

Abstract
Deficits in memory for everyday activities are common complaints among healthy and demented older adults. The medial temporal lobes and dorsolateral prefrontal cortex are both affected by aging and early-stage Alzheimer's disease, and are known to influence performance on laboratory memory tasks. We investigated whether the volume of these structures predicts everyday memory. Cognitively healthy older adults and older adults with mild Alzheimer's-type dementia watched movies of everyday activities and completed memory tests on the activities. Structural MRI was used to measure brain volume. Medial temporal but not prefrontal volume strongly predicted subsequent memory. Everyday memory depends on segmenting activity into discrete events during perception, and medial temporal volume partially accounted for the relationship between performance on the memory tests and performance on an event-segmentation task. The everyday-memory measures used in this study involve retrieval of episodic and semantic information as well as working memory updating. Thus, the current findings suggest that during perception, the medial temporal lobes support the construction of event representations that determine subsequent memory.
Keywords: perception, memory, cognitive neuroscience, aging
Subjective memory complaints commonly reported by older adults include forgetting the location of items such as keys, forgetting directions to familiar locations, and repeating stories (Gilewski, Zelinski, & Schaie, 1990). Further, caregivers report that older adults with dementia have trouble remembering addresses and phone numbers, and knowing how to work familiar machines (Jorm & Jacomb, 1989). These subjective impressions are often accurate: Older adults show objective declines in memory for things such as people's names, items on a grocery list, and information in a news program (West, Crook, & Barron, 1992).
Everyday memory1 involves remembering information from events that occur naturalistically in day-to-day life. It can be distinguished from memory as assessed by typical laboratory tasks (e.g., learning word lists, picture lists, or word pairs) on two related dimensions: (a) The memoranda are richer and more structured than those in laboratory tasks, and (b) knowledge about everyday events often is encoded into preexisting knowledge structures.
What are the cognitive and neural mechanisms that influence everyday memory? First, general memory systems, including those measured by traditional laboratory tests of episodic and working memory, surely play an important role. Performance on such measures declines with age (R. T. Zacks, Hasher, & Li, 2000), and these declines could lead to individual differences in older adults' everyday memory. Second, everyday memory may depend on mechanisms that deal specifically with everyday memory's rich and knowledge-driven character.
Older adults have demonstrated better memory on tests that provide more contextual support and structure (for a review, see R. T. Zacks et al., 2000). Crystallized knowledge increases with age (Horn & Cattell, 1967), so older adults should be able to use this knowledge on everyday-memory tasks. In fact, scripts and schemas (Hess, 1990) and expertise (Morrow, Leirer, & Altieri, 1992) help older adults' (and younger adults') memory performance. To the degree that older adults can capitalize on structure and knowledge, this should facilitate remembering. For example, when people try to remember a news report about a birth at the zoo, their success may depend on the degree to which they can form and maintain episodic long-term memories and manipulate phrases (“baby elephant,” “40 hours in labor”) in working memory. But their success also may depend on the degree to which they can construct effective representations of the structured events in the report and use their knowledge about zoos and births to populate those representations.
Neurophysiologically, the dorsolateral prefrontal cortex (DLPFC) is known to support working memory (Goldman-Rakic, 1995). Thus, differences in DLPFC volume may be related to differences in everyday-memory performance. Another brain region that may support everyday memory is the medial temporal lobe (MTL). The MTL is important for episodic long-term memory (e.g., Eichenbaum, Yonelinas, & Ranganath, 2007). In addition, recent evidence has demonstrated that the MTL—in particular, the hippocampus—may be critical for the retrieval of recently presented information (Hannula, Tranel, & Cohen, 2006; Oztekin, Davachi, & McElree, 2010; Swallow et al., 2011; but see Jeneson & Squire, 2012). Both the DLPFC and the MTL atrophy with age, and these declines have been linked to cognitive deficits in older adults (Golomb et al., 1993; Raz, Dixon, Head, Dupuis, & Acker, 1998). Alzheimer's disease is associated with further atrophy in both structures, with the MTL being particularly affected early in the progression (McDonald et al., 2009). Could individual differences in DLPFC and MTL atrophy lead to differences in everyday memory?
Event-Segmentation Theory
Event-segmentation theory (EST; J. M. Zacks, Speer, Swallow, Braver, & Reynolds, 2007) provides a framework for characterizing the contributions of these cognitive and neural mechanisms to everyday-memory performance. According to EST, as people observe ongoing activity, they spontaneously segment it into discrete events. Information relevant to the current event is captured in an event model, which is a working memory representation of “what is happening now.” An event model represents information that is invariant across an event, but it must be updated when one event ends and another begins. EST posits that when an event model is updated, an event boundary is perceived. Event boundaries serve as memory anchors in that they help people chunk complex activity into meaningful events. Individuals' ability to identify event boundaries is related to their memory for events (Kurby & Zacks, 2011; J. M. Zacks, Speer, Vettel, & Jacoby, 2006).
Thus, everyday-memory performance is predicted by one's ability to segment and organize activity as one perceives it. Given that working memory is central to this process, the quality of event segmentation likely depends on the brain structures that support working memory functioning, namely, the DLPFC and MTL. Updating of event models also is influenced by episodic memories of previous events, which may also depend on the MTL. Finally, updating of event models is influenced by semantic knowledge about events (i.e., knowledge divorced from personal experience of the events), which may depend particularly on the prefrontal cortex, including the DLPFC and the medial prefrontal cortex (Grafman, 1995; Krueger, Barbey, & Grafman, 2009).
The Present Study
We assessed everyday memory in cognitively healthy older adults and in older adults with mild Alzheimer's-type dementia. This latter population is ideal for examining everyday memory because individuals vary widely in their self-assessed and objectively assessed everyday-memory ability (Jorm & Jacomb, 1989; West et al., 1992), and because Alzheimer's disease selectively affects some of the mechanisms that may be involved in everyday memory. Older adults in the earliest stages of Alzheimer's disease show decreased MTL volume (Stoub, Rogalski, Leurgans, Bennett, & deToledo-Morrell, 2010) and MTL function (Golby et al., 2005). Further, Alzheimer's patients show changes in prefrontal regions, including increased amyloid deposition (Klunk et al., 2004) and abnormal prefrontal activation (Becker et al., 1996), compared with cognitively healthy older adults. These neurophysiological changes are associated with the hallmark memory impairments of Alzheimer's disease (Golby et al., 2005; Stoub et al., 2010).
Our sample of older adults watched and segmented movies of everyday activities (e.g., preparing breakfast) and then completed tests of memory for the activities. Volumetric data from structural MRI scans also were obtained for the participants. Our main goal was to evaluate whether event segmentation predicted memory for everyday events, and whether this relationship was dependent on the integrity of the DLPFC, the MTL, or both.
Method
Participants
All participants were recruited through the Knight Alzheimer's Disease Research Center at Washington University in St. Louis. The presence of dementia was assessed according to the Clinical Dementia Rating (CDR) scale (Morris, 1993). The CDR is based on a 90-min clinical interview of the participant and a collateral source (often a spouse, child, or close friend). This interview assesses changes in cognitive and functional abilities in the areas of memory, orientation, judgment and problem solving, community affairs, hobbies, and personal care. CDR scores range from 0 to 2, with 0 indicating no dementia, 0.5 indicating questionable or very mild dementia, 1 indicating mild dementia, and 2 indicating moderate dementia. Participants were excluded if they had any of a wide range of neurological disorders (e.g., Parkinson's disease, Huntington's disease), neurological damage (e.g., due to seizures or head trauma), other types of dementia (e.g., vascular, Lewy bodies), cerebrovascular disease, or depression. We scores of 0.5 (15 females, 26 males), and 21 individuals with CDR scores of 1 (7 females, 14 males).
Materials
Event segmentation
Participants watched four movies: one practice and three experimental movies. The practice movie involved a male actor building a ship out of Legos (duration = 155 s). The experimental movies involved a female actor preparing breakfast (329 s), a male actor decorating for a party (376 s), and a female actor checking out a book at a library (249 s). Participants watched and segmented each movie twice—once at a coarse grain and once at a fine grain. As they watched the movies, participants were instructed to press the space bar on a computer keyboard each time they thought one large (coarse grain) or small (fine grain) meaningful unit of activity ended and another began (see Segmentation Procedure in the Supplemental Material available online). We measured segmentation agreement, the extent to which a participant's segmentation locations correlated with the normative2 segmentation locations of another sample. Segmentation agreement was calculated with point-biserial correlations that were scaled to control for individual differences in the number of segmentation locations identified (see Kurby & Zacks, 2011). Segmentation agreement can range from 0 to 1, with larger values indicating better agreement with the group, and was averaged across grains.
Everyday memory
Participants completed three measures of memory for the events in each movie. The first measure was recall. For this test, participants orally described the activity in the movie. Each movie was broken down into activity units according to the Action Coding System described by Schwartz (1991), and recall was scored as the number of units reported. Units were finegrained parts of an activity, termed “A1” units by Schwartz (e.g., “she walked to the sink,” “turned on the water,” “put soap on her hands,” etc.). The second measure was a forced-choice recognition task in which on each trial, participants were presented with two still photographs— one from the movie they had watched and one from a similar but different movie. The test for each movie had 20 trials, and recognition was scored as the proportion of correct responses. Finally, participants completed an order-memory task. For each movie, they were given 12 still photographs that were taken from the movie but were out of temporal order. Participants were instructed to rearrange the photographs in the order in which they had occurred in the movie. Order memory was scored as an error measure, which was the mean absolute deviation of each picture from the correct position.
MRI acquisition and analysis
Participation at the Knight Alzheimer's Disease Research Center includes structural MRI scans every other year. When a participant had undergone multiple scans, we used the most recent scan that had occurred prior to the behavioral testing. However, some participants (n = 28) had never completed a scan, so the sample included volume estimates for 28 participants with CDR scores of 0, 31 participants with CDR scores of 0.5, and 15 participants with CDR scores of 1.3 T1-weighted scans, with MP-RAGE (magnetization-prepared rapid acquisition with gradient echo) imaging, were obtained for each of these subjects (repetition time, or TR = 9.7 ms; echo time, or TE = 4 ms; inversion time, or TI = 20 ms; 1 mm × 1 mm × 1.25 mm resolution).
Gray matter volume estimates were obtained using FreeSurfer 5.1 image analysis suite (http://surfer.nmr.mgh .harvard.edu/; Fischl, 2012), and regions of interest (ROIs) were based on the Desikan–Killiany atlas (Desikan et al., 2006). ROIs potentially associated with everyday memory were the DLPFC, which was defined as the rostral middle frontal gyrus,4 and MTL, which was made up of the entorhinal cortex, hippocampus, and parahippocampal gyrus regions (which include the perirhinal and parahippocampal cortex). Volumes were summed across hemispheres and then normalized to control for intracranial volume using linear regression (e.g., Buckner et al., 2004).
Cognitive ability battery
A cognitive ability battery (ELSMEM; Storandt, Balota, & Salthouse, 2009) designed to assess a broad spectrum of abilities was administered to all participants, usually 1 to 2 weeks after their annual clinical assessment. We obtained participants' scores on the episodic memory tasks: the sum of scores on the three free-recall trials from the Selective Reminding Test (Grober & Buschke, 1987), the Verbal Paired Associates score from the Wechsler Memory Scale (WMS; Wechsler & Stone, 1973), and the immediate- and delayed-recall scores from the WMS Logical Memory test. (See Table S1 in the Supplemental Material for descriptive statistics.)
Procedure
Participants were seated at a laptop computer and practiced the coarse-grained event-segmentation task using the example movie. The experimenter answered any questions and restated the instructions. Then, participants segmented the breakfast, party, and library movies, respectively. Following each movie, participants completed the recall task, the forced-choice recognition task, and finally the order-memory task. After the third movie, they completed a short form of the Naturalistic Action Test (Schwartz, Segal, Veramonti, Ferraro, & Buxbaum, 2002), followed by a script-knowledge test based on the procedure described by Rosen, Caplan, Sheesley, Rodriguez, and Grafman (2003). Next, participants watched each movie—including the example movie—a second time and segmented the events at a fine grain. (No memory tests followed this viewing.) Finally, participants provided a saliva sample for DNA analyses. (The Naturalistic Action Test, script-knowledge test, and DNA analyses were for separate projects and are not discussed further.)
Data preparation
We screened each variable for values more than 3.5 standard deviations away from the sample mean (univariate outliers); 2 values (< 0.1% of the data) met this criterion. We replaced these extreme values and missing values (< 2% of the data) using the expectation maximization (EM) procedure in SPSS 19.0. All variables were approximately normally distributed (skewness < |1.5|, kurtosis < |1.5|).
Results
Descriptive statistics for the event-segmentation and memory measures, as well as volume estimates for the ROIs, are presented by CDR status in Table 1. On all behavioral measures, participants with CDR scores of 0 performed best, and those with CDR scores of 1 performed worst, ts(54–79) = 4.86–8.11, ps < .001.
Table 1. Descriptive Statistics for Age, Segmentation Agreement, Everyday Memory, and Brain Volume.
| Variable | CDR score = 0 | CDR score = 0.5 | CDR score = 1 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
||||||||||
| M | Median | Range | d (CDR = 0 vs. 0.5)a | M | Median | Range | d (CDR =0.5 vs. l)a | M | Median | Range | d (CDR = 0 vs. l)a | |
| Age (years) | 74.65 | 75.00 | 65–86 | −0.55 | 77.83 | 78.00 | 66–86 | 0.05 | 77.55 | 78.00 | 63–90 | −0.47 |
| Segmentation agreement | ||||||||||||
| Breakfast | .38 | .34 | .24–.58 | 0.20 | .36 | .33 | .07–.59 | 0.62 | .28 | .29 | .00–.46 | 0.84 |
| Library | .45 | .45 | .27–.61 | 0.72 | .39 | .40 | .18–.64 | 0.72 | .30 | .32 | .05–.52 | 1.33 |
| Party | .43 | .42 | .19–.68 | 0.59 | .36 | .35 | .03–.59 | 0.78 | .28 | .29 | .04–.40 | 1.36 |
| Recall | ||||||||||||
| Breakfast | 22.18 | 21.00 | 1–61 | 0.88 | 13.10 | 13.00 | 0–31 | 1.26 | 4.24 | 3.00 | 0–23 | 1.94 |
| Library | 13.03 | 12.50 | 1–34 | 0.64 | 6.78 | 6.00 | 0–24 | 1.29 | 1.81 | 2.00 | 0–10 | 1.86 |
| Party | 26.65 | 22.50 | 1–74 | 0.77 | 13.24 | 10.00 | 0–57 | 1.00 | 2.76 | 1.00 | 0–12 | 1.65 |
| Recognition | ||||||||||||
| Breakfast | .71 | .70 | .40–.90 | 0.50 | .65 | .65 | .30–.90 | 1.09 | .50 | .50 | .30–.80 | 1.60 |
| Library | .77 | .80 | .20–.95 | 0.33 | .72 | .70 | .35–1.0 | 1.12 | .55 | .53 | .25–.80 | 1.58 |
| Party | .88 | .90 | .20–.1.0 | 0.66 | .78 | .80 | .40–1.0 | 1.26 | .60 | .60 | .40–.90 | 1.90 |
| Order memory (errors) | ||||||||||||
| Breakfast | 0.66 | 0.33 | 0.00–4.5 | −0.74 | 1.41 | 1.00 | 0.00–4.5 | −1.44 | 3.24 | 3.00 | 0.83–5.5 | −2.20 |
| Library | 1.48 | 1.50 | 0.00–4.3 | −0.99 | 2.58 | 2.50 | 0.50–4.5 | −1.05 | 3.70 | 3.83 | 1.0–5.0 | −2.21 |
| Party | 0.12 | 0.00 | 0.00–0.83 | −0.37 | 0.23 | 0.17 | 0.00–1.8 | −1.36 | 1.54 | 1.33 | 0.00–4.7 | −1.51 |
| Volume (mm3) | ||||||||||||
| Entorhinal cortex | 3,926 | 3,927 | 2,821–5,203 | 0.71 | 3,469 | 3,397 | 2,084–4,597 | 1.38 | 2,544 | 2,574 | 1,615–3,721 | 2.18 |
| Hippocampus | 7,526 | 7,528 | 5,836–9,056 | 1.18 | 6,491 | 6,316 | 5,001–8,403 | 0.99 | 5,671 | 5,685 | 4,730–6,734 | 2.71 |
| Parahippocampal gyrus | 3,662 | 3,645 | 2,814–4,693 | 0.34 | 3,478 | 3,473 | 2,052–4,542 | 0.87 | 2,987 | 2,965 | 1,871–3,846 | 1.38 |
| DLPFC | 27,294 | 27,304 | 23,511–32,853 | 0.46 | 25,951 | 25,467 | 19,441–31,890 | −0.15 | 26,382 | 26,385 | 22,659–32,247 | 0.35 |
Note: CDR = Clinical Dementia Rating scale (Morris, 1993); DLPFC = dorsolateral prefrontal cortex.
