

Abstract
Purpose
To investigate the roles of an adrenomedullin receptor antagonist (adrenomedullin22–52) on high-glucose-induced human retinal endothelial cell (HREC) in vitro cell biology.
Methods
HRECs were cultured with different concentrations of glucose and adrenomedullin22–52. The proliferation of HRECs was evaluated by a cell counting kit-8 assay. Cell migration was assessed by scratch wound assay, and cell sprouting was detected by tube formation assay. The mRNA levels of adrenomedullin (ADM), vascular endothelial growth factor (VEGF), ADAMTS-1, and TSP-1 were measured by reverse-transcription polymerase chain reaction (RT–PCR). The VEGF and phosphatidylinositol 3′ kinase (PI3K) pathway protein expression levels were assessed by western blot analysis.
Results
Compared with 5 mM normal glucose treatment, 30 mM glucose significantly promoted the migration of HRECs, which was attenuated by 1 μg/ml adrenomedullin22–52. The proliferation of HRECs was also suppressed by 1 μg/ml adrenomedullin22–52. Furthermore, compared with other groups, 5 μg/ml of adrenomedullin22–52 was shown to suppress high-glucose-induced tube formation of HRECs. With adrenomedullin22–52 treatment, the mRNA level of ADAMTS-1 was significantly increased. Moreover, western blot and RT–PCR analyses showed that HRECs treated with 30 mM glucose exhibited increased VEGF and PI3K pathway protein levels, while the expression levels were suppressed by 5 μg/ml of adrenomedullin22–52.
Conclusions
Our study indicated that adrenomedullin22–52 mediated the migration, proliferation and tube formation after HRECs were exposed to high levels of glucose, which may be related to its ability to affect the expression of VEGF through the PI3K pathway.
Introduction
Diabetic retinopathy (DR) is one of the most common and severe micro-vascular complications of diabetes and can eventually lead to blindness. Early clinical signs of DR include micro aneurysms and hemorrhages, while the later signs are dilated, tortuous, irregular and narrow retinal blood vessels called retinal ischemia that ultimately result in neovascularization. At this stage, abnormal new blood vessels form at the back of the eye as a part of proliferative diabetic retinopathy (PDR) [1]. The incidence of DR is increasing every year, and it is one of the main causes of blindness. Therefore, it is necessary to develop effective measures for treating and preventing retinal neovascularization.
Adrenomedullin (ADM), a 52-amino-acid polypeptide, is a kind of expansion of blood vessels and diuretic active peptide originally isolated from human pheochromocytoma [2]. ADM belongs to a large family of peptides that includes calcitonin gene-related peptide (CGRP), calcionin (CT), and amylin [3]. It acts through the G protein-coupled receptor calcitonin receptor-like receptor (CRLR), with specificity for ADM being conferred by the receptor activity modifying proteins 2 (RAMP2) and 3 (RAMP3) [4].
ADM and its receptors are expressed in several tissues and organs, including the heart, lungs, kidney, blood plasma, spleen, endocrine glands, brain, and thymus [5]. Published studies have shown that vascular endothelial cells and vascular smooth muscle cells actively synthesize and secrete ADM [6,7]. Moreover, a recent study observed that each cell type in the retina could secrete ADM [8]. ADM has widely biologic functions in the vasculature, such as vasodilatation, regulation of vascular permeability, vascular protection, and promotion of angiogenesis [9]. ADM also may be involved in several ocular pathologies. The expression of ADM was increased in the vitreous humor of patients with diabetes [10,11], in patients with primary open-angle glaucoma [12], and in the aqueous humor of patients with uveitis and vitreoretinal disorders [13]; patients with retinitis pigmentosa have increased plasma ADM [14]. Adrenomedullin22–52 is a truncated peptide derived from ADM and a selective ADM receptor antagonist. Several independent studies have suggested that ADM could significantly promote the proliferation, migration and tube formation of human umbilical vein endothelial cells [15-17], while those functions could be suppressed by adrenomedullin22–52 [16,17]. A recent study reported that ADM increased the area of CNV in a laser-induced CNV model, which can be significantly blocked by both ADM antagonist and ADM antibody [18].
During the formation of new blood vessels, endothelial cells are stimulated to migrate, proliferate, and invade the surrounding tissues to form capillaries [15]. This process is closely associated with the activation of several signal pathways. Evidence suggests that ADM-induced angiogenesis is mediated by phosphatidylinositol 3′ kinase (PI3K/Akt), mitogen-activated protein kinase (MAPK), focal adhesion kinase (p125FAK), and extracellular signal-regulated kinase 1/2 (ERK1/2), while these effects are counteracted by the ADM receptor antagonist (adrenomedullin22–52) [19-21]. Additionally, there are several factors involved in the process of new blood vessel formation in the eyes. Vascular endothelial growth factor (VEGF) is considered to be a main stimulating factor among the various factors of pro-angiogenesis. VEGF is closely related to the ocular angiogenesis platelet protein that is an important regulatory factor in the process of blood vessel formation; it can promote vascular endothelial cell migration and proliferation, so it stimulates the generation of new blood vessels [22]. Several studies have found that the angiogenesis role of ADM is closely related with VEGF. Maki and colleagues [23] reported that ADM promoted the generation of new blood vessels via VEGF in a mice brain hypoperfusion model. In vitro, ADM can encourage the proliferation and tube formation of human umbilical vein endothelial cells and human saphenous vein endothelial cells by promoting the expression of VEGF [23,24].
Based on these studies, we speculated that ADM may be involved in the development of DR. However, the role of adrenomedullin22–52 on high-glucose-induced HREC in vitro cell biology has remained obscure. Therefore, we investigated the role of adrenomedullin22–52 on high- glucose-induced HREC proliferation, migration, and tube formation.
Methods
Reagents and antibodies
Human retinal endothelial cell lines (HRECs) were purchased from Ya Ji biologic technologies (Shanghai, China). Dulbecco’s modified Eagle medium (DMEM) was obtained from Thermo scientific (Hyclone, UT). Fetal Bovine Serum came from PAA laboratories (Cölbe, Germany). Trypsin-EDTA, D-glucose, and adrenomedullin22–52 were originated from Sigma-Aldrich (St Louis, MO). The cell counting kit-8 was purchased from Dojindo (Kumamoto, Japan). Matrigel was bought from Becton Dickinson Labware (Bedford, MA). The primers were synthesized by the Shanghai biologic engineering technology service (Shanghai, China). Traces of RNA extraction kit (RNeasy Mini Kit) and reverse transcription Kit (Ominiscript RT Kit) came from Germany Qiagen Company (Qiagen, Hilden, Germany). Rabbit anti VEGF antibody was shipped from Santa Cruz Biotechnology (Santa Cruz, CA). Anti-PI3K P110 alpha (Phosphatidylinositol 3 kinase catalyticalpha) was purchased from biolegend (San Diego, CA), and GAPDH antibody was obtained from the Beyotime Institute of Biotechnology (Shanghai, China).
Cell culture
HRECs were cultured in a humidified atmosphere of 5% CO2 and 95% air at 37 °C in DMEM medium supplemented with 10% fetal bovine serum. Cells were passaged 1:2 at conference by release with 0.25% Trypsin-EDTA and subcultured in culture flasks. The HRECs were split at 90% confluence, and the culture media were changed every 2–3 days.
Treatment of HRECs
HRECs were divided into several groups: 5 mM normal glucose, 30 mM glucose, and 30 mM glucose plus different concentrations of adrenomedullin22–52. The HRECs were split at 90% confluence and subcultured in 96-well plates or 6-well plates according to the appropriate assay conditions.
Cell migration assay
The effect of adrenomedullin22–52 on high-glucose-induced migration of HRECs was evaluated by scratch wound assay. In brief, the HRECs were starved with serum-free medium for 12 h when the HRECs had grown to 90% confluence in the 6-well plates. When the HRECs had grown to over-confluence, a wound of appropriate width was made with a 200 μl tip. Then, the HRECs were washed with sterile 1×PBS three times to remove the floating cells, incubated with serum-free media containing different concentrations of glucose with or without adrenomedullin22–52, and then cultured in the 6-well plate in at 37 °C in a 5% CO2 incubator. The migration monolayer was photographed at 0 h, 12 h, 24 h, and 48 h. Five fields were photographed for each well. The migration distance was measured by using Photoshop CS2. The experiment was performed at least three times with similar results.
Cell proliferation assay
CCK-8 assay was used to evaluate the high-glucose-induced proliferation of HRECs with different concentrations of adrenomedullin22–52. In brief, 3 ×103 of HRECs were seeded into each well of a 96–well plate and allowed to adhere for 24 h. After cells were adherent to the bottom of the plate, the cells were cultured in serum-free media for starvation for 24 h. Then, the cells were treated with different concentrations of glucose and adrenomedullin22–52 (10 ng/ml-1000 ng/ml) for 24 h, and the proliferative activity was determined by CCK-8 assay according to the manufacturer’s instructions. In brief, 10 μl CCK-8 were added to each well followed by incubation for an additional 2–4 h. When the media changed from red to yellow, the absorbance value at a wavelength of 450 nm was detected. The experiment was performed at least three times with similar results.
Tube formation assay
The morphogenesis assay on Matrigel was conducted to evaluate the effect of adrenomedullin22–52 on high-glucose induced HRECs tube formation. According to the manufacturer’s instructions, the Matrigel was placed in a refrigerator at 4 °C overnight to thaw. Once it had thawed, 60 μl Matrigel was added to a pre-cooled 96-well plate and the plate was immediately placed in a humidified CO2 incubator at 37 °C for 30 min to solidify the Matrigel. HRECs that had been cultured under different media with or without adrenomedullin22–52 were seeded immediately on the solidified Matrigel at a density of 1.5×104 cells per well. Then, the plates were placed in a humidified atmosphere of 5% CO2 and 95% air at 37 °C for 8 h to allow the formation of a capillary-like structure. The pictures were photographed, and the number of capillaries formed was qualitatively assessed using Image-Pro Plus 6.0 software. The experiment was repeated at least three times under the same conditions.
RNA isolation and cDNA synthesis
When the HRECs had grown to 90% confluence in the culture flasks, the cells were harvested, seeded onto 6-well culture plates, and cultured in a humidified atmosphere of 5% CO2 and 95% air at 37 °C in DMEM medium supplemented with 10% fetal bovine serum. When the HRECs had grown to over-confluence of the bottom, the cells were treated with different concentrations of adrenomedullin22–52 for 6–8 h. Cells were harvested, and total RNA was isolated using the RNeasy Mini Kit according to the manufacturer’s instructions. Reverse transcription of RNA was performed in a 20 μl final volume using a Reverse Transcription Kit (Ominiscript RT Kit) and a cDNA Synthesis Kit according to the manufacturer’s instructions. Finally, the cDNA was stored at −20 °C before its use in RT–PCR.
RT–PCR
RT–PCR was performed to detect the expression of mRNA for ADM, VEGF, ADAMTS-1, and TSP-1. These primers used are summarized in Table 1. A 2 μl aliquot of cDNA was amplified by PCR in a total volume of 25 μl containing 17.575 μl ddH2O, 2.5 μl 10 ×PCR buffer, 2 μl dNTP, 0.125 μl rTaq, and 2 μl of each primer. The PCR was performed under the following conditions: an initial denaturation step at 94 °C for 2 min followed by 35 or 27 cycles of standard PCR (30 s denaturation at 94 °C, 30 s annealing at 58 °C, 30 s extension at 72 °C), and re-extension at 72 °C for 10 min. Levels of VEGF, ADAMTS-1, TSP-1, and ADM mRNA were normalized to those of GAPDH mRNA.
Table 1. Specific set of primers and conditions of PCR.
| Gene | Primmer sequences (5′-3′) | Annealing temp(°C) | PCR cycles | PCR product (bp) |
|---|---|---|---|---|
|
VEGF |
F: TGGTCCCAGGCTGCACCCAT |
58 |
35 |
184 |
| R: CGCATCGCATCAGGGGCACA |
||||
|
ADAMTS-1 |
F: CTCCGCCTGCACGCCTTTGA |
58 |
35 |
171 |
| R: ATCGCCATTCACGGTGCCGG |
||||
|
ADM |
F: ATGAAGCTGGTTCCGTCG |
58 |
35 |
350 |
| R: ATCCTAAAGAA AGTGGGGAGCACTTC |
||||
|
TSP-1 |
F: GGTCGGCCTGCACTGTCACC |
58 |
35 |
179 |
| R: GGGGAAGCTGCTGCACTGGG |
In the table, “F” indicates forward primer and “R” indicates reverse primer.
Western blot analysis
HRECs were split at 90% confluence and cultured in a humidified atmosphere of 5% CO2 and 95% air at 37 °C with different concentrations of adrenomedullin22–52 for 24 h. Cells were harvested and lysed in 100 μl of lysis buffer with protease inhibitor cocktail for total protein extraction. The lysates were then centrifuged from the samples at 13,800 ×g at 4 °C for 10 min. The supernatant was removed, placed in a fresh tube, mixed with 6×SDS sample buffer and heated for 10 min at 100 °C. The protein samples were separated with 10% SDS–PAGE and transferred onto a nitrocellulose membrane. The membrane was blocked for 1 h in PBS containing 0.05% Tween-20 (PBST) and 5% nonfat dry milk powder at room temperature. The membrane was then incubated with the primary antibody at 4 °C overnight, washed three times with PBST containing 0.05% Tween-20 for 10 min, and incubated with horseradish peroxidase-conjugated secondary antibodies for 1 h at room temperature. The blot was visualized using chemiluminescence detection reagents (Cell Signing Technology, Danvers, MA), and the pictures were photographed using the ChemiDocTM MP Imaging System (BIO-RAD, Hercules, CA). The data were analyzed by Image J software. GAPDH was used as a loading control. The experiment was performed three times with similar results.
Statistical analysis
All data are expressed as mean ±SD. Data were analyzed statistically by one way ANOVA (ANOVA) or two-tailed Student t test with the Statistical Package for the Social Sciences (SPSS) 13.0 software. A p value <0.05 was accepted as statistically significant.
Results
Effect of high glucose on ADM mRNA expression in HRECs
Published results have shown that ADM is increased in the plasma of DR patients [25]. RT–PCR was performed to observe whether the high glucose level would affect the HRECs’ ADM secretion. As shown in Figure 1, when compared with 5 mM glucose conditions, ADM expression was upregulated in HRECs under 30 mM glucose conditions. Therefore, the high glucose condition appeared to upregulate the expression of ADM in HRECs.
Figure 1.
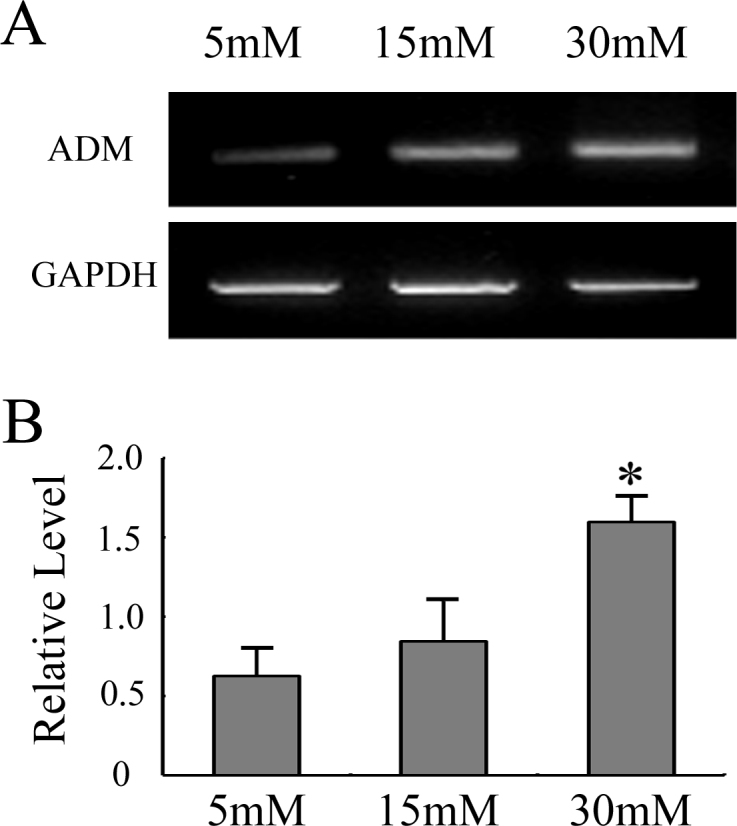
High glucose promoted the expression of ADM. After HRECs were treated with different concentrations of glucose (5 mM, 15 mM, and 30 mM) for 48 h, RT–PCR was performed to assess the mRNA expression of ADM. A: Representative results from three independent trials of RT–PCR. B: Ratios of ADM to GAPDH were determined by RT–PCR. The quantitative data of the ratio were from three independent experiments. All values represent Mean±SD (n=3). * p<0.05 versus the 5 mM group.
Adrenomedullin22–52 inhibited HREC proliferation induced by high glucose
To assess the effect of adrenomedullin22–52 on the proliferation of HRECs, in vitro CCK-8 assay was performed to measure the proliferation ability of HRECs at different concentrations of glucose and adrenomedullin22–52 (10 ng/ml-1000 ng/ml). The results shown in Figure 2 indicate that compared with the 5 mM glucose treatment, the proliferation ability of HRECs had no significant effect when treated with 30 mM glucose. The proliferation ability of HRECs was inhibited by the treatment with 30 mM glucose and 1000 ng/ml of adrenomedullin22–52 compared to 30 mM glucose.
Figure 2.

Adrenomedullin22–52 inhibited HREC proliferation induced by high glucose. After the HRECs were treated with high glucose or high glucose plus different concentrations of adrenomedullin22–52 for 24 h, cell proliferation was measured by CCK-8 assay. The absorbance value was detected using an enzyme-linked immunity instrument at a wavelength of 450 nm. All values represent Mean±SD (n=3). * p<0.05 versus the 30 mM group.
Adrenomedullin22–52 inhibited HREC migration induced by high glucose
To evaluate whether adrenomedullin22–52 had an effect on the migration of HRECs, we performed an in vitro scratch wound test to measure the migration ability of HRECs at different concentrations of glucose and adrenomedullin22–52. As shown in Figure 3, a significantly accelerated wound closure was observed in the group treated with 30 mM glucose compared with the 5 mM glucose treatment group. However, the wound area was still wide in the HRECs with high glucose co-treatment with 1 μg/ml adrenomedullin22–52 at 24 h. These results indicated that adrenomedullin22–52 could effectively inhibit the migration ability of HRECs cultured in 30 mM glucose.
Figure 3.

Adrenomedullin22–52 inhibited HREC migration induced by high glucose. HRECs were analyzed for the ability to migrate into a wound area using a scratch wound assay. A: After HRECs were wounded using a 200 μl yellow micropipette tip, they were incubated with serum-free media containing high glucose with or without adrenomedullin22–52 for 24 h. The re-endothelialized monolayer area was photographed. A magnification of × 200 was used to image the re-endothelialized monolayer area. B: The quantitative data of the migration distance were from three independent experiments. All values represent Mean±SD (n=3). * p<0.05 versus the 30 mM group. # p<0.05 versus the 5 mM group.
Effect of high glucose and adrenomedullin22–52 on capillary tube formation of HRECs
To explore the effect of high glucose and adrenomedullin22–52 on angiogenesis, we measured HREC tube formation by Matrigel assay. As shown in Figure 4, the number of capillary-like structures was significantly enhanced under the high glucose condition compared with the 5 mM glucose group. However, the treatment with 5 ug/ml of adrenomedullin22–52 caused significantly fewer capillary-like structures than those produced by high glucose. These results provide further proof that adrenomedullin22–52 can inhibit the ability of high glucose-induced capillary-like formation.
Figure 4.

Adrenomedullin22–52 inhibited high-glucose-induced tube formation. The tube formation was determined by Matrigel assay. A: After culture under appropriate glucose conditions for 24 h, HRECs (1.5×104 cells/well) were added to 96-well culture plates and incubated for 8 h to allow the formation of capillary-like structures. A magnification of × 200 was used to image the cells. B: The quantitative data of the capillary-like structures were obtained from three independent experiments. All values represent Mean±SD (n=3). * p<0.05 versus the 30 mM group. # p<0.05 versus the 5 mM group.
Effect of high glucose and adrenomedullin22–52 on VEGF, ADAMTS-1, and TSP-1 mRNA expression in HRECs
The effect of adrenomedullin22–52 on the mRNA expression of VEGF, ADAMTS-1, and TSP-1 in high-glucose-induced HRECs was determined next. As shown in Figure 5, the expression of VEGF was significantly increased in 30 mM glucose conditions compared with 5 mM glucose, while the level was significantly suppressed by 5 μg/ml adrenomedullin22–52. Following treatment with 5 μg/ml adrenomedullin22–52, the mRNA level of ADAMTS-1 was significantly increased.
Figure 5.

Angiogenic factors expression. After the HRECs were treated with high glucose or high glucose plus different concentrations of adrenomedullin22–52 for 8 h, RT–PCR analysis was performed to detect pro-angiogenic and anti-angiogenic factor gene expression in the HRECs. A: Representative results from three independent runs of RT–PCR. B: Ratios of target gene expression to GAPDH were determined by RT–PCR. The quantitative data of the ratios were from three independent experiments. All values represent Mean±SD (n=3). * p<0.05 versus the 30 mM group. # p<0.05 versus the 5 mM group.
Effect of high glucose and adrenomedullin22–52 on VEGF and PI3K pathway protein expression in HRECs
Consistent with our RT–PCR data, the western blot shown in Figure 6 indicated that compared with 5 mM glucose, the 30 mM glucose treatment markedly increased the VEGF protein expression, while this increase was significantly blocked by 5 μg/ml adrenomedullin22–52. This result suggested that 5 μg/ml adrenomedullin22–52 could inhibit VEGF secretion and significantly inhibited the levels of VEGF in the media of HRECs under the high glucose condition. The western blot results regarding the PI3K pathway shown in Figure 7 further supported the VEGF result.
Figure 6.

Adrenomedullin22–52 inhibited high-glucose-induced VEGF expression. After HRECs were treated with high glucose or high glucose plus different concentrations of adrenomedullin22–52 for 24 h, the protein level of VEGF was determined by western blot assay. A: Representative results from three independent trials of western blot. B: Ratios of VEGF to GAPDH were determined by western blot. The quantitative data of the ratios were from three independent experiments. All values represent Mean±SD (n=3). * p<0.05 versus the 30 mM group. # p<0.05 versus the 5 mM group.
Figure 7.

Adrenomedullin22–52 inhibited the activation of PI3K by high glucose. After HRECs were treated with high glucose or high glucose plus different concentrations of adrenomedullin22–52 for 24 h, the protein level of PI3K was determined by western blot assay. A: Representative results from three independent runs of western blot. B: Ratios of PI3K to GAPDH were determined by western blot. The quantitative data of the ratios were collected from three independent experiments. All values represent Mean±SD (n=3). * p<0.05 versus the 30 mM group. # p<0.05 versus the 5 mM group.
Discussion
High glucose is considered to be an important risk factor in the development of DR. It could activate the polyol pathway of glucose metabolism. It generates cellular stress, injury of vascular pericytes and endothelial cells, and the development of abnormal capillaries [26,27]. