

Abstract
The membrane of red blood cells consists of a phospholipid bilayer with embedded membrane proteins and is associated on the cytoplasmatic side with a network of proteins, the membrane skeleton. Band3 has an important role as centre of the functional complexes e.g. gas exchange complex and as element of attachment for the membrane skeleton maintaining membrane stability and flexibility. Up to now it is unclear if band3 is involved in the morphology change of red blood cells after contact with radiographic contrast media. The study revealed for the first time that Iopromide induced markedly more severe alterations of the membrane skeleton compared to Iodixanol whose effects were similar to erythrocytes suspended in autologous plasma. A remarkable clustering of band3 was found associated with an accumulation of band3 in spicules and also a sequestration of band3 to the extracellular space. This was evidently accompanied by a gross reduction of functional band3 complexes combined with a dissociation of spectrin from band3 leading to a loss of homogeneity of the spectrin network. It could be demonstrated for the first time that RCM not only induced echinocyte formation but also exocytosis of particles at least coated with band3.
Introduction
Radiographic contrast media (RCM) are widely used to visualize arteries e.g. in interventional cardiology. Their effects on blood vessels should be minimal [1] so that the object of the measurement – blood cells and vessels - are not influenced by the RCM molecules. However, all RCMs exhibit a more or less strong effect on endothelial cells [2]–[5] as well as on erythrocytes [6]–[14]. Up to now it is unclear which organelles or parts of the cells are effected by RCM. In a former study it was shown that RCM did not attach to or intercalate in the erythrocyte membrane and thus effect echinocytec formation as was discussed for a long time [6], [15]. There is a lot of evidence that echinocyte formation is not possible without participation of the membrane cytoskeleton. Reinhart et al. could show that the presence of spectrin is essential for echinocyte formation [16]. In addition, Becker & Lux could show that dysfunctional band3 resulted clinically in hereditary spherocytosis [17]. Therefore, it can be speculated that both constituents of the membrane cytoskeleton might interact and effect echinocyte formation what has never been shown so far.
The erythrocyte membrane consists of a phospholipid bilayer with embedded membrane proteins and is associated on the cytoplasmatic side with a network of proteins, the membrane cytoskeleton [18]. The bicarbonate/chloride exchanger band3 (Anion Exchanger 1, AE1) is the most abundant protein in the erythrocyte membrane [19]. It has an important role e.g. in gas exchange, senescence and removal of cells from the circulation, anchoring motifs for the glycolytic enzymes (GE) and functions as a point of attachment for the cytoskeleton maintaining the mechanical and osmotic properties of the erythrocyte [20], [21]. Band3 is found in three distinct protein complexes associated with the erythrocyte membrane: an ankyrin-dependent tetrameric band3 complex, a dimeric band3 complex bound to the protein 4.1-glycophorin C junctional complex and freely diffusing dimeric band3 complexes [22], [23].
It appears that an intact cytoskeleton is vital for normal cell shape. Defects or deficiencies in band3 were described to lead to a decrease in cohesion between the lipid bilayer and membrane skeleton resulting in loss of membrane surface area and a pathology termed hereditary spherocytosis [24]. Also in some haemolytic anemias, the erythrocytes can not only become spherical but are also extremely fragile, implying that the membrane skeleton furnishes stability and elasticity to the cells [17], [25]–[29]. It was shown that in mice with a knock out of band3 immediate post-natal death of most pups coincided with a high fragility of the erythrocyte membrane [30].
Reinhart et al. reported that echinocytes did not occur in spectrin-deficient knock-out mice [16]. Therefore, it can be assumed that spectrin is also a key element of erythrocytic shape changes. Also the link between the membrane skeleton and the membrane seems to be important for normal shape as rupture of the interaction of the linking proteins prevents any shape changes and reduces deformability of red cell membranes [31], [32].
It was reported that under physiological conditions the spectrin net is linked to band3 via multiple high affinity protein-protein interactions (e.g. ankyrin, adducin, protein 4.1, glycophorin [20]).
It is unknown whether the distances change between binding partners, especially spectrin and band3, during echinocytic shape changes. Closely spaced elements like band3 (stained red) or spectrin (stained green) will be subjected to a colour shift from red/green to yellow when these two elements approach one another and become co-localized. Therefore, a co-localization study was performed based on immuno-cytochemical double staining of erythrocytes. This would allow to prove whether RCM induce intracellular effects mediated by spectrin and band3 and whether both RCM act in the same way.
Materials and Methods
The purpose of the investigation was to examine whether radiographic contrast media provoked echinocyte formations are accompanied by a reorganisation of band3 and/or spectrin filaments.
Ethics Statement
The study protocol conformed to the ethical guidelines of the 1975 Declaration of Helsinki as reflected in a priori approval by the institutional review committee of the Medical Faculty of the Heinrich Heine University Düsseldorf (registry number: 3522). Written informed consent was obtained from each subject before entry into the study.
Radiographic Contrast Media
Two radiographic contrast media with variant Iodine concentrations and approved for intra-arterial application were examined: (Iodixanol 320 mg Iodine/ml, GE Healthcare, München, Germany; Iopromide 370 mg Iodine/ml Bayer/Schering, Berlin, Germany).
Blood Collection
Venous blood (20 ml) was collected in a standardized manner from the cubital veins of n = 6 healthy adults anticoagulated with potassium EDTA according to actual International guidelines [33]. The samples were stored in sealed polystyrene tubes.