The effect size between the two indicated CDR groups was calculated using Cohen's d
Correlations and factor analyses
Table 2 presents the correlations among the measures of event segmentation, memory, and brain volume. As shown in the table, the segmentation-agreement variables correlated positively with each other, as did the everyday-memory variables. (For correlations among the individual memory measures, see Table S2 in the Supplemental Material.) We entered these variables into an exploratory factor analysis, and two clearly interpretable factors emerged (Table 3). The MTL volume variables correlated positively with each other (rs = .48–.69), and when these variables were entered into an exploratory factor analysis, a single factor accounted for 74.1% of the variance. (DLPFC volume correlated weakly with MTL volume.)
Table 2. Correlations Between Segmentation Agreement, Everyday Memory, and Brain Volume.
| Variable | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|
| Segmentation agreement | |||||||||
| 1. Breakfast | — | ||||||||
| 2. Library | .71 | — | |||||||
| 3. Party | .53 | .57 | — | ||||||
| Everyday memory | |||||||||
| 4. Breakfast | .41 | .43 | .26 | — | |||||
| 5. Library | .38 | .37 | .32 | .82 | — | ||||
| 6. Party | .38 | .40 | .32 | .87 | .81 | — | |||
| Brain volume | |||||||||
| 7. Entorhinal cortex | .16 | .26 | .30 | .49 | .55 | .53 | — | ||
| 8. Hippocampus | .18 | .39 | .24 | .45 | .45 | .47 | .66 | — | |
| 9. Parahippocampal gyrus | .14 | .23 | .19 | .47 | .39 | .47 | .69 | .48 | — |
| 10. Dorsolateral prefrontal cortex | .09 | .10 | .14 | .16 | .10 | .12 | .17 | .22 | .27 |
Note: N = 74. For segmentation agreement, performance was averaged across the coarse-grained and fine-grained segmentation tasks for each movie. For everyday memory, performance across the recall, recognition, and order-memory tests was averaged for each movie. Significant correlations (p < .05) are in boldface.
Table 3. Results of the Exploratory Factor Analysis.
| Variable | Factor loading | h2 | |
|---|---|---|---|
|
| |||
| Factor 1 | Factor 2 | ||
| Everyday memory | |||
| Breakfast | .00 | .94 | .76 |
| Library | .02 | .86 | .81 |
| Party | .00 | .94 | .84 |
| Segmentation agreement | |||
| Breakfast | .80 | .02 | .64 |
| Library | .88 | .00 | .69 |
| Party | .65 | .00 | .15 |
Note: The exploratory factor analysis was conducted using Principal axis as the extraction method and Promax as the rotation method. h2 = communality estimate. For everyday memory, performance across the recall, recognition, and order-memory tests was averaged for each movie. For segmentation agreement, performance was averaged across the coarse-grained and fine-grained segmentation tasks for each movie.
On the basis of these results, we created unit-weighted composite variables by averaging the z scores. The segmentation-agreement composite was the average of the z-scored agreement values from the breakfast, library, and party movies. The everyday-memory composite was the average of the recall, recognition, and order-memory z scores for the three movies. Finally, the MTL composite consisted of the average z-scored volumes for the entorhinal cortex, parahippocampal gyrus, and hippocampus. Reliability estimates for the composites were acceptably high (αs > .79).
Regression analyses
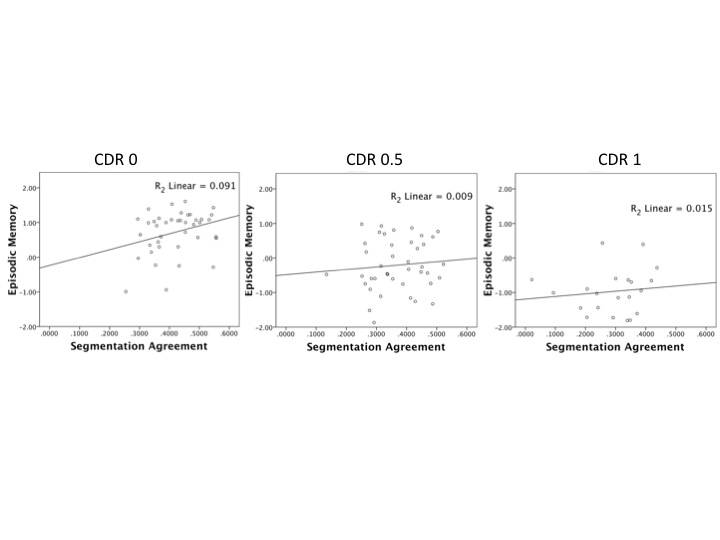
Segmentation agreement correlated significantly with everyday memory across all participants (r = .45, p < .001; for scatter plots by CDR group, see Fig. S1 in the Supplemental Material; see Fig. S2 in the Supplemental Material for scatter plots illustrating the relationship between segmentation agreement and episodic memory in the three CDR groups). MTL volume also correlated significantly with everyday memory (r = .58, p < .001). By contrast, DLPFC volume correlated weakly with everyday memory (r = .13, p = .13). We performed regression analyses to determine whether MTL volume accounted for the relationship between segmentation agreement and everyday memory.5 In the first analysis, we regressed everyday memory on segmentation agreement. In the second analysis, we regressed everyday memory on segmentation agreement after entering MTL volume into the model. In the third analysis, we regressed everyday memory on MTL volume after entering segmentation agreement in the model.
The results are summarized in Figure 1a. Segmentation agreement accounted for 20.2% of the variance in everyday memory. This value dropped to 7.2% in the model statistically controlling for MTL volume, a 64% reduction, F(1, 71) = 11.57, p < .001. Regression analyses also were conducted to examine the effect of the individual medial temporal regions on the individual memory measures. Each region accounted for a large portion of the relationship between segmentation agreement and each memory measure (see Table S4 in the Supplemental Material). In addition, everyday memory was significantly and independently predicted by segmentation agreement (7.2%), F(1, 71) = 8.68, p = .004, and MTL volume (21.1%), F(1, 71) = 25.58, p < .001.
Fig. 1.

Venn diagrams depicting relationships among (a) segmentation agreement, everyday memory, and medial temporal lobe (MTL) volume and (b) segmentation agreement, everyday memory, and score on the Clinical Dementia Rating (CDR) scale (Morris, 1993).
Given the high correlation between CDR score and MTL volume (r = −.64, p = .001), we repeated the regression analyses replacing MTL volume with CDR status.
The results are displayed in Figure 1b. The variance in everyday memory accounted for by segmentation agreement (20.2%) dropped to 4.3% in the model statistically controlling for CDR status, a 79% reduction, F(1, 71) = 14.51, p < .001. In addition, everyday memory was significantly and independently predicted by segmentation agreement (4.3%), F(1, 71) = 5.78, p = .019, and CDR status (27.2%), F(1, 71) = 36.66, p < .001.
Discussion
The two main goals of the current study were to evaluate whether (a) event segmentation predicts everyday memory and (b) MTL or DLPFC volume accounts for this relationship. Individual differences in segmentation agreement predicted individual differences in everyday memory. Participants whose segmentation agreement was more normative remembered more of the activity in the movies. These results replicate those of J. M. Zacks et al. (2006) and extend them to a more demented group of older adults.
The finding that event segmentation predicted everyday memory in the most demented group (see Fig. S1 in the Supplemental Material) is important because these participants sometimes had difficulty remembering any information from the movies even after the short retention intervals. Approximately 2.5% of the cognitively healthy group and 9.8% of the very mildly demented group recalled no accurate information from the movies, whereas 52.3% of the most demented group recalled no accurate information. Although there was a restricted range in everyday-memory performance in the most demented group, we did observe some variability, and event segmentation accounted for a significant portion of this variability. These findings suggest that the ability to organize an activity during perception may offset the everyday-memory decrements observed in demented older adults. Thus, interventions aimed at improving event segmentation may be beneficial for everyday-memory performance for both cognitively normal and demented adults.
Individual differences in MTL volume accounted for about two thirds of the relationship between event segmentation and memory. Clinical dementia status also significantly accounted for this relationship. Together, MTL volume and dementia status accounted for almost half of the variance in event memory, F(2, 71) = 32.25, p < .001. One possible explanation for why MTL volume and CDR status accounted for shared variance in the relationship between segmentation and memory is that Alzheimer's disease neuropathology in its early stages affects the MTL disproportionately (Golby et al., 2005; Stoub et al., 2010), and that this leads to declines in memory, orientation, judgment, and problem solving. These cognitive declines, in turn, produce higher scores on the CDR scale.
What role might the MTL play in event segmentation and memory? There are several possible mechanistic explanations. First, the MTL could be responsible for episodic representations of previous events that aid in the encoding of current activities. A wealth of research has demonstrated that the MTL supports retrieval from long-term memory (for a review, see Eichenbaum et al., 2007). Preexisting knowledge gained through related experiences should help with the organization and comprehension of novel stimuli. For instance, memories of previous experiences dining out (as well as schemas and scripts) help people predict what a hostess will say when they enter a new restaurant or when they watch a movie of someone else entering a restaurant.
Second, the MTL, and particularly the hippocampus, may be involved in pattern separation (e.g., Bakker, Kirwan, Miller, & Stark, 2008). Dementia-related atrophy in the hippocampus may lead to decreased ability to differentiate between two events, and thereby impair everyday memory.
Third, the MTL has been implicated in binding features together during on-line perception (Hannula, Ryan, Tranel, & Cohen, 2007; Hannula et al., 2006). It is possible that this binding, and thus the MTL, is necessary for constructing effective event models. Impaired construction of event models would lead to less effective event segmentation.
Finally, the MTL may contribute to the maintenance and updating of current event models (i.e., working memory for events). Oztekin et al. (2010) demonstrated that the hippocampus is activated during retrieval tasks that engage working memory. More specifically, they found that the hippocampus is activated during the retrieval of all items on a list except the final item. The final item presumably is maintained in the focus of attention, whereas the other items have been displaced to secondary memory (Unsworth & Engle, 2007) or activated long-term memory (Cowan, 2001). Jeneson and Squire (2012) have argued that the MTL supports retrieval over short retention intervals in the case of information that has recently been displaced from working memory to long-term memory—whether because working memory capacity has been exceeded, attention has been diverted, or there has been a lack of rehearsal.
An important contribution of the present study is that it shows that MTL volume predicts memory for everyday activities. Studies have demonstrated repeatedly that the MTL plays an important role in performance of laboratory tests of episodic memory (see Eichenbaum et al., 2007). In the current study, the correlations between MTL volume and everyday memory (r = .58) were similar to the correlations between MTL volume and performance on laboratory episodic memory tasks (word-list recall: r = .62; logical memory: r = .47; associative learning: r = .53; zs < 0.91, ps > .18).
It is notable that DLPFC volume was not significantly related to event segmentation or to memory (Table 1), which was surprising given the known role of this region in working memory maintenance. One possible explanation for this result is that the DLPFC is not important either for segmenting events or for long-term memory encoding. We think this is unlikely. Another possibility is that our sample did not vary greatly in the degree of DLPFC atrophy. The fact that the three clinical groups did not differ significantly in DLPFC volume, F(2, 73) = 1.59, p = .21, is consistent with this possibility. The lack of group differences in this measure is not particularly surprising because faster rates of prefrontal atrophy are observed during later stages of Alzheimer's disease (McDonald et al., 2009), whereas our participants were at most only mildly demented. It may be that in populations older or more demented than our sample, individual differences in DLPFC atrophy would have more opportunity to show their influence.
Conclusion
Progression of dementia of the Alzheimer's type is associated with a decrease in MTL volume (Dickerson et al., 2009). The current results suggest that MTL neuropathology may lead to a decreased ability to segment and remember everyday activities. However, normative event segmentation was associated with better everyday memory even among mildly demented older adults, and even in those with small MTL volume. This suggests that interventions aimed at strengthening event-segmentation ability may help to ameliorate everyday-memory deficits associated with normal aging and even the early stages of dementing neuropathology.
Supplementary Material
{kind=link}
Acknowledgments
Funding: This research was supported by National Institutes of Health Grant R01 AG031150, Principal Investigator Jeffrey M. Zacks; National Institutes of Health Grant F32 AG039162, Principal Investigator Heather R. Bailey; National Institute on Aging Grants P50 AG05681, P01 AG03991, and P01 AG26276, Principal Investigator John C. Morris; and the generous support of Fred Simmons and Olga Mohan.
Footnotes
Notes: Historically, everyday memory has been a controversial term. Some people use it to refer to memory assessed in field studies involving naturally occurring memoranda, which have been criticized for lack of experimental control (e.g., Banaji & Crowder, 1989). More broadly, the field has used the term in connection with tasks and materials that include features of memory that approximate those important for memory for everyday events. This is how we use the term.
Nearly two thirds of the participants in this study by definition are nonnormative. Thus, we used normative segmentation locations, obtained from a group of healthy older adults, reported in a previous study (Kurby, Sargent, Bailey, & Zacks, 2012).
Images for 7 participants were collected on a Siemens 1.5-T Vision scanner, whereas images for 67 participants were collected on a Siemens 3-T Trio scanner. Volume estimates did not differ by scanner type. Also, controlling for scanner type and the Scanner Type × Volume interaction did not change the amount of variance in the cognitive variables that the volume measures accounted for. Thus, scanner type was not included in any further analyses.
We examined nearby regions (i.e., caudal middle frontal gyrus and the inferior frontal gyrus: pars orbitalis, pars triangularis, and pars opercularis), but volume estimates from these prefrontal regions were moderately correlated with one another, and they did not predict segmentation agreement or everyday memory.
Although our primary focus was on the DLPFC and the MTL, we also examined several regions that EST proposes are involved in everyday memory. In particular, EST proposes that memory updating is triggered by dopaminergic projections from the midbrain, by error monitoring performed by the anterior cingulate cortex, and by event-model maintenance that may depend on posterior association cortex. Structural imaging of the midbrain dopamine system is technically challenging (D'Ardenne, McClure, Nystrom, & Cohen, 2008; Düzel et al., 2009), but identification of its targets in the striatum is straightforward; therefore, we examined caudate and putamen volume. Anterior cingulate cortex was defined as the caudal and rostral anterior cingulate regions in FreeSurfer. Posterior association cortex was defined as the posterior cingulate, cuneus, and precuneus. However, we observed no differences between CDR groups in any of these regions, nor were volumes of these regions correlated with segmentation agreement or everyday-memory performance.
Controlling for age in the regression analyses did not alter the conclusions presented here (see Table S3 in the Supplemental Material).
Declaration of Conflicting Interests: The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Supplemental Material: Additional supporting information may be found at http://pss.sagepub.com/content/by/supplemental-data
References
- Bakker A, Kirwan B, Miller M, Stark CE. Pattern separation in the human hippocampal CA3 and dentate gyrus. Science. 2008;319:1640–1642. doi: 10.1126/science.1152882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banaji MR, Crowder RG. The bankruptcy of everyday memory. American Psychologist. 1989;44:1185–1193. [Google Scholar]
- Becker JT, Mintun MA, Aleva K, Wiseman MB, Nichols T, DeKosky ST. Compensatory reallocation of brain resources supporting verbal episodic memory in Alzheimer's disease. Neurology. 1996;46:692–700. doi: 10.1212/wnl.46.3.692. [DOI] [PubMed] [Google Scholar]
- Buckner RL, Head D, Parker J, Fotenos AF, Marcus D, Morris JC, Snyder AZ. A unified approach for morphometric and functional data analysis in young, old, and demented adults using automated atlas-based head size normalization: Reliability and validation against manual measurement of total intracranial volume. NeuroImage. 2004;23:724–738. doi: 10.1016/j.neuroimage.2004.06.018. [DOI] [PubMed] [Google Scholar]
- Cowan N. The magical number 4 in short-term memory: A reconsideration of mental storage capacity. Behavioral and Brain Sciences. 2001;24:87–114. doi: 10.1017/s0140525x01003922. [DOI] [PubMed] [Google Scholar]
- D'Ardenne K, McClure S, Nystrom L, Cohen J. BOLD responses reflecting dopaminergic signals in the human ventral tegmental area. Science. 2008;319:1264–1267. doi: 10.1126/science.1150605. [DOI] [PubMed] [Google Scholar]
- Desikan RS, Segonne F, Fischl B, Quinn BT, Dickerson BC, Blacker D, Killiany RJ. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. NeuroImage. 2006;31:968–980. doi: 10.1016/j.neuroimage.2006.01.021. [DOI] [PubMed] [Google Scholar]
- Dickerson BC, Feczko E, Augustinack JC, Pacheco J, Morris JC, Fischl B, Buckner RL. Differential effects of aging and Alzheimer's disease on medial temporal lobe cortical thickness and surface area. Neurobiology of Aging. 2009;30:432–440. doi: 10.1016/j.neurobiolaging.2007.07.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Düzel E, Bunzeck N, Guitart-Masip M, Wittmann B, Schott BH, Tobler PN. Functional imaging of the human dopaminergic midbrain. Trends in Neurosciences. 2009;32:321–328. doi: 10.1016/j.tins.2009.02.005. [DOI] [PubMed] [Google Scholar]
- Eichenbaum H, Yonelinas AP, Ranganath C. The medial temporal lobe and recognition memory. Annual Review of Neuroscience. 2007;30:123–152. doi: 10.1146/annurev.neuro.30.051606.094328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischl B. Freesurfer. NeuroImage. 2012;62:774–781. doi: 10.1016/j.neuroimage.2012.01.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilewski MJ, Zelinski EM, Schaie KW. The Memory Functioning Questionnaire for assessment of memory complaints in adulthood and old age. Psychology and Aging. 1990;5:482–490. doi: 10.1037//0882-7974.5.4.482. [DOI] [PubMed] [Google Scholar]
- Golby A, Silverberg G, Race E, Gabrieli S, O'Shea J, Knierim K, Gabrieli J. Memory encoding in Alzheimer's disease: An fMRI study of explicit and implicit memory. Brain. 2005;128:773–787. doi: 10.1093/brain/awh400. [DOI] [PubMed] [Google Scholar]
- Goldman-Rakic PS. Cellular basis of working memory. Neuron. 1995;14:477–485. doi: 10.1016/0896-6273(95)90304-6. [DOI] [PubMed] [Google Scholar]
- Golomb J, de Leon MJ, Kluger A, George AE, Tarshish C, Ferris SH. Hippocampal atrophy in normal aging: An association with recent memory impairment. Archives of Neurology. 1993;50:967–973. doi: 10.1001/archneur.1993.00540090066012. [DOI] [PubMed] [Google Scholar]
- Grafman J. Similarities and distinctions among current models of prefrontal cortical functions. Annals of the New York Academy of Sciences. 1995;769:337–368. doi: 10.1111/j.1749-6632.1995.tb38149.x. [DOI] [PubMed] [Google Scholar]
- Grober E, Buschke H. Genuine memory deficits in dementia. Developmental Neuropsychology. 1987;38:13–36. [Google Scholar]
- Hannula DE, Ryan JD, Tranel D, Cohen NJ. Rapid onset relational memory effects are evident in eye movement behavior, but not in hippocampal amnesia. Journal of Cognitive Neuroscience. 2007;19:1690–1705. doi: 10.1162/jocn.2007.19.10.1690. [DOI] [PubMed] [Google Scholar]
- Hannula DE, Tranel D, Cohen NJ. The long and the short of it: Relational memory impairments in amnesia, even at short lags. The Journal of Neuroscience. 2006;26:8352–8359. doi: 10.1523/JNEUROSCI.5222-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hess TM. Aging and schematic influences on memory. In: Hess TM, editor. Aging and cognition: Knowledge, organization, and utilization. Amsterdam, The Netherlands: North-Holland: 1990. pp. 93–160. [Google Scholar]
- Horn JL, Cattell RB. Age differences in fluid and crystallized intelligence. Acta Psychologica. 1967;26:107–129. doi: 10.1016/0001-6918(67)90011-x. [DOI] [PubMed] [Google Scholar]
- Jeneson A, Squire LR. Working memory, long-term memory, and medial temporal lobe function. Learning & Memory. 2012;19:15–25. doi: 10.1101/lm.024018.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jorm AF, Jacomb PA. The Informant Questionnaire on Cognitive Decline in the Elderly (IQCODE): Sociode-mographic correlates, reliability, validity and some norms. Psychological Medicine. 1989;19:1015–1022. doi: 10.1017/s0033291700005742. [DOI] [PubMed] [Google Scholar]
- Klunk WE, Engler H, Nordberg A, Wang Y, Blomqvist G, Holt DP, Langstrom B. Imaging brain amyloid in Alzheimer's disease with Pittsburgh Compound-B. Annals of Neurology. 2004;55:306–319. doi: 10.1002/ana.20009. [DOI] [PubMed] [Google Scholar]
- Krueger F, Barbey A, Grafman J. The medial pre-frontal cortex mediates social event knowledge. Trends in Cognitive Sciences. 2009;13:103–109. doi: 10.1016/j.tics.2008.12.005. [DOI] [PubMed] [Google Scholar]
- Kurby CA, Sargent JQ, Bailey HR, Zacks JM. Event segmentation and memory in younger and older adults: An fMRI investigation. Poster presented at the 2012 Cognitive Aging Conference; Atlanta, GA. 2012. Apr, [Google Scholar]
- Kurby CA, Zacks JM. Age differences in the perception of hierarchical structure. Memory & Cognition. 2011;39:75–91. doi: 10.3758/s13421-010-0027-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDonald CR, McEvoy LK, Gharapetian L, Fennema-Notestine C, Hagler DJ, Jr, Holland D, Dale AM. Regional rates of neocortical atrophy from normal aging to early Alzheimer disease. Neurology. 2009;73:457–465. doi: 10.1212/WNL.0b013e3181b16431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris JC. The Clinical Dementia Rating (CDR): Current version and scoring rules. Neurology. 1993;43:2412–2414. doi: 10.1212/wnl.43.11.2412-a. [DOI] [PubMed] [Google Scholar]
- Morrow DG, Leirer VO, Altieri PA. Aging, expertise, and narrative processing. Psychology and Aging. 1992;7:376–388. doi: 10.1037//0882-7974.7.3.376. [DOI] [PubMed] [Google Scholar]
- Oztekin I, Davachi L, McElree B. Are representations in working memory distinct from representations in long-term memory? Neural evidence in support of a single store. Psychological Science. 2010;21:1123–1133. doi: 10.1177/0956797610376651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raz N, Dixon FM, Head DP, Dupuis JH, Acker JD. Neuroanatomical correlates of cognitive aging: Evidence from structural MRI. Neuropsychology. 1998;12:95–114. doi: 10.1037//0894-4105.12.1.95. [DOI] [PubMed] [Google Scholar]
- Rosen VM, Caplan L, Sheesley L, Rodriguez R, Grafman J. An examination of daily activities and their scripts across the adult lifespan. Behavior Research Methods, Instruments, & Computers. 2003;35:32–48. doi: 10.3758/bf03195495. [DOI] [PubMed] [Google Scholar]
- Schwartz MF. The quantitative description of action disorganization after brain damage: A case. Cognitive Neuropsychology. 1991;8:381–414. [Google Scholar]
- Schwartz MF, Segal M, Veramonti T, Ferraro M, Buxbaum LJ. The Naturalistic Action Test: A standardized assessment for everyday action impairment. Neuropsychological Rehabilitation. 2002;24:311–339. [Google Scholar]
- Storandt M, Balota DA, Salthouse TA. ELSMEM: A computerized battery to assess Executive, Linguistic, Spatial, and MEMory abilities. 2009 Retrieved from http://www.psych.wustl.edu/coglab/index.html.
- Stoub TR, Rogalski EJ, Leurgans S, Bennett DA, deToledo-Morrell L. Rate of entorhinal and hip-pocampal atrophy in incipient and mild AD: Relation to memory function. Neurobiology of Aging. 2010;31:1089–1098. doi: 10.1016/j.neurobiolaging.2008.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swallow KM, Barch DM, Head D, Maley CJ, Holder D, Zacks JM. Changes in events alter how people remember recent information. Journal of Cognitive Neuroscience. 2011;23:1052–1064. doi: 10.1162/jocn.2010.21524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Unsworth N, Engle RW. On the division of short-term and working memory: An examination of simple and complex span and their relation to higher order abilities. Psychological Bulletin. 2007;133:1038–1066. doi: 10.1037/0033-2909.133.6.1038. [DOI] [PubMed] [Google Scholar]
- Wechsler D, Stone CP. Wechsler Memory Scale. New York, NY: Psychological Corp; 1973. [Google Scholar]
- West RL, Crook TH, Barron KL. Everyday memory performance across the life span: Effects of age and noncognitive individual differences. Psychology and Aging. 1992;7:72–82. doi: 10.1037//0882-7974.7.1.72. [DOI] [PubMed] [Google Scholar]
- Zacks JM, Speer NK, Swallow KM, Braver TS, Reynolds JR. Event perception: A mind/brain perspective. Psychological Bulletin. 2007;133:273–293. doi: 10.1037/0033-2909.133.2.273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zacks JM, Speer NK, Vettel JM, Jacoby LL. Event understanding and memory in healthy aging and dementia of the Alzheimer type. Psychology and Aging. 2006;21:466–482. doi: 10.1037/0882-7974.21.3.466. [DOI] [PubMed] [Google Scholar]
- Zacks RT, Hasher L, Li KZH. Human memory. In: Salthouse TA, Craik FIM, editors. The handbook of aging and cognition. 2nd. Mahwah, NJ: Erlbaum; 2000. pp. 293–357. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.