, et al. Global, regional, and national causes of child mortality in 2008: a systematic analysis. Lancet. 2010;375:1969–87. doi: 10.1016/S0140-6736(10)60549-1. [DOI] [PubMed] [Google Scholar]
- 9.Kurinczuk JJ, White-Koning M, Badawi N. Epidemiology of neonatal encephalopathy and hypoxicischaemic encephalopathy. Early human development. 2010;86:329–38. doi: 10.1016/j.earlhumdev.2010.05.010. [DOI] [PubMed] [Google Scholar]
- 10.van Vliet EOG, de Kieviet JF, Oosterlaan J, van Elburg RM. Perinatal Infections and Neurodevelopmental Outcome in Very Preterm and Very Low-Birth-Weight Infants A Meta- Analysis. JAMA Pediatr. 2013;167:662–8. doi: 10.1001/jamapediatrics.2013.1199. [DOI] [PubMed] [Google Scholar]
- 11.Badawi N, Kurinczuk JJ, Keogh JM, et al. Intrapartum risk factors for newborn encephalopathy: the Western Australian case-control study. Bmj. 1998;317:1554–8. doi: 10.1136/bmj.317.7172.1554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wu YW, Croen LA, Shah SJ, Newman TB, Najjar DV. Cerebral palsy in a term population: risk factors and neuroimaging findings. Pediatrics. 2006;118:690–7. doi: 10.1542/peds.2006-0278. [DOI] [PubMed] [Google Scholar]
- 13.Eklind S, Mallard C, Arvidsson P, Hagberg H. Lipopolysaccharide induces both a primary and a secondary phase of sensitization in the developing rat brain. Pediatric research. 2005;58:112–6. doi: 10.1203/01.PDR.0000163513.03619.8D. [DOI] [PubMed] [Google Scholar]
- 14.Vontell R, Supramaniam V, Thornton C, et al. Toll-Like Receptor 3 Expression in Glia and Neurons Alters in Response to White Matter Injury in Preterm Infants. Developmental neuroscience. 2013 doi: 10.1159/000346158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Stridh L, Ek CJ, Wang X, Nilsson H, Mallard C. Regulation of Toll-Like Receptors in the Choroid Plexus in the Immature Brain After Systemic Inflammatory Stimuli. Translational stroke research. 2013;4:220–7. doi: 10.1007/s12975-012-0248-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Fleiss B, Gressens P. Tertiary mechanisms of brain damage: a new hope for treatment of cerebral palsy? Lancet neurology. 2012;11:556–66. doi: 10.1016/S1474-4422(12)70058-3. [DOI] [PubMed] [Google Scholar]
- 17.Gucuyener K, Atalay Y, Aral YZ, Hasanoglu A, Turkyilmaz C, Biberoglu G. Excitatory amino acids and taurine levels in cerebrospinal fluid of hypoxic ischemic encephalopathy in newborn. Clin Neurol Neurosurg. 1999;101:171–4. doi: 10.1016/s0303-8467(99)00035-9. [DOI] [PubMed] [Google Scholar]
- 18.Jensen FE. Developmental factors regulating susceptibility to perinatal brain injury and seizures. Current Opinion in Pediatrics. 2006;18:628–33. doi: 10.1097/MOP.0b013e328010c536. [DOI] [PubMed] [Google Scholar]
- 19.Buonocore G, Perrone S, Bracci R. Free radicals and brain damage in the newborn. Biology of the Neonate. 2001;79:180–6. doi: 10.1159/000047088. [DOI] [PubMed] [Google Scholar]
- 20.Hagberg H, Gressens P, Mallard C. Inflammation during fetal and neonatal life: implications for neurologic and neuropsychiatric disease in children and adults. Annals of Neurology. 2012;71:444–57. doi: 10.1002/ana.22620. [DOI] [PubMed] [Google Scholar]
- 21.Chavez-Valdez R, Martin LJ, Northington FJ. Programmed Necrosis: A Prominent Mechanism of Cell Death following Neonatal Brain Injury. Neurology Research International. 2012;2012:257563. doi: 10.1155/2012/257563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ikonomidou C, Turski L. Why did NMDA receptor antagonists fail clinical trials for stroke and traumatic brain injury? Lancet Neurology. 2002;1:383–6. doi: 10.1016/s1474-4422(02)00164-3. [DOI] [PubMed] [Google Scholar]
- 23.Costantine MM, Drever N. Antenatal exposure to magnesium sulfate and neuroprotection in preterm infants. Obstetrics and Gynecology Clinics of North America. 2011;38:351–66. xi. doi: 10.1016/j.ogc.2011.02.019. [DOI] [PubMed] [Google Scholar]
- 24.Wilhelm W, Hammadeh ME, White PF, Georg T, Fleser R, Biedler A. General anesthesia versus monitored anesthesia care with remifentanil for assisted reproductive technologies: effect on pregnancy rate. Journal of Clinical Anesthesia. 2002;14:1–5. doi: 10.1016/s0952-8180(01)00331-2. [DOI] [PubMed] [Google Scholar]
- 25.Ma D, Lim T, Xu J, et al. Xenon preconditioning protects against renal ischemic-reperfusion injury via HIF-1alpha activation. J Am Soc Nephrol. 2009;20:713–20. doi: 10.1681/ASN.2008070712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yang T, Zhuang L, Rei Fidalgo AM, et al. Xenon and sevoflurane provide analgesia during labor and fetal brain protection in a perinatal rat model of hypoxia-ischemia. PLoS One. 2012;7:e37020. doi: 10.1371/journal.pone.0037020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Thoresen M, Hobbs CE, Wood T, Chakkarapani E, Dingley J. Cooling combined with immediate or delayed xenon inhalation provides equivalent long-term neuroprotection after neonatal hypoxiaischemia. Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism. 2009;29:707–14. doi: 10.1038/jcbfm.2008.163. [DOI] [PubMed] [Google Scholar]
- 28.Palmer C, Towfighi J, Roberts RL, Heitjan DF. Allopurinol administered after inducing hypoxia-ischemia reduces brain injury in 7-day-old rats. Pediatric Research. 1993;33:405–11. doi: 10.1203/00006450-199304000-00018. [DOI] [PubMed] [Google Scholar]
- 29.Benders MJ, Bos AF, Rademaker CM, et al. Early postnatal allopurinol does not improve short term outcome after severe birth asphyxia. Archives of Disease in Childhood Fetal and Neonatal Edition. 2006;91:F163–5. doi: 10.1136/adc.2005.086652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kaandorp JJ, Benders MJ, Rademaker CM, et al. Antenatal allopurinol for reduction of birth asphyxia induced brain damage (ALLO-Trial); a randomized double blind placebo controlled multicenter study. BMC pregnancy and childbirth. 2010;10:8. doi: 10.1186/1471-2393-10-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Buonocore G, Groenendaal F. Anti-oxidant strategies. Seminars in fetal & neonatal medicine. 2007;12:287–95. doi: 10.1016/j.siny.2007.01.020. [DOI] [PubMed] [Google Scholar]
- 32.Yu L, Derrick M, Ji H, et al. Neuronal nitric oxide synthase inhibition prevents cerebral palsy following hypoxia-ischemia in fetal rabbits: comparison between JI-8 and 7-nitroindazole. Developmental Neuroscience. 2011;33:312–9. doi: 10.1159/000327244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Loren DJ, Seeram NP, Schulman RN, Holtzman DM. Maternal dietary supplementation with pomegranate juice is neuroprotective in an animal model of neonatal hypoxic-ischemic brain injury. Pediatric Research. 2005;57:858–64. doi: 10.1203/01.PDR.0000157722.07810.15. [DOI] [PubMed] [Google Scholar]
- 34.West T, Atzeva M, Holtzman DM. Pomegranate polyphenols and resveratrol protect the neonatal brain against hypoxic-ischemic injury. Developmental Neuroscience. 2007;29:363–72. doi: 10.1159/000105477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Zhang W, Hu X, Yang W, Gao Y, Chen J. Omega-3 polyunsaturated fatty acid supplementation confers long-term neuroprotection against neonatal hypoxic-ischemic brain injury through anti-inflammatory actions. Stroke. 2010;41:2341–7. doi: 10.1161/STROKEAHA.110.586081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ramanantsoa N, Fleiss B, Bouslama M, et al. Bench to Cribside: the Path for Developing a Neuroprotectant. Translational Stroke Research. 2013;4:258–77. doi: 10.1007/s12975-012-0233-2. [DOI] [PubMed] [Google Scholar]
- 37.Welin AK, Svedin P, Lapatto R, et al. Melatonin reduces inflammation and cell death in white matter in the mid-gestation fetal sheep following umbilical cord occlusion. Pediatric Research. 2007;61:153–8. doi: 10.1203/01.pdr.0000252546.20451.1a. [DOI] [PubMed] [Google Scholar]
- 38.Villapol S, Fau S, Renolleau S, Biran V, Charriaut-Marlangue C, Baud O. Melatonin promotes myelination by decreasing white matter inflammation after neonatal stroke. Pediatric Research. 2011;69:51–5. doi: 10.1203/PDR.0b013e3181fcb40b. [DOI] [PubMed] [Google Scholar]
- 39.Robertson NJ, Faulkner S, Fleiss B, et al. Melatonin augments hypothermic neuroprotection in a perinatal asphyxia model. Brain. 2013;136:90–105. doi: 10.1093/brain/aws285. [DOI] [PubMed] [Google Scholar]
- 40.Yu X, Shacka JJ, Eells JB, et al. Erythropoietin receptor signalling is required for normal brain development. Development. 2002;129:505–16. doi: 10.1242/dev.129.2.505. [DOI] [PubMed] [Google Scholar]
- 41.Mazur M, Miller RH, Robinson S. Postnatal erythropoietin treatment mitigates neural cell loss after systemic prenatal hypoxic-ischemic injury. Journal of neurosurgery Pediatrics. 2010;6:206–21. doi: 10.3171/2010.5.PEDS1032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Digicaylioglu M, Lipton SA. Erythropoietin-mediated neuroprotection involves cross-talk between Jak2 and NF-kappaB signalling cascades. Nature. 2001;412:641–7. doi: 10.1038/35088074. [DOI] [PubMed] [Google Scholar]
- 43.Sun Y, Calvert JW, Zhang JH. Neonatal hypoxia/ischemia is associated with decreased inflammatory mediators after erythropoietin administration. Stroke. 2005;36:1672–8. doi: 10.1161/01.STR.0000173406.04891.8c. [DOI] [PubMed] [Google Scholar]
- 44.Juul SE, Beyer RP, Bammler TK, McPherson RJ, Wilkerson J, Farin FM. Microarray analysis of high-dose recombinant erythropoietin treatment of unilateral brain injury in neonatal mouse hippocampus. Pediatr Res. 2009;65:485–92. doi: 10.1203/PDR.0b013e31819d90c8. [DOI] [PubMed] [Google Scholar]
- 45.Zacharias R, Schmidt M, Kny J, et al. Dose-dependent effects of erythropoietin in propofol anesthetized neonatal rats. Brain Res. 2010;1343:14–9. doi: 10.1016/j.brainres.2010.04.081. [DOI] [PubMed] [Google Scholar]
- 46.Kumral A, Tugyan K, Gonenc S, et al. Protective effects of erythropoietin against ethanol-induced apoptotic neurodegenaration and oxidative stress in the developing C57BL/6 mouse brain. Brain Res Dev Brain Res. 2005;160:146–56. doi: 10.1016/j.devbrainres.2005.08.006. [DOI] [PubMed] [Google Scholar]
- 47.Chattopadhyay A, Choudhury TD, Bandyopadhyay D, Datta AG. Protective effect of erythropoietin on the oxidative damage of erythrocyte membrane by hydroxyl radical. Biochem Pharmacol. 2000;59:419–25. doi: 10.1016/s0006-2952(99)00277-4. [DOI] [PubMed] [Google Scholar]
- 48.Iwai M, Cao G, Yin W, Stetler RA, Liu J, Chen J. Erythropoietin promotes neuronal replacement through revascularization and neurogenesis after neonatal hypoxia/ischemia in rats. Stroke. 2007;38:2795–803. doi: 10.1161/STROKEAHA.107.483008. [DOI] [PubMed] [Google Scholar]
- 49.Osredkar D, Sall JW, Bickler PE, Ferriero DM. Erythropoietin promotes hippocampal neurogenesis in in vitro models of neonatal stroke. Neurobiol Dis. 2010;38:259–65. doi: 10.1016/j.nbd.2010.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kellert BA, McPherson RJ, Juul SE. A comparison of high-dose recombinant erythropoietin treatment regimens in brain-injured neonatal rats. Pediatr Res. 2007;61:451–5. doi: 10.1203/pdr.0b013e3180332cec. [DOI] [PubMed] [Google Scholar]
- 51.Gonzalez FF, Abel R, Almli CR, Mu D, Wendland M, Ferriero DM. Erythropoietin sustains cognitive function and brain volume after neonatal stroke. Dev Neurosci. 2009;31:403–11. doi: 10.1159/000232558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Demers EJ, McPherson RJ, Juul SE. Erythropoietin protects dopaminergic neurons and improves neurobehavioral outcomes in juvenile rats after neonatal hypoxia-ischemia. Pediatric Research. 2005;58:297–301. doi: 10.1203/01.PDR.0000169971.64558.5A. [DOI] [PubMed] [Google Scholar]
- 53.Iwai M, Stetler RA, Xing J, et al. Enhanced oligodendrogenesis and recovery of neurological function by erythropoietin after neonatal hypoxic/ischemic brain injury. Stroke. 2010;41:1032–7. doi: 10.1161/STROKEAHA.109.570325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Reitmeir R, Kilic E, Kilic U, et al. Post-acute delivery of erythropoietin induces stroke recovery by promoting perilesional tissue remodelling and contralesional pyramidal tract plasticity. Brain : a journal of neurology. 2011;134:84–99. doi: 10.1093/brain/awq344. [DOI] [PubMed] [Google Scholar]
- 55.Juul S. Erythropoietin in anemia of prematurity. The journal of maternal-fetal & neonatal medicine : the official journal of the European Association of Perinatal Medicine, the Federation of Asia and Oceania Perinatal Societies, the International Society of Perinatal Obstetricians. 2012;25:80–4. doi: 10.3109/14767058.2012.716987. [DOI] [PubMed] [Google Scholar]
- 56.van der Kooij MA, Groenendaal F, Kavelaars A, Heijnen CJ, van Bel F. Neuroprotective properties and mechanisms of erythropoietin in in vitro and in vivo experimental models for hypoxia/ischemia. Brain Res Rev. 2008;59:22–33. doi: 10.1016/j.brainresrev.2008.04.007. [DOI] [PubMed] [Google Scholar]
- 57.Juul SE, McPherson RJ, Bauer LA, Ledbetter KJ, Gleason CA, Mayock DE. A phase I/II trial of highdose erythropoietin in extremely low birth weight infants: pharmacokinetics and safety. Pediatrics. 2008;122:383–91. doi: 10.1542/peds.2007-2711. [DOI] [PubMed] [Google Scholar]
- 58.Fauchere JC, Dame C, Vonthein R, et al. An approach to using recombinant erythropoietin for neuroprotection in very preterm infants. Pediatrics. 2008;122:375–82. doi: 10.1542/peds.2007-2591. [DOI] [PubMed] [Google Scholar]
- 59.Wu YW, Bauer LA, Ballard RA, et al. Erythropoietin for neuroprotection in neonatal encephalopathy: safety and pharmacokinetics. Pediatrics. 2012;130:683–91. doi: 10.1542/peds.2012-0498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Almeida RD, Manadas BJ, Melo CV, et al. Neuroprotection by BDNF against glutamate-induced apoptotic cell death is mediated by ERK and PI3-kinase pathways. Cell death and differentiation. 2005;12:1329–43. doi: 10.1038/sj.cdd.4401662. [DOI] [PubMed] [Google Scholar]
- 61.Feng Y, Rhodes PG, Bhatt AJ. Neuroprotective effects of vascular endothelial growth factor following hypoxic ischemic brain injury in neonatal rats. Pediatric research. 2008;64:370–4. doi: 10.1203/PDR.0b013e318180ebe6. [DOI] [PubMed] [Google Scholar]
- 62.Xiao BG, Lu CZ, Link H. Cell biology and clinical promise of G-CSF: immunomodulation and neuroprotection. J Cell Mol Med. 2007;11:1272–90. doi: 10.1111/j.1582-4934.2007.00101.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Fathali N, Lekic T, Zhang JH, Tang J. Long-term evaluation of granulocyte-colony stimulating factor on hypoxic-ischemic brain damage in infant rats. Intensive Care Med. 2010;36:1602–8. doi: 10.1007/s00134-010-1913-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Schabitz WR, Laage R, Vogt G, et al. AXIS: a trial of intravenous granulocyte colony-stimulating factor in acute ischemic stroke. Stroke; a journal of cerebral circulation. 2010;41:2545–51. doi: 10.1161/STROKEAHA.110.579508. [DOI] [PubMed] [Google Scholar]
- 65.Popken GJ, Hodge RD, Ye P, et al. In vivo effects of insulin-like growth factor-I (IGF-I) on prenatal and early postnatal development of the central nervous system. Eur J Neurosci. 2004;19:2056–68. doi: 10.1111/j.0953-816X.2004.03320.x. [DOI] [PubMed] [Google Scholar]
- 66.Kannan S, Dai H, Navath RS, et al. Dendrimer-based postnatal therapy for neuroinflammation and cerebral palsy in a rabbit model. Science translational medicine. 2012;4:130ra46. doi: 10.1126/scitranslmed.3003162. [DOI] [PMC free article] [PubMed] [Google Scholar]