Sample Processing
Immediately after sampling, plasma and erythrocytes were separated by centrifugation (500 g, 5 min). Plasma was harvested and the plasma/radiographic contrast media mixtures required for suspension of the erythrocytes were prepared. Iodixanol or Iopromide in the concentration of 30% v/v were added to the plasma. Then, the red blood cells – without the buffy coat - were suspended in autologous plasma (control group) or in the RCM/plasma mixtures and incubated for 5 minutes at 37°C.
Staining of Components of the Membrane Cytoskeleton
After the incubation of erythrocytes in autologous plasma or in different RCM/plasma mixtures (30% v/v Iodixanol or Iopromide, respectively), conventional blood smears were prepared on glass substrates and air dried. For each of the 3 groups 18 slides (3 from every donor) were layered with cells.
In the follow up the air dried samples were postfixed in 2% paraformaldehyde. After short rinsing in isotonic phosphate buffered saline (PBS ) at room temperature the samples were transferred into cold acetone (−20°C) for 2 minutes to render the cell membranes permeable for the antibodies. Components of the membrane cytoskeleton were double stained in consecutive steps to display the distribution of the components in the cytoskeleton. The components were stained either in red (band3; first antibody: SLC4A1 rabbit anti human polyclonal antibody (Biozol, Eching, Germany; dilution 1∶50 in PBS), second antibody: anti rabbit IgG TRITC conjugated (dilution 1∶30 in PBS)) or in green (spectrin; first antibody: mouse anti human spectrin IgG (dilution 1∶30 in PBS), second antibody: anti mouse IgG FITC conjugated (dilution 1∶30 in PBS)).
In case of a very close proximity of actin and spectrin, confocal laser scanning microscopy allows to visualise a merger of the colours red and green resulting in a colour shift towards yellow colour tones. Vice versa, if there should be a dissociation of earlier closely spaced components then there would be a colour shift from yellow coloration to red and green [34], [35]. The cells were visualized using confocal laser scanning microscopy at a primary magnification of 1∶63 (TCS SP5, Leica, Wetzlar, Germany). The appearance of pixels of preselected yellow tones in the images was quantified using an image analysis system (MacIntosh, Intelcore I7 with Adobe Photoshop, extended version). Those yellow colour tones (merger of green and red colours) were selected which were found in the roots of exocytotic-like protrusions from erythrocytes [36], [37]. 10 samples per group were examined and 4 fields of view per sample with more than 200 cells were analyzed and the mean numbers of pre-selected yellow tone pixels and the number of red dots (condensed band 3) were quantified.
For all samples mean value and standard deviation are given. For multi-sample comparisons a variance analysis for repeated measurements was performed. To isolate the group or groups that differed from the others, Student-Newman-Keuls test for paired comparisons was used. A probability value of less than 0.05 was accepted as significant.
Results
Figure 1 shows double stained erythrocytes suspended in autologous plasma in the red or the green channel of the confocal laser scanning microscope.
Figure 1. Double stained erythrocytes in autologous plasma.

Band3: left image in red; spectrin: middle image in green; right image merger of red (band3) and green (spectrin). Co-localized structures are coloured in yellow tones; (primary magnification 1∶63; zoom factor 1∶2.1).
Discocytes revealed more band3 stain in central cell parts compared to the cell periphery. In erythrocyte aggregates the stain was enhanced in contact regions of the cells. Differing from band3 stain the spectrin stain was distributed over the cells more homogenously and there was no enhancement of spectrin stain in contact regions of aggregated erythrocytes.
The merger of red and green colour channels shows a yellow shift in a few cases (see arrow) where band3 and spectrin were co-localized. On an average 40.5±5.5 yellow pixels per field of view were found.
Incubation of Erythrocytes in an Iodixanol/plasma-mixture (30% v/v)
Most erythrocytes were found as single cells, a few in the majority non-branched linear aggregates (rouleaux) were observed (more or less comparable to erythrocytes in autologous plasma).
Figure 2 shows erythrocytes which were suspended in a Iodixanol/plasma-mixture (30% v/v). There were only few cells showing echinocytic shape change. Cells with spicules showed increased amounts of stained band3 in these spicules. At the same time less spectrin stain was found there indicating that spectrin did not seem to enter the spicules (therefore, in the spectrin stain it is difficult to identify echinocytes).
Figure 2. Double stained erythrocytes after incubation in plasma supplemented with Iodixanol320 (30% v/v).

Band3 is stained in red (left image), spectrin in green (middle image). The merger of both colour channels is presented in the right image with co-localized structures coloured in yellow tones (30% v/v; primary magnification 1∶63; zoom factor 1∶1).
However, other echinocytes with well rounded protrusions exhibited spectrin staining. In the few linearly aggregated erythrocytes (rouleaux) the green stain was enhanced in the contacting regions. It can be assumed that in these contacting regions spectrin was concentrated hinting to a condensation of the spectrin network.
Compared to cells in autologous plasma the distribution of band3 and spectrin in cells suspended in Iodixanol/plasma-mixture was similar but the grain-like pattern of red-green coloured areas (merger of red and green channels in Figure 2 ) was a bit coarser hinting to a slight condensation of band3 and of spectrin where a tendency to condense seemed to be a bit stronger in band3.
