

Abstract
Interleukin-15 (IL-15) is a cytokine produced in the normal brain that acts on its specific receptor IL-15Rα and co-receptors IL-2Rβ and IL-2Rγ in neuronal cells. The functions of the cerebral IL-15 system, however, are not yet clear. To test the hypothesis that IL-15Rα regulates metabolic activity and body temperature, we quantified the specific metabolic phenotype of IL-15Rα knockout mice. These normal-appearing mice were leaner with lower fat composition. During the entire circadian cycle, the knockout mice had a significantly higher acrophase in locomotor activity and heat dissipation. During the light phase, there was significantly greater food intake, oxygen consumption, and carbon dioxide production. The difference in the dark and light phases suggests that IL-15Rα participates in circadian rhythm regulation. The higher oxygen consumption in the light phase indicates adaptive thermogenesis in the knockout mice. The body temperature of the receptor knockout mice was significantly higher than the control in the light phase, and this was mainly caused by a large difference occurring between 0600 and 0900 h. In addition to the metabolic chamber studies and circadian rhythm analyses, qPCR of hypothalamic homogenates indicated higher mRNA expression of orexin and transient receptor potential vanilloid 4 cation channels. Consistent with a direct role of IL-15Rα in the hypothalamus, IL-15 treatment of the wild-type mice induced c-Fos expression in the preoptic area. We conclude that activation of hypothalamic neurons by IL-15 in mice contributes to thermoregulation and modifies the metabolic phenotype.
Keywords: IL-15, Temperature regulation, Locomotor activity, Hypothalamus, Metabolism, Circadian rhythm, Chronobiology
Introduction
Interleukin-15 (IL-15) is a 14 kD cytokine produced by many types of parenchymal cells in different organs as well as by immune cells. It belongs to the four-helical bundle family of cytokines, binds to its specific receptor IL-15Rα which can recruit the co-receptors IL-2Rβ and IL-2Rγ to form a high-affinity complex and initiates cellular signaling by activation of Janus kinase (JAK)-1 and JAK-2 and Signal Transducer and Activator of Transcription (STAT)-3 and STAT-5 proteins (Waldmann et al. 1998). Most studies of IL-15 have focused on its immunoregulatory functions. Its central nervous system (CNS) actions have been insufficiently addressed. Nonetheless, we recently showed that IL-15 crosses the blood-brain barrier, a process that can be upregulated by administration of lipopolysaccharide (Pan et al. 2008c) but diminished by induction of experimental autoimmune encephalomyelitis (Hsuchou et al. 2009b). IL-15 mRNA and its receptor subunits are also constitutively present in various brain regions (Hanisch et al. 1997; Hsuchou et al. 2009b). These findings indicate active communication between blood-borne IL-15 and the CNS, and suggest a functional role of the IL-15 system in CNS disease processes.
Cytokines exert a broad spectrum of actions in the CNS, including regulation of cognitive function, mood, sleep, feeding, thermoregulation, and many other aspects of neuroendocrine control (Banks et al. 1995). After intracerebroventricular (icv) injection, high doses of IL-15 (100 and 500 ng) increase the number of non-rapid eye movement sleep episodes in rabbits while decreasing their duration. At 500 ng/rabbit, IL-15 also increases brain temperature (Kubota et al. 2001). It is not yet clear how IL-15 affects thermoregulation. Here, we determined whether the cerebral IL-15 system plays a major role in thermoregulation and metabolic activity by behavioral tests and neurochemical analyses of IL-15Rα knockout (KO) mice. The results suggest that the hypothalamus can mediate some of the effects of IL-15 in decreasing body temperature, mediated in part by transient receptor potential vanilloid (TRPV)4 cation channels. The direct involvement of the IL-15 system in the hypothalamus is further supported by studies on wild-type mice after IL-15 treatment.
