

Abstract
Background
Tinnitus is an auditory sensation frequently following hearing loss. After cochlear injury, deafferented neurons become sensitive to neighbouring intact edge-frequencies, guiding an enhanced central representation of these frequencies. As psychoacoustical data [1-3] indicate enhanced frequency discrimination ability for edge-frequencies that may be related to a reorganization within the auditory cortex, the aim of the present study was twofold: 1) to search for abnormal auditory mismatch responses in tinnitus sufferers and 2) relate these to subjective indicators of tinnitus.
Results
Using EEG-mismatch negativity, we demonstrate abnormalities (N = 15) in tinnitus sufferers that are specific to frequencies located at the audiometrically normal lesion-edge as compared to normal hearing controls (N = 15). Groups also differed with respect to the cortical locations of mismatch responsiveness. Sources in the 90–135 ms latency window were generated in more anterior brain regions in the tinnitus group. Both measures of abnormality correlated with emotional-cognitive distress related to tinnitus (r ~ .76). While these two physiological variables were uncorrelated in the control group, they were correlated in the tinnitus group (r = .72). Concerning relationships with parameters of hearing loss (depth and slope), slope turned out to be an important variable. Generally, the steeper the hearing loss is the less distress related to tinnitus was reported. The associations between slope and the relevant neurophysiological variables are in agreement with this finding.
Conclusions
The present study is the first to show near-to-complete separation of tinnitus sufferers from a normal hearing control group based on neurophysiological variables. The finding of lesion-edge specific effects and associations with slope of hearing loss corroborates the assumption that hearing loss is the basis for tinnitus development. It is likely that some central reorganization follow a damage to hearing receptors, even though the paradoxical results indicate that they most likely are somewhat different than originally assumed (see Background). One partial explanation might lie in the involvement of top-down (presumably frontal-lobe) controlled processes. A better comprehension of the exact mechanisms leading to the present results could have a broad impact on the understanding and perhaps treatment of tinnitus.
Background
Tinnitus is defined as perception of sound in absence of an identifiable physical source. This condition, which can negatively affect normal everyday functioning, is diagnosed solely on an individual's report. Approximately 10 % of the population experience this condition, 10 % of whom claim to have moderate to severe distress caused by the tinnitus, such as sleeping problems, concentration difficulties, enhanced anxiety or depression [4]. For the European Union, this figure leads to 5 million people. Tinnitus frequently follows malfunction of hearing, e.g. presbyacusis, acute hearing loss, or acoustic trauma [5]. Contemporary views of tinnitus emphasize the role of the central auditory system [5,6]. Perception of phantom sound has been attributed to enhanced correlated spontaneous neural activity, interpreted as sound by the auditory cortex [6,7].
Several animal studies could demonstrate that circumscribed damage to hearing receptors alters the receptive field of the deafferented neurons. The characteristic frequency of these neurons moves toward intact lesion-edge frequencies [8,9], so that the edge-frequencies are overrepresented. In humans, Mühlnickel et al. [10] observed a shift of the cortical representation of the matched tinnitus frequency into an area adjacent to the expected tonotopic location. However in this study, subjects with an audiometrically detectable hearing loss were excluded from the study. Investigating subjects with high frequency hearing-loss, Dietrich et al. [11] were able to demonstrate an enhanced source activity for lesion-edge frequencies as compared to control frequencies well in the normal hearing range.
However, the authors could not report any significant correlations between the dipole moment and measures of tinnitus distress. It has to be emphasized though that tinnitus was not the focus of the study. Nevertheless, the general theoretical approach, assuming alteration of functional organization in the central auditory system, still remains appealing, suggesting that tinnitus may be the auditory analogue of somatosensory phantom pain: cortical reorganization following amputation has been shown to be correlated with the amount of reorganization in primary somatosensory cortex [12]. Irrespective of how useful this metaphor is, the briefly outlined animal research [7-9] suggests that generation of tinnitus is related to neuroplastic processes triggered by receptor damage.
The auditory system incorporates several efferent pathways, allowing opportunities for top-down processing. There is evidence that corticofugal pathways can alter subcortical best frequencies [13], and even hair cell activity [14,15]. Studying the negative difference wave (Nd), Jacobson et al. [16] found an enhancement of this component in tinnitus patients, suggesting increased attentional resources drawn to processing of auditory information. A few neuroimaging studies exist, in which tinnitus patients were able to modulate their tinnitus intensity via facial movements. From these studies it follows that among others primary [5,17], secondary auditory cortical areas [17-19] and limbic system structures, including the frontal cortex [17,19,20] are part of a tinnitus related neural network. In this study, we report a strong correlation of lesion-edge effects with the emotional distress caused by tinnitus, pointing to the potential role of top-down modulation of auditory cortex functioning.
