

Significance
Has natural selection led to adaptations of Lévy flight foraging, as stated on the respective Wikipedia page? Random walks with scale-free jump length distributions were indeed shown to optimize the search for sparse targets as supported by extensive movement data of many animal species and humans. Here we demonstrate that small variations of the search conditions strongly modify these claims: In the presence of a bias, underwater currents for sea predators or winds for airborne searchers, a Lévy searcher easily overshoots the target, and Brownian strategies become advantageous. Even in the absence of a bias, there exist conditions for which a Brownian strategy may effect faster target localization. Our results show clear limitations for the universality of Lévy flight foraging.
Keywords: search optimization, stochastic processes, Lévy foraging hypothesis
Abstract
It is generally believed that random search processes based on scale-free, Lévy stable jump length distributions (Lévy flights) optimize the search for sparse targets. Here we show that this popular search advantage is less universal than commonly assumed. We study the efficiency of a minimalist search model based on Lévy flights in the absence and presence of an external drift (underwater current, atmospheric wind, a preference of the walker owing to prior experience, or a general bias in an abstract search space) based on two different optimization criteria with respect to minimal search time and search reliability (cumulative arrival probability). Although Lévy flights turn out to be efficient search processes when the target is far from the starting point, or when relative to the starting point the target is upstream, we show that for close targets and for downstream target positioning regular Brownian motion turns out to be the advantageous search strategy. Contrary to claims that Lévy flights with a critical exponent α = 1 are optimal for the search of sparse targets in different settings, based on our optimization parameters the optimal α may range in the entire interval (1, 2) and especially include Brownian motion as the overall most efficient search strategy.
How do you find the proverbial needle in the haystack or an enemy submarine in the vast expanse of the sea? Scientists have studied the dynamics and optimization of search processes for decades, their interests ranging from military tasks such as locating enemy vessels or mines in the ocean and search strategies of animals for food to diffusion control of molecular processes in biological cells (1–4). Without prior knowledge about the location of the target, a searcher randomly explores the search space. However, as already argued by Shlesinger and Klafter (5), instead of performing a Brownian walk a better search strategy for sparse targets is that of a Lévy flight (LF): The agent moves randomly with a power-law distribution 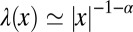 of relocation lengths. Owing to their scale-free, fractal character, LFs combine local exploration with decorrelating, long-range excursions. These effect a reduced oversampling compared with Brownian search (i.e., the forager is less likely to return to previously visited sites). In the field of movement ecology (6), such random jump-like search processes are often referred to as “blind search” using saltatory motion, which is typical for predators hunting at spatial scales that exceed their sensory range (7–10). Such blind search, inter alia, was observed for fully aquatic marine vertebrate predators including plankton-feeding basking sharks (Cetorhinus maximus) (11), jellyfish predators, leatherback turtles (Dermochelys coriacea) (12), and southern elephant seals (13). This is the kind of search motion we investigate here.
of relocation lengths. Owing to their scale-free, fractal character, LFs combine local exploration with decorrelating, long-range excursions. These effect a reduced oversampling compared with Brownian search (i.e., the forager is less likely to return to previously visited sites). In the field of movement ecology (6), such random jump-like search processes are often referred to as “blind search” using saltatory motion, which is typical for predators hunting at spatial scales that exceed their sensory range (7–10). Such blind search, inter alia, was observed for fully aquatic marine vertebrate predators including plankton-feeding basking sharks (Cetorhinus maximus) (11), jellyfish predators, leatherback turtles (Dermochelys coriacea) (12), and southern elephant seals (13). This is the kind of search motion we investigate here.
How commonly are LFs actually observed in nature? Apart from the flight of the albatross (14, 15), power-law relocation statistics were reported for a variety of species including mussels (16), spider monkeys (17), jackals (18), marine predators (8, 9), and even for human motion patterns (19, 20). Also for microscopic creatures LF search may be found; for instance, for the movement of Escherichia coli owing to the power-law statistics underlying their flagella rotation switching (21, 22). In engineering the optimal search behavior of robots was identified to follow LF behavior (23). However, in several cases the reports of LFs are debated. Thus, spider monkeys actually move deterministically (24), and mussels are believed to have multimodal and not power-law relocation patterns (25, 26). Similarly, plant lice have Lévy movements at the population level but not for the motion of individuals (27). However, the discussion of the LF nature of the flight of the albatross recently saw an interesting twist. Whereas a reanalysis of albatross flights showed that they generally are not LFs (28), strong evidence was presented according to which LFs are indeed a search pattern for individuals (29).
Despite this ongoing controversy the LF search hypothesis is a widely accepted dogma in the statistical and biological physics communities and corroborated by a number of careful studies using extensive movement data. Viswanathan et al. (4) indeed promote the LF foraging hypothesis: “Superdiffusive motion governed by fat-tailed propagators optimize encounter rates under specific (but common) circumstances: hence some species must have evolved mechanisms that exploit these properties […].” How justified is this dogma? We here investigate the generic problem of the efficiency of LF search based on complementary optimization criteria such as the search time and the search reliability (cumulative arrival probability) for a single target in a large (infinite) one-dimensional search space. We consider different initial separations between target and searcher and explicitly allow for an external bias in the search process. The latter occurs naturally due to underwater currents, atmospheric winds, or a tilt in a complex algorithmic search space. Such a bias could also mimic the directional preference of a searcher based on previous experience. In our approach we assume that the target is fixed in space, for instance, we could envisage a bird of prey searching for a mouse or snake in a field on a windy day, or search robots trying to locate a shipwreck on the sea floor in the presence of underwater streams.
Based on the statistics of first arrival of one-dimensional LFs to the target we show here that the optimal search strategy crucially depends on the initial searcher–target distance and the presence of even a small bias, in particular, on the direction of the bias with respect to the target location relative to the initial position of the searcher. We find that Brownian search may be more efficient than LF search when the stream is toward the target or, alternatively, when the target happens to be close to the searcher. Conversely, LF search wins out when the target is difficult to locate. Our results shed new light on the long-standing question of optimization in random search processes and challenge the generic role of the LF foraging hypothesis. Without any prior knowledge it might actually be advantageous to use Brownian search methods.