Materials and Methods
Basic Metabolic Parameters
All studies were conducted following a protocol approved by the Institutional Animal Care and Use Committee. Mice were purchased from Jackson Laboratories (Bar Harbor, ME) and housed in the animal care facility for at least 2 weeks before study. IL-15Rα KO mice (IL-15RKO; B6;129X1-IL-15ratm1Ama/J, stock number 003723) were studied when 2.5–3.5 months old along with their matched controls (B6.129SF2/J, stock number 101045). Separate studies on male and female mice showed that the patterns of changes were not different between genders. Since IL-15 is involved in autoimmune disorders that are more common in females, data from female mice are presented in this report. The mice were housed four per cage and fed with regular rodent chow. The ambient room temperature was 23°C. The light span was 0700–1900 h in a light–dark 12:12 lighting regimen. Body weight and 24-h food intake were recorded.
Metabolic Chamber
The metabolic profile of the mice was measured by Oxymax open circuit calorimeter (Columbus Instruments, Columbus, OH). Since this consists of 16 chambers, seven control mice and eight RKO mice were randomly selected to undergo the measurement. One chamber was left empty as a negative control. The mice were allowed to habituate to single housing 4 days before entering the metabolic chambers. Mice were then individually housed in each metabolic chamber for 7 days. After another habituation period of 4 days within the chamber to avoid any possible confounding anxiety related to a novel environment, data recorded from the subsequent 3 days were used for analyses. Oxygen consumption (VO2), carbon dioxide production (VCO2), respiratory exchange quotient (RER, VO2/VCO2), activity level, and heat dissipation were measured every 18 min for 7 consecutive days.
Temperature Telemetry
Temperature recording was conducted on separate groups of RKO and control mice of 3 m of age (n=3/group). The Mini Mitter Company (Bend, OR) telemetry system was used. Mice were briefly anesthetized by isofluorane inhalation. The gas-sterilized E-mitters were implanted intraperitoneally, and the mice were allowed to recover for 7 days and then single-caged for another 3 days. Recording was initiated after the cages were placed on corresponding ER-4000 receivers and calibrated for each mouse. Data were sampled every minute and captured by VitalView software (Mini Mitter Company). The reading at each hour was used to generate the circadian plot. In separate batches of mice after being used for the metabolic chamber study, rectal temperature was also sampled once between 1100–1200 h (n=9/group).
Nuclear Magnetic Resonance
The body composition of fat, lean mass, and water were measured in a Bruker minispec live mouse analyzer (Bruker Optics, The Woodslands, TX), as described previously (Pan and Kastin 2007). The percent of fat and lean mass were normalized to the body weight of the mouse.
Immunohistochemistry
To determine the effect of IL-15 treatment on hypothalamic thermoregulatory gene expression, B6 mice were anesthetized by intraperitoneal urethane and received IL-15 (2 μg/mouse, R & D Systems, Minneapolis, MN) by intravenous injection through the left jugular vein. The control mice received phosphate buffered saline (PBS) only (50 μl/mouse). Three hours later, the mice were perfused intracardially with sterile PBS and 3% paraformaldehyde and processed for immunohistochemistry as described previously (Pan et al. 2008a; Hsuchou et al. 2009a). A rabbit polyclonal anti-c-fos antibody (Santa Cruz Biotechnology, sc-52, 1 μg/ml) was used as an indicator of cellular activation with increased expression of the immediate early gene.
Real-Time RT-PCR
The samples were collected at 1100–1200 h. The hypothalamus was dissected, and homogenized in RNA lysis buffer. Reverse transcription and real-time RT-PCR (qPCR) were performed by the SYBR Green amplification method as described previously (Pan et al. 2008b). The primers are listed in Table 1. The mRNAs for TRPV4 and TRPV1 cation channels were measured because TRPVs are known to participate in thermoregulation (Caterina 2007; Plant and Strotmann 2007). The mRNA for orexin was determined because this peptide is involved in sleep and appetite regulation (Ganjavi and Shapiro 2007).
Table 1. SYBR Green primer list for qPCR in mouse hypothalamus.
| Gene | Forward primers | Reverse primers |
|---|---|---|
| TRPV1 | FP: TTGTCAATGCCAGCTACACAG | RP:TTTCAATGGCAATGTGTAATGCT |
| TRPV4 | FP: CGAGAACCGCCATGAGATG | RP: CTTACGCCACTTGTCTCTCAACA |
| Orexin | FP: TGGACCACTGCACTGAAGAGA | RP: CAGGGAACCTTTGTAGAAGGAAAG |
| GAPDH | FP: TGTGTCCGTCGTGGATCTGA | RP: CCTGCTTCACCACCTTCTTGA |
Statistics
For single measurements, means are presented with their standard errors. One-way analysis of variance was performed to determine differences between the groups. For rhythmic data, population mean cosinor analyses were used. The percent rhythm, the midline estimating statistic of rhythm (MESOR, a rhythm-adjusted mean), amplitude, and acrophase were determined as described previously (Bingham et al. 1982; Katinas et al. 2001; Pan et al. 2002). Parametric tests were performed to determine the difference between the control and KO groups, taking into consideration not only the amplitude but also the acrophase.
Results
Basic Metabolic Phenotype of the IL15RKO Mice
The RKO mice were normal in appearance, and their body weight was not significantly different from the control group (Fig. 1a). Their fat composition, however, was significantly decreased (Fig. 1b, p<0.05). There was a corresponding increase of lean body mass (p<0.05). In support of a higher metabolism, these RKO mice showed an average of 0.8°C elevation of body temperature that was measured by rectal probe at 11:00 a.m., 4 h after the onset of the light cycle (Fig. 2a, p<0.01). Telemetry monitoring showed that the major difference in body temperature occurred between 0600–0900 h, around the onset of the light span (Fig. 2b).
Figure 1.

Basic metabolic phenotype of IL15Rα KO mice. a There was no difference in body weight. b There was a significant reduction of fat composition and greater lean mass. *p<0.05. n=9/group
Figure 2.

IL15Rα KO mice had higher body temperature. a The difference of resting temperature at 1100–1200 h was 0.8 C. **p<0.01. n=9/group. b The difference in body temperature was most pronounced around the transition from the dark phase to the light phase, occurring at 0600–0900 h
Metabolic Chamber Analyses
The metabolic activities of two groups of RKO and control mice were determined after 4 days habituation in the Oxymax metabolic chamber. Locomotor activity and its circadian variation were determined by measurement of interruption of horizontal and vertical infrared beams. The RKO mice had a significantly higher MESOR activity both in the horizontal plane (p<0.01, Fig. 3a) and in the vertical plane (rearing, p<0.05). The RKO mice were hyperactive during both the light and dark spans (Fig. 3b). Concurrently, there was greater heat dissipation in the RKO mice (p<0.05 for MESOR, Fig. 3c).
Figure 3.

IL15Rα KO mice had higher locomotor activity both during the dark and light phase (a–b), with greater heat dissipation (c). Both variables showed a significant difference in the MESOR by cosinor analysis (p<0.05). Total horizontal plane activity was significantly higher in both the dark and light phases (***p<0.005)
The RKO group also showed a trend of increased oxygen consumption when all 3 days were analyzed (p=0.08, Fig. 4a). The increase of oxygen consumption in the light phases was significant (p<0.05, Fig. 4b). The production of carbon dioxide tended to increase concomitantly although not quite reaching statistical significance during the entire cycle (p=0.06, Fig. 4c), but was significantly different from the control group in the light phase (p<0.05, Fig. 4d). As a result of the concurrent increase of VO2 and VCO2, the respiratory quotient RER was unchanged (p=0.19). The RER ranged from 0.8 to 1. This indicates a greater propensity for carbohydrate metabolism than fat oxidation. There was a significant increase in food intake in the light phase (Fig. 5a–b). This contrasts with the lack of significant increase of food intake in the dark phase.
Figure 4.

IL15Rα KO mice had a trend toward a higher MESOR in both oxygen consumption (a VO2, p=0.08) and carbon dioxide production (c VCO2, p=0.06) (n=7/group). The difference in the light phase was significant for both VO2 (b) and VCO2 (d). *p<0.05
Figure 5.

