

Abstract
Advanced pancreatic cancer still has a poor prognosis, even with the approval of several drugs, such as gemcitabine. Therefore, developing effective and safe antitumor agents is urgently needed. 6-Shogaol, a phenol extracted from ginger, has been linked to suppression of proliferation and survival of cancer with different mechanisms. In the present study, we investigated whether 6-shogaol could suppress pancreatic cancer progress and potentiate pancreatic cancer to gemcitabine treatment in vitro and in vivo. We found that 6-shogaol prevented the activation of toll like receptor 4 (TLR4)/NF-κB signaling. The modulation of NF-κB signaling by 6-shogaol was ascertained by electrophoretic mobility shift assay and western blot analysis. The suppression of NF-κB signaling and key cell survival regulators including COX-2, cyclinD1, survivin, cIAP-1, XIAP, Bcl-2, and MMP-9 brought the anti-proliferation effects in pancreatic cancer cells and sensitized them to gemcitabine treatment. Furthermore, in a pancreatic cancer xenograft model, we found a decreased proliferation index (Ki-67) and increased apoptosis by TUNEL staining in 6-shogaol treated tumors. It was also shown that 6-shogaol combined with gemcitabine treatment was more effective than drug alone, consistent with the downregulation of NF-κB activity along with its target genes COX-2, cyclinD1, survivin, cIAP-1, and XIAP. Overall, our results suggest that 6-shogaol can inhibit the growth of human pancreatic tumors and sensitize them to gemcitabine by suppressing of TLR4/NF-κB-mediated inflammatory pathways linked to tumorigenesis.
Electronic supplementary material
The online version of this article (doi:10.1208/s12248-013-9558-3) contains supplementary material, which is available to authorized users.
Key words: chemosensitization, gemcitabine, pancreatic cancer, TLR4/NF-κB, 6-shogaol
INTRODUCTION
Pancreatic cancer is a highly aggressive malignant disease with a 5-year survival rate of less than 5%. According to the statistics published by the National Cancer Institute, 43,920 new pancreatic cancer cases were reported and 37,390 individuals died of pancreatic cancer in the year 2012 in USA (http://www.cancer.gov/cancertopics/types/pancreatic). Conventional therapeutic strategies have yielded limited success with a great proportion of dismal outcomes in pancreatic cancer treatment. The limitations necessitate the development of less toxic drugs as well as adjuvants that can selectively sensitize pancreatic tumor cells to chemotherapy drugs.
Toll like receptors (TLRs) are pattern recognition receptors, playing a key role in inflammatory response by recognizing pathogen-associated molecular patterns (PAMPs) and endogenous damage-associated molecular patterns (DAMPs). Mounting evidence has suggested the reflexive relationship between inflammation and cancer, including pancreatic cancer (1,2). Specifically, lipopolysaccharide (LPS) from intestinal gram-negative bacteria can promote TLR4/NF-κB activation in pancreatic cancer cells which connects inflammation with cancer progression (3). In fact, clinical studies found that TLR4 and NF-κB were overexpressed in pancreatic adenocarcinoma compared to surrounding tissues. Patients with an absence of TLR4 expression in tumor tissues survived significantly longer than those with TLR4 expression (4). Furthermore, NF-κB, an essential downstream component of TLR4 signal pathway, has been shown to be involved in pancreatic cancer progression and chemoresistance. All these studies suggest that TLR4/NF-κB is a potential therapeutic target for the treatment of pancreatic cancer.
Although gemcitabine is an agent approved by the US Food and Drug Administration (FDA) to treat pancreatic cancer, the outcome is not fully satisfactory mainly due to toxicity to normal tissues and acquisition of resistance by tumor tissues. Multiple lines of evidence have suggested that activation of NF-κB and its target genes could increase cell survival and contribute to cellular resistance to gemcitabine. Therefore, NF-κB inhibitors could reduce the chemoresistance to gemcitabine and perhaps be adopted in combination with gemcitabine as a novel therapeutic treatment regimen for pancreatic cancer (5).
6-Shogaol is a phenolic alkanone derived from ginger. As one of the most active ingredients of ginger, 6-shogaol has been reported to exert potent antitumor activity in a variety of human cancer cell lines in vitro (6–8). Previous studies have demonstrated that 6-shogaol induced apoptosis of hepatocellular carcinoma and cervical cancer via endoplasmic reticulum stress (9,10). Additionally, increasing evidences have demonstrated that this plant polyphenol is a potent blocker of the NF-κB pathway (6,11,12). These studies suggest that 6-shogaol can be useful as an adjuvant to potentiate the effects of gemcitabine in pancreatic cancer.
In the present study, we investigated the effects of 6-shogaol on the growth of human pancreatic cancer cells. We also tested whether 6-shogaol could sensitize human pancreatic tumors to gemcitabine in vitro and in vivo. We demonstrated that 6-shogaol inhibited the proliferation of pancreatic cancer cells, enhanced gemcitabine-induced apoptosis, and potentiated the antitumor activity of gemcitabine against subcutaneously implanted human pancreatic tumors through the downregulation of TLR4/NF-κB signaling and NF-κB-regulated gene products.
MATERIALS AND METHODS
Cell Lines and Cell Culture
The human pancreatic cancer cell lines PANC-1 and BxPC-3 were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). PANC-1 cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal bovine serum (Gibco) and BxPC-3 cells were cultured in RPMI 1640 containing 10% FBS. All cell lines were incubated at 37°C in a humidified atmosphere of 5% CO2.
Materials
Antibodies against TLR4, COX-2, and β-actin were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Antibodies against cIAP-1, XIAP, survivin, laminB, phosphor-NF-κB p65, NF-κB p65, MMP-9, cyclinD1, Bcl-2, pro-caspase 3, cleaved-caspase 3, and cleaved-poly-ADP-ribose polymerase (PARP) were from Cell Signaling Technology (Beverly, MA). 6-Shogaol (>95% pure) was extracted from ginger using a supercritical fluid extraction method. We calculated molarities according to the amount of active agent in the material and dissolved 6-shogaol in dimethylsulfoxide (DMSO) to make 100 mM stock solution as described previously (9). Gemcitabine (Gemzar) was purchased from Eli Lilly (Indianapolis, USA) and stored at 4°C, which was dissolved in sterile phosphate buffered solution (PBS) on the day of use.
Animals
All procedures in animal experiments were approved by the Institutional Animal Care and Use Committee (IACUC) of the China Pharmaceutical University (Nanjing, China). Six-week-old pathogen-free male BALB/c immunodeficient nude mice were obtained from Shanghai Slac Laboratory Animal Limited Company (Shanghai, China). Animals were housed in sterile filter-capped microisolator cages and provided with sterilized diet and water.
Proliferation Assay
First, we examined the anti-proliferation of 6-shogaol. In brief, PANC-1 and BxPC-3 cells (3,000 per well) were seeded in 96-well plates and supplemented with fresh medium containing different concentrations of 6-shogaol (0, 15, 20, and 25 μM) on the next day. After incubating for 24, 48, and 72 h, 3−(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) solution (5 mg/mL) was added to each well. After incubation at 37°C for an additional 4 h, MTT formazan formed by viable cells was dissolved in DMSO, and absorbance was measured at 570 nm by the Universal Microplate Reader (BIO-TEK instruments, Inc., Vermont, MA). To further examine chemosensitization of 6-shogaol, cells were plated as described above and cultured with medium containing 6-shogaol (10–20 μM) for 24 h. Then, these cells were subsequently incubated with gemcitabine (0.1–10 μM) for an additional 72 h. Cells were exposed to 6-shogaol for 96 h and gemcitabine for 72 h as a single-agent treatment control.
Drug Interaction Analysis
Drug interaction between 6-shogaol and gemcitabine was assessed using the combination index (CI), where CI < 1, CI = 1, and CI > 1 indicate synergistic, additive, and antagonistic effects, respectively (13). The CI value was calculated as follows:
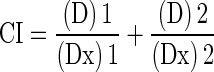 |
where (Dx)1 and (Dx)2 are the 50% inhibitory concentration of cell growth (IC50) of 6-shogaol and gemcitabine, respectively, and (D)1 and (D)2 are the drug concentrations in combination treatments that also inhibit cell growth by 50%. The IC50 was calculated by non-linear least squares curve fitting.
Apoptosis Assay
Cell death detection enzyme-linked immunosorbent assay (ELISA) kit (Roche, Mannheim, Germany) was used to detect apoptosis in PANC-1 and BxPC-3 cells according to the manufacturer’s protocols.
Clonogenic Assay
PANC-1 and BxPC-3 cells were plated (200,000 per well) in a six-well plate and incubated overnight at 37°C. After different concentrations of 6-shogaol (0, 15, and 20 μM) treatment for 24 h, cells were trypsinized and viable cells were counted using trypan blue exclusion method. Then 1,000 viable cells were plated in 100-mm Petri dishes to determine the plating efficiency as well as assess the effects of treatment on clonogenic survival. Cells were incubated for 14 days in a humidified incubator containing 5% CO2 in air at 37°C. After 4% paraformaldehyde fixing for 15 min, colonies were stained with 2% crystal violet and then counted.
Protein Extraction and Western Blot Analysis
Protein samples were prepared by incubation of cells or tumor tissues in radioimmunoprecipitation assay (RIPA) buffer containing protease inhibitor cocktail and phosphatase inhibitor cocktail (Roche, Mannheim, Germany). The concentration of total proteins was determined using enhanced BCA (bicinchonininc acid) protein assay reagent (Beyotime, Jiangsu, China). Protein samples were separated by 10–12% SDS-PAGE and transferred to nitrocellulose (NC) membranes. Membranes were blocked with 5% nonfat dried milk in 1× Tris-buffered saline (TBS) containing 0.05% Tween 20 under room temperature for 2 h and then incubated with primary antibodies at 4°C overnight. Protein bands were detected by incubation with horseradish peroxidase-conjugated secondary antibodies (ZSGB-BIO, Beijing, China), and visualized by enhanced chemiluminescence reagent (Amersham, USA).
Electrophoretic Mobility Shift Assay
Electrophoretic mobility shift assays (EMSA) was used to assess NF-κB activation. Briefly, nuclear extracts prepared from treated cells and tumor samples with Nuclear and Cytoplasmic Extraction Reagents (Pierce, IL, USA) were incubated with biotin labeled double-stranded NF-κB oligonucleotide (5′-TTGTTACAAGGGACTTTCCGCTGGGGACTTTCCAGGGGGAGGCGTGG-3′; underline indicates NF-κB-binding sites) for 30 min at 37°C (14), and EMSA was performed followed the instructions of LightShift chemiluminescent EMSA kit (Pierce).
Native gel PAGE
To perform Native PAGE analysis, native proteins were prepared using ice-cold isotonic buffer [50 mM Tris (pH 7.4), 150 mM NaCl, 1% NP-40, 0.25% sodium deoxycholate] containing protease inhibitor cocktail and phosphatase inhibitor cocktail. An amount of 50 μg of protein was loaded onto 6% SDS-free, PAGE gels. Proteins were transferred to NC membranes and immunoblotted with TLR4 antibody as described in the western blotting part.
Plasmid and Small Interference RNA Transient Transfection
The p65 plasmid was purchased from Sino Biological Inc. (Beijing, China). The TLR4 small interference RNA (siRNA) and the scrambled siRNA were purchased from Santa Cruz Biotechnology. Transfection of DNA (plasmids) and RNA (siRNA) was performed using Lipofectamine™ 2000 (Invitrogen) according to the manufacturer’s instructions.
Xenograft Animal Experiment
After an acclimatization period of 1 week, mice (n = 24) were injected with PANC-1 cells (1 × 107) with matrigel (100 μL) to the right flank of each mice. When tumor volume reached 100 mm3, mice were randomly divided into the following four treatment groups with six animals each: (a) Vehicle group; (b) 6S group (50 mg/kg 6-shogaol once daily by intraperitoneal [i.p.] injection); (c) Gem group (25 mg/kg gemcitabine twice weekly by i.p. injection); (d) 6S + Gem group (50 mg/kg 6-shogaol once daily by i.p. injection and 25 mg/kg gemcitabine twice weekly by i.p. injection). Tumor volume and body weight were measured every other day. The volume was calculated using the formula V = 0.52 a b2, with a as the larger diameter and b as the smaller diameter. Therapy was continued for 4 weeks and mice were sacrificed at 24 h after the last administration of compound. Tumor samples were excised and weighed. Half of the tumor tissues were formalin fixed. The other half was snap frozen in liquid nitrogen and stored at −80°C.
Immunohistochemistry Staining
Tumor samples were embedded in paraffin and sectioned for immunohistochemical staining for Ki-67 and terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) and observed with a microscope (Nikon, Japan). Results were expressed as percentage of Ki-67/TUNEL-positive cells per ×40 magnification. A total of ten ×40 fields were examined from three tumors of each of the treatment groups.
Statistical Analysis
All data was represented as mean ± standard deviation (SD) for at least three independent experiments and representative examples are shown. Statistical analysis of multiple-group comparisons was performed by one-way analysis of variance (ANOVA). Comparisons between two groups were analyzed using Student tests. A p value < 0.05 was considered statistically significant.
RESULTS
6-Shogaol Inhibited Proliferation and Induced Apoptosis of Pancreatic Cancer Cells
To examine the anti-proliferative effects of 6-shogaol on human pancreatic cancer cells, we used PANC-1 cells which exhibit K-Ras and p53 mutations, and BxPC-3 cells with wild-type K-Ras. The viability of cells treated with 6-shogaol at various concentrations (0, 15, 20, and 25 μM) for 24, 48, and 72 h was determined by MTT assay. As shown in Fig. 1a, 6-shogaol inhibited proliferation of pancreatic cancer cells irrespective of their genetic background in a dose- and time-dependent manner. To further evaluate the anti-proliferative activity of 6-shogaol, colony-formation assay was performed. Treatment with 6-shogaol (15 and 20 μM) for 24 h resulted in reduction of colony number and colony size in PANC-1 and BxPC-3 cells compared to control (Fig. 1b). Furthermore, to determine the mechanism of the observed proliferation inhibition by 6-shogaol treatment, apoptosis was evaluated using histone-DNA ELISA assay. As shown in Fig. 1c, there was a significant increase in the number of apoptotic cells when cells were exposed to 6-shogaol. Cleaved active components of caspase 3 and its downstream substrate PARP increased dose-dependently upon 6-shogaol treatment (Fig. 1d). All the results indicated that the anti-proliferative effects of 6-shogaol were correlated to cell apoptosis.
Fig. 1.
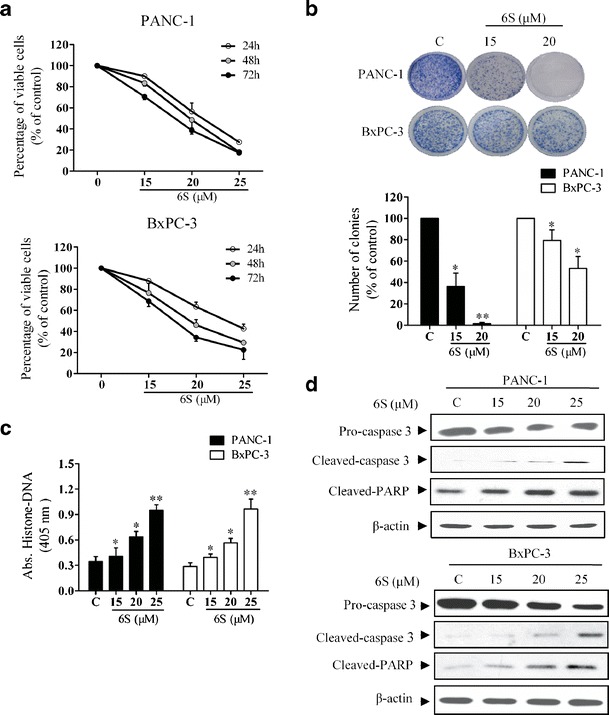
6-Shogaol reduces the survival of pancreatic cancer cells in vitro. a 6-Shogaol inhibited proliferation of pancreatic cancer cells in a dose- and time-dependent manner. PANC-1 and BxPC-3 cells (3,000 per well) were seeded into a 96-well plate, incubated for 24 h and then treated with 6-shogaol (0, 15, 20, and 25 μM) for 24, 48, and 72 h. Cell viability was determined by MTT assay. b 6-Shogaol inhibited pancreatic cancer cells colony-formation ability. PANC-1 and BxPC-3 cells were plated (200,000 per well) in a six-well plate, incubated for 24 h and then treated with 6-shogaol (0, 15, and 20 μM) for 24 h. Then 1,000 viable cells were plated in 100 mm Petri dish and cells were incubated for 14 days. Colonies were stained with 2% crystal violet and the images of the representative colonies shown were taken using camera (Nikon). Statistical analysis showed the percentage of cells that formed colonies relative to the controls (control cells = 100%). c Effects of 6-shogaol on PANC-1 and BxPC-3 apoptosis. PANC-1 and BxPC-3 cells were seeded into a 96-well plate, incubated for 24 h and then treated with 6-shogaol (0, 15, 20, and 25 μM) for 24 h. Histone-DNA ELISA was performed. d Western blot analysis of pro-caspase 3, cleaved-caspase 3, and cleaved-PARP in pancreatic cancer cells. PANC-1 and BxPC-3 cells were plated (200,000 per well) in a six-well plate, incubated for 24 h and then treated with 6-shogaol (0, 15, 20, and 25 μM) for 24 h. Protein samples were prepared and western blot analysis was performed. *p < 0.05 or **p < 0.01, treatment vs control. C control, 6S 6-shogaol
6-Shogaol Inhibited TLR4/NF-κB Signaling in Pancreatic Cancer Cells
To investigate the mechanism that 6-shogaol induced apoptosis in pancreatic cancer cells, NF-κB activity was evaluated given that the signal is constitutive activated in most pancreatic cancer (15). As shown in Fig. 2a, 6-shogaol treatment resulted in a decreased p65 phosphorylation. We then checked the subcellular localization of p65, as shown in Fig. 2b, 6-shogaol sequestered p65 in the cytoplasm in a concentration-dependent manner. These observations were further supported by the finding that 6-shogaol treatment attenuated NF-κB-DNA-binding activity as assessed by EMSA (Fig. 2c). Subsequently, the NF-κB-regulated gene products were analyzed by PCR and western blot. We found that 6-shogaol suppressed the constitutive expression of anti-apoptotic proteins (cIAP-1, survivin, XIAP, and Bcl-2), proliferative proteins (COX-2 and cyclin D1) and metastatic protein (MMP-9) in a concentration-dependent manner in PANC-1 cells (supplemental 1 and Fig. 2d). These findings indicated that 6-shogaol induced apoptosis in association with inhibition of NF-κB and its downstream effectors.
Fig. 2.
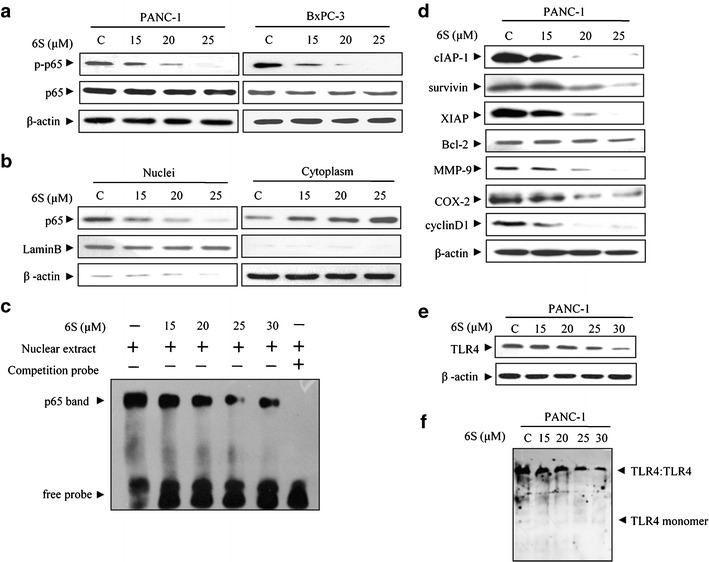
6-Shogaol inhibits TLR4/NF-κB signaling in pancreatic cancer cells. a 6-Shogaol decreased phosphorylation of p65 Ser 536. PANC-1 and BxPC-3 cells were treated with 0, 15, 20, and 25 μM 6-shogaol for 24 h and cell lysates were prepared and subjected to western blot analysis using antibodies against p-p65 (ser536), p65 and β-actin. b 6-Shogaol inhibited NF-κB nuclear translocation. After PANC-1 cells were treated with 6-shogaol (0, 15, 20, and 25 μM) for 24 h, the cytoplasmic and nuclear fractions of the cells were analyzed by western blotting for the p65 protein, with β-actin and lamin B as cytoplasmic and nuclear loading controls, respectively. c 6-Shogaol decreased NF-κB DNA-binding activity. Cells were incubated with 6-shogaol (0, 15, 20, 25, and 30 μM) for 24 h, and DNA binding was determined in nuclear extracts using EMSA. d 6-Shogaol suppressed the constitutive expression of NF-κB-regulated gene products involved in anti-apoptosis, proliferation, and metastasis. The PANC-1 cells were treated with indicated concentrations of 6-shogaol for 24 h. The whole-cell lysates was prepared and western blot was performed. e 6-Shogaol had minor effects on the expression of TLR4 at the concentration of 15 and 20 μM, but at higher concentration (25 and 30 μM), 6-shogaol decreased the total TLR4 protein. PANC-1 cells were treated with 6-shogaol (15, 20, 25, and 30 μM) for 24 h. Total proteins were prepared and subjected to western blot analysis using antibody against TLR4 and β-actin. f 6-Shogaol inhibited TLR4 dimerization. PANC-1 cells were treated with 6-shogaol (0, 15, 20, 25, and 30 μM) for 3 h. Native proteins were prepared and subjected to Native PAGE analysis using antibody against TLR4. C control, 6S 6-shogaol
We next investigated which components in NF-κB signaling 6-shogaol affected. TLR4 protein can activate the NF-κB pathway and increase p65 Ser536 phosphorylation (16). Dimerization of TLR4 triggers a series of cascade to activate its downstream signaling which ultimately results in activation of NF-κB (17,18). TAK-242 (resatorvid), a small-molecule-specific inhibitor of TLR4 signaling, decreased the expression of cIAP-1, survivin, XIAP, and p-p65, while LPS, a TLR4 ligand, increased the expression of these genes (supplemental 2). Both pancreatic cancer cell lines (PANC-1 and BxPC-3) expressed TLR4 (supplemental 3). Furthermore, 6-shogaol had minor effects on the expression of TLR4 at the concentration of 15 and 20 μM, but at higher concentration (25 and 30 μM), 6-shogaol decreased the total TLR4 protein (Fig. 2e). In addition, 6-shogaol inhibited the dimerization of TLR4 (Fig. 2f). Together, these findings demonstrated that TLR4 might be an important effective target for 6-shogaol in the treatment of pancreatic cancer.
6-Shogaol Sensitized Pancreatic Cancer Cells to Gemcitabine Treatment
Although gemcitabine was approved for treating pancreatic cancer by FDA, the outcome is still poor due to the formation of drug resistance, a process in which NF-κB regulation is involved (5). Since 6-shogaol can attenuate NF-κB activity, we next investigated whether 6-shogaol could augment cytotoxicity of gemcitabine in pancreatic cancer cells. The effects of gemcitabine on cell viability were determined by MTT assay (supplemental 4). Subsequent studies were undertaken to examine whether cells pretreated with 6-shogaol could be more sensitive to gemcitabine. Treatment with either 6-shogaol (15 μM, 96 h) or gemcitabine (1 μM, 72 h) alone resulted in a 25∼30% loss of viability of PANC-1 and BxPC-3 cells. However, pretreatment with 6-shogaol (15 μM for 24 h) followed by gemcitabine (1 μM, 72 h) treatment resulted in an approximately 55∼60% inhibition of proliferation in all cell types (Fig. 3a). This anti-proliferative activity was further confirmed by colony-formation assay (Fig. 3b). It was shown that pretreatment with 6-shogaol (15 μM for 24 h) followed by gemcitabine (1 μM, 72 h) treatment induced apoptosis in a greater proportion when compared to treatment with either drug alone (Fig. 3c). Both cleaved-caspase 3 and cleaved-PARP were also increased (Fig. 3d). Multiple-dose combinations of 6-shogaol (10–20 μM) and gemcitabine (0.1–10 μM) were also investigated. As shown in supplemental 5, the cell viability was distinctly decreased in all pancreatic cancer cell lines investigated on the combination of 6-shogaol and gemcitabine treatment. Moreover, to confirm synergism, the CI values were determined for combination treatment group, at concentration ratio of 2:1, 1:1, and 1:2. We found that cells pretreated with 6-shogaol showed synergistic loss of the cell viability when combined with gemcitabine (Table I, II). These findings indicated that 6-shogaol increased sensitivity toward gemcitabine in pancreatic cancer cells.
Fig. 3.
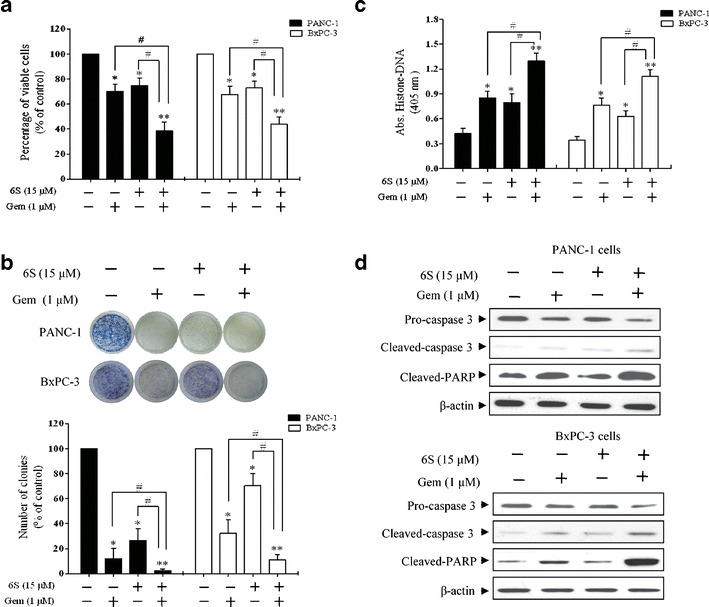
6-Shogaol pretreatment enhances the antitumor effects of gemcitabine. PANC-1 and BxPC-3 cells were pretreated with 6-shogaol (15 μM) for 24 h and then cells were incubated with gemcitabine (1 μM) for an additional 72 h for a, c and d. a MTT assay results showed that 6-shogaol potentiated gemcitabine-induced cytotoxicity. b The combination of 6-shogaol and gemcitabine was more effective in inhibiting colony formation. After 6-shogaol (15 μM) treatment for 24 h, PANC-1 and BxPC-3 cells were treated with gemcitabine for another 24 h and clonogenic assay was performed. Statistical analysis showed the percentage of cells that formed colonies relative to the controls (control cells = 100%). c Increased apoptosis by 6-shogaol pretreatment was determined using histone-DNA ELISA. d Western blot showed that 6-shogaol and gemcitabine together inhibited the expression of pro-caspase 3 and increased the expression of cleaved-caspase 3 and cleaved-PARP in PANC-1 and BxPC-3 cells. *p < 0.05, **p < 0.01, control vs treatment groups. # p < 0.05, ## p < 0.01, monotherapy (6S or Gem) vs combination (6S + Gem). 6S 6-shogaol, Gem gemcitabine
Table I.
IC50 of 6-shogaol (96 h) and Gemcitabine (72 h)
| IC50 (μM) | ||
|---|---|---|
| Cell lines | 6S | Gem |
| PANC-1 | 18.0 | 36.87 |
| BxPC-3 | 16.9 | 6.60 |
6S 6-shogaol, Gem gemcitabine
Table II.
CI Values at Concentration Ratio of 2:1, 1:1, and 1:2
| Cell lines | 6S/Gem | IC50(μM) | CI |
|---|---|---|---|
| PANC-1 | 2:1 | 13.04 | 0.601 |
| 1:1 | 14.40 | 0.595 | |
| 1:2 | 19.21 | 0.703 | |
| BxPC-3 | 2:1 | 9.772 | 0.879 |
| 1:1 | 9.116 | 0.960 | |
| 1:2 | 7.123 | 0.860 |
IC50-values (μM) for various dose combinations of 6-shogaol and gemcitabine
6S 6-shogaol, Gem gemcitabine
6-Shogaol Potentiated the Effects of Gemcitabine by Downregulating TLR4/NF-κB Signaling
Because 6-shogaol sensitized pancreatic cancer cells to gemcitabine, we investigated whether this property was dependent on inactivation of NF-κB. PANC-1 cells were exposed to 6-shogaol (15 μM) for 24 h followed by treatment with gemcitabine (1 μM) for another 72 h and nuclear extracts from these cells were subjected to EMSA. Consistent with previously published data (19), gemcitabine treatment induced DNA-binding ability of NF-κB. However, pretreatment with 6-shogaol inhibited gemcitabine-induced activation of NF-κB (Fig. 4a). These results were further confirmed by immunoblot of target proteins regulated by NF-κB in PANC-1 cells (Fig. 4b). We then determined whether overexpression of p65 or silencing of TLR4 could affect the treatment of 6-shogaol on pancreatic cancer cells. Firstly, the overexpression efficiency of p65 and specific suppression of TLR4 were confirmed by western blotting (Fig. 4c, d). After transfection, PANC-1 cells were pretreated with 6-shogaol (15 μM for 24 h) followed by gemcitabine (1 μM, 72 h) and cell viability was determined. Either overexpression of p65 or silence of TLR4 attenuated the 6-shogaol effects (Fig. 4e, f), indicating TLR4/NF-κB signaling might be the main functional target of 6-shogaol in pancreatic cancer cells.
Fig. 4.
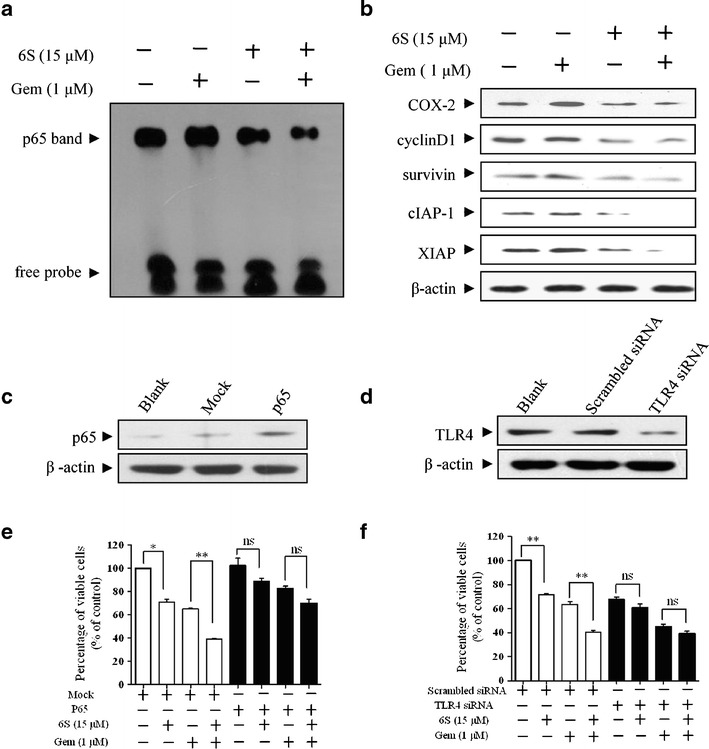
6-Shogaol enhances the antitumor effects of gemcitabine against the TLR4/NF-κB signaling in pancreatic cancer cells. a Detection of NF-κB DNA-binding activity using EMSA showed the inhibition of NF-κB by 6-shogaol. PANC-1 cells were pretreated with 6-shogaol (6S, 15 μM) for 24 h and then treated with gemcitabine (Gem, 1 μM) for 72 h. Nuclear extracts were prepared and subjected to EMSA. b Western blot showed that 6-shogaol and gemcitabine together inhibited the expression of NF-κB-dependent gene products. After the same treatment as (a), whole-cell lysates were obtained and subjected to western blot analysis using antibodies against cyclinD1, COX-2, survivin, cIAP-1, XIAP, and β-actin. c PANC-1 cells, either untransfected (Blank) or transfected with vector control (Mock) or p65 for 24 h, were assessed for p65 protein expression. d PANC-1 cells, either untransfected (Blank) or transfected with scrambled siRNA or TLR4 siRNA for 24 h, were assessed for TLR4 protein expression. e–f Overexpression of p65 or silence of TLR4 affected the treatment of 6-shogaol on pancreatic cancer cells. After overexpression of p65 or silence of TLR4 for 24 h, PANC-1 cells were pretreated with 6-shogaol (15 μM) for 24 h and then cells were incubated with gemcitabine (1 μM) for an additional 72 h and cell viability was determined. *p < 0.05, **p < 0.01, control vs 6S group or Gem group vs combination (6S + Gem) group. 6S 6-shogaol, Gem gemcitabine, ns no significant difference
Therapeutic Effects of Gemcitabine Were Enhanced by 6-Shogaol In Vivo
We further evaluated in vivo antitumor activity of 6-shogaol in a xenograft pancreatic tumor model. The experimental protocol is depicted in Fig. 5a. Tumor growth was significantly inhibited in mice injected with 6-shogaol (p < 0.05) or gemcitabine (p < 0.05) compared with the vehicle group, while the tumor growth of 6S + Gem group was slower than any single agent (p < 0.05) (Fig. 5b, c). As shown in Fig. 5d, treatment with 6-shogaol or gemcitabine alone caused 43% (p < 0.05) and 54% (p < 0.05) reduction in mean tumor weight respectively relative to vehicle group. Treatment of the combination of the two resulted in 71% decrease in tumor weight, a result which was superior to that observed with 6-shogaol (p < 0.05) or gemcitabine (p < 0.05) alone. The administration of 6-shogaol did not result in severe weight loss in mice (Fig. 5e). Moreover, pathologic changes in heart, liver, spleen, lung, kidney, and pancreas were not observed (supplemental 6), suggesting that 6-shogaol did not exert detrimental health effects within the parameters used in our study.
Fig. 5.
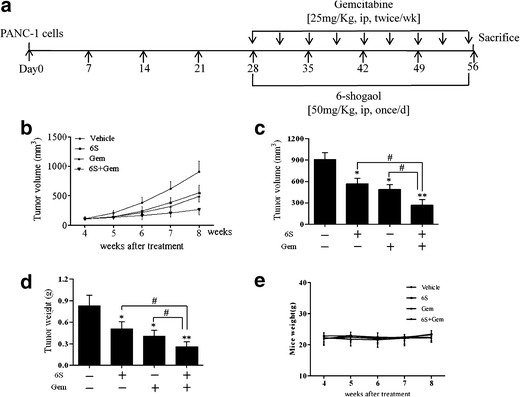
6-Shogaol potentiates the effects of gemcitabine in inhibiting the growth of pancreatic cancer in vivo. a Flow chart for in vivo experimental design and treatment schedule. b Average tumor volume of vehicle control mice and treatment group mice at various time points (n = 6). c Tumor volumes in mice measured on the last day of the experiment (n = 6). d Isolated tumor weight between different groups of mice (n = 6). e Body weight changes of mice during the treatment (n = 6). *p < 0.05, **p < 0.01, vehicle controls vs treatment groups. # p < 0.05, ## p < 0.01, monotherapy (6S or Gem) vs combination (6S + Gem). 6S 6-shogaol, Gem gemcitabine
6-Shogaol Exhibited Antitumor Activity in Xenografts of PANC-1 Cells by Downregulation of NF-κB Signaling
We then investigated whether treatment with 6-shogaol or the combination of 6-shogaol and gemcitabine could effectively target NF-κB in tumor tissues. Results showed that NF-κB activity was downregulated by 6-shogaol treatment, and gemcitabine-induced activation of NF-κB was attenuated when gemcitabine and 6-shogaol were given together (Fig. 6a). Expression of NF-κB-regulated gene products in tumor samples was also downregulated by 6-shogaol (Fig. 6b). All the results are evidence in favor of the hypothesis that inhibition of NF-κB signaling is one of the molecular mechanisms whereby 6-shogaol potentiates the antitumor activity of gemcitabine in vivo.
Fig. 6.
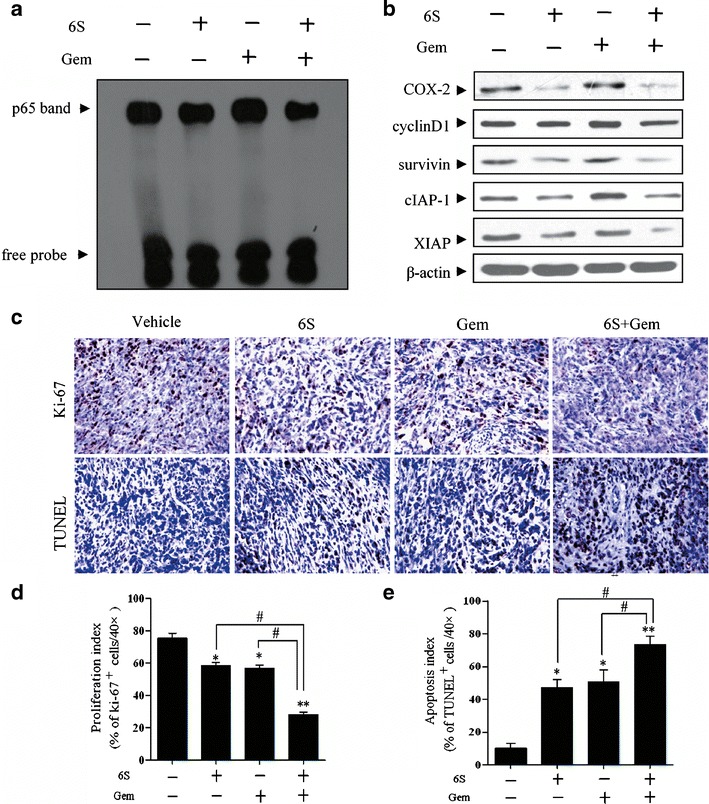
6-Shogaol potentiates the effects of gemcitabine though downregulation NF-κB and NF-κB-regulated gene products in vivo. a Detection of NF-κB DNA-binding activity by EMSA in xenografted tumor tissue samples showed the inhibition of NF-κB by 6-shogaol. b Western blot showed that combination treatment of 6-shogaol and gemcitabine inhibited the expression of NF-κB-dependent gene products in pancreatic tumor tissues. Tumor lysates from each group were performed western blot analysis using antibodies against cyclinD1, COX-2, survivin, cIAP-1, XIAP, and β-actin. c Immunohistochemical analysis of Ki-67 and TUNEL in tumor tissue sections from different treatment groups showed the decreased proliferation and increased apoptosis by either 6-shogaol alone or in combination with gemcitabine treatment. The photographs were taken at the magnification of ×20. d Quantification of Ki-67- or TUNEL-positive cells in pancreatic cancer tumors. A total of ten ×40 fields were examined from three tumors of each of the treatment groups. *p < 0.05, **p < 0.01, vehicle controls vs treatment groups. # p < 0.05, ## p < 0.01, monotherapy (6S or Gem) vs combination (6S + Gem). 6S 6-shogaol, Gem gemcitabine
Immunohistochemistry revealed significant reduction of Ki-67 positive cells in tumors derived from mice treated with 6-shogaol and gemcitabine. However, the combination of the two was most effective (p < 0.05). The TUNEL-positive cells were significantly increased in tumors from the combination group relative to single-agent-treated animals (Fig. 6c–e). Altogether, these data concurred with the decreased expression of genes involved in proliferation and survival within tumor tissues.
DISCUSSION
Pancreatic cancer remains one of the most challenging malignancies, partially due to the advanced disease stage at diagnosis and intrinsic drug resistance (20). Therefore, agents that inhibit proliferation, survival, angiogenesis, and overcome chemoresistance are highly desirable. In the present study, we investigated the antitumor activity of 6-shogaol, as well as its chemosensitization effects to gemcitabine against human pancreatic cancer.
6-Shogaol has an α, β-unsaturated carbonyl group which has been documented to react with biological nucleophiles such as sulfhydryl groups of cysteine residues by a Michael addition (21). Previous reports have shown that certain phytochemicals with α, β-unsaturated carbonyl group structural motif could disrupt the disulfide bonds, thus inhibiting the prerequisite step for TLR4 activation–homodimerization (22–24). Additionally, it was reported that in macrophage and monocytic cells, 6-shogaol could disrupt TLR4 activation (12,25). However, whether 6-shogaol functions similarly in a solid tumor is not known, and the corresponding in vivo data are lacking. Our results show that 6-shogaol both decreased the total TLR4 protein (Fig. 2e) and inhibited endogenous TLR4 dimerization (Fig. 2f).
Furthermore, multiple lines of evidence have suggested that NF-κB, the major transcription factor known to be activated by TLR4 signaling, plays important roles in pancreatic tumor progression, metastasis and chemoresistance (15,26,27). Therefore, we speculated that 6-shogaol might inhibit proliferation and induce apoptosis of pancreatic cancer cells through inhibition of TLR4/NF-κB signaling. In the present study, 6-shogaol was shown to downregulate constitutive activation of NF-κB as well as their target gene products, including cyclin D1, COX-2, survivin, Bcl-2, XIAP, cIAP-1, and MMP-9 in pancreatic cancer (Fig. 2d), leading to growth arrest and apoptosis induction (Fig. 1a–d). These observations are valuable for several reasons. Firstly, COX-2, which is overexpressed in pancreatic cancer, plays a significant role in cancer cell survival and angiogenesis (28,29). Additionally, Bcl-2 has been reported to contribute to pancreatic cancer chemoresistance (30). Moreover, a large body of data indicates that survivin, XIAP, and cIAP-1, members of IAP proteins are constitutively activated in pancreatic adenocarcinoma tissue samples and correlate with resistance to chemotherapy (31,32). Therefore, targeting of these proteins leads to enhanced chemosensitivity of pancreatic cancer.
TLR4 and its downstream transcription factor NF-κB have been detected constitutively active not only in pancreatic cancer cells, but also in gastric, breast, and colon cancer cells (33–35). Since we found that 6-shogaol could inhibit TLR4/NF-κB signaling cascade in pancreatic cancer cells, we propose that 6-shogaol can also inhibit the growth of other cancer cells in which TLR4 is highly active, and these can be investigated in future studies.
Owing to de novo and acquired chemoresistance, pancreatic cancer is highly resistant to conventional chemotherapy. One feature that chemotherapeutic treatments of cancer have in common is their activation of NF-κB, which mediates cell survival. In recent years, combinatorial approach has been used to overcome this problem by the application of rational chemotherapeutic combinations, such as combination of NF-κB inhibitors and chemotherapy drugs (36). In our study, 6-shogaol was used in combination with gemcitabine to test its efficacy against pancreatic cancer. Results from our experiments showed that pretreatment with 6-shogaol was effective in sensitizing pancreatic cancer cells to gemcitabine-induced apoptosis. Further, to confirm synergism, IC50 was calculated (IC50 = 36.87 and 6.60 μM for PANC-1 and BxPC-3 cells) when cells were incubated with gemcitabine for 72 h. These are within the IC50 range reported by others (37). Actually, even for one particular cell line, the IC50 values range from a few nanomole to several millimole (38,39). This mainly relates to gemcitabine chemoresistance in pancreatic cancer and IC50 calculation method. Cell status and treatment deviation also caused wide range of IC50. In general, it would not affect synergism analysis. Interestingly, pretreatment with 6-shogaol resulted in an inhibition of constitutive and gemcitabine-induced NF-κB activity and exerted its chemosensitization effects to gemcitabine. Similarly, several dietary chemopreventive compounds have also been shown to sensitize pancreatic cancer cells to gemcitabine through the downregulation of NF-κB (40,41).
In the xenograft mouse model, we found that 6-shogaol effectively suppressed the growth of pancreatic cancer and the extent of inhibition was comparable to that of gemcitabine alone. Most interestingly, the combination treatment of 6-shogaol with gemcitabine was found to be much more effective in inhibiting growth of tumors than either agent alone. In vivo results verified that 6-shogaol was instrumental in suppressing the constitutive activation as well as gemcitabine-induced activation of NF-κB. These results further corroborated the hypothesis that 6-shogaol could induce chemotherapeutic sensitization of pancreatic cancer to gemcitabine treatment.
CONCLUSION
Our present findings showed that 6-shogaol had significant potential for the treatment of pancreatic cancer. Firstly, 6-shogaol inhibited TLR4 signaling resulting in a reduced activation of NF-κB, which led to a delayed growth of pancreatic cancer. Moreover, pretreatment with 6-shogaol resulted in an inhibition of constitutive and gemcitabine-induced NF-κB activity and exerted its chemosensitization effects to gemcitabine. Our results provided evidence to support further development of 6-shogaol as an adjuvant to conventional chemotherapeutics for the treatment of pancreatic tumors as well as other cancer cells with high TLR4 expression.
Electronic Supplementary Material
(DOCX 1615 kb)
Acknowledgments
This work was supported by the National Science Foundation of China (No. 81001618), the National Science &Technology Pillar Program during the Twelfth Five-year Plan Period (2012BAI29B07), and program for New Century Excellent Talents in University (NCET-12-0976).
Footnotes
Ling Zhou and Lianwen Qi contributed equally to this work.
Contributor Information
Xiaojun Xu, Phone: +86-25-83271379, FAX: +86-25-83271379, Email: xiaojunxu2000@163.com.
Ping Li, Phone: +86-25-83271379, FAX: +86-25-83271379, Email: liping2004@126.com.
References
- 1.Coussens LM, Werb Z. Inflammation and cancer. Nature. 2002;420(6917):860–7. doi: 10.1038/nature01322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Yoshimoto S, Loo TM, Atarashi K, Kanda H, Sato S, Oyadomari S, et al. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome. Nature. 2013;499(7456):97–101. doi: 10.1038/nature12347. [DOI] [PubMed] [Google Scholar]
- 3.Ikebe M, Kitaura Y, Nakamura M, Tanaka H, Yamasaki A, Nagai S, et al. Lipopolysaccharide (LPS) increases the invasive ability of pancreatic cancer cells through the TLR4/MyD88 signaling pathway. J Surg Oncol. 2009;100(8):725–31. doi: 10.1002/jso.21392. [DOI] [PubMed] [Google Scholar]
- 4.Zhang J-J, Wu H-S, Wang L, Tian Y, Zhang J-H, Wu H-L. Expression and significance of TLR4 and HIF-1 alpha in pancreatic ductal adenocarcinoma. World J Gastroenterol. 2010;16(23):2881–8. doi: 10.3748/wjg.v16.i23.2881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Arlt A, Gehrz A, Muerkoster S, Vorndamm J, Kruse ML, Folsch UR, et al. Role of NF-kappa B and Akt/PI3K in the resistance of pancreatic carcinoma cell lines against gemcitabine-induced cell death. Oncogene. Oncogene. 2003;22(21):3243–51. doi: 10.1038/sj.onc.1206390. [DOI] [PubMed] [Google Scholar]
- 6.Hung J-Y, Hsu Y-L, Li C-T, Ko Y-C, Ni W-C, Huang M-S, et al. 6-Shogaol, an active constituent of dietary ginger, induces autophagy by inhibiting the AKT/mTOR pathway in human non-small cell lung cancer A549 cells. J Agric Food Chem. 2009;57(20):9809–16. doi: 10.1021/jf902315e. [DOI] [PubMed] [Google Scholar]
- 7.Ishiguro K, Ando T, Maeda O, Ohmiya N, Niwa Y, Kadomatsu K, et al. Ginger ingredients reduce viability of gastric cancer cells via distinct mechanisms. Biochem Biophys Res Commun. 2007;362(1):218–23. doi: 10.1016/j.bbrc.2007.08.012. [DOI] [PubMed] [Google Scholar]
- 8.Tan BS, Kang O, Mai CW, Tiong KH, Khoo AS-B, Pichika MR, et al. 6-Shogaol inhibits breast and colon cancer cell proliferation through activation of peroxisomal proliferator activated receptor gamma (PPARgamma) Cancer Lett. 2013;336(1):127–39. doi: 10.1016/j.canlet.2013.04.014. [DOI] [PubMed] [Google Scholar]
- 9.Hu R, Zhou P, Peng Y-B, Xu X, Ma J, Liu Q, et al. 6-Shogaol induces apoptosis in human hepatocellular carcinoma cells and exhibits anti-tumor activity in vivo through endoplasmic reticulum stress. PLoS One. 2012;7(6):e39664. doi: 10.1371/journal.pone.0039664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Liu Q, Peng Y-B, Qi L-W, Cheng X-L, Xu X-J, Liu L-L, et al. The cytotoxicity mechanism of 6-shogaol-treated HeLa human cervical cancer cells revealed by label-free shotgun proteomics and bioinformatics analysis. Evid Based Complement Alternat Med. 2012;2012:278652. doi: 10.1155/2012/278652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ling H, Yang H, Tan SH, Chui WK, Chew EH. 6-Shogaol, an active constituent of ginger, inhibits breast cancer cell invasion by reducing matrix metalloproteinase-9 expression via blockade of nuclear factor-kappa B activation. Br J Pharmacol. 2010;161(8):1763–77. doi: 10.1111/j.1476-5381.2010.00991.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Park S-J, Lee M-Y, Son B-S, Youn H-S. TBK1-targeted suppression of TRIF-dependent signaling pathway of toll-like receptors by 6-shogaol, an active component of ginger. Biosci, Biotechnol, Biochem. 2009;73(7):1474–8. doi: 10.1271/bbb.80738. [DOI] [PubMed] [Google Scholar]
- 13.Chou T-C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol Rev. 2006;58(3):621–81. doi: 10.1124/pr.58.3.10. [DOI] [PubMed] [Google Scholar]
- 14.Harikumar KB, Kunnumakkara AB, Sethi G, Diagaradjane P, Anand P, Pandey MK, et al. Resveratrol, a multitargeted agent, can enhance antitumor activity of gemcitabine in vitro and in orthotopic mouse model of human pancreatic cancer. Int J Cancer. 2010;127(2):257–68. doi: 10.1002/ijc.25041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Liptay S, Weber CK, Ludwig L, Wagner M, Adler G, Schmid RM. Mitogenic and antiapoptotic role of constitutive NF-kappa B/Rel activity in pancreatic cancer. Int J Cancer. 2003;105(6):735–46. doi: 10.1002/ijc.11081. [DOI] [PubMed] [Google Scholar]
- 16.Wang J, Cai Y, Shao L-J, Siddiqui J, Palanisamy N, Li R, et al. Activation of NF-kappa B by TMPRSS2/ERG fusion isoforms through toll-like receptor-4. Cancer Res. 2011;71(4):1325–33. doi: 10.1158/0008-5472.CAN-10-2210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kawai T, Akira S. Signaling to NF-kappa B by toll-like receptors. Trends Mol Med. 2007;13(11):460–9. doi: 10.1016/j.molmed.2007.09.002. [DOI] [PubMed] [Google Scholar]