In what follows we first define the LF model and the associated first arrival process. We then define our measures to gauge the optimization of the search process in terms of the search efficiency and the search reliability (cumulative success probability). In Results we present our results for the cases without and with an external drift. In Discussion we analyze these results in detail. We also comment on the relation of the simplified LF approach to other search models such as intermittent search or Lévy walk (LW) models. Finally, we also argue that the one-dimensional analysis presented here is relevant to the case of (effectively) 2D search.
Model
LFs and First Arrival Time.
Consider the scenario sketched in Fig. 1: A walker searches one-dimensionally for a stationary target by performing random jumps, with a prescribed distribution of jump lengths x. In the blind search scenario with saltatory motion studied herein, location of the target occurs when at the end of a jump the walker hits the target. For LFs the distribution of jump lengths is of the power-law form 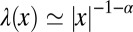 with
with 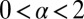 (30, 31). The associated variance
(30, 31). The associated variance 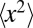 of the jump lengths diverges, such that the resulting scale-free jump process features occasional, extremely long jumps. In contrast to Brownian search with frequent returns to previously visited points in space, as originally pointed out by Shlesinger and Klafter (5) these long jumps improve the efficiency of the search process by leading the random walker to uncorrelated areas of the search space. We recall that for LFs long leap-overs with length distribution
of the jump lengths diverges, such that the resulting scale-free jump process features occasional, extremely long jumps. In contrast to Brownian search with frequent returns to previously visited points in space, as originally pointed out by Shlesinger and Klafter (5) these long jumps improve the efficiency of the search process by leading the random walker to uncorrelated areas of the search space. We recall that for LFs long leap-overs with length distribution 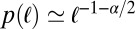 across a point may frequently occur, and thus the probability of actually arriving at a point is significantly smaller than the passage of the walker across this point (32–34). Leap-overs are crucial to the understanding of the first arrival of LFs. They occur radially in all dimensions.
across a point may frequently occur, and thus the probability of actually arriving at a point is significantly smaller than the passage of the walker across this point (32–34). Leap-overs are crucial to the understanding of the first arrival of LFs. They occur radially in all dimensions.
Fig. 1.
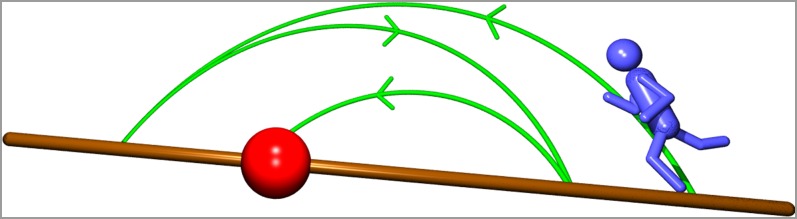
Scheme of the blind random search process. A walker performs random jumps in the search space until he hits the target. Here, the search is biased by a drift away from the target. Such an uphill drift caused, for instance, by underwater streams or above-ground winds affects the search efficiency. Note that for LFs the walker may overshoot the target (the so-called leap-overs) such that the first arrival to the target is less efficient than the passage across the target.
As discussed here, the jumps may be biased by a drift representing an external bias (wind or water current) or by the searcher’s previous experience. We refer to the bias as “downhill” when the target lies in the direction of the stream as seen from the initial position of the walker, and vice versa. Discovery of the target then corresponds to the process of first arrival of the walker at the target position. The basis for our description of the first arrival process for LFs is the fractional Fokker–Planck equation (FFPE) for LFs in the presence of the bias (drift velocity) v (30, 31):
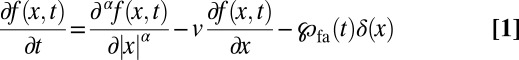 |
defined for 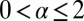 . The distribution f(x, t) is the density function to find the walker at position x at time t, for which we assume the initial position x0, i.e.,
. The distribution f(x, t) is the density function to find the walker at position x at time t, for which we assume the initial position x0, i.e., 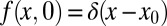 . In our rescaled units, we note that we encounter a unit (generalized) diffusivity in Eq. 1. The fractional derivative
. In our rescaled units, we note that we encounter a unit (generalized) diffusivity in Eq. 1. The fractional derivative 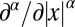 is simply defined in terms of its Fourier transform,
is simply defined in terms of its Fourier transform, 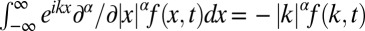 , where
, where 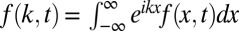 is the Fourier transform of f(x, t). Intrinsically, this is a nonlocal operator reflecting the spatial correlations introduced by the scale-free nature of the jump length distribution
is the Fourier transform of f(x, t). Intrinsically, this is a nonlocal operator reflecting the spatial correlations introduced by the scale-free nature of the jump length distribution 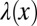 . In the limit α = 2 we recover the standard second-order derivative
. In the limit α = 2 we recover the standard second-order derivative 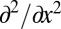 of the Brownian diffusion term of the Fokker–Planck equation. In Eq. 1, we introduced rescaled, dimensionless variables, such that v is a measure for the amplitude of the drift (see SI Text for the rescaling of the FFPE). Eq. 1 contains a point sink at
of the Brownian diffusion term of the Fokker–Planck equation. In Eq. 1, we introduced rescaled, dimensionless variables, such that v is a measure for the amplitude of the drift (see SI Text for the rescaling of the FFPE). Eq. 1 contains a point sink at 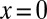 representing the target: the random walker is removed when the target is hit, and
representing the target: the random walker is removed when the target is hit, and 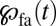 is the density of first arrival. Eq. 1 generalizes the first arrival dynamics in absence of a drift of ref. 32. Owing to the sink term,
is the density of first arrival. Eq. 1 generalizes the first arrival dynamics in absence of a drift of ref. 32. Owing to the sink term, 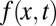 is not normalized, that is, the cumulative survival
is not normalized, that is, the cumulative survival 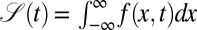 is a decreasing function of time. Using the properties of the fractional derivative, integration of Eq. 1 over the position coordinate x delivers the first arrival density,
is a decreasing function of time. Using the properties of the fractional derivative, integration of Eq. 1 over the position coordinate x delivers the first arrival density, 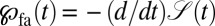 .
.
The solution of Eq. 1 can be obtained via Fourier–Laplace transform, and for the first arrival density 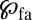 we find
we find
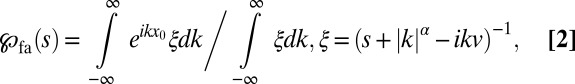 |
where the Laplace transform with respect to time of the function 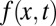 is defined through
is defined through 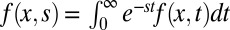 . The result of Eq. 2 instantly shows an important feature: For discontinuous LFs with
. The result of Eq. 2 instantly shows an important feature: For discontinuous LFs with 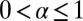 , the quantity
, the quantity 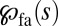 vanishes, because the integral in the denominator diverges while the integral in the numerator converges. Thus, Lévy search for a point-like target will never succeed for
vanishes, because the integral in the denominator diverges while the integral in the numerator converges. Thus, Lévy search for a point-like target will never succeed for 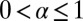 . This property reflects the transience of LFs with
. This property reflects the transience of LFs with 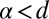 , where d is the embedding spatial dimension (35); we refer also to the discussion of higher dimensions in Discussion. In this sense the value
, where d is the embedding spatial dimension (35); we refer also to the discussion of higher dimensions in Discussion. In this sense the value 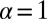 obtained for optimal search for sparse targets in drift-free search (14, 36–41) is to be seen as a limiting point for α from above unity. We obtained analytical results for the first arrival behavior encoded in Eq. 2. in the limit of a small bias, discussed in SI Text . In the following we combine numerical analysis and complementary definitions of the search efficiency to study the optimal random search of Brownian versus LF strategies.
obtained for optimal search for sparse targets in drift-free search (14, 36–41) is to be seen as a limiting point for α from above unity. We obtained analytical results for the first arrival behavior encoded in Eq. 2. in the limit of a small bias, discussed in SI Text . In the following we combine numerical analysis and complementary definitions of the search efficiency to study the optimal random search of Brownian versus LF strategies.
Search Efficiency and Reliability.
What is a good measure for the efficiency of a search mechanism? There are two frequently used definitions of search efficiency: counting the number of successfully located targets either per traveled unit distance or per number of steps (7). These definitions work well when there is a finite target density, as is often assumed in the literature. Here we are interested in the more natural problem of the search for a single target, a countable number of targets, or a finite target area. In such cases the average search time 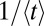 diverges, and we thus need a modified definition for the search efficiency. We advocate the average over inverse search times,
diverges, and we thus need a modified definition for the search efficiency. We advocate the average over inverse search times,
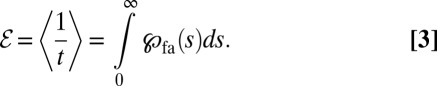 |
Owing to this definition of ℰ as the mean of the inverse first arrival times, contributions from short and intermediate times dominate the efficiency. To demonstrate the usefulness of the definition in Eq. 3 we determined ℰ for a Brownian searcher for both downhill and uphill situations with arbitrary v and 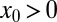 . We find, respectively,
. We find, respectively,
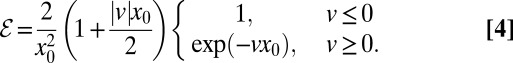 |
Consistently we observe that the search efficiency increases with v when the stream pushes the searcher toward the target, whereas the efficiency decreases exponentially in the uphill case. The latter can be interpreted as an activation barrier for target detection. In the absence of a drift the efficiency is just the inverse mean diffusion time (on average, the diffusive scaling 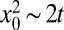 holds in dimensionless units).
holds in dimensionless units).
Combining expressions Eq. S4 from SI Text and Eq. 3 we obtain the search efficiency for an LF in the presence of a weak bias,
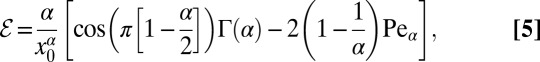 |
for 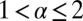 . Here we introduced the generalized Péclet number
. Here we introduced the generalized Péclet number 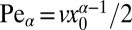 that quantifies the relative strength of the external drift v versus the (unit) diffusivity. The length scale is set by the distance x0 between the walker’s initial position and the target. Note that
that quantifies the relative strength of the external drift v versus the (unit) diffusivity. The length scale is set by the distance x0 between the walker’s initial position and the target. Note that 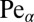 is in fact dimensionless, owing to the rescaling of variables (see Eq. 1). This is our first main result. In the Brownian limit
is in fact dimensionless, owing to the rescaling of variables (see Eq. 1). This is our first main result. In the Brownian limit 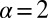 the efficiency is
the efficiency is 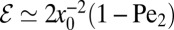 , which corresponds to the small v expansion in Eq. 4. For
, which corresponds to the small v expansion in Eq. 4. For 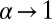 and with
and with 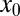 fixed the efficiency drops to zero. Although
fixed the efficiency drops to zero. Although 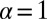 is the optimal parameter for LF search of sparse but finite target density, for the case considered here the transition to discontinuous LFs at
is the optimal parameter for LF search of sparse but finite target density, for the case considered here the transition to discontinuous LFs at 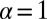 means that the target can no longer be detected. Already these simple observations show that the standard dogmas on the efficiency of random search processes are more specific than usually assumed.
means that the target can no longer be detected. Already these simple observations show that the standard dogmas on the efficiency of random search processes are more specific than usually assumed.
A complementary measure for the quality of a given search process is the cumulative probability
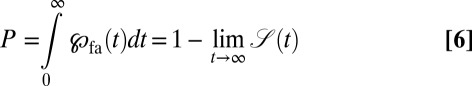 |
that the walker ever arrives at the target. For some purposes this search reliability may be more relevant than the efficiency ℰ. A large value of P for given parameters corresponds to a high success probability to eventually locate the target. Even for Brownian search, when the drift points away from the target with respect to the walker’s initial position, the search reliability may be smaller than unity (i.e., there is a finite residual probability that the walker never locates the target).
Results
Drift-Free Case.
Let us analyze the efficiency of LF search in more detail, starting with the case of vanishing drift strength v. This situation is usually analyzed in the search literature. Because the time to reach the target grows substantially with initial distance 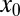 , we compare the search efficiency for different values of
, we compare the search efficiency for different values of 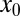 . In Fig. 2 we show the dependence of the relative efficiency
. In Fig. 2 we show the dependence of the relative efficiency 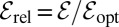 on the stable index α defining how steep the power-law tail of the jump length distribution
on the stable index α defining how steep the power-law tail of the jump length distribution 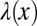 is. Here
is. Here 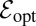 is the maximal value of ℰ for given
is the maximal value of ℰ for given 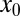 attained at the optimal stable index
attained at the optimal stable index 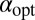 . We observe that when the starting point of the walker
. We observe that when the starting point of the walker 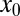 is close to the target, the optimal search strategy is Brownian. This is intuitively clear: Brownian motion with its local jumps cannot overshoot the target and therefore leads to quick target localization. For more distant targets the oversampling of Brownian walks, that is, the tendency to multiply return to previously visited points, reduces the Brownian efficiency and LFs win out. This is shown for the larger
is close to the target, the optimal search strategy is Brownian. This is intuitively clear: Brownian motion with its local jumps cannot overshoot the target and therefore leads to quick target localization. For more distant targets the oversampling of Brownian walks, that is, the tendency to multiply return to previously visited points, reduces the Brownian efficiency and LFs win out. This is shown for the larger 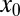 values in Fig. 2. Interestingly, the behavior of the relative efficiency
values in Fig. 2. Interestingly, the behavior of the relative efficiency 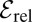 can acquire a nonmonotonic dependence on α and becomes sharper for increasing
can acquire a nonmonotonic dependence on α and becomes sharper for increasing 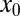 . In the limit of very large
. In the limit of very large 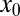 the optimal value of the stable index α tends to unity, consistent with previous observations (14, 36–41). The nonmonotonicity of
the optimal value of the stable index α tends to unity, consistent with previous observations (14, 36–41). The nonmonotonicity of 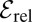 is another central result in the present work that has crucial consequences for the optimization of a given blind random search process.
is another central result in the present work that has crucial consequences for the optimization of a given blind random search process.
Fig. 2.

Relative efficiency 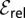 for LFs with vanishing drift
for LFs with vanishing drift 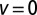 , as a function of the stable index α, Eq. 5. The curves are drawn for the initial positions
, as a function of the stable index α, Eq. 5. The curves are drawn for the initial positions 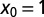 (dashed green),
(dashed green), 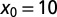 (dotted red),
(dotted red), 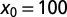 (dashed blue), and
(dashed blue), and 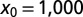 (solid black). With growing
(solid black). With growing 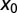 the functional shape changes from a monotonic to nonmonotonic shape and then becomes sharper, the maxima shifting toward unity. Thus, for
the functional shape changes from a monotonic to nonmonotonic shape and then becomes sharper, the maxima shifting toward unity. Thus, for 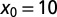 we find
we find 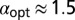 , whereas for
, whereas for 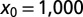 ,
, 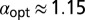 .
.
At fixed starting position 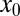 in the drift-free case the implicit expression to determine the optimal stable index
in the drift-free case the implicit expression to determine the optimal stable index 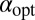 follows from the extremal condition
follows from the extremal condition 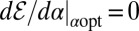 . The result reads
. The result reads
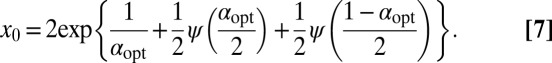 |
Here ψ denotes the digamma function. Eq. 7 allows us to plot 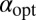 as a function of the initial position of the LF searcher shown in Fig. 3. Interestingly, if for our dimensionless units the initial position is closer to the target than
as a function of the initial position of the LF searcher shown in Fig. 3. Interestingly, if for our dimensionless units the initial position is closer to the target than 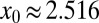 , then the optimal search strategy is Brownian; otherwise, it corresponds to LFs with
, then the optimal search strategy is Brownian; otherwise, it corresponds to LFs with 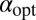 < 2 (in math mode).
< 2 (in math mode).
Fig. 3.

Optimal stable index 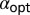 as function of the initial position
as function of the initial position 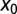 of the walker, as obtained from Eq. 7. For
of the walker, as obtained from Eq. 7. For 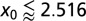 , the optimal search strategy turns out to be Brownian (shaded area).
, the optimal search strategy turns out to be Brownian (shaded area).
Search with External Drift.
Once an external drift biases the walker, the behavior of the arrival to the target as a function of the initial position 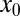 and the drift strength v changes dramatically. In particular, there may exist a finite residual survival probability
and the drift strength v changes dramatically. In particular, there may exist a finite residual survival probability 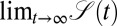 mirroring cases when the walker is being pushed away from the target and never reaches it. From dimensional analysis it is straightforward to show that the search reliability P solely depends on the single parameter
mirroring cases when the walker is being pushed away from the target and never reaches it. From dimensional analysis it is straightforward to show that the search reliability P solely depends on the single parameter 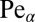 , and is thus a meaningful characteristic for the search process. The search reliability
, and is thus a meaningful characteristic for the search process. The search reliability 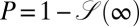 for the biased case is displayed for a large range of the generalized Péclet number
for the biased case is displayed for a large range of the generalized Péclet number 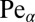 in Fig. 4, Left. In addition, Fig. 4, Right shows a blow-up for the behavior at small
in Fig. 4, Left. In addition, Fig. 4, Right shows a blow-up for the behavior at small 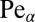 . These results are obtained from numerical solution of Eq. 2 and are thus not restricted to small
. These results are obtained from numerical solution of Eq. 2 and are thus not restricted to small 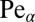 values. We confirmed the FFPE results with simulation for
values. We confirmed the FFPE results with simulation for 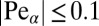 (Fig. 4, Right) based on the Langevin Eq. S3 in SI Text with very good agreement. Each simulations point was obtained as a ratio of particles to the overall number of 10,000 runs. We estimated the error bars by computation of the deviations P for each of 1,000 consecutive runs. We note that higher values of
(Fig. 4, Right) based on the Langevin Eq. S3 in SI Text with very good agreement. Each simulations point was obtained as a ratio of particles to the overall number of 10,000 runs. We estimated the error bars by computation of the deviations P for each of 1,000 consecutive runs. We note that higher values of 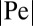 require longer Langevin equation simulations times.
require longer Langevin equation simulations times.
Fig. 4.

(Left) Dependence of the search reliability 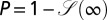 on the generalized Péclet number
on the generalized Péclet number 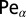 . The residual value P quantifies how likely it is that the walker eventually localizes the target. Lines are from numerical solution of Eq. 2. The blue (solid) line corresponds to Brownian search
. The residual value P quantifies how likely it is that the walker eventually localizes the target. Lines are from numerical solution of Eq. 2. The blue (solid) line corresponds to Brownian search 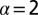 , the green (dashed) line represents LF search with
, the green (dashed) line represents LF search with 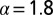 , and the red (dotted) line stands for
, and the red (dotted) line stands for 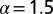 . Finally, the black (dashed-dotted) line is for
. Finally, the black (dashed-dotted) line is for 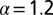 . (Right) Same for small values of
. (Right) Same for small values of 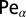 . In addition to the numerical solution of Eq. 2, the symbols and error bars are obtained from Langevin equation simulations of LF trajectories based on Eq. S3.
. In addition to the numerical solution of Eq. 2, the symbols and error bars are obtained from Langevin equation simulations of LF trajectories based on Eq. S3.
As seen from Fig. 4, in the downhill case when the searcher is pushed toward the target by the stream (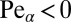 ) the best strategy in terms of the search reliability P is always that of Brownian search, in which case
) the best strategy in terms of the search reliability P is always that of Brownian search, in which case 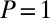 for all values of
for all values of 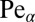 . The LF searcher in this regime always fares worse (
. The LF searcher in this regime always fares worse (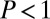 ), the discrepancy increasing for smaller values of the stable index. This is due to the occurrence of leap-overs across the target for LFs. In the presence of a strong drift, the search reliability P becomes considerably smaller. The opposite tendency is observed for the uphill case when the walker needs to move against the stream toward the target (
), the discrepancy increasing for smaller values of the stable index. This is due to the occurrence of leap-overs across the target for LFs. In the presence of a strong drift, the search reliability P becomes considerably smaller. The opposite tendency is observed for the uphill case when the walker needs to move against the stream toward the target (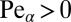 ). Now, LFs with a smaller stable index perform better, owing to the possibility of approaching the target faster with fewer jumps. We note, however, that the absolute gain of LF versus Brownian search in the uphill case is considerably smaller than the loss in the downhill scenario.
). Now, LFs with a smaller stable index perform better, owing to the possibility of approaching the target faster with fewer jumps. We note, however, that the absolute gain of LF versus Brownian search in the uphill case is considerably smaller than the loss in the downhill scenario.
The search efficiency ℰ is affected by the external stream even more dramatically than the search reliability P, as shown in Fig. 5. Here, the initial position is fixed at 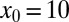 in Fig. 5, and at
in Fig. 5, and at 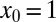 in Fig. 5, Inset. Black (full) lines correspond to the downhill case with
in Fig. 5, Inset. Black (full) lines correspond to the downhill case with 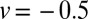 and red (dashed) curves to the uphill case with
and red (dashed) curves to the uphill case with 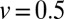 . The neutral case
. The neutral case 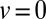 is shown by the blue (dotted) line. For
is shown by the blue (dotted) line. For 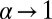 the curves converge at zero efficiency, in the case
the curves converge at zero efficiency, in the case 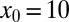 they almost coincide below
they almost coincide below 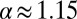 . In the case without bias and
. In the case without bias and 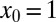 , consistent with our observations in the drift-free case above, the optimal strategy remains Brownian (Fig. 3). In contrast, for the larger initial separation
, consistent with our observations in the drift-free case above, the optimal strategy remains Brownian (Fig. 3). In contrast, for the larger initial separation 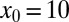 in the downhill case the optimal search strategy is also Brownian, whereas without bias we found
in the downhill case the optimal search strategy is also Brownian, whereas without bias we found 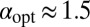 . In the uphill case the optimal stable index is shifted to
. In the uphill case the optimal stable index is shifted to 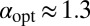 . The delicate behaviors of ℰ, P, and
. The delicate behaviors of ℰ, P, and 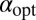 constitute our other important finding.
constitute our other important finding.
Fig. 5.

Search efficiency as a function of the stable index α for initial positions 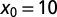 and
and 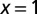 (Inset, same axes). We show the downhill case (
(Inset, same axes). We show the downhill case ( , solid black curve), neutral case (
, solid black curve), neutral case (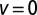 , blue dotted curve), and uphill case (
, blue dotted curve), and uphill case (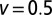 , red dashed curve). For a short distance between the initial position and target, the Brownian search always has the maximal efficiency, whereas for larger separation
, red dashed curve). For a short distance between the initial position and target, the Brownian search always has the maximal efficiency, whereas for larger separation 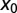 in the uphill (
in the uphill (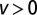 ) and neutral (
) and neutral (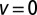 ) cases an optimum value different from 2 for the stable index α exists.
) cases an optimum value different from 2 for the stable index α exists.
Discussion
What is now the best random search strategy? As we showed here the answer to this question depends on what is more important—to reach the target quickly or to locate it most reliably—and on the physical situation: the presence of an external bias and the initial separation between searcher and target. The main message from this study is that LF search and its optimization is sensitive to the exact conditions.
Specifically, we investigated the performance of LF search models along or against an external stream. Defining the search efficiency as the average inverse arrival time 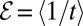 to the target, we obtained a versatile measure to quantify search processes when the search space does not have a constant target density. The search efficiency ℰ reproduces the features of Brownian search and works well for both unbiased and biased search processes, unlike the similar but frequently applied construct
to the target, we obtained a versatile measure to quantify search processes when the search space does not have a constant target density. The search efficiency ℰ reproduces the features of Brownian search and works well for both unbiased and biased search processes, unlike the similar but frequently applied construct 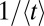 . In terms of this efficiency we investigate the optimal search strategy, comparing Lévy and Brownian search processes. Without an external bias, it turns out that the optimal strategy depends on the initial separation between the searcher and the target: For small separations Brownian motion is the most efficient way of finding the target. On increasing this separation LFs become more and more efficient in comparison with Brownian search: Consistent with previous findings in different scenarios, the stable index α decreases toward unity in the limit of very large initial searcher–target separation. In particular, we find that despite the frequent claim that LFs with
. In terms of this efficiency we investigate the optimal search strategy, comparing Lévy and Brownian search processes. Without an external bias, it turns out that the optimal strategy depends on the initial separation between the searcher and the target: For small separations Brownian motion is the most efficient way of finding the target. On increasing this separation LFs become more and more efficient in comparison with Brownian search: Consistent with previous findings in different scenarios, the stable index α decreases toward unity in the limit of very large initial searcher–target separation. In particular, we find that despite the frequent claim that LFs with 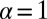 are most efficient for sparse targets, depending on the parameters of the search space the optimal stable index may range in the whole interval between unity and 2, and thus may include Brownian motion as the most efficient strategy.
are most efficient for sparse targets, depending on the parameters of the search space the optimal stable index may range in the whole interval between unity and 2, and thus may include Brownian motion as the most efficient strategy.