IL15Rα KO mice showed greater food intake in the light phase (a–b). n=7/group. ***p<0.005
mRNA of TRPV4, TRPV1, and Orexin in the Hypothalamus
In the hypothalamus of the RKO mice, there was a significant increase of TRPV4 mRNA (Fig. 6a). This is in contrast to the lack of significant changes of TRPV1 mRNA (not shown). The mRNA for orexin (both A and B isoforms with primers synthesized for the shared sequences) also showed a trend toward an increase (p=0.08, Fig. 6b).
Figure 6.

IL15Rα KO mice showed a significantly higher mRNA for TRPV4 (a p<0.05) and a trend toward increased orexin mRNA (b p= 0.08) in the hypothalamus. n=5/group
c-Fos Immunoreactivity in B6 Mice after IL-15 Treatment
In the hypothalamus of a control mouse that received intravenous injection of PBS 3 h earlier, the basal c-fos immunoreactivity was scattered dorsal to the preoptic area. In a mouse 3 h after IL-15 intravenous injection, there was an increase in the number of c-fos immunoreactive cells, mainly in the medial preoptic area (arrows, Fig. 7).
Figure 7.
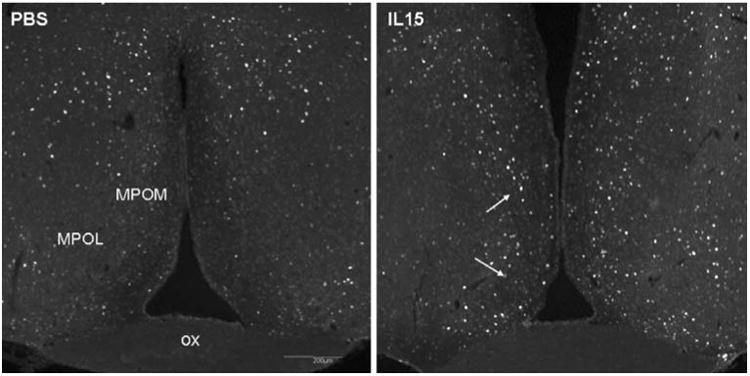
The IL15-treated mice had a higher number of c-fos immunoreactive cells in the medial portion of the preoptic area 3 h after iv injection of IL15
Discussion
For unknown reasons, the functions of cerebral IL-15 have eluded attention from most neuroscientists. Lee and colleagues first reported the presence of IL-15 mRNA and protein in enriched cultures of human fetal astrocytes and microglia, and showed their upregulation by inflammatory stimuli (Lee et al. 1996). IL-15 and its receptors are detectable in both neonatal and adult mouse brains. IL-15 acts as an autocrine factor for microglia and affects nitric oxide production and cell growth in culture (Hanisch et al. 1997). The mRNA for IL-15 and IL-15Rα isoforms has higher expression in the hippocampus and cerebellum than in the cerebral cortex and thalamus of the fetal brain (Kurowska et al. 2002), but the levels of expression have not been studied in the hypothalamus. Nonetheless, the increase of brain temperature in response to icv IL-15 (Kubota et al. 2001) suggests that IL-15 may also act on the hypothalamus to regulate body temperature and metabolic activity.
The IL-15Rα deletion in the KO mice was generated by replacing a 3.0 kb fragment of the IL-15Rα genomic clone containing exons 2 and 3 with a phosphoglyceraldehyde-driven neomycin resistance gene casette flanked by lox sequences (ploxNeolox), ligation to a phosphoglyceraldehyde-driven thymidine kinase gene casette (pgkTK), and selection by G418 and 2′ fluoro-5-iodo-arabinofuranosyluracil (Lodolce et al. 1998). The mutation abolishes the production of any known functional alternative splice variants of IL-15Rα. The IL-15Rα KO mice show decreased proliferation and homing of lymphocytes to peripheral lymph nodes, resulting in lymphopenia despite grossly normal T and B lymphocyte development (Lodolce et al. 1998). Even though the abnormality in the immune system becomes symptomatic after 3 months of age, there was no obvious structural abnormality in the hypothalamic region of these mice found in this study.
The RKO mice showed normal growth and development, with the same body weight as the strain background controls. However, nuclear magnetic resonance (NMR) analysis showed a significant reduction of fat composition and higher lean body mass. Fat utilization was similar to the controls, reflected by RER. There was also higher food intake. Thus, we hypothesized that these mice must have a faster metabolism to maintain their homeostasis.