After incubation of erythrocytes in a Iodixanol/plasma-mixture an increase in the number of pixels with yellow tone occurred (83.0±53.3 pixels on an average; p<0.05). The co-localization associated yellow colour tones appeared at the periphery of these erythrocytes incubated in Iodixanol/plasma-mixture and not in central parts as those which were found in erythrocytes incubated in autologous plasma.
Incubation of Erythrocytes in a Iopromide/plasma-mixture (30% v/v)
Figure 3 reveals that band3 stain was arranged in a rectangular fashion unlike the arrangement of band3 stain in erythrocytes suspended in autologous plasma or in Iodixanol/plasma-mixture. The formation of spicules can be observed in band3 stained erythrocytes showing a clear enhancement of band3 in these spicules (arrow in Figure 3 merged image), especially in very pointed ones, often revealing band3 in the tip of the spicule (arrow in Figure 4).
Figure 3. Double staining of erythrocytes after incubation in plasma supplemented with Iopromide370 (30% v/v).

Band3 (left image) is coloured in red; spectrin (middle image) in green. The merger of red (band3) and green (spectrin) channels is presented in the right image with co-localized structures in yellow tones (primary magnification 1∶63; zoom factor 1∶1). Arrow in the middle image points at a cell with condensed spectrin bands in the central part of the cell, arrow in the right image points at condensed band3 (red dot) in spicule tip.
Figure 4. Co-localization of spectrin and band3 at the root of a spicule.

Band3 is coloured in red, spectrin in green. The co-localization of both proteins at the root of a spicule (revealed by the yellow colour shift) is marked by an arrow (primary magnification 1∶63; zoom factor 1∶15).
Compared to erythrocytes in autologous plasma or in a Iodixanol/plasma-mixture a homogenous spectrin stain was no longer found in erythrocytes suspended in a Iopromide/plasma-mixture (Figure 3 middle). Most of the cells were found in linear branched or un-branched or in non-linear branched aggregates (rosettes or cross-like aggregates) where the spectrin stain was mainly concentrated at the border regions of contacting cells and often running through central parts of cells as a fibre-like network (arrow in Figure 3 middle). One of the most obvious findings and most clear to recognize was the rectangular arrangement of the spectrin network – completely different from the homogenous distribution and more or less round arrangement of spectrin in erythrocytes suspended in autologous plasma or in a Iodixanol/plasma-mixture.
The merger of red (band3) and green (spectrin) channels showed that co-localizations were rarely seen (44.0±34.2 pixels on an average) and that there was a multitude of relatively fine grained red dots revealing a concentration of band3 in echinocytic spicule tips (arrows in Figure 3 merger of red and green channels, Figure 4) or outside of cells possibly extruded from the spicule tips (see Figure 5).
Figure 5. Condensed band3 outside of erythrocytes.

Details from figure 3 right (image electronically magnified): condensed band3 stain outside of erythrocytes (left image) or loosely connected to the erythrocyte membrane (right image) (primary magnification 1∶63; zoom factor 1∶10).
Figure 4 shows an erythrocyte in detail with condensed band3 in spicule tip marked by an arrow in Figure 3. Co-localizations of spectrin and band3 (see yellow colour in Figure 4 marked by arrow) appeared almost exclusively at the roots of the spicules and not in central parts of erythrocytes (as in cells incubated in autologous plasma).
Figure 5 demonstrates for the first time that particles at least coated with band3 were found outside of erythrocytes and gives evidence that not only echinocyte formation with spicules occurred after RCM contact but also exocytosis with the export of band3 from erythrocytes.
Quantitative Image Analysis
An image-analysis showed that the appearance of pixels with selected yellow tones in red/green merged images, due to a co-localization of band3 (red stained structures) and spectrin (green stained structures), differed significantly between the three groups (Kruskal Wallis test: p = 0.003). Using a post-hoc analysis (Tukey tests) it could be shown, that cells in a Iodixanol/plasma mixture showed significantly more co-localized spectrin and band3 (83.9±53.3 yellow tone pixels) than cells suspended in autologous plasma (40.4±5.5 yellow pixels; p<0.05) or cells suspended in a Iopromid/plasma mixture (44.4±34.2 pixels; p<0.05).
Also the number of condensed band3 dots inside of erythrocytes was counted. In cells suspended in autologous plasma a mean number of 5.5±1.6 condensed band3 dots was found, in cells suspended in a Iodixanol/plasma mixture 27.5±4.9 and in cells suspended in a Iopromide/plasma mixture 16.8±5.1 condensed band3 dots (p<0.001) were found. The number of dots in the Iodixanol/plasma mixture differed significantly from the number in cells in autologous plasma and as well from the number in Iopromide/plasma mixture (p<0.05 each).
In a third analysis the numbers of band3 particles outside of erythrocytes were counted. Here, the three groups also differed significantly (p<0.001). After suspending erythrocytes in a Iopromide/plasma mixture 6.7±1.4 band3 particles were found, after suspending erythrocytes in a Iodixanol/plasma mixture 1.3±1.2 particles and in cells suspended in autologous plasma 0.17±0.4 particles were found. The number of band3 particles in the Iopromide/plasma mixture differed significantly from the number of such particles in autologous plasma and as well from the number in a Iodixanol/plasma mixture (p<0.05 each).
Discussion
The study revealed that the shape changes or the aggregation of erythrocytes in contact with radiographic contrast media (RCM) clearly coincided with changes in the structure of the membrane skeleton depending on the type of RCM used. This was most clearly demonstrated by the abolishment of the homogenous spectrin distribution with a relocation of spectrin primarily to the rim of erythrocytes and accompanied by appearance of some thick bundles of spectrin crossing central parts of cells suspended in a Iopromide/plasma-mixture but not in a Iodixanol/plasma-mixture (in accordance with the results from a former study [36]). Another remarkable finding was the formation of rosette-like erythrocyte aggregates only in Iopromide/plasma-mixtures. This seems to imply an activation of complement according to Luginbühl [38], which was found to occur in vivo after application of hyperosmolar RCM (+70.2%, p<0.00001) but only weakly (+9%, p<0.026) [39] or not at all [40] after application of isoosmolar RCM. This is in good agreement with former studies describing a weak increase in linear erythrocyte aggregates but not the formation of rosettes [9], [36], [41].
Remarkable differences appeared with respect to the formation of protrusions and spicules which contained band3. Small amounts of protrusions and spicules were demonstrated in erythrocytes suspended in autologous plasma or in a Iodixanol/plasma-mixture (slightly more than in autologous plasma). Considerably more protrusions and spicules were found after suspension of erythrocytes in a Iopromide/plasma-mixture. These results correlate very well with findings in trans-illuminated microscopical studies [10], [42]. It has to be kept in mind that in the study reported here the silhouettes of the membrane cytoskeleton were displayed and not the shapes of the cell membrane.
The major structural member of the erythrocytic membrane skeletal network is the spectrin tetramer [43]. In this study changes in the membrane skeleton of erythrocytes were characterized (see Figure 6) by a transition from a homogenous distribution of the spectrin network (cells in autologous plasma, or in a Iodixanol/plasma-mixture) to the generation of spectrin bundles mostly concentrated at the cell rim and sometimes found also in central parts of the erythrocytes (cells in a Iopromide/plasma-mixture). This transition was described to be caused by disruption of the ankyrin-band3 binding [44], coinciding with a loss of nearness of spectrin dimers necessary for their self-association [45]. The tetrameric state of spectrin is assumed to be essential in order to maintain the normal membrane mechanical stability [13], [43], [46]. Shear elasticity of the red cell membrane was reported to be regulated by extension of spectrin tetramers and a reversible unfolding of constituent triple helical repeats [18]. The adducin-band3-spectrin bridge - more recently described [20] - probably contributes also to membrane stability.
Figure 6. Merger of red (band3) and green (spectrin) channels after examination in a confocal laser scanning microscope.
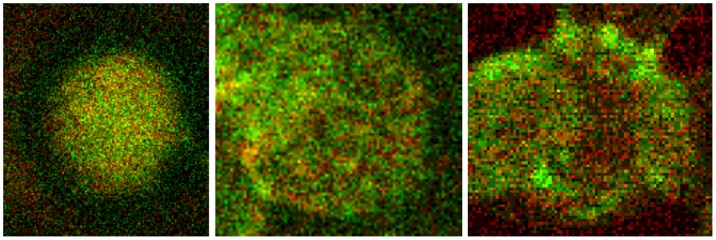
Double stained erythrocytes with antibodies against band3 (red) and spectrin (green, details from Figures 1 , 2 , 3 ). Left: erythrocyte suspended in autologous plasma, middle: erythrocyte suspended in a Iodixanol/plasma-mixture (30% v/v), right: erythrocyte suspended in a Iopromide/plasma-mixture (30% v/v; primary magnification 1∶63; zoom factor 1∶10).
Figure 6 left shows a representative image of erythrocytes suspended in autologous plasma displaying a more or less homogeneous distribution of band3 and spectrin. Erythrocytes suspended in Iodixanol/plasma mixtures (middle) showed a little less homogeneous distribution of band3 and spectrin with a few small aggregates of spectrin found in the majority in central parts of the cells. Yellow tone areas due to co-localization of band3 and spectrin appeared in the periphery. The right image shows an erythrocyte suspended in a Iopromide/plasma-mixture. The most important aspect is the arrangement of few very coarse aggregates of spectrin almost completely found at the periphery of cells.
Erythrocytes in autologous plasma displayed a more or less homogeneous distribution of fine-grained band3 (Figure 7, left). Erythrocytes in Iodixanol/plasma-mixture showed a band3 distribution which was less homogeneous with a few coarser band3 aggregates in central parts of the cell as well as in the periphery. Erythrocytes in Iopromide/plasma-mixture displayed an inhomogeneous pattern of band3 with almost no band3 aggregates in central parts and very few coarse band3 aggregates concentrated at the cell rim.
Figure 7. Representative erythrocytes immunologically stained with antibodies against band3 (details from Figures 1, 2, 3).

Left: erythrocyte suspended in autologous plasma, middle: erythrocyte suspended in a Iodixanol/plasma mixture (30% v/v), right: erythrocyte suspended in a Iopromide/plasma mixture (30% v/v; primary magnification 1∶63; zoom factor 1∶10).
In a very recent study Franke et al. [36] reported that upon Iopromide addition the spectrin net usually multiply linked to the erythrocyte membrane abandoned its well- rounded shape near the cell membrane now forming a box-like structure almost rectangular and linked to the erythrocyte membrane via very few – about 8 to 10– junctional elements [36], which affords the dissociation of the vast majority of connections between the spectrin network and the membrane. Such a box-like structure of the spectrin net after addition of Iopromide to the plasma is shown in Figure 8B and C with numbers from 1–7 depicting the corners of such a box-like structured spectrin configuration (Figure 8C; where corner number 8 is hidden behind number 3).
Figure 8. Band3 and spectrin structures after Iopromide contact.

Band3 aggregates (A) and box-like spectrin configuration (B, C) of single erythrocytes after contact with Iopromide (primary magnification 1∶63; zoom factor 1∶10).
It might be hypothesized that the very few coarse band3 aggregates shown in Figure 7C and Figure 8A are the junctional elements binding the rectangular box-like spectrin configuration to the erythrocyte membrane.
Band3 is described to constitute a centre organized into complexes for gas transport [47]–[50], for cation efflux pumping (anion exchanger) [51]–[53], for glycolysis [54], [55], for the control of cell volume [56] and of erythrocyte life span (senescence) [57], where most probably further complexes exist to cope with mechanical deformation [32], [58], and haemostatic stimuli [59]. The drastic reorganization of the band3 - spectrin network effected by suspension of erythrocytes in a Iopromide/plasma-mixture coincided with a strong alteration of the spectrin network and a clustering of band3 in very few highly condensed centres at the rim of the cells. This is most probably accompanied by a strong increase in membrane stiffness (contributing to microcirculatory disorders shown in patients with coronary artery disease after Iopromide application during coronary angiography [60], [61]). Also a strong decrease of centres for gas transport can be expected contributing to hypo-oxygenation of the tissue as was shown after Iopromide application in the myocardium of pigs [62] but not after Iodixanol application [63]. The clustering of band3 is assumed to lead to a shortening of the lifespan and removal of erythrocytes by offering senescence signals.
This study revealed for the first time a remarkable loss of band3 after contact of erythrocytes with Iopromide demonstrated by a sequestration of band3 probably out of the tips of the spicules giving evidence that the echinocyte formation provoked by Iopromide seems to be associated with an exocytotic-like process – a mechanism well known from somatic cells containing nuclei [64], [65], but up to now not described in erythrocytes. As band3 is a key element of binding the membrane cytoskeleton to the erythrocyte membrane, the loss of band3 could be associated with an irreversible reduction of membrane cytoskeleton binding potential. This irreversible reduction might be the reason that the echinocyte formation was shown to be reversible only for a certain fraction of echinocytes after their resuspension in autologous plasma [66]. It might be hypothesized that particularly older erythrocytes – with less cell volume [67], and decreases in haemoglobin [67] and cell function [68] – could be the prime targets.
Conclusion
Differences in the formation of echinocytes after their suspension in RCM/plasma-mixtures were reported earlier [9]–[13], [41] and are in good agreement with the results from this study. This co-localization study, however, goes far beyond as differences exerted by Iodixanol versus Iopromide in the formation of protrusions and spicules and of echinocytes could be attributed to subcellular phenomena, however, in different ways. It could be demonstrated for the first time that Iopromide induced echinocyte formation coinciding with a more or less rectangularly arranged spectrin configuration mainly concentrated at the border regions of contacting cells and often running through central parts of cells as a fibre-like network – completely different from the homogenous spectrin distribution of cells in autologous plasma or in a Iodixanol/plasma-mixture. In addition, also a process resembling exocytosis of particles at least coated with band3 could be demonstrated after incubation of erythrocytes in Iopromide/plasma-mixture.
Iopromide, evidently, induced different and markedly more severe alterations of the membrane skeleton compared to Iodixanol whose effects were similar to erythrocytes suspended in autologous plasma.
Funding Statement
The authors thank the Bundesministerium für Bildung und Forschung (BMBF) for funding within project number 0315696A “Poly4Bio BB”. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Aspelin P, Stacul F, Thomsen HS, Morcos SK, van der Molen AJ, et al. (2006) Effects of iodinated contrast media on blood and endothelium. Eur Radiol 16: 1041–1049. [DOI] [PubMed] [Google Scholar]
- 2. Franke RP, Fuhrmann R, Hiebl B, Jung F (2011) Influence of radiographic contrast media (iodixanol und iomeprol) on the morphology of human arterial and venous endothelial cells on extracellular matrix in vitro. Clin Hemorheol Microcirc 48: 41–56. [DOI] [PubMed] [Google Scholar]
- 3. Franke RP, Fuhrmann R, Park JW, Rickert D, Hiebl B, et al. (2007) The effect of radiographic contrast media on the morphology of human venous endothelial cells. Clin Hemorheol Microcirc 37: 329–338. [PubMed] [Google Scholar]
- 4. Ramponi S, Grotti A, Morisetti A, Vultaggio S, Lorusso V (2007) Effects of iodinated contrast media on endothelium: An in vitro study. Toxicol In Vitro 21: 191–196. [DOI] [PubMed] [Google Scholar]
- 5. Sumimura T, Sendo T, Itoh Y, Oka M, Oike M, et al. (2003) Calcium-dependent injury of human microvascular endothelial cells induced by a variety of iodinated radiographic contrast media. Invest Radiol 38: 366–374. [PubMed] [Google Scholar]
- 6. Reinhart WH, Pleisch B, Harris LG, Lutolf M (2005) Influence of contrast media (iopromide, ioxaglate, gadolinium-DOTA) on blood viscosity, erythrocyte morphology and platelet function. Clin Hemorheol Microcirc 32: 227–239. [PubMed] [Google Scholar]
- 7. Aspelin P (1979) Effect of ionic and non-ionic contrast media on red cell deformability in vitro. Acta Radiol Diagn (Stockh) 20: 1–12. [DOI] [PubMed] [Google Scholar]
- 8.Aspelin P, Nilsson PE, Schmid-Schonbein H, Schroder S, Simon R (1987) Effect of four non-ionic contrast media on red blood cells in vitro. I. Morphology. Acta Radiol Suppl 370: 79–83. [PubMed]
- 9. Hardeman MR, Goedhart P, Koen IY (1991) The effect of low-osmolar ionic and nonionic contrast media on human blood viscosity, erythrocyte morphology, and aggregation behavior. Invest Radiol 26: 810–819. [DOI] [PubMed] [Google Scholar]
- 10. Jung F, Mrowietz C, Rickert D, Hiebl B, Park JW, et al. (2008) The effect of radiographic contrast media on the morphology of human erythrocytes. Clin Hemorheol Microcirc 38: 1–11. [PubMed] [Google Scholar]
- 11. Kerl JM, Nguyen SA, Lazarchick J, Powell JW, Oswald MW, et al. (2008) Iodinated contrast media: effect of osmolarity and injection temperature on erythrocyte morphology in vitro. Acta Radiol 49: 337–343. [DOI] [PubMed] [Google Scholar]
- 12. Lerche D, Hennicke G (1992) The effects of different ionic and non-ionic X-ray contrast media on the morphological and rheological properties of human red blood cells. Clinical Hemorheology and Microcirculation 12: 341–355. [Google Scholar]
- 13. Losco P, Nash G, Stone P, Ventre J (2001) Comparison of the effects of radiographic contrast media on dehydration and filterability of red blood cells from donors homozygous for hemoglobin A or hemoglobin S. Am J Hematol. 68: 149–158. [DOI] [PubMed] [Google Scholar]
- 14. Strickland NH, Rampling MW, Dawson P, Martin G (1992) Contrast Media-Induced Effects on Blood Rheology and Their Importance in Angiography. Clinical Radiology 45: 240–242. [DOI] [PubMed] [Google Scholar]
- 15. Franke R, Minkow A, Hiebl B, Fuhrmann R, Mrowietz C, et al. (2010) Embedding of radiographic media molecules in the membrane of erythrocytes. Clin Hemorheol Microcirc 46: 225–232. [DOI] [PubMed] [Google Scholar]
- 16. Reinhart WH, Sung LP, Sung KL, Bernstein SE, Chien S (1988) Impaired echinocytic transformation of ankyrin- and spectrin-deficient erythrocytes in mice. Am J Hematol 29: 195–200. [DOI] [PubMed] [Google Scholar]
- 17. Becker PS, Lux SE (1985) Hereditary spherocytosis and related disorders. Clin Haematol 14: 15–43. [PubMed] [Google Scholar]
- 18. Bennett V, Baines AJ (2001) Spectrin and ankyrin-based pathways: metazoan inventions for integrating cells into tissues. Physiol Rev 81: 1353–1392. [DOI] [PubMed] [Google Scholar]
- 19. Jiang J, Magilnick N, Tsirulnikov K, Abuladze N, Atanasov I, et al. (2013) Single particle electron microscopy analysis of the bovine anion exchanger 1 reveals a flexible linker connecting the cytoplasmic and membrane domains. PLoS One 8: e55408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Anong WA, Franco T, Chu H, Weis TL, Devlin EE, et al. (2009) Adducin forms a bridge between the erythrocyte membrane and its cytoskeleton and regulates membrane cohesion. Blood 114: 1904–1912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Benninghoff A (1994) Anatomie Band 1.; Drenckhahn D, Zenker W, editors: Urban & Schwarzenberg, München - Baltimore.
- 22. Nestor MW, Cai X, Stone MR, Bloch RJ, Thompson SM (2011) The actin binding domain of betaI-spectrin regulates the morphological and functional dynamics of dendritic spines. PLoS One 6: e16197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Wolny M, Grzybek M, Bok E, Chorzalska A, Lenoir M, et al. (2011) Key amino acid residues of ankyrin-sensitive phosphatidylethanolamine/phosphatidylcholine-lipid binding site of betaI-spectrin. PLoS One 6: e21538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Stefanovic M, Markham NO, Parry EM, Garrett-Beal LJ, Cline AP, et al. (2007) An 11-amino acid beta-hairpin loop in the cytoplasmic domain of band 3 is responsible for ankyrin binding in mouse erythrocytes. Proc Natl Acad Sci U S A 104: 13972–13977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Connes P, Machado R, Hue O, Reid H (2011) Exercise limitation, exercise testing and exercise recommendations in sickle cell anemia. Clin Hemorheol Microcirc 49: 151–163. [DOI] [PubMed] [Google Scholar]
- 26. Lamarre Y, Petres S, Hardy-Dessources MD, Sinnapah S, Romana M, et al. (2012) Abnormal flow adhesion of sickle red blood cells to human placental trophoblast extracellular matrix. Clin Hemorheol Microcirc 51: 229–234. [DOI] [PubMed] [Google Scholar]
- 27. Low PS, Waugh SM, Zinke K, Drenckhahn D (1985) The role of hemoglobin denaturation and band 3 clustering in red blood cell aging. Science 227: 531–533. [DOI] [PubMed] [Google Scholar]
- 28. Palek J, Sahr KE (1992) Mutations of the red blood cell membrane proteins: from clinical evaluation to detection of the underlying genetic defect. Blood 80: 308–330. [PubMed] [Google Scholar]
- 29. Reinhart WH (2011) Peculiar red cell shapes: Fahraeus Lecture 2011. Clin Hemorheol Microcirc 49: 11–27. [DOI] [PubMed] [Google Scholar]
- 30. Southgate CD, Chishti AH, Mitchell B, Yi SJ, Palek J (1996) Targeted disruption of the murine erythroid band 3 gene results in spherocytosis and severe haemolytic anaemia despite a normal membrane skeleton. Nat Genet 14: 227–230. [DOI] [PubMed] [Google Scholar]
- 31. Khan AA, Hanada T, Mohseni M, Jeong JJ, Zeng L, et al. (2008) Dematin and adducin provide a novel link between the spectrin cytoskeleton and human erythrocyte membrane by directly interacting with glucose transporter-1. J Biol Chem 283: 14600–14609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Salomao M, Zhang X, Yang Y, Lee S, Hartwig JH, et al. (2008) Protein 4.1R-dependent multiprotein complex: new insights into the structural organization of the red blood cell membrane. Proc Natl Acad Sci U S A 105: 8026–8031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Baskurt OK, Boynard M, Cokelet GC, Connes P, Cooke BM, et al. (2009) New guidelines for hemorheological laboratory techniques. Clin Hemorheol Microcirc 42: 75–97. [DOI] [PubMed] [Google Scholar]
- 34. Jackson CJ, Garbett PK, Nissen B, Schrieber L (1990) Binding of human endothelium to Ulex europaeus I-coated Dynabeads: application to the isolation of microvascular endothelium. J Cell Sci 96: 257–262. [DOI] [PubMed] [Google Scholar]
- 35. Noria S, Xu F, McCue S, Jones M, Gotlieb AI, et al. (2004) Assembly and reorientation of stress fibers drives morphological changes to endothelial cells exposed to shear stress. Am J Pathol 164: 1211–1223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Franke RP, Scharnweber T, Fuhrmann R, Mrowietz C, Jung F (2013) Effect of radiographic contrast media (Iodixanol, Iopromide) on the spectrin/actin-network of the membranous cytoskeleton of erythrocytes. Clin Hemorheol Microcirc 54: 273–285. [DOI] [PubMed] [Google Scholar]
- 37. Nightingale TD, Cutler DF, Cramer LP (2012) Actin coats and rings promote regulated exocytosis. Trends Cell Biol 22: 329–337. [DOI] [PubMed] [Google Scholar]
- 38. Luginbühl A, Nikolic M, Beck HP, Wahlgren M, Lutz HU (2007) Complement factor D, albumin, and immunoglobulin G anti-band 3 protein antibodies mimic serum in promoting rosetting of malaria-infected red blood cells. Infect Immun 75: 1771–1777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Deftereos S, Giannopoulos G, Kossyvakis C, Raisakis K, Kaoukis A, et al. (2009) Effect of radiographic contrast media on markers of complement activation and apoptosis in patients with chronic coronary artery disease undergoing coronary angiography. J Invasive Cardiol 21: 473–477. [PubMed] [Google Scholar]
- 40.Vik H, Froysa A, Sonstevold A, Toft K, Stov PS, et al.. (1995) Complement activation and histamine release following administration of roentgen contrast media. Acta Radiol Suppl 399: 83–89. [DOI] [PubMed]
- 41. Aspelin P, Schmid-Schonbein H (1978) Effect of ionic and non-ionic contrast media on red cell aggregation in vitro. Acta Radiol Diagn (Stockh) 19: 766–784. [DOI] [PubMed] [Google Scholar]
- 42. Mrowietz C, Franke RP, Jung F (2012) Influence of different radiographic contrast media on the echinocyte formation of human erythrocytes. Clin Hemorheol Microcirc 50: 35–47. [DOI] [PubMed] [Google Scholar]
- 43. Chasis JA, Mohandas N (1986) Erythrocyte membrane deformability and stability: two distinct membrane properties that are independently regulated by skeletal protein associations. J Cell Biol 103: 343–350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Blanc L, Salomao M, Guo X, An X, Gratzer W, et al. (2010) Control of erythrocyte membrane-skeletal cohesion by the spectrin-membrane linkage. Biochemistry 49: 4516–4523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Mohandas N, Gallagher PG (2008) Red cell membrane: past, present, and future. Blood 112: 3939–3948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Johnson CP, Tang HY, Carag C, Speicher DW, Discher DE (2007) Forced unfolding of proteins within cells. Science 317: 663–666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Bruce LJ, Beckmann R, Ribeiro ML, Peters LL, Chasis JA, et al. (2003) A band 3-based macrocomplex of integral and peripheral proteins in the RBC membrane. Blood 101: 4180–4188. [DOI] [PubMed] [Google Scholar]
- 48. Endeward V, Cartron JP, Ripoche P, Gros G (2008) RhAG protein of the Rhesus complex is a CO2 channel in the human red cell membrane. FASEB J 22: 64–73. [DOI] [PubMed] [Google Scholar]
- 49. Sowah D, Casey JR (2011) An intramolecular transport metabolon: fusion of carbonic anhydrase II to the COOH terminus of the Cl(-)/HCO(3)(-)exchanger, AE1. Am J Physiol Cell Physiol 301: C336–346. [DOI] [PubMed] [Google Scholar]
- 50. Sterling D, Reithmeier RA, Casey JR (2001) A transport metabolon. Functional interaction of carbonic anhydrase II and chloride/bicarbonate exchangers. J Biol Chem 276: 47886–47894. [DOI] [PubMed] [Google Scholar]
- 51. Chu H, Puchulu-Campanella E, Galan JA, Tao WA, Low PS, et al. (2012) Identification of cytoskeletal elements enclosing the ATP pools that fuel human red blood cell membrane cation pumps. Proc Natl Acad Sci U S A 109: 12794–12799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Hoffman JF, Dodson A, Proverbio F (2009) On the functional use of the membrane compartmentalized pool of ATP by the Na+ and Ca++ pumps in human red blood cell ghosts. J Gen Physiol 134: 351–361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Tiffert T, Lew VL (2011) Elevated intracellular Ca2+ reveals a functional membrane nucleotide pool in intact human red blood cells. J Gen Physiol 138: 381–391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Campanella ME, Chu H, Low PS (2005) Assembly and regulation of a glycolytic enzyme complex on the human erythrocyte membrane. Proc Natl Acad Sci U S A 102: 2402–2407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Puchulu-Campanella E, Chu H, Anstee DJ, Galan JA, Tao WA, et al. (2013) Identification of the components of a glycolytic enzyme metabolon on the human red blood cell membrane. J Biol Chem 288: 848–858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Sega MF, Chu H, Christian J, Low PS (2012) Interaction of deoxyhemoglobin with the cytoplasmic domain of murine erythrocyte band 3. Biochemistry 51: 3264–3272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Ferru E, Giger K, Pantaleo A, Campanella E, Grey J, et al. (2011) Regulation of membrane-cytoskeletal interactions by tyrosine phosphorylation of erythrocyte band 3. Blood 117: 5998–6006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Lopes de Almeida JP, Freitas-Santos T, Saldanha C (2012) Erythrocyte deformability dependence on band 3 protein in an in-vitro model of hyperfibrinogenemia. Clin Hemorheol Microcirc 50: 213–219. [DOI] [PubMed] [Google Scholar]
- 59. Sprague RS, Stephenson AH, Ellsworth ML (2007) Red not dead: signaling in and from erythrocytes. Trends Endocrinol Metab 18: 350–355. [DOI] [PubMed] [Google Scholar]
- 60. Bach R, Jung F, Scheller B, Hummel B, Ozbek C, et al. (1996) Influence of a non-ionic radiography contrast medium on the microcirculation. Acta Radiol 37: 214–217. [DOI] [PubMed] [Google Scholar]
- 61. Spitzer S, Munster W, Sternitzky R, Bach R, Jung F (1999) Influence of Iodixanol-270 and Iopentol-150 on the microcirculation in man: influence of viscosity on capillary perfusion. Clin Hemorheol Microcirc 20: 49–55. [PubMed] [Google Scholar]
- 62. Jung F, Matschke K, Mrowietz C, Tugtekin SM, Geissler T, et al. (2003) Influence of radiographic contrast media on myocardial tissue oxygen tension: NaCl-controlled, randomised, comparative study of iohexol versus iopromide in an animal model. Clin Hemorheol Microcirc 29: 53–61. [PubMed] [Google Scholar]
- 63. Matschke K, Gerk U, Mrowietz C, Park JW, Jung F (2007) Influence of radiographic contrast media on myocardial oxygen tension: a randomized, NaCl-controlled comparative study of iodixanol versus iomeprol in pigs. Acta Radiol 48: 292–299. [DOI] [PubMed] [Google Scholar]
- 64. Antonelou MH, Kriebardis AG, Stamoulis KE, Trougakos IP, Papassideri IS (2011) Apolipoprotein J/clusterin in human erythrocytes is involved in the molecular process of defected material disposal during vesiculation. PLoS One 6: e26033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Bell AJ, Satchwell TJ, Heesom KJ, Hawley BR, Kupzig S, et al. (2013) Protein distribution during human erythroblast enucleation in vitro. PLoS One 8: e60300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Mrowietz C, Hiebl B, Franke RP, Park JW, Jung F (2008) Reversibility of echinocyte formation after contact of erythrocytes with various radiographic contrast media. Clin Hemorheol Microcirc 39: 281–286. [PubMed] [Google Scholar]
- 67.Gershon D, Glass GA, Gershon H (1988) The Effect of Host and Cell Age on the Rat Erythrocyte: Biochemical Aspects. In: Platt D, editor. Blood Cells, Rheology, and Aging: Springer Berlin Heidelberg. 42–50.
- 68.Platt D, Rieck W (1988) Red Cell Membrane Proteins, Glycoproteins, and Aging. In: Platt D, editor. Blood Cells, Rheology, and Aging: Springer Berlin Heidelberg. 29–41.