On a behavioral (psychoacoustic) level a damage to inner-ear hearing receptors is known to go along with abnormally finer frequency-discrimination threshold at lesion-edge (LE) frequencies in high-frequency hearing-loss subjects [1-3]. The authors of these studies interpreted their results that LE-frequencies have an enlarged representation in the auditory cortex. Based on these findings from neuroscience and psychoacoustics an auditory mismatch design seemed appropriate to investigate whether processing of LE frequencies is abnormal. The auditory mismatch response refers to an evoked potential that is elicited when slightly deviant stimuli are interspersed in a series of standard tones. Here the standard tone (85%) in one condition was chosen to be a frequency at the audiometrically normal edge of the hearing-loss slope. In a control condition the standard was set to be an octave below the LE, i.e. well in the normal hearing range. Besides the standards three deviant frequencies – 1, 2 and 4% lower than the respective standard frequency – were presented each with a 5% probability of appearance. A focus of the data analysis was to investigate the source related mismatch response which was defined as the source activity in response to the standard subtracted from the one to a deviant. Statistical analysis was performed in a latency range 90–225 ms post stimulus onset reflecting the approximate time window for the mismatch response [21,22]. In order to capture possible dynamic processes (e.g., around the N1), the observation time was splitted into three equally long time windows (90–135 ms, 135–180 ms, 180–225 ms; see Methods for further details).
Assuming that edge-frequencies have enlarged representational zones [11], we tested if this would lead to enlarged mismatch responses when tones were in this frequency range. Another goal consisted in studying if different neuronal generators are involved in the processing of the auditory stimuli as compared to normal hearing controls. Effects seen on a group comparison level were related with tinnitus related distress which was assed using a standard German questionnaire (Tinnitus Fragebogen; [23]). Furthermore, as hearing loss is assumed to be fundamental for the development of tinnitus, questionnaire data and relevant neurophysiological variables were correlated with parameters of hearing loss (depth and steepness / slope).
Results
The 4 % deviant produces the largest amplitude differences relative to the standard (F2,56 = 22.46, p < .001). For the LE condition, Figure 1 depicts a striking abnormality in the tinnitus group that appears in the 90–135 ms time-window. Whereas the control group shows a marked deviance-dependency (1% < 2% < 4%), this is not the case for the tinnitus group. The amplitude following the 1% deviant is enhanced, while the amplitude of the 2% deviant appears to be reduced. For the LE condition, but not for the control frequency, an interaction between Group and Deviant (F2,56 = 6.73, p < .003) was found. Post-hoc analyses of the within factor Deviance using Tukey-Kramer HSD show a significant difference in the tinnitus group between 2% and 4% (mean difference: -.17, critical difference: .12); statistical significance were attained in the control group for the difference between 1% and 4% (mean difference: -.34, critical difference: .17), 2% and 4% (mean difference: -.19). An unpaired t-test yields a significant difference between tinnitus and controls for the 1% deviant (mean difference: .14; t28 = 2.71, p < .02) and 2% deviant (mean difference: -.11; t28 = -2.27, p < .04), but not for the 4% deviant (mean difference: -.13; t28 = -1.54, p > .1).
Figure 1.
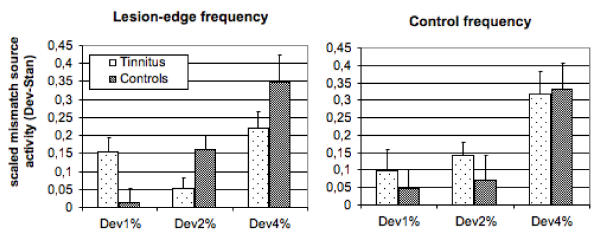
Mismatch related source activity. Responses to standards were subtracted from those to deviants whereby positive values indicate a greater neuronal response following the deviant. Normalized amplitudes (source strength) of the mismatch negativity are displayed for the time segment from 90–135 ms.
Source localization of the N1 showed a significant differences between groups in the inferior-superior (tinnitus: Mean = 65.17 mm / Standard Error = 2.92, controls: 52.55 / 4.85; F1,28 = 4.97, p < .04) and posterior-anterior direction (tinnitus: 4.38 / 3.89, controls: -5.51 / 1.69; F1,28 = 5.44, p < .03). Groups did not differ in the medial lateral direction (tinnitus: 43.88/ 2.08, controls: 45.17 / 1.58; F1,28 = 0.23).