As expected from this hypothesis, the RKO mice had a higher body temperature. The most pronounced change occurred at the transition from the dark phase to light phase, coinciding with higher locomotor activity in both the horizontal plane and rearing. Thus, elevation of body temperature may be a result of increased locomotor activity; but how much is contributed by hypothalamic thermoregulation? In the periphery, IL-15 increases uncoupling protein expression and lipid oxidation, and eventually reduces the mass of both brown and white adipose tissue (Almendro et al. 2008). However, the RKO mice show an increase of oxygen consumption (VO2) in the light phase, consistent with adaptive thermogenesis (DeRuisseau et al. 2004). Since IL-15Rα deletion results in an increase in body temperature, this suggests a hypothermic effect of IL-15 signaling. Therefore, the higher body temperature in the RKO mice suggests it is a central mechanism, rather than antagonism of a peripheral mechanism, that plays a major role in thermoregulation of these mice.
The control of body temperature is an integration of input from the core and body surface thermal afferents by the hypothalamic thermostat which has a regulatory set point. The output by the sympathetic nervous system affects vasomotor tone, sweat secretion, and activities of the adrenal and thyroid glands. Output via the somatomotor nervous system alters skeletal muscle shivering and mouse behavior. The increase of locomotor activity indicates that IL-15Rα knockout mice have motor activation that probably results in an elevation of body temperature. In addition, the involvement of the hypothalamus in IL-15Rα-mediated thermoregulation is supported by elevation of the vallinoid receptor TRPV4 and of the thermogenic peptide orexin in hypothalamic extracts. It is known that TRPV1 induces hypothermia (Gavva et al. 2007; Gavva 2008). In contrast to a role of TRPV1 in the periphery that indirectly acts on thermoregulation (McGaraughty et al. 2009), TRPV4 is located in anterior hypothalamic structures involved in temperature sensation and the integration of thermal and osmotic information (Guler et al. 2002). Besides the sleep-inducing role of orexin-A, this neuropeptide also promotes sympathetic and thermogenic activation, thus increasing energy expenditure (Monda et al. 2003). The slight increase of orexin mRNA in the hypothalamus (its primary site of production) also tends to support involvement of the hypothalamus in the regulation of body temperature in IL-15Rα KO mice. Furthermore, IL-15 treatment in wild-type mice directly activated c-Fos in the preoptic area, a region of the hypothalamus known to play a crucial role in thermoregulation (McGaraughty et al. 2009). Therefore, the results indicate that hypothalamic IL-15Rα is crucially involved in thermoregulation and maintenance of energy expenditure.
In summary, IL-15Rα knockout mice have decreased body fat and increased lean mass, without altered body weight. This can be explained by their increased body temperature, locomotor activity, food intake, and heat dissipation associated with increased adaptive thermogenesis. We conclude that disruption of IL-15Rα signaling is involved in excessive motor activation, disrupted circadian rhythm of thermoregulation, and altered metabolic phenotype.
Acknowledgments
Supported by NIH (NS62291 and NS45751 to WP and DK54880 to AJK).
Contributor Information
Yi He, Pennington Biomedical Research Center, LSU System, 6400 Perkins Road, Baton Rouge, LA 70808, USA.
Xiaojun Wu, Pennington Biomedical Research Center, LSU System, 6400 Perkins Road, Baton Rouge, LA 70808, USA.
Reas S. Khan, Pennington Biomedical Research Center, LSU System, 6400 Perkins Road, Baton Rouge, LA 70808, USA
Abba J. Kastin, Pennington Biomedical Research Center, LSU System, 6400 Perkins Road, Baton Rouge, LA 70808, USA
Germaine G. Cornelissen-Guillaume, Chronobiology Laboratories, University of Minnesota, Minneapolis, MN 55455, USA
Hung Hsuchou, Pennington Biomedical Research Center, LSU System, 6400 Perkins Road, Baton Rouge, LA 70808, USA.
Barry Robert, Pennington Biomedical Research Center, LSU System, 6400 Perkins Road, Baton Rouge, LA 70808, USA.