- 18.Akira S, Takeda K. Toll-like receptor signalling. Nat Rev Immunol. 2004;4(7):499–511. doi: 10.1038/nri1391. [DOI] [PubMed] [Google Scholar]
- 19.Banerjee S, Zhang Y, Wang Z, Che M, Chiao PJ, Abbruzzese JL, et al. In vitro and in vivo molecular evidence of genistein action in augmenting the efficacy of cisplatin in pancreatic cancer. Int J Cancer. 2007;120(4):906–17. doi: 10.1002/ijc.22332. [DOI] [PubMed] [Google Scholar]
- 20.El-Rayes BF, Shields AF, Vaitkevicius V, Philip PA. Developments in the systemic therapy of pancreatic cancer. Cancer Invest. 2003;21(1):73–86. doi: 10.1081/CNV-120016406. [DOI] [PubMed] [Google Scholar]
- 21.Ishiguro K, Ando T, Watanabe O, Goto H. Specific reaction of alpha, beta-unsaturated carbonyl compounds such as 6-shogaol with sulfhydryl groups in tubulin leading to microtubule damage. FEBS Lett. 2008;582(23–24):3531–6. doi: 10.1016/j.febslet.2008.09.027. [DOI] [PubMed] [Google Scholar]
- 22.Youn HS, Saitoh SI, Miyake K, Hwang DH. Inhibition of homodimerization of Toll-like receptor 4 by curcumin. Biochem Pharmacol. 2006;72:62–9. doi: 10.1016/j.bcp.2006.03.022. [DOI] [PubMed] [Google Scholar]
- 23.Youn HS, Lee JK, Choi YJ, Saitoh SI, Miyake K, Hwang DH, et al. Cinnamaldehyde suppresses toll-like receptor 4 activation mediated through the inhibition of receptor oligomerization. Biochem Pharmacol. 2008;75(2):494–502. doi: 10.1016/j.bcp.2007.08.033. [DOI] [PubMed] [Google Scholar]
- 24.Youn H-S, Lim HJ, Lee HJ, Hwang D, Yang M, Jeon R, et al. Garlic (Allium sativum) extract inhibits lipopolysaccharide-induced toll-like receptor 4 dimerization. Biosci, Biotechnol, Biochem. 2008;72(2):368–75. doi: 10.1271/bbb.70434. [DOI] [PubMed] [Google Scholar]
- 25.Ahn S-I, Lee J-K, Youn H-S. Inhibition of homodimerization of toll-like receptor 4 by 6-shogaol. Mol Cells. 2009;27(2):211–5. doi: 10.1007/s10059-009-0026-y. [DOI] [PubMed] [Google Scholar]
- 26.Wang WX, Abbruzzese JL, Evans DB, Larry L, Cleary KR, Chiao PJ. The nuclear factor-kappa B RelA transcription factor is constitutively activated in human pancreatic adenocarcinoma cells. Clin Cancer Res. 1999;5(1):119–27. [PubMed] [Google Scholar]
- 27.Fujioka S, Sclabas GM, Schmidt C, Frederick WA, Dong QG, Abbruzzese JL, et al. Function of nuclear factor kappa B in pancreatic cancer metastasis. Clin Cancer Res. 2003;9(1):346–54. [PubMed] [Google Scholar]
- 28.Albazaz R, Verbeke CS, Rahman SH, McMahon MJ. Cyclooxygenase-2 expression associated with severity of PanIN lesions: a possible link between chronic pancreatitis and pancreatic cancer. Pancreatology. 2005;5(4–5):361–9. doi: 10.1159/000086536. [DOI] [PubMed] [Google Scholar]
- 29.Yip-Schneider MT, Barnard DS, Billings SD, Cheng L, Heilman DK, Lin A, et al. Cyclooxygenase-2 expression in human pancreatic adenocarcinomas. Carcinogenesis. 2000;21(2):139–46. doi: 10.1093/carcin/21.2.139. [DOI] [PubMed] [Google Scholar]
- 30.Bai JR, Sui JH, Demirjian A, Vollmer CM, Marasco W, Callery MP. Predominant Bcl-XL knockdown disables antiapoptotic mechanisms: tumor necrosis factor-related apoptosis-inducing ligand-based triple chemotherapy overcomes chemoresistance in pancreatic cancer cells in vitro. Cancer Res. 2005;65(6):2344–52. doi: 10.1158/0008-5472.CAN-04-3502. [DOI] [PubMed] [Google Scholar]
- 31.Tamm I, Kornblau SM, Segall H, Krajewski S, Welsh K, Kitada S, et al. Expression and prognostic significance of IAP-family genes in human cancers and myeloid leukemias. Clin Cancer Res. 2000;6(5):1796–803. [PubMed] [Google Scholar]
- 32.Lee MA, Park GS, Lee HJ, Jung JH, Kang JH, Hong YS, et al. Survivin expression and its clinical significance in pancreatic cancer. BMC Cancer. 2005;5:127. doi: 10.1186/1471-2407-5-127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.de Oliveira JG, Silva AE. Polymorphisms of the TLR2 and TLR4 genes are associated with risk of gastric cancer in a Brazilian population. World J Gastroenterol. 2012;18(11):1235–42. doi: 10.3748/wjg.v18.i11.1235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ahmed A, Wang JH, Redmond HP. Silencing of TLR4 increases tumour progression and lung metastasis in a murine model of breast cancer. Br J Surg. 2011;98:26–6. doi: 10.1245/s10434-012-2595-9. [DOI] [PubMed] [Google Scholar]
- 35.Huang B, Zhao J, Li HX, He KL, Chen YB, Mayer L, et al. Toll-like receptors on tumor cells facilitate evasion of immune surveillance. Cancer Res. 2005;65(12):5009–14. doi: 10.1158/0008-5472.CAN-05-0784. [DOI] [PubMed] [Google Scholar]
- 36.Bold RJ, Virudachalam S, McConkey DJ. Chemosensitization of pancreatic cancer by inhibition of the 26S proteasome. J Surg Res. 2001;100(1):11–7. doi: 10.1006/jsre.2001.6194. [DOI] [PubMed] [Google Scholar]
- 37.Soukaina R, Christelle B, Celine P, Amor H. Gemcitabine-based chemogene therapy for pancreatic cancer using Ad-dCK::UMK GDEPT and TS/RR siRNA strategies. Neoplasia. 2009;11(7):637–50. doi: 10.1593/neo.81686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mark SD, Hiromichi I, Michael JZ, Stanley WA, Edward EW. Inhibition of Src tyrosine kinase impairs inherent and acquired gemcitabine resistance in human pancreatic adenocarcinoma cells. Clin Cancer Res. 2004;10(7):2307–18. doi: 10.1158/1078-0432.CCR-1183-3. [DOI] [PubMed] [Google Scholar]
- 39.Rosemary AF, Blake B, Christine G, Angusg D. Mechanisms underlying gemcitabine resistance in pancreatic cancer and sensitisation by the iMiD™ lenalidomide. Anticancer Res. 2011;31(11):3747–56. [PubMed] [Google Scholar]
- 40.Banerjee S, Kaseb AO, Wang Z, Kong D, Mohammad M, Padhye S, et al. Antitumor activity of gemcitabine and oxaliplatin is augmented by thymoquinone in pancreatic cancer. Cancer Res. 2009;69(13):5575–83. doi: 10.1158/0008-5472.CAN-08-4235. [DOI] [PubMed] [Google Scholar]
- 41.Banerjee S, Zhang YX, Ali S, Bhuiyan M, Wang ZW, Chiao PJ, et al. Molecular evidence for increased antitumor activity of gemcitabine by genistein in vitro and in vivo using an orthotopic model of pancreatic cancer. Cancer Res. 2005;65(19):9064–72. doi: 10.1158/0008-5472.CAN-05-1330. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX 1615 kb)