Taken together, differences were found for the center of cortical responsivity around 100 ms and for the 1% and 2% mismatch response in the 90–135 ms period of the LE condition. These variables were used to assess associations with tinnitus related psychological distress (measured using a standard German questionnaire, [23]) by calculating simple linear regressions. As one subject (mti011) scored zero points on this subscale, thus representing an extreme, we excluded this case for the main calculation with neurophysiological variables (results are however also reported when this case is included). We found that source localizations in the posterior-anterior direction and the mismatch related source strength to the 2% deviant are strongly correlated with distress variables, i.e.: higher distress scores are associated with more anterior sources (Figure 2a; r = .76, F1,13 = 16.60, p < .002; with mti011: r = .53, F1,14 = 5.01, p < .04) and stronger mismatch activation for the 2% deviant (r = .74, F1,13 = 14.33, p < .003; with mti011: r = .64, F1,14 = 9.72, p < .009). The association between the distress scores and source localitzation in inferior-superior direction were not comparable in magnitude (range: .37 <r < .47) and also failed to reach statistical significance.
Figure 2.
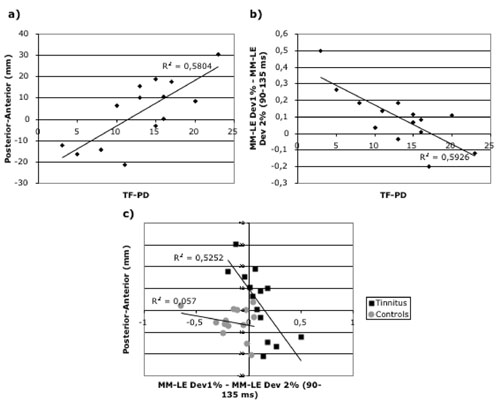
Scatterplot of neurophysiological variables. Associations between Psychological Distress (i.e., Emotional + Cognitive Distress; measured with the Tinnitus Questionnaire) and a) source localization in the anterior-posterior direction and b) the difference between the mismatch response of the LE condition to the 1% and 2% deviant in the early time window (Difference Index; negative values indicating a more normal mismatch pattern, i.e. higher responses for the 2%-Deviant). c) The correlation between the two neurophysiological variables. Note that they are highly correlated for tinnitus but not for control subjects, and that both groups are almost perfectly separated by these two variables. Concerning all scatterplots: straigth line corresponds to fit without mti011 (circle in a) and b), square in c)), broken line to the fit including this subject (see text for details).
Since an enhanced activation for the 1% deviant in the LE condition and a reduced activity for the 2% deviant seemed to be at the core of the abnormal response in tinnitus, we tested whether the difference between mismatch-related source activity for the 1% and 2% deviant would also show significant relationships with distress variables. This was indeed the case, i.e. the more negative this index (and therefore, the more "normal") the stronger the distress (see Figure 2b; r = -.77, F1,13 = 17.43, p < .002; with mti011: r = -.72, F1,14 = 13.68, p < .003). In Figure 2c it can be seen that the mismatch and source localization related variables are well correlated for the tinnitus group (r = .72, F1,13 = 13.24, p < .004; with mti011: r = -.69, F1,14 = 11.09, p < .006) but not for the control group (r = .23, n.s.). In the tinnitus group, the more normal the mismatch in the early time window of the LE condition (i.e., the more negative the difference index), the more anterior the focus of neuronal activity.
We also examined the possibility that the anterior shift of source localization and its considerable variability may be caused by an enhanced frontal activation, relative to the expected temporal activation. This was suggested by the difference in the source distribution between controls and tinnitus subjects using Minimum Norm solution (see Methods), as shown in Figure 3 for the 112 ms latency (maximum difference; upper panel). The results (see Figure 3 lower panel) suggest that a linear relationship exists between the two variables for the majority of cases. However, there is a subset of four patients with the most posterior source localizations. These patients exhibit strong frontal activation, thus causing the overall association to be quadratic. This relationship is statistically significant for the right hemisphere only (F2,14 = 6.32, p < 02). Interestingly, these four patients are exactly the ones who reported the lowest distress values (see Figure 2a).
Figure 3.
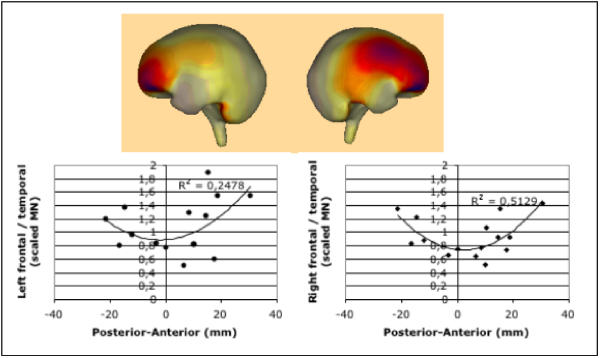
Upper panel: The minimum-norm solution for the difference between the Grand Means of the tinnitus and the control groups indicates a greater prefrontal activity (dark shading) in tinnitus patients following auditory stimulation. The darkest shading corresponds to 100% of the maximum activation (the ligther, the stronger the decrease relative to the maximum). This exploratory analysis was the basis for the post-hoc notion that the source localization in the anterior-posterior direction may be associated with prefrontal activity. Lower Panel: Correlating relative frontal activity (as compared to temporal sources) of each individual with the source location of the regional source results in a quadratic trend that is more pronounced for the right hemisphere.