Franz Halberg, Chronobiology Laboratories, University of Minnesota, Minneapolis, MN 55455, USA.
Weihong Pan, Email: panw@pbrc.edu, Pennington Biomedical Research Center, LSU System, 6400 Perkins Road, Baton Rouge, LA 70808, USA.
References
- Almendro V, Fuster G, Busquets S, Ametller E, Figueras M, Argiles JM, Lopez-Soriano FJ. Effects of IL-15 on rat brown adipose tissue: uncoupling proteins and PPARs. Obesity (Silver Spring) 2008;16:285–289. doi: 10.1038/oby.2007.47. [DOI] [PubMed] [Google Scholar]
- Banks WA, Kastin AJ, Broadwell RD. Passage of cytokines across the blood-brain barrier. Neuroimmunomodulation. 1995;2:241–248. doi: 10.1159/000097202. [DOI] [PubMed] [Google Scholar]
- Bingham C, Arbogast B, Guillaume GC, Lee JK, Halberg F. Inferential statistical methods for estimating and comparing cosinor parameters. Chronobiologia. 1982;9:397–439. [PubMed] [Google Scholar]
- Caterina MJ. Transient receptor potential ion channels as participants in thermosensation and thermoregulation. Am J Physiol Regul Integr Comp Physiol. 2007;292:R64–R76. doi: 10.1152/ajpregu.00446.2006. [DOI] [PubMed] [Google Scholar]
- DeRuisseau LR, Parsons AD, Overton JM. Adaptive thermogenesis is intact in B6 and A/J mice studied at thermoneutrality. Metabolism. 2004;53:1417–1423. doi: 10.1016/j.metabol.2004.06.007. [DOI] [PubMed] [Google Scholar]
- Ganjavi H, Shapiro CM. Hypocretin/Orexin: a molecular link between sleep, energy regulation, and pleasure. J Neuropsych Clin Neurosci. 2007;19:413–419. doi: 10.1176/jnp.2007.19.4.413. [DOI] [PubMed] [Google Scholar]
- Gavva NR. Body-temperature maintenance as the predominant function of the vanilloid receptor TRPV1. Trends Pharmacol Sci. 2008;29:550–557. doi: 10.1016/j.tips.2008.08.003. [DOI] [PubMed] [Google Scholar]
- Gavva NR, Bannon AW, Surapaneni S, Hovland DN, Jr, Lehto SG, Gore A, Juan T, Deng H, Han B, Klionsky L, Kuang R, Le A, Tamir R, Wang J, Youngblood B, Zhu D, Norman MH, Magal E, Treanor JJ, Louis JC. The vanilloid receptor TRPV1 is tonically activated in vivo and involved in body temperature regulation. J Neurosci. 2007;27:3366–3374. doi: 10.1523/JNEUROSCI.4833-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guler AD, Lee H, Iida T, Shimizu I, Tominaga M, Caterina M. Heat-evoked activation of the ion channel, TRPV4. J Neurosci. 2002;22:6408–6414. doi: 10.1523/JNEUROSCI.22-15-06408.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanisch UK, Lyons SA, Prinz M, Nolte C, Weber JR, Kettenmann H, Kirchhoff F. Mouse brain microglia express interleukin-15 and its multimeric receptor complex functionally coupled to Janus kinase activity. J Biol Chem. 1997;272:28853–28860. doi: 10.1074/jbc.272.46.28853. [DOI] [PubMed] [Google Scholar]
- Hsuchou H, He Y, Kastin AJ, Tu H, Markadakis EN, Rogers RC, Fossier PB, Pan W. Obesity induces functional astrocytic leptin receptors in hypothalamus. Brain. 2009a;132:889–902. doi: 10.1093/brain/awp029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsuchou H, Pan W, Wu X, Kastin AJ. Cessation of blood-to-brain influx of interleukin-15 during development of EAE. J Cereb Blood Flow Metab. 2009b;29:1568–1578. doi: 10.1038/jcbfm.2009.79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katinas GS, Halberg F, Cornelissen G, Hawkins D, Bueva MV, Korzhevsky DE, Sapozhnikova LR, Rhodus N, Schaffer E. About 8- and approximately 84-h rhythms in endotheliocytes as in endothelin-1 and effect of trauma. Peptides. 2001;22:647–659. doi: 10.1016/s0196-9781(01)00375-8. [DOI] [PubMed] [Google Scholar]