The analysis in terms of the complementary search reliability 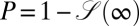 shows that when the initial position of the searcher with respect to the target is along the stream, the optimal search strategy is always Brownian, owing to the combined effect of biased motion and absence of the leap-overs. This result is even true for a small bias. The average search time in this case is simply given in terms of the classic ratio of initial searcher–target separation and the drift velocity. When the searcher needs to reach the target against the stream, in line with naive expectations, LFs indeed provide the better search strategy. Remarkably, the absolute gain from using the Brownian strategy instead of LFs in the downhill scenario is significantly larger than the loss from using Brownian motion instead of LFs in the uphill case. Depending on the details of the search space, without prior knowledge of the strength and direction of external streams, the choice of a Brownian strategy might therefore be advantageous overall, in contrast to the LF hypothesis. This point is further illustrated in terms of the search efficiency as function of the initial searcher–target separation in the presence of a bias in Fig. 6. These observations may be of particular importance to swimming or airborne searchers, because streams occur most naturally there, or when the searcher itself prefers one direction, for instance owing to prior experience. Our findings may also be relevant for computational search algorithms in biased landscapes (42). Of course, more quantitative statements need a detailed investigation for a given, concrete system.
shows that when the initial position of the searcher with respect to the target is along the stream, the optimal search strategy is always Brownian, owing to the combined effect of biased motion and absence of the leap-overs. This result is even true for a small bias. The average search time in this case is simply given in terms of the classic ratio of initial searcher–target separation and the drift velocity. When the searcher needs to reach the target against the stream, in line with naive expectations, LFs indeed provide the better search strategy. Remarkably, the absolute gain from using the Brownian strategy instead of LFs in the downhill scenario is significantly larger than the loss from using Brownian motion instead of LFs in the uphill case. Depending on the details of the search space, without prior knowledge of the strength and direction of external streams, the choice of a Brownian strategy might therefore be advantageous overall, in contrast to the LF hypothesis. This point is further illustrated in terms of the search efficiency as function of the initial searcher–target separation in the presence of a bias in Fig. 6. These observations may be of particular importance to swimming or airborne searchers, because streams occur most naturally there, or when the searcher itself prefers one direction, for instance owing to prior experience. Our findings may also be relevant for computational search algorithms in biased landscapes (42). Of course, more quantitative statements need a detailed investigation for a given, concrete system.
Fig. 6.

Search efficiency versus initial position 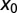 for
for 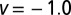 (i.e., negative initial walker–target separations
(i.e., negative initial walker–target separations 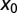 correspond to uphill motion).
correspond to uphill motion).
Our analysis was performed for a blind, saltatory random LF search in one spatial dimension. What would be expected if these conditions were relaxed? First, moving from one to two spatial dimensions, regular Brownian motion remains recurrent, that is, the sample path is fully connected and thus space-filling in both one and two dimensions. LFs with 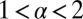 are recurrent in one dimension but always transient in two dimensions. We therefore expect the results discovered herein to be equally relevant in two dimensions. In both one and two dimensions (linearly or radially) LFs are distinguished by the occurrence of leap-overs, owing to which the target location may become less efficient than for Brownian search. Most search processes indeed fall in the category of (effectively) one or two dimensions. For instance, they are one-dimensional in streams, along coastlines, or at forest–meadow and other borders. For unbounded search processes as performed by birds or fish of prey, the motion in the vertical dimension shows a much smaller span than the radial horizontal motion, and thus becomes effectively two dimensional. Second, when we modify the condition of blind search and allow the walker to look out for prey while relocating, in one dimension this would obviously completely change the picture in favor of LFs with their long unidirectional steps. However, in two dimensions the radial leap-overs would still hamper the detection of the target as long as it is not exactly crossed during a step, and our qualitative results remain relevant.
are recurrent in one dimension but always transient in two dimensions. We therefore expect the results discovered herein to be equally relevant in two dimensions. In both one and two dimensions (linearly or radially) LFs are distinguished by the occurrence of leap-overs, owing to which the target location may become less efficient than for Brownian search. Most search processes indeed fall in the category of (effectively) one or two dimensions. For instance, they are one-dimensional in streams, along coastlines, or at forest–meadow and other borders. For unbounded search processes as performed by birds or fish of prey, the motion in the vertical dimension shows a much smaller span than the radial horizontal motion, and thus becomes effectively two dimensional. Second, when we modify the condition of blind search and allow the walker to look out for prey while relocating, in one dimension this would obviously completely change the picture in favor of LFs with their long unidirectional steps. However, in two dimensions the radial leap-overs would still hamper the detection of the target as long as it is not exactly crossed during a step, and our qualitative results remain relevant.
Third we address the question as to what will change if we relax the saltatory motion condition and introduce a finite velocity of propagation. This corresponds to the picture of LWs, random walks with a spatiotemporal coupling (43). The coupling penalizes long jumps by connecting these with long waiting times. The resulting, non-Markovian process can be thought of as a random change between velocity modes with different directions. In the simplest case the probability density function of LWs are characterized by two counter propagating δ peaks with time-decreasing weight. In between these peaks a Lévy stable propagator emerges. Thus, at longer times, we would expect the same advantages to LW search as we know it from LF search, as, for instance, observed in ref. 36. A detailed study of leap-overs and the connected effects to search for LWs remains, however, elusive.
There exist alternative random search models, including intermittent processes switching between local diffusive search and blind, ballistic relocations (44–47), or persistent random walk models with finite-ranged directional correlations (48). However, the central advantage of LF strategies is their robustness: Although other models perform equally well compared with LFs when their parameters are optimized for specific, fixed environmental conditions (e.g., the target density), owing to their scale-free nature LFs remain close to optimal even when these conditions change (36), whereas the other search strategies then fare significantly worse. Moreover, owing to their connection with critical phenomena (49), physicists tend to promote LFs for efficient search (14, 50, 51). For their wide popularity, it is important to put the LF search efficiency and reliability into perspective, as is done here.
It will be interesting to extend our findings to scenarios with a finite search space, in which LFs eventually fill the entire area and thus the search always becomes successful (compare ref. 52) and to generalize this approach to higher dimensions and processes with spatiotemporal coupling. Additionally, our results for a single or few targets should be compared with the situation of a constant target density.
Supplementary Material
Acknowledgments
V.V.P. acknowledges discussions with A. Cherstvy and financial support from Deutsche Forschungsgemeinschaft Project PA 2042/1-1. R.M. acknowledges financial support from the Academy of Finland (FiDiPro scheme).
Footnotes
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1320424111/-/DCSupplemental.
References
- 1.Bénichou O, Loverdo C, Moreau M, Voituriez R. Intermittent search strategies. Rev Mod Phys. 2011;83(1):81–129. doi: 10.1103/PhysRevE.80.031146. [DOI] [PubMed] [Google Scholar]
- 2.Bressloff PC, Newby JM. Stochastic models of intracellular transport. Rev Mod Phys. 2013;85(1):135–196. [Google Scholar]
- 3.Shlesinger MF. Random searching. J Phys A. 2009;42:434001. [Google Scholar]
- 4.Viswanathan GE, da Luz MGE, Raposo EP, Stanley HE. The Physics of Foraging: An Introduction to Random Searches and Biological Encounters. New York: Cambridge Univ Press; 2011. [Google Scholar]
- 5.Shlesinger MF, Klafter J. In: On Growth and Form. Stanley HE, Ostrowsky N, editors. Dordrecht, The Netherlands: Martinus Nijhoff; 1986. [Google Scholar]
- 6.Nathan R, et al. A movement ecology paradigm for unifying organismal movement research. Proc Natl Acad Sci USA. 2008;105(49):19052–19059. doi: 10.1073/pnas.0800375105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.James A, Pitchford JW, Plank MJ. Efficient or inaccurate? Analytical and numerical modelling of random search strategies. Bull Math Biol. 2010;72(4):896–913. doi: 10.1007/s11538-009-9473-z. [DOI] [PubMed] [Google Scholar]
- 8.Sims DW, et al. Scaling laws of marine predator search behaviour. Nature. 2008;451(7182):1098–1102. doi: 10.1038/nature06518. [DOI] [PubMed] [Google Scholar]
- 9.Humphries NE, et al. Environmental context explains Lévy and Brownian movement patterns of marine predators. Nature. 2010;465(7301):1066–1069. doi: 10.1038/nature09116. [DOI] [PubMed] [Google Scholar]
- 10.Burrow JF, Baxter PD, Pitchford JW. Lévy processes, saltatory foraging, and superdiffusion. Math Model Nat Phenom. 2008;3(3):115–130. [Google Scholar]
- 11.Sims DW, Witt MJ, Richardson AJ, Southall EJ, Metcalfe JD. Encounter success of free-ranging marine predator movements across a dynamic prey landscape. Proc Biol Sci. 2006;273(1591):1195–1201. doi: 10.1098/rspb.2005.3444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Houghton JD, Doyle TK, Wilson MW, Davenport J, Hays GC. Jellyfish aggregations and leatherback turtle foraging patterns in a temperate coastal environment. Ecology. 2006;87(8):1967–1972. doi: 10.1890/0012-9658(2006)87[1967:jaaltf]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 13.Bradshaw CJA, Hindell MA, Sumner MD, Michael KJ. Loyalty pays: Potential life history consequences of fidelity to marine foraging regions by southern elephant seals. Anim Behav. 2004;68:1349–1360. [Google Scholar]
- 14.Viswanathan GM, et al. Optimizing the success of random searches. Nature. 1999;401(6756):911–914. doi: 10.1038/44831. [DOI] [PubMed] [Google Scholar]
- 15.Viswanathan GM, et al. Lévy flight search patterns of wandering albatrosses. Nature. 1996;381(6581):413–415. [Google Scholar]
- 16.de Jager M, Weissing FJ, Herman PMJ, Nolet BA, van de Koppel J. Lévy walks evolve through interaction between movement and environmental complexity. Science. 2011;332(6037):1551–1553. doi: 10.1126/science.1201187. [DOI] [PubMed] [Google Scholar]
- 17.Ramos-Fernández G, et al. Lévy walk patterns in the foraging movements of spider monkeys (Ateles geoffroyi) Behav Ecol Sociobiol. 2004;55(3):223–230. [Google Scholar]
- 18.Atkinson RPD, Rhodes CJ, MacDonald DW, Anderson RM. Scale-free dynamics in the movement patterns of jackals. Oikos. 2002;98(1):134–140. [Google Scholar]
- 19.González MC, Hidalgo CA, Barabási A-L. Understanding individual human mobility patterns. Nature. 2008;453(7196):779–782. doi: 10.1038/nature06958. [DOI] [PubMed] [Google Scholar]
- 20.Brockmann D. Following the money. Phys World. 2010;23(2):31–34. [Google Scholar]
- 21.Korobkova E, Emonet T, Vilar JMG, Shimizu TS, Cluzel P. From molecular noise to behavioural variability in a single bacterium. Nature. 2004;428(6982):574–578. doi: 10.1038/nature02404. [DOI] [PubMed] [Google Scholar]
- 22.Tu Y, Grinstein G. How white noise generates power-law switching in bacterial flagellar motors. Phys Rev Lett. 2005;94(20):208101. doi: 10.1103/PhysRevLett.94.208101. [DOI] [PubMed] [Google Scholar]
- 23.van Dartel M, Postma E, van den Herik J, de Croon G. Macroscopic analysis of robot foraging behaviour. Connect Sci. 2004;16(3):169–181. [Google Scholar]
- 24.Boyer D, et al. Scale-free foraging by primates emerges from their interaction with a complex environment. Proc Biol Sci. 2006;273(1595):1743–1750. doi: 10.1098/rspb.2005.3462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Jansen VAA, Mashanova A, Petrovskii S. Comment on “Lévy walks evolve through interaction between movement and environmental complexity”. Science. 2012;335(6071):918. doi: 10.1126/science.1215747. author reply 918. [DOI] [PubMed] [Google Scholar]
- 26.Mashanova A, Oliver TH, Jansen VAA. Evidence for intermittency and a truncated power law from highly resolved aphid movement data. J R Soc Interface. 2010;7(42):199–208. doi: 10.1098/rsif.2009.0121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Petrovskii S, Mashanova A, Jansen VAA. Variation in individual walking behavior creates the impression of a Levy flight. Proc Natl Acad Sci USA. 2011;108(21):8704–8707. doi: 10.1073/pnas.1015208108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Edwards AM, et al. Revisiting Lévy flight search patterns of wandering albatrosses, bumblebees and deer. Nature. 2007;449(7165):1044–1048. doi: 10.1038/nature06199. [DOI] [PubMed] [Google Scholar]
- 29.Humphries NE, Weimerskirch H, Queiroz N, Southall EJ, Sims DW. Foraging success of biological Lévy flights recorded in situ. Proc Natl Acad Sci USA. 2012;109(19):7169–7174. doi: 10.1073/pnas.1121201109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Metzler R, Klafter J. The random walk’s guide to anomalous diffusion: A fractional dynamics approach. Phys Rep. 2000;339:1–77. [Google Scholar]
- 31.Metzler R, Klafter J. The restaurant at the end of the random walk: Recent developments in the description of anomalous transport by fractional dynamics. J Phys A. 2004;37:R161–R208. [Google Scholar]
- 32.Chechkin AV, Metzler R, Gonchar VY, Klafter J, Tanatarov LV. First passage time density for Lévy flight processes and the failure of the method of images. J Phys A. 2003;36:L537–L544. [Google Scholar]
- 33.Koren T, Lomholt MA, Chechkin AV, Klafter J, Metzler R. Leapover lengths and first passage time statistics for Lévy flights. Phys Rev Lett. 2007;99(16):160602. doi: 10.1103/PhysRevLett.99.160602. [DOI] [PubMed] [Google Scholar]
- 34.Koren T, Chechkin AV, Klafter J. On the first passage time and leapover properties of Lévy motions. Physica A. 2007;379(1):10–22. [Google Scholar]
- 35.Sato K-I. Lévy Processes and Infinitely Divisible Distributions. Cambridge, UK: Cambridge Univ Press; 1999. [Google Scholar]
- 36.Lomholt MA, Koren T, Metzler R, Klafter J. Lévy strategies in intermittent search processes are advantageous. Proc Natl Acad Sci USA. 2008;105(32):1055–11059. [Google Scholar]
- 37.Lomholt MA, Ambjörnsson T, Metzler R. Optimal target search on a fast-folding polymer chain with volume exchange. Phys Rev Lett. 2005;95(26):260603. doi: 10.1103/PhysRevLett.95.260603. [DOI] [PubMed] [Google Scholar]
- 38.Bartumeus F, da Luz MGE, Viswanathan GM, Catalan J. Animal search strategies: A quantitative random-walk analysis. Ecology. 2005;86(1):3078–3087. [Google Scholar]
- 39.Bartumeus F, Levin SA. Fractal reorientation clocks: Linking animal behavior to statistical patterns of search. Proc Natl Acad Sci USA. 2008;105(49):19072–19077. doi: 10.1073/pnas.0801926105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Reynolds AM. Optimal random Lévy-loop searching: New insights into the searching behaviours of central-place foragers. EPL. 2008;82(2):20001. [Google Scholar]
- 41.Reynolds AM. Scale-free animal movement patterns: Lévy walks outperform fractional Brownian motions and fractional Lévy motions in random search scenarios. J Phys A. 2009;42:434006. [Google Scholar]
- 42.Pavlyukevich I. Lévy flights, non-local search and simulated annealing. J Comput Phys. 2007;226(2):1830–1844. [Google Scholar]
- 43.Klafter J, Blumen A, Shlesinger MF. Stochastic pathway to anomalous diffusion. Phys Rev A. 1987;35(7):3081–3085. doi: 10.1103/physreva.35.3081. [DOI] [PubMed] [Google Scholar]
- 44.Bénichou O, Coppey M, Moreau M, Suet PH, Voituriez R. Optimal search strategies for hidden targets. Phys Rev Lett. 2005;94(19):198101. doi: 10.1103/PhysRevLett.94.198101. [DOI] [PubMed] [Google Scholar]
- 45.Loverdo C, Bénichou O, Moreau M, Voiturierz R. Enhanced reaction kinetics in biological cells. Nat Phys. 2008;4(2):134–137. [Google Scholar]
- 46.Benhamou S. How many animals really do the Lévy walk? Ecology. 2007;88(8):1962–1969. doi: 10.1890/06-1769.1. [DOI] [PubMed] [Google Scholar]
- 47.Reynolds A. Adaptive Lévy walks can outperform composite Brownian walks in non-destructive random walk scenarios. Physica A. 2009;388(5):561–569. [Google Scholar]
- 48.Bovet P, Benhamou S. Spatial analysis of animals’ movements using a correlated random walk model. J Theor Biol. 1988;131(4):419–433. [Google Scholar]
- 49.Stanley HE. Introduction to Phase Transitions and Critical Phenomena. New York: Oxford Univ Press; 1988. [Google Scholar]
- 50.Bartumeus F, Catalan J, Fulco UL, Lyra ML, Viswanathan GM. Optimizing the encounter rate in biological interactions: Lévy versus Brownian strategies. Phys Rev Lett. 2002;88(9):097901. doi: 10.1103/PhysRevLett.88.097901. [DOI] [PubMed] [Google Scholar]
- 51.Barabási A-L. Bursts: The Hidden Patterns Behind Everything We Do, from Your E-mail to Bloody Crusades. New York: Dutton; 2010. [Google Scholar]
- 52.Vahabi M, Schulz JHP, Shokri B, Metzler R. Area coverage of radial Lévy flights with periodic boundary conditions. Phys Rev E Stat Nonlin Soft Matter Phys. 2013;87(4):042136. doi: 10.1103/PhysRevE.87.042136. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.