Finally, concerning the role of the hearing loss parameters (depth and steepness), steepness appears to be relevant. This audiometric variable is significantly correlated with tinntitus related distress (see Figure 4c; r = -.64, F1,13 = 8.64, p < .02; with mti011: r = -.44, F1,14 = 3.24, p < .09). The direction of the correlation implies that steeper hearing losses are related to lower distress values. Furthermore steepness showed a significant association with the difference index of the mismatch at LE (see Figure 4b; r = .68, F1,13 = 10.49, p < .007; with mti011: r = .64, F1,14 = 9.18, p < .01), while being uncorrelated (r = .19) with the analog difference index for CO. Although the correlation with source localization in the posterior-anterior direction was also significant (see Figure 4a; r = -.54, F1,13 = 4.97, p < .05; with mti011: r = -.56, F1,14 = 5.80, p < .03), this effect seemed to be driven by the four subjects with the steepest hearing losses. All associations with depth of hearing loss were not significant (all Fs < 2). Steepness and depth of hearing loss were uncorrelated (r = -.16, F1,14 = 0.31, p < .59).
Figure 4.
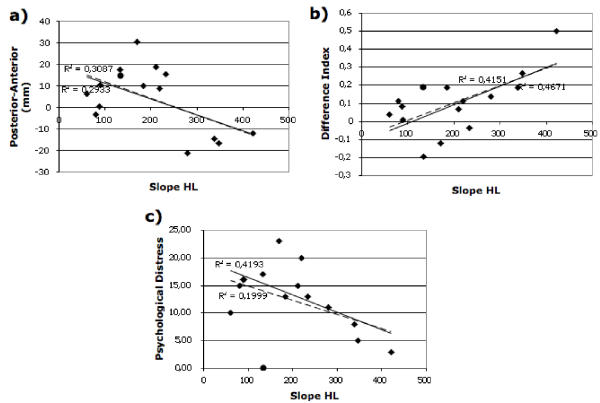
Scatterplot of steepness of hearing loss (larger values corresponding to steeper slopes; see Methods for details). a) The correlation of slope and source localization in posterior-anterior direction. Though being statisticilly the association is not very convincing and seems to be largely driven by four subjects with the steepest hearing losses. b) Association of steepness of hearing loss with the difference index of the mismatch (see also Figure 2b) shows a strong positive correlation. c) Hearing loss slope is negatively associated with tinnitus related distress. Concerning all scatterplots: straigth line corresponds to fit without mti011 (as circle), broken line to the fit including this subject (see text for details).
Discussion
Overall, the notion of a general enhancement of the auditory mismatch response at lesion edge frequencies could not be confirmed. This is not necessarily a contradiction to Dietrich et al. [11], as our study investigated the difference between two responses. The response pattern is generally more complex incorporating enhanced (1% deviant) and reduced (2% deviant) neuronal activity. It is noteworthy to emphasize that there was no difference to the control group in the control condition. The reason for this paradox result can not be derived from the present study. However it is difficult to argument with peripheral mechanisms, as the LE frequency was chosen by visual inspection from the audiogram: this measure does not offer detailed information about the type and extent of damage to receptors in the inner ear. It is striking that the time window of the effect is almost identical as the one reported by Jacobson et al. [16] on the enhanced negative difference wave (100–140 ms) in tinnitus, indicating an increased attention in this group. It is also conceivable for our study that (involuntary) attentional mechanisms were activated differently in the tinnitus group as compared to the control group; mechanisms, that might not be elicited in a classic N1 design as employed by Dietrich et al. [11]. Additionally, the relationship between the mismatch response and tinnitus related distress indicates that a more normal mismatch pattern is associated with enhanced distress scores. Subjects with the most abnormal mismatch patterns were those to exhibit lowest distress values. However, the more normal mismatch pattern in distressed subjects is not comparable with the pattern seen in normal hearing controls, as there is a considerable shift of the focus of neuronal activation in an anterior direction. Based on this finding, we hypothesize that a damage to hearing receptors of the inner-ear trigger reorganization processes leading to an abnormal mismatch pattern as found in the patients with low distress. With an increase of the distress level however, frontal brain regions involved in emotional and attentional regulation modulate this mismatch response. Prefrontal activity has been shown to be involved in processing auditory information previously[24], also for the auditory mismatch response [25,26]. Our data indicate a possible involvement of especially right prefrontal areas. Yet, caution has to be taken due to the low spatial resolution offered by EEG. A further aspect to consider is that a feature of the audiogram, namely slope of the hearing loss, is associated to both psychological distress and the mismatch difference index. Together with the specifity of the mismatch effects for the LE, this finding underlines the importance of seeing tinnitus in conjunction with a damage of the hearing system and not as two independent phenomena. Steepness of the hearing loss is positively associated with the abnormality of the mismatch response at LE as reflected in the difference index. Both variables are negatively related with tinntitus related distress, i.e. high scores are correlated with a more normal mismatch pattern and flatter hearing loss slopes. In summary, these results imply that a too simple reorganization logic (i.e., peripheral damage leading to expanded edge frequency representation and this resulting in tinnitus) that also underlied the present study is perhaps too naive, at least in the context of tinnitus. A better understanding of how auditory representational areas reorganize following a damage to receptors and how these processes act in concert with top-down mechanisms seems to be of big importance for a better understanding of this auditory phantom perception in general.