- Kubota T, Brown RA, Fang J, Krueger JM. Interleukin-15 and interleukin-2 enhance non-REM sleep in rabbits. Am J Physiol Regul Integr Comp Physiol. 2001;281:R1004–R1012. doi: 10.1152/ajpregu.2001.281.3.R1004. [DOI] [PubMed] [Google Scholar]
- Kurowska M, Rudnicka W, Maslinska D, Maslinski W. Expression of IL-15 and IL-15 receptor isoforms in select structures of human fetal brain. Ann N Y Acad Sci. 2002;966:441–445. doi: 10.1111/j.1749-6632.2002.tb04245.x. [DOI] [PubMed] [Google Scholar]
- Lee YB, Satoh J, Walker DG, Kim SU. Interleukin-15 gene expression in human astrocytes and microglia in culture. Neuro-Report. 1996;7:1062–1066. doi: 10.1097/00001756-199604100-00022. [DOI] [PubMed] [Google Scholar]
- Lodolce JP, Boone DL, Chai S, Swain RE, Dassopoulos T, Trettin S, Ma A. IL-15 receptor maintains lymphoid homeostasis by supporting lymphocyte homing and proliferation. Immunity. 1998;9:669–676. doi: 10.1016/s1074-7613(00)80664-0. [DOI] [PubMed] [Google Scholar]
- McGaraughty S, Segreti JA, Fryer RM, Brown BS, Faltynek CR, Kym PR. Antagonism of TRPV1 receptors indirectly modulates activity of thermoregulatory neurons in the medial preoptic area of rats. Brain Res. 2009;1268:58–67. doi: 10.1016/j.brainres.2009.02.018. [DOI] [PubMed] [Google Scholar]
- Monda M, Viggiano A, De Luca V. Haloperidol reduces the sympathetic and thermogenic activation induced by orexin A. Neurosci Res. 2003;45:17–23. doi: 10.1016/s0168-0102(02)00191-8. [DOI] [PubMed] [Google Scholar]
- Pan W, Kastin AJ. Mahogany, blood-brain barrier, and fat mass surge in AVY mice. Int J Obes. 2007;31:1030–1032. doi: 10.1038/sj.ijo.0803536. [DOI] [PubMed] [Google Scholar]
- Pan W, Cornelissen G, Halberg F, Kastin AJ. Selected contribution: circadian rhythm of tumor necrosis factor-alpha uptake into mouse spinal cord. J Appl Physiol. 2002;92:1357–1362. doi: 10.1152/japplphysiol.00915.2001. [DOI] [PubMed] [Google Scholar]
- Pan W, Hsuchou H, He Y, Sakharkar A, Cain C, Yu C, Kastin AJ. Astrocyte leptin receptor (ObR) and leptin transport in adult-onset obese mice. Endocrinology. 2008a;149:2798–2806. doi: 10.1210/en.2007-1673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pan W, Hsuchou H, Tu H, Kastin AJ. Developmental changes of leptin receptors in cerebral microvessels: unexpected relation to leptin transport. Endocrinology. 2008b;149:877–885. doi: 10.1210/en.2007-0893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pan W, Hsuchou H, Yu C, Kastin AJ. Permeation of blood-borne IL15 across the blood-brain barrier and the effect of LPS. J Neurochem. 2008c;106:313–319. doi: 10.1111/j.1471-4159.2008.05390.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plant TD, Strotmann R. TRPV4. Handb Exp Pharmacol. 2007;179:189–205. doi: 10.1007/978-3-540-34891-7_11. [DOI] [PubMed] [Google Scholar]
- Waldmann T, Tagaya Y, Bamford R. Interleukin-2, interleukin-15, and their receptors. Int Rev Immunol. 1998;16:205–226. doi: 10.3109/08830189809042995. [DOI] [PubMed] [Google Scholar]