

Abstract
Phagocytic leukocytes, predominantly macrophages, not only ingest and destroy invading pathogens, but are charged with clearing dead and dying host cells. The process of engulfing apoptotic cells is called efferocytosis and has long been appreciated for its role in the resolution of inflammation. New evidence is emerging that efferocytosis represents a double-edged sword in microbial immunity. While efferocytosis of influenza and Mycobacterium tuberculosis-infected cells results in pathogen destruction, efferocytosis of Leishmania-infected neutrophils may promote infection. Understanding how macrophages, dendritic cells (DC) and neutrophils process pathogens encased within a dying cell could lead to the development of novel therapeutics that simultaneously suppress inflammation and promote pathogen clearance.
The Many Jobs of the Macrophage
Macrophages function primarily as phagocytes during innate immune responses, but perform diverse actions in homeostasis and disease. During infection, detection of pathogen-associated molecular patterns (PAMPs) by macrophages leads to the production of cytokines and chemokines, promoting the recruitment of other cells and triggering an immune response. By degrading pathogen-derived antigens and presenting them to T cells, macrophages can promote or perpetuate adaptive immunity [1]. Moreover, by engulfing and degrading bacteria, macrophages sterilize tissue, resolve inflammation, and prevent further stimulation of the immune system.
In addition to their roles in the immune response to infection, macrophages serve a second, possibly more important function. They are the cell type primarily responsible for efferocytosis, the process of engulfing and eliminating apoptotic cells. Apoptosis, or programmed cell death, is necessary for the development of multicellular organisms, and even in adults, thousands of cells die and are replaced every second [2]. Two key processes ensure that apoptosis is a non-inflammatory form of cell death: 1) the dying cell’s contents are packaged into apoptotic bodies with intact plasma membranes [3]; and 2) the apoptotic bodies are engulfed by phagocytes. Without their swift removal via efferocytosis, apoptotic bodies disintegrate and release their intracellular contents. This process, known as secondary necrosis, leads to inflammation and can contribute to autoimmunity [4].
Efferocytosis: A Distinct Process
Although efferocytosis resembles phagocytosis, it is a distinct process, mediated by specific receptors, bridging molecules, and downstream signaling pathways. Phagocytosis by macrophages has been extensively reviewed [5,6]. Briefly, it is the process by which macrophages capture and engulf small particles or microbes. The engulfment of microbes relies on the signaling of Rho family GTPases, specifically RhoA, and subsequent actin cytoskeleton rearrangement [7]. The macrophage forms actin protrusions around the microbe, which fuse, resulting in complete engulfment (Figure 1). The resulting intracellular vesicle is termed a phagosome and initially possesses an environment similar to the extracellular milieu. Maturation of the phagosome is a step-wise process involving Rab GTPase-driven fusion of endosomes and lysosomes. These small organelles dock, releasing hydrolytic and catabolic enzymes. The resulting phagolysosome continues to acidify to a pH of nearly 4, leading to the destruction of the microbe [8].
Figure 1. Efferocytosis is a distinct process from phagocytosis.

In phagocytosis, increased levels of RhoA result in actin polymerization and stress fiber formation, allowing receptor-mediated uptake of microbes. Fusion of the actin projections leads to internalization of the bacteria in a phagosome, which undergoes stepwise maturation, leading to degradation of the pathogen. This process can lead to both tissue sterilization and antigen presentation. In contrast, efferocytosis results from increased levels of Rac1, causing formation of lamellipodia and membrane ruffles upon actin polymerization, leading to the internalization of infected and ‘double-wrapped’ apoptotic bodies in an efferosome. From here, the ‘double-wrapped’ pathogen can be destroyed if the efferosome undergoes acidification, similar to the process of phagolysosome fusion. However, some pathogens appear to be able to use the process of efferocytosis to diseminate and disperse.
The engulfment of apoptotic cells by efferocytosis more resembles macropinocytosis [9]. The receptors that recognize an apoptotic cell and mediate efferocytosis are distinct from those that mediate phagocytosis (see below), and once an apoptotic cell is engulfed, the intracellular signaling further diverges. During efferocytosis, RhoA activity is suppressed and Rac1 activity coordinates the engulfment of the apoptotic body and formation of a spacious phagosome, termed an efferosome, which surrounds the newly engulfed apoptotic cell (Figure 1). While new GTPases are being identified that have a role in efferosome maturation, the end result is similar to phagocytosis: successive lysosome fusion events deliver hydrolytic enzymes to the maturing efferosome. This together with its gradual acidification ultimately establishes harsh conditions that destroy the apoptotic cell [8,10].
Find Me, Catch Me, Eat Me
The process of efferocytosis is highly regulated and begins with the apoptotic cell. An early event during apoptosis is the exposure of phosphatidylserine (PS) to the exofacial leaflet of the phospholipid membrane (Figure 2A). Another early event is the release of chemokines [7,11], or “Find Me” signals by the dying cell that recruits macrophages to the site of cell death (Figure 2A). In response, the macrophage increases the expression of bridge molecules and receptors that act as tethers for the “Catch Me” step (Figure 2B). Finally, the “Eat Me” step occurs when the macrophage engulfs the dying cell and generates the large spacious efferosome (Figure 2C). Efferocytosis is said to be anti-inflammatory as macrophages produce IL-10, TGF-β, and PGE2 during the engulfment of apoptotic cells in model systems [12–15]. Thus, efferocytosis is important for the resolution of inflammation not only because it removes dead cells, relieves tissue congestion, and prevents release of phlogistic cellular contents, but also promotes tissue repair and wound healing [16,17].
Figure 2. Efferocytosis involves the regulated uptake and degradation of apoptotic bodies.
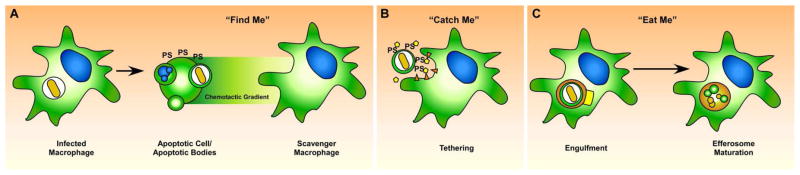
When an infected cell undergoes apoptosis (A), the pathogen and its antigens are presumably packaged along with other intracellular contents into apoptotic bodies which have intact cellular membranes. During apoptosis, the dying cell produces “Find Me” signals, such as chemokines that recruit macrophages and other phagocytic cells to the site of the dying cell, along with the accumulation of of phosphatidylserine (PS) on the exofacial leaflet. In the “Catch Me” phase, recruited macrophages express specific receptors and bridge molecules that bind to PS or other distinct ligands on the apoptotic cell (B). Binding between the apoptotic cell and the scavenger macrophage activates Rac1, leading to actin reorganization and production of projections that surround the infected apoptotic cell, ulimately surrounding and engulfing the apoptotic cell in the efferosome (C). The efferosome undergoes step-wise maturation that includes fusion with lysosomes and endosomes, leading to rapid degradation of both the apoptotic cell and the intracellular pathogen.
Efferocytosis and Pathogen Clearance
Some pathogens have evolved mechanisms to survive intracellularly as a means of immune evasion. These pathogens have a remarkable diversity of virulence mechanisms that target and modify host physiological processes to persist within their host cell [18]. Host cells have a variety of ways to deal with intracellular infection, one of them being apoptosis. Activation of either the intrinsic (caspase-9-dependent) and extrinsic (caspase-8-dependent) cell death pathways can lead to apoptosis of the infected cell. Cytotoxic T lymphocytes can induce apoptosis in infected cells through exocytosis of cytotoxic granules that contain perforin and granzymes or via the Fas/FasL pathway [19–21]. Given the abundance of cell death that accompanies intracellular infection, researchers have begun to investigate the fate of microbes inhabiting a cell that undergoes apoptosis and is engulfed by a phagocyte. Two possibilities have been described: the pathogen can be destroyed along with the engulfed apoptotic cell (host antimicrobial activity) or the pathogen can use the process of efferocytosis to disperse into new cellular hosts (“Trojan Horse” model). The intersection of efferocytosis and intracellular pathogen fate has only been studied in a few diseases. However any pathogen that relies on intracellular infection, particularly where apoptosis or cytolysis is linked to control, should be investigated for the role efferocytosis may play.
Examples of Efferocytosis and Microbial Immunity
During Leishmania major infection, efferocytosis of infected neutrophils may promote infection by the “Trojan Horse” model of disease. The infectious sand fly bite recruits neutrophils that rapidly phagocytose L. major metacyclic promastigotes and this is crucial for the establishment of infection. Parasite infection induces neutrophils apoptosis, which then attracts macrophages and DCs. In vitro models demonstrate that macrophages efferocytose these infected apoptotic neutrophils [22–25]. In vivo, DCs have also been found to engulf these infected apoptotic neutrophils [26]. Unfortunately, whether L. major survived after DC efferocytosis of infected neutrophils was not assessed. As it appears that L. major uses apoptotic neutrophils as a Trojan Horse to gain entry into it’s preferred terminal host cells, the macrophage, it is important to understand how the parasite survives in the efferosome.
Mycobacterium tuberculosis typically avoids inducing apoptotic death of the macrophage it infects and instead skews the death modality towards a necrotic fate, which facilitates its spread into neighboring cells and avoids immune clearance [27]. For reasons not entirely understood, some M. tuberculosis-infected macrophages die by apoptosis, which has long been known to be associated with suppression of bacterial growth [28,29]. M. tuberculosis-infected macrophages that die by apoptosis are rapidly engulfed by uninfected macrophages via efferocytosis and this process is responsible for the bactericidal effect associated with apoptosis [30]. Blocking efferocytosis both in vitro and in vivo abrogates bacterial control. M. tuberculosis killing post-efferocytosis is due to lysosome fusion as both the internalized apoptotic bodies and the bacteria are degraded simultaneously by the degradative enzymes delivered via the lysosomes. It is interesting to speculate why efferocytosis allows lysosome recruitment to the M. tuberculosis-containing phagosome while M. tuberculosis typically arrests phagosome maturation following phagocytosis of naked bacteria. The mechanism of efferocytic killing remains an important avenue for further research and will be discussed more in the next section. Similarly, Streptococcus pneumoniae infection of macrophages causes apoptosis and is linked with pathogen control [19]. This may suggest that efferocytosis is involved in control of the infection.
Mycobacterium marinum is a natural marine pathogen, and efferocytosis has similarly been described as a means to control this microbe in a zebra fish model of innate M. tuberculosis infection [31]. M. marinum-infected macrophages dying by apoptosis are engulfed by recruited neutrophils. Following efferocytosis of the M. marinum-infected macrophage, the neutrophil generates an NADPH-dependent oxidative burst, which kills M. marinum [32]. What is unknown is what is stimulating the ROS production. Oxidative burst is a mechanism used by innate cells to control pathogens; however, its upregulation is dependent upon pathogen recognition [33]. It is unknown whether an efferocytic cell senses the pathogen within the apoptotic cell. Alternatively, all cells generate an NADPH-dependent oxidative burst following apoptotic cell engulfment in response to the large ‘meal’ of oxidized lipids [34]. The cell employs many compensatory mechanisms to ensure that an apoptotic cell can be engulfed without damaging or triggering apoptosis in the efferocytic cell [35].
Influenza A induces apoptosis of pulmonary epithelial cells and in cultured cell lines. Co-culture of infected epithelial cells or in vitro cell lines with macrophages leads to efferocytosis [36]. Importantly, the macrophages were found to contain viral particles inside their efferosomes after efferocytosis. Blocking apoptosis inhibits efferocytosis of these virus-infected cells. During efferocytosis, there was a decline in viral titers, suggesting that for influenza A, efferocytosis is an antiviral mechanism. These data supports the idea that efferocytosis may have microbicidal activity against multiple pathogens [37].
How Efferocytosis Mediates Pathogen Clearance: Potential Mechanisms
Why efferocytosis leads to pathogen killing while the antimicrobial effect of phagocytosis and phagolysosome fusion is frequently circumvented is an important avenue for research. A few potential mechanisms have been put forward to explain the bactericidal effects of efferocytosis (Fig.3). Many pathogens actively interfere with phagosome maturation. In the case of M. tuberculosis, several bioactive lipids decorate the surface of the bacterium, which interferes with endosome and lysosome recruitment, thus arresting phagosome maturation and enabling M. tuberculosis to live within a non-caustic and protected vesicle [38,39]. Perhaps ‘double-wrapped’ behind both an apoptotic cell membrane and efferosome sequesters M. tuberculosis’ virulence factors from finding their targets in the cytosol. Many intracellular pathogens, such as Legionella, Coxiella, and Salmonella, similarly rely on secreted proteins to exert control over their host cells and thus ‘double-wrapping’ may contribute to the clearance of other infections [40–42]. Additionally, a different subset of small GTPases regulates the maturation of an efferosome than a phagosome [8,43]. Perhaps these molecules are not the targets of pathogens’ arsenal and thus efferosome maturation can continue uninterrupted with the successive acidification and destruction of pathogen and apoptotic cell alike. Leishmania, however, may avoid efferocytic killing [22]. L. major amastigotes subsist within acidic lysosomes and thus remain immune to killing via this strategy [44]. Leishmania promastigotes extensively remodel the phagosome in which it resides, by altering the composition of the lipid bilayer, preventing lysosome fusion and manipulating the surrounding actin cytoskeletal network [45,46]. Perhaps the mechanisms of phagosome disruption employed by L. major are so thorough that efferosome maturation is similarly inhibited. Furthermore, autophagy contributes to clearance of many intracellular pathogens, and LC3 and other autophagic components may localize to the efferosome [47–49]. More work is required to understand how autophagy may contribute to control following efferocytosis.
Figure 3. Four Proposed Mechanisms of Efferocytic Control.

Following efferocytosis of an infected apoptotic body there are 4 potential mechanisms that contribute to microbial killing. (A) A microbe’s secreted virulence factors, which typically penetrate the phagosome, are trapped behind additional membrane from the apoptotic cell; consequently, they cannot reach their host targets. As it has been proposed that the machinery involved in efferosome maturation is different than phagosome maturation, (B), microbial virulence factors are ineffective at inhibiting efferosome maturation. Following efferocytosis, autophagic machinery can be recruited to the nascent efferosome (C). A, B, and C all ultimately allow lysosome fusion. In (D), reactive oxygen species generated in response to efferocytosis indirectly kills the pathogen.
Little is known about the signaling within macrophages following uptake of an infected apoptotic cell. Efferocytosis typically evokes anti-inflammatory cascades, such as IL-10 and TGF-β production by the engulfing cell, as a means to dampen any potential self-directed immunity generated from the presence of dying cells [12,50]. Efferocytosis of infected apoptotic cells presents the phagocyte with two simultaneously conflicting signals: PAMPs that drive inflammation, and apoptotic cell ligands that trigger an anti-inflammatory program. Efferocytosis of an infected apoptotic cell by DC elicits a different response than is triggered when a macrophage engulfs an infected apoptotic cell [51]. IL-6 and TGF-β production from DCs following efferocytosis of an infected apoptotic cell drive differentiation of Th17 responses [52]. The unique signals produced by macrophages and neutrophils following infected apoptotic cell efferocytosis remain unknown.
Further Directions
The role of efferocytosis in microbial immunity is just now emerging. We envision efferocytosis as a critical part of the immune system’s tool kit for pathogen destruction. Some microbes are easily killed by phagocytosis such as extracellular bacteria (Fig. 4a). For other more virulent pathogens, the macrophage requires help from immune cells. For example, T cell cytokines activate the macrophage to enhance the expression of bactericidal functions (Fig. 4b). Finally, if infection overwhelms the macrophage, cytotoxic T cells can recognize heavily infected cells and induce them to die by apoptosis, or the infected cell may undergo apoptosis on its own because of increased stress. Either way, this now allows recruited macrophages to engulf and destroy the pathogens (Fig. 4c)
Figure 4. Efferocytosis is one mechanism that the host can use to control infection.

Macrophages autonomous control (A) frequently happens with low virulence or extracellular pathogens following phagocytosis. For pathogens that are more adapted to the intracellular environment of the macrophage, the macrophage needs help and collaborates with T cells (B). T cell recognition of the macrophage leads to the elaboration of T cell cytokines such as IFNγ and TNF, which activate the macrophage and enhance phagolysomal fusion but also stimulates other antimicrobial pathways, which augments the ability of the macrophage to kill intracellular bacteria. Some pathogens can overwhelm macrophage defenses. Under these conditions, the macrophage may die by apoptosis (C) if it is recognized by CTL; alternately, cell intrinsic factors may trigger an apoptotic death. Death by apoptosis ultimately allows for efferocytosis.
Efferocytosis, as both a means of pathogen clearance and inflammation suppression may be amenable for pharmacological targeting. In fact, the concept of promoting efferocytosis is already being considered by the atherosclerosis field as a way to alleviate plaque formation [17]. Harnessing the body’s own means and mechanisms of pathogen killing and immunoregulation is an attractive way to combat disease. Especially at a time when drug resistance by bacteria and viruses is increasing, the ability to provide new treatments that augment the body’s own defenses without targeting the microbe may alleviate the looming drug resistance crisis and be effective against otherwise recalcitrant pathogens.
Highlights.
Apoptotic cell death commonly follows intracellular pathogen infection.
Macrophages engulf and digest dying cells via the process of efferocytosis.
Efferocytosis of an infected apoptotic cell can result in pathogen destruction
This process represents a novel innate immune antimicrobial function
Acknowledgments
Funding. This work was supported by R01 AI 098637awarded to S.M.B.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Neefjes J, Jongsma ML, Paul P, Bakke O. Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat Rev Immunol. 2011;11 :823–836. doi: 10.1038/nri3084. [DOI] [PubMed] [Google Scholar]
- 2.Savill J, Fadok V, Henson P, Haslett C. Phagocyte recognition of cells undergoing apoptosis. Immunol Today. 1993;14:131–136. doi: 10.1016/0167-5699(93)90215-7. [DOI] [PubMed] [Google Scholar]
- 3.Taylor RC, Cullen SP, Martin SJ. Apoptosis: controlled demolition at the cellular level. Nat Rev Mol Cell Biol. 2008;9:231–241. doi: 10.1038/nrm2312. [DOI] [PubMed] [Google Scholar]
- 4.Rodriguez-Manzanet R, Sanjuan MA, Wu HY, Quintana FJ, Xiao S, Anderson AC, Weiner HL, Green DR, Kuchroo VK. T and B cell hyperactivity and autoimmunity associated with niche-specific defects in apoptotic body clearance in TIM-4-deficient mice. Proc Natl Acad Sci U S A. 2010;107:8706–8711. doi: 10.1073/pnas.0910359107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Jaumouille V, Grinstein S. Receptor mobility, the cytoskeleton, and particle binding during phagocytosis. Curr Opin Cell Biol. 2011;23:22–29. doi: 10.1016/j.ceb.2010.10.006. [DOI] [PubMed] [Google Scholar]
- 6**.Underhill DM, Goodridge HS. Information processing during phagocytosis. Nat Rev Immunol. 2012;12:492–502. doi: 10.1038/nri3244. This excellent review addresses the ability of the host cell to modulate its internalization of a foreign particle or microbe and subsequent signaling based on the type of particle or microbe. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Korns D, Frasch SC, Fernandez-Boyanapalli R, Henson PM, Bratton DL. Modulation of macrophage efferocytosis in inflammation. Front Immunol. 2011;2:57. doi: 10.3389/fimmu.2011.00057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kinchen JM, Ravichandran KS. Phagosome maturation: going through the acid test. Nat Rev Mol Cell Biol. 2008;9:781–795. doi: 10.1038/nrm2515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9*.Hoffmann PR, deCathelineau AM, Ogden CA, Leverrier Y, Bratton DL, Daleke DL, Ridley AJ, Fadok VA, Henson PM. Phosphatidylserine (PS) induces PS receptor-mediated macropinocytosis and promotes clearance of apoptotic cells. J Cell Biol. 2001;155:649–659. doi: 10.1083/jcb.200108080. This review provides a detailed list of the GTPases involved in efferocytosis, from actin polymerization to phagosome maturation. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lu N, Zhou Z. Membrane trafficking and phagosome maturation during the clearance of apoptotic cells. Int Rev Cell Mol Biol. 2012;293:269–309. doi: 10.1016/B978-0-12-394304-0.00013-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cullen SP, Henry CM, Kearney CJ, Logue SE, Feoktistova M, Tynan GA, Lavelle EC, Leverkus M, Martin SJ. Fas/CD95-induced chemokines can serve as “find-me” signals for apoptotic cells. Mol Cell. 2013;49:1034–1048. doi: 10.1016/j.molcel.2013.01.025. [DOI] [PubMed] [Google Scholar]
- 12.Fadok VA, Bratton DL, Konowal A, Freed PW, Westcott JY, Henson PM. Macrophages that have ingested apoptotic cells in vitro inhibit proinflammatory cytokine production through autocrine/paracrine mechanisms involving TGF-beta, PGE2, and PAF. J Clin Invest. 1998;101:890–898. doi: 10.1172/JCI1112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kim S, Elkon KB, Ma X. Transcriptional suppression of interleukin-12 gene expression following phagocytosis of apoptotic cells. Immunity. 2004;21 :643–653. doi: 10.1016/j.immuni.2004.09.009. [DOI] [PubMed] [Google Scholar]
- 14.Mukundan L, Odegaard JI, Morel CR, Heredia JE, Mwangi JW, Ricardo-Gonzalez RR, Goh YP, Eagle AR, Dunn SE, Awakuni JU, et al. PPAR-delta senses and orchestrates clearance of apoptotic cells to promote tolerance. Nat Med. 2009;15:1266–1272. doi: 10.1038/nm.2048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Xiao YQ, Freire-de-Lima CG, Schiemann WP, Bratton DL, Vandivier RW, Henson PM. Transcriptional and translational regulation of TGF-beta production in response to apoptotic cells. J Immunol. 2008;181:3575–3585. doi: 10.4049/jimmunol.181.5.3575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Vandivier RW, Henson PM, Douglas IS. Burying the dead: the impact of failed apoptotic cell removal (efferocytosis) on chronic inflammatory lung disease. Chest. 2006;129:1673–1682. doi: 10.1378/chest.129.6.1673. [DOI] [PubMed] [Google Scholar]
- 17.Tabas I. Macrophage death and defective inflammation resolution in atherosclerosis. Nat Rev Immunol. 2010;10:36–46. doi: 10.1038/nri2675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18**.Baxt LA, Garza-Mayers AC, Goldberg MB. Bacterial subversion of host innate immune pathways. Science. 2013;340:697–701. doi: 10.1126/science.1235771. This excellent review highlights the many ways intracellular pathogens in particular seek to control their hosts. [DOI] [PubMed] [Google Scholar]
- 19.Dockrell DH, Lee M, Lynch DH, Read RC. Immune-mediated phagocytosis and killing of Streptococcus pneumoniae are associated with direct and bystander macrophage apoptosis. J Infect Dis. 2001;184:713–722. doi: 10.1086/323084. [DOI] [PubMed] [Google Scholar]
- 20*.Ashida H, Mimuro H, Ogawa M, Kobayashi T, Sanada T, Kim M, Sasakawa C. Cell death and infection: a double-edged sword for host and pathogen survival. J Cell Biol. 2011;195:931–942. doi: 10.1083/jcb.201108081. This review highlights the many ways pathogens induce death of host cells via apoptosis, necrosis or pyroptosis. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Barry M, Bleackley RC. Cytotoxic T lymphocytes: all roads lead to death. Nat Rev Immunol. 2002;2:401–409. doi: 10.1038/nri819. [DOI] [PubMed] [Google Scholar]
- 22.Peters NC, Egen JG, Secundino N, Debrabant A, Kimblin N, Kamhawi S, Lawyer P, Fay MP, Germain RN, Sacks D. In vivo imaging reveals an essential role for neutrophils in leishmaniasis transmitted by sand flies. Science. 2008;321:970–974. doi: 10.1126/science.1159194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Laskay T, van Zandbergen G, Solbach W. Neutrophil granulocytes--Trojan horses for Leishmania major and other intracellular microbes? Trends Microbiol. 2003;11:210–214. doi: 10.1016/s0966-842x(03)00075-1. [DOI] [PubMed] [Google Scholar]
- 24.Peters NC, Sacks DL. The impact of vector-mediated neutrophil recruitment on cutaneous leishmaniasis. Cell Microbiol. 2009;11:1290–1296. doi: 10.1111/j.1462-5822.2009.01348.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.van Zandbergen G, Klinger M, Mueller A, Dannenberg S, Gebert A, Solbach W, Laskay T. Cutting edge: neutrophil granulocyte serves as a vector for Leishmania entry into macrophages. J Immunol. 2004;173:6521–6525. doi: 10.4049/jimmunol.173.11.6521. [DOI] [PubMed] [Google Scholar]
- 26.Ribeiro-Gomes FL, Peters NC, Debrabant A, Sacks DL. Efficient capture of infected neutrophils by dendritic cells in the skin inhibits the early anti-leishmania response. PLoS Pathog. 2012;8:e1002536. doi: 10.1371/journal.ppat.1002536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Behar SM, Divangahi M, Remold HG. Evasion of innate immunity by Mycobacterium tuberculosis: is death an exit strategy? Nat Rev Microbiol. 2010;8:668–674. doi: 10.1038/nrmicro2387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Oddo M, Renno T, Attinger A, Bakker T, MacDonald HR, Meylan PR. Fas ligand-induced apoptosis of infected human macrophages reduces the viability of intracellular Mycobacterium tuberculosis. J Immunol. 1998;160:5448–5454. [PubMed] [Google Scholar]
- 29.Molloy A, Laochumroonvorapong P, Kaplan G. Apoptosis, but not necrosis, of infected monocytes is coupled with killing of intracellular bacillus Calmette-Guerin. J Exp Med. 1994;180:1499–1509. doi: 10.1084/jem.180.4.1499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30**.Martin CJ, Booty MG, Rosebrock TR, Nunes-Alves C, Desjardins DM, Keren I, Fortune SM, Remold HG, Behar SM. Efferocytosis is an innate antibacterial mechanism. Cell Host Microbe. 2012;12:289–300. doi: 10.1016/j.chom.2012.06.010. This paper shows that efferocytosis is what contributes to Mtb killing following apoptosis** [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Davis JM, Clay H, Lewis JL, Ghori N, Herbomel P, Ramakrishnan L. Real-time visualization of mycobacterium-macrophage interactions leading to initiation of granuloma formation in zebrafish embryos. Immunity. 2002;17 :693–702. doi: 10.1016/s1074-7613(02)00475-2. [DOI] [PubMed] [Google Scholar]
- 32**.Yang CT, Cambier CJ, Davis JM, Hall CJ, Crosier PS, Ramakrishnan L. Neutrophils exert protection in the early tuberculous granuloma by oxidative killing of mycobacteria phagocytosed from infected macrophages. Cell Host Microbe. 2012;12:301–312. doi: 10.1016/j.chom.2012.07.009. This paper demonstrates that neutrophils also exert anti-mycobacterial effect** [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gonzalez-Cortes C, Reyes-Ruvalcaba D, Diez-Tascon C, Rivero-Lezcano OM. Apoptosis and oxidative burst in neutrophils infected with Mycobacterium spp. Immunol Lett. 2009;126:16–21. doi: 10.1016/j.imlet.2009.07.006. [DOI] [PubMed] [Google Scholar]
- 34.Lee HN, Surh YJ. Resolvin D1-mediated NOX2 inactivation rescues macrophages undertaking efferocytosis from oxidative stress-induced apoptosis. Biochem Pharmacol. 2013;86:759–769. doi: 10.1016/j.bcp.2013.07.002. [DOI] [PubMed] [Google Scholar]
- 35.Yvan-Charvet L, Pagler TA, Seimon TA, Thorp E, Welch CL, Witztum JL, Tabas I, Tall AR. ABCA1 and ABCG1 protect against oxidative stress-induced macrophage apoptosis during efferocytosis. Circ Res. 2010;106:1861–1869. doi: 10.1161/CIRCRESAHA.110.217281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kosmider B, Messier EM, Janssen WJ, Nahreini P, Wang J, Hartshorn KL, Mason RJ. Nrf2 protects human alveolar epithelial cells against injury induced by influenza A virus. Respir Res. 2012;13:43. doi: 10.1186/1465-9921-13-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Fujimoto I, Pan J, Takizawa T, Nakanishi Y. Virus clearance through apoptosis-dependent phagocytosis of influenza A virus-infected cells by macrophages. J Virol. 2000;74:3399–3403. doi: 10.1128/jvi.74.7.3399-3403.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Vergne I, Fratti RA, Hill PJ, Chua J, Belisle J, Deretic V. Mycobacterium tuberculosis phagosome maturation arrest: mycobacterial phosphatidylinositol analog phosphatidylinositol mannoside stimulates early endosomal fusion. Mol Biol Cell. 2004;15:751–760. doi: 10.1091/mbc.E03-05-0307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Vergne I, Chua J, Lee HH, Lucas M, Belisle J, Deretic V. Mechanism of phagolysosome biogenesis block by viable Mycobacterium tuberculosis. Proc Natl Acad Sci U S A. 2005;102:4033–4038. doi: 10.1073/pnas.0409716102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hubber A, Roy CR. Modulation of host cell function by Legionella pneumophila type IV effectors. Annu Rev Cell Dev Biol. 2010;26:261–283. doi: 10.1146/annurev-cellbio-100109-104034. [DOI] [PubMed] [Google Scholar]
- 41.Steele-Mortimer O. The Salmonella-containing vacuole: moving with the times. Curr Opin Microbiol. 2008;11:38–45. doi: 10.1016/j.mib.2008.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Barry AO, Mege JL, Ghigo E. Hijacked phagosomes and leukocyte activation: an intimate relationship. J Leukoc Biol. 2011;89:373–382. doi: 10.1189/jlb.0510270. [DOI] [PubMed] [Google Scholar]
- 43.Kinchen JM, Doukoumetzidis K, Almendinger J, Stergiou L, Tosello-Trampont A, Sifri CD, Hengartner MO, Ravichandran KS. A pathway for phagosome maturation during engulfment of apoptotic cells. Nat Cell Biol. 2008;10:556–566. doi: 10.1038/ncb1718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lewis DH, Peters W. The resistance of intracellular Leishmania parasites to digestion by lysosomal enzymes. Ann Trop Med Parasitol. 1977;71:295–312. doi: 10.1080/00034983.1977.11687192. [DOI] [PubMed] [Google Scholar]
- 45.Spath GF, Garraway LA, Turco SJ, Beverley SM. The role(s) of lipophosphoglycan (LPG) in the establishment of Leishmania major infections in mammalian hosts. Proc Natl Acad Sci U S A. 2003;100:9536–9541. doi: 10.1073/pnas.1530604100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46*.Moradin N, Descoteaux A. Leishmania promastigotes: building a safe niche within macrophages. Front Cell Infect Microbiol. 2012;2:121. doi: 10.3389/fcimb.2012.00121. This is a good review of Leshmaina virulence mechanisms* [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47**.Li W, Zou W, Yang Y, Chai Y, Chen B, Cheng S, Tian D, Wang X, Vale RD, Ou G. Autophagy genes function sequentially to promote apoptotic cell corpse degradation in the engulfing cell. J Cell Biol. 2012;197:27–35. doi: 10.1083/jcb.201111053. Authors show that autophagy is involved following efferocytosis* [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48*.Choy A, Roy CR. Autophagy and bacterial infection: an evolving arms race. Trends Microbiol. 2013;21:451–456. doi: 10.1016/j.tim.2013.06.009. This is an excellent review of autophagy and microbial immunity** [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Martinez J, Almendinger J, Oberst A, Ness R, Dillon CP, Fitzgerald P, Hengartner MO, Green DR. Microtubule-associated protein 1 light chain 3 alpha (LC3)-associated phagocytosis is required for the efficient clearance of dead cells. Proc Natl Acad Sci U S A. 2011;108:17396–17401. doi: 10.1073/pnas.1113421108. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 50.Green DR, Ferguson T, Zitvogel L, Kroemer G. Immunogenic and tolerogenic cell death. Nat Rev Immunol. 2009;9:353–363. doi: 10.1038/nri2545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Blander JM, Medzhitov R. Toll-dependent selection of microbial antigens for presentation by dendritic cells. Nature. 2006;440:808–812. doi: 10.1038/nature04596. [DOI] [PubMed] [Google Scholar]
- 52.Torchinsky MB, Garaude J, Martin AP, Blander JM. Innate immune recognition of infected apoptotic cells directs T(H)17 cell differentiation. Nature. 2009;458:78–82. doi: 10.1038/nature07781. [DOI] [PubMed] [Google Scholar]