Conclusions
1. The major contribution of this study are the identfication of neurophysiological variables that are associated with the perceived distress level and to demonstrate a close-to-perfect separation of the tinnitus from the normal hearing control group on a physiological basis. So far diagnosis of tinnitus has solely relied on reports made by the subject. If the measures found here prove to be replicable, this could implicate first steps in the direction of an objectification of tinnitus and its related distress. They could serve e.g. as dependent variables in evaluation of therapies that currently lack objective markers. At this point however we have to emphasize the correlative nature of this finding. We do not want to imply that the relationships with neurophysiological variables resemble 'the' tinnitus itself.
2. Based on the odd response and association pattern with distress found in the tinnitus group for the LE condition, we state that tinnitus is overall a more complex phenomenon than a pure result of reorganization processes of neuronal response properties in the auditory cortex following damage to hearing receptors. This impression could arise when focussing on the animal 'tinnitus' literature that links reorganization of auditory representational areas with hearing loss. Our data suggests that these 'basic' neuroplastic processes (of which ones are relevant to tinnitus has still to be found out) have to be seen in conjunction with top-down processes in order to have the chance to grasp this partly disabling condition in humans. The fact that neurophysiological responses for LE frequencies are correlated with distress level in our study, however not in the Dietrich et al. one [11] raises the question for upcoming studies concerning the importance of the LE region.
3. The effects of steepness of hearing loss and the specifity of the mismatch effects for LE corroborate the notion that tinnitus has to be seen in a context with a underlying peripheral damage, rather than seeing these two phenomena as independent.
4. Even though the identification of neurophysiological correlates of tinnitus is a success in general, the exact mechanisms underlying our results remain unclear. An understanding of these mechanisms however may be a key for a better comprehension of tinnitus, potentially having implications for the treatment of this condition.
5. Our temporary conclusion concerning treatment is twofold: a) The fact, that our results can not be solely explained on the basis of activity from the auditory cortex alone but probably needs to take regions involved in emotional and attentional regulation into account, implies that therapeutic appraoches to tinnitus need to reduce potentially harming top-down effects (e.g., negative cognitions, fears) that direct the attention of the affected person to the phantom sound. These may include, elements already realized in therapeutic strategies such as Tinnitus Retraining Therapy [27] or Cognitive-Behavioral Therapy [28]. b) However, also effects closely related to the individual hearing-loss were observed. Thus, one of the aims in a therapy should also include the reversal of abnormal functioning of neurons in the auditory cortex following a damage. These might include improved hearing-aids and challenging hearing tasks in the affected frequency regions. Yet the problem here is that we have barely understood the neurscientificbasis of tinnitus.
Methods
Participants
The tinnitus and control group were matched by sex (13 males, 2 females) and age (tinnitus: mean age (range) = 47.27 (24–65); controls: 46.67 (24–63). All members of the tinnitus group experienced either unilateral or bilateral moderate to severe high-frequency hearing loss, and a tonal tinnitus. Four tinnitus sufferers would not recall the exact elicitors for their condition, nine of them reported to have had a long professional exposure to noise and two mentioned noise trauma as the origin of their disorder. No participant was aware of the hearing-loss (not uncommon in cases of high-frequency hearing loss) prior to the experience of tinnitus.
Stimuli and procedure
Prior to the experiment written informed consent was obtained and a tinnitus questionnaire [23] completed. For the mismatch experiment the patient was placed in a sound-attenuated chamber. All stimuli were sinusoidal tones of 70 ms duration (10 ms rise and fall). A mismatch procedure using one standard (85% probability) and three deviants (3 × 5% probability) was employed. Two conditions were defined based on visual inspection of a high-resolution audiogram of each tinnitus subject: In the Lesion-Edge (LE) condition, the standard was defined as the lowest frequency before hearing threshold started to deteriorate (i.e., audiometrically normal frequency prior to the downward slope in the audiogram). Stimuli were presented monaurally via headphones (HD520II, Sennheiser). Stimulation was presented to the ear in which the participant localized the tinnitus (ipsilateral tinnitus) or the one in which the tinnitus was perceived to be stronger (bilateral tinnitus with a dominant side). For the Control (CO) condition, the standard was set one octave below the LE frequency. Deviants were chosen to be 1, 2 and 4% lower in frequency than the standard in each condition. For each tinnitus patient, a control subject was matched by age and gender. The respective stimulus was presented to each patient's control subject. Three runs of each condition were alternated, with the starting condition balanced across subjects. Overall, 3701 stimuli were presented per condition with an ITI of 700 ms. During this time, participants were instructed to read a book of their choice. Prior to the actual experiment, the stimuli were matched in loudness to a 1000 Hz sinusoid tone set at 65 dB SPL.
Data acquisition and analysis
Neuroelectric signals were recorded (A/D rate: 500 Hz; filter DC-100 Hz; SynAmps amplifier, Neuroscan) from 65 locations on the scalp with Ag/AgCl electrodes mounted on an electrode cap (electrode impedances < 5 kΩ). At the end of the EEG-measurement electrode positions, relative to anatomical landmarks of the head, were digitized with a position indicator system (ISOTRAK II, Polhemus). Raw data were corrected for vertical (recorded above and below the right eye) and horizontal eye-movements (recorded at the outer canthi of both eyes), and for eyeblinks [29]. For source analysis BESA2000 (version 4.2.26, MEGIS) was used. A time-window of 30 ms around the peak in the global field power of each individual was selected and two symmetric (ipsilateral and contralateral to stimulation) regional sources were fitted to the data. Source localizations obtained from this analysis were used to obtain the source waveforms of each stimulus type (standard and deviants) for each condition (LE and CO) separately. Orientations of the dipoles were adjusted to the maximum value of the global field power (peak of N1 in all cases). To minimize the possibility of certain subjects extreme values and effects, source waveforms were standardized using the vector scaling method proposed by McCarthy and Wood [30]. The source-related mismatch activity was defined as the difference in activity between the respective deviant and its standard. To test whether the effect found on the posterior-anterior axis (localizations further anterior in tinnitus and high correlation with distress scores), were a cosequence of enhanced frontal activity the Minimum Norm Estimate (MNE; [31,32]) was employed (computations done with BESA2000). This method allows the analysis of distributed source configurations, by estimating the current strength of a multitude of dipoles (here: 162) located on a sphere. The basic computation consists in the multiplication of the pseudo-inverse of the lead-field matrix (i.e., the sensitivity of the sensors to the sources) with the obtained data. Prior to MNE, the grand average of each individual was projected onto a common standard electrode configuration. Following this, the overall grand average of the normal-hearing control group was subtracted from each tinnitus individual, i.e. the control group served as a kind of baseline. These data were then used for the MNE analysis, and the resulting source waveforms were analoguously scaled as described above. The results was subsequently averaged between 90–135 ms (i.e., the N1 time-window), thus resulting in a vector with 162 values (one per dipole) for each tinnitus individual. Since it was assumed that the source localizations observed using the symmetrical dipoles could have been the consequence of activity stemming mainly from frontal and temporal sources, the mean scaled activity values of clusters of dipoles in these regions were divided (i.e., frontal / temporal) for both hemispheres. This index, gives some information on how much more active frontal regions were as compared to temporal. Based on the notion that enhanced frontal activity would draw the localization of the fitted dipole further anterior, we expected a linear relationship.
Hearing loss parameters
Depth of hearing loss was defined as the absolute difference between the threshold (in dB HL) at LE to the steepest point of the hearing loss slope. Steepness of the hearing loss slope was defined as the regression-coefficient of following formula:
h~a*(log10(f)) + b,
where h is the hearing threshold, a is the slope (corresponding to steepness of hearing loss), f is the relevant frequency in the audiogram and b the intercept.
Author's contributions
NW designed the study, carried out the experiments, performed the data analysis and drafted the manuscript. SV assisted in collecting data. PB gave advice in the use of the data-analysis software (BESA). TE participated in the discussion and general conclusions.
Acknowledgments
Acknowledgments
This work was supported by a grant of the Deutsche Forschungsgemeinschaft (EL101/20). We thank Christina Robert for comments on early versions of the manuscript and H. Böttcher from ProAkustik for providing a high-precision audiometer.
Contributor Information
Nathan Weisz, Email: Nathan.Weisz@uni-konstanz.de.
Sandra Voss, Email: Sandra.Voss@switech.ch.
Patrick Berg, Email: Patrick.Berg@uni-konstanz.de.
Thomas Elbert, Email: Thomas.Elbert@uni-konstanz.de.
References
- McDermott HJ, Lech M, Kornblum MS, Irvine DR. Loudness perception and frequency discrimination in subjects with steeply sloping hearing loss: possible correlates of neural plasticity. J Acoust Soc Am. 1998;104:2314–2325. doi: 10.1121/1.423744. [DOI] [PubMed] [Google Scholar]
- Thai-Van H, Micheyl C, Norena A, Collet L. Local improvement in auditory frequency discrimination is associated with hearing-loss slope in subjects with cochlear damage. Brain. 2002;125:524–537. doi: 10.1093/brain/awf044. [DOI] [PubMed] [Google Scholar]
- Thai-Van H, Micheyl C, Moore BC, Collet L. Enhanced frequency discrimination near the hearing loss cut-off: a consequence of central auditory plasticity induced by cochlear damage? Brain. 2003;126:2235–2245. doi: 10.1093/brain/awg228. [DOI] [PubMed] [Google Scholar]
- Pilgramm M, Rychlik R, Lebisch H, Siedentop H, Goebel G, Kirchhoff D. Tinnitus in der Bundesrepublik Deutschland - eine repräsentative epidemiologische Studie. HNO aktuell. 1999;7:261–265. [Google Scholar]
- Lockwood AH, Salvi RJ, Burkard RF. Tinnitus. N Engl J Med. 2002;347:904–910. doi: 10.1056/NEJMra013395. [DOI] [PubMed] [Google Scholar]
- Eggermont JJ. Central tinnitus. Auris Nasus Larynx. 2003;30 Suppl:7–12. doi: 10.1016/S0385-8146(02)00122-0. [DOI] [PubMed] [Google Scholar]
- Norena AJ, Eggermont JJ. Changes in spontaneous neural activity immediately after an acoustic trauma: implications for neural correlates of tinnitus. Hear Res. 2003;183:137–153. doi: 10.1016/S0378-5955(03)00225-9. [DOI] [PubMed] [Google Scholar]
- Irvine DR, Rajan R, Brown M. Injury- and use-related plasticity in adult auditory cortex. Audiol Neurootol. 2001;6:192–195. doi: 10.1159/000046831. [DOI] [PubMed] [Google Scholar]
- Rajan R, Irvine DR. Neuronal responses across cortical field A1 in plasticity induced by peripheral auditory organ damage. Audiol Neurootol. 1998;3:123–144. doi: 10.1159/000013786. [DOI] [PubMed] [Google Scholar]
- Mühlnickel W, Elbert T, Taub E, Flor H. Reorganization of auditory cortex in tinnitus. Proc Natl Acad Sci U S A. 1998;95:10340–10343. doi: 10.1073/pnas.95.17.10340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dietrich V, Nieschalk M, Stoll W, Rajan R, Pantev C. Cortical reorganization in patients with high frequency cochlear hearing loss. Hear Res. 2001;158:95–101. doi: 10.1016/S0378-5955(01)00282-9. [DOI] [PubMed] [Google Scholar]
- Flor H, Elbert T, Knecht S, Wienbruch C, Pantev C, Birbaumer N, Larbig W, Taub E. Phantom-limb pain as a perceptual correlate of cortical reorganization following arm amputation. Nature. 1995;375:482–484. doi: 10.1038/375482a0. [DOI] [PubMed] [Google Scholar]
- Zhang Y, Suga N, Yan J. Corticofugal modulation of frequency processing in bat auditory system. Nature. 1997;387:900–903. doi: 10.1038/43180. [DOI] [PubMed] [Google Scholar]
- Xiao Z, Suga N. Modulation of cochlear hair cells by the auditory cortex in the mustached bat. Nat Neurosci. 2002;5:57–63. doi: 10.1038/nn786. [DOI] [PubMed] [Google Scholar]
- Maison S, Micheyl C, Collet L. Influence of focused auditory attention on cochlear activity in humans. Psychophysiology. 2001;38:35–40. doi: 10.1017/S0048577201990109. [DOI] [PubMed] [Google Scholar]
- Jacobson GP, Calder JA, Newman CW, Peterson EL, Wharton JA, Ahmad BK. Electrophysiological indices of selective auditory attention in subjects with and without tinnitus. Hear Res. 1996;97:66–74. doi: 10.1016/0378-5955(96)00055-X. [DOI] [PubMed] [Google Scholar]
- Mirz F, Gjedde A, Sodkilde-Jrgensen H, Pedersen CB. Functional brain imaging of tinnitus-like perception induced by aversive auditory stimuli. Neuroreport. 2000;11:633–637. doi: 10.1097/00001756-200002280-00039. [DOI] [PubMed] [Google Scholar]
- Giraud AL, Chery-Croze S, Fischer G, Fischer C, Vighetto A, Gregoire MC, Lavenne F, Collet L. A selective imaging of tinnitus. Neuroreport. 1999;10:1–5. doi: 10.1097/00001756-199901180-00001. [DOI] [PubMed] [Google Scholar]
- Mirz F, Pedersen B, Ishizu K, Johannsen P, Ovesen T, Stodkilde-Jorgensen H, Gjedde A. Positron emission tomography of cortical centers of tinnitus. Hear Res. 1999;134:133–144. doi: 10.1016/S0378-5955(99)00075-1. [DOI] [PubMed] [Google Scholar]
- Lockwood AH, Salvi RJ, Coad ML, Towsley ML, Wack DS, Murphy BW. The functional neuroanatomy of tinnitus: evidence for limbic system links and neural plasticity. Neurology. 1998;50:114–120. doi: 10.1212/wnl.50.1.114. [DOI] [PubMed] [Google Scholar]
- Picton TW, Alain C, Otten L, Ritter W, Achim A. Mismatch negativity: different water in the same river. Audiol Neurootol. 2000;5:111–139. doi: 10.1159/000013875. [DOI] [PubMed] [Google Scholar]
- Näätänen R, Alho K. Mismatch negativity--the measure for central sound representation accuracy. Audiol Neurootol. 1997;2:341–353. doi: 10.1159/000259255. [DOI] [PubMed] [Google Scholar]
- Goebel G, Hiller W. Tinnitus-Fragebogen (TF): Ein Instrument zur Erfassung von Belastung und Schweregrad bei Tinnitus. Göttingen, Hogrefe; 1998. [Google Scholar]
- Romanski LM, Goldman-Rakic PS. An auditory domain in primate prefrontal cortex. Nat Neurosci. 2002;5:15–16. doi: 10.1038/nn781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jemel B, Achenbach C, Muller BW, Ropcke B, Oades RD. Mismatch negativity results from bilateral asymmetric dipole sources in the frontal and temporal lobes. Brain Topogr. 2002;15:13–27. doi: 10.1023/A:1019944805499. [DOI] [PubMed] [Google Scholar]
- Alho K, Woods DL, Algazi A, Knight RT, Naatanen R. Lesions of frontal cortex diminish the auditory mismatch negativity. Electroencephalogr Clin Neurophysiol. 1994;91:353–362. doi: 10.1016/0013-4694(94)00173-1. [DOI] [PubMed] [Google Scholar]
- Jastreboff PJ, Jastreboff MM. Tinnitus Retraining Therapy (TRT) as a method for treatment of tinnitus and hyperacusis patients. J Am Acad Audiol. 2000;11:162–177. [PubMed] [Google Scholar]
- Andersson G. Psychological aspects of tinnitus and the application of cognitive-behavioral therapy. Clin Psychol Rev. 2002;22:977–990. doi: 10.1016/S0272-7358(01)00124-6. [DOI] [PubMed] [Google Scholar]
- Berg P, Scherg M. A multiple source approach to the correction of eye artifacts. Electroencephalogr Clin Neurophysiol. 1994;90:229–241. doi: 10.1016/0013-4694(94)90094-9. [DOI] [PubMed] [Google Scholar]
- McCarthy G Wood CC. Scalp distributions of event-related potentials: an ambiguity associated with analysis of variance models. Electroencephalogr Clin Neurophysiol. 1985;62:203–208. doi: 10.1016/0168-5597(85)90015-2. [DOI] [PubMed] [Google Scholar]
- Hämäläinen M, Hari R, Ilmoniemi RJ, Knuutila J, Lounasmaa OV. Magnetencephalography - theory, instrumentation, and applications to noninvasive studies of the working human brain. Reviews of Modern Physics. 1993;65:413–497. doi: 10.1103/RevModPhys.65.413. [DOI] [Google Scholar]
- Hauk O, Keil A, Elbert T, Müller MM. Comparison of data transformation procedures to enhance topographical accuracy in time-series analysis of the human EEG. J Neurosci Methods. 2002;113:111–122. doi: 10.1016/S0165-0270(01)00484-8. [DOI] [PubMed] [Google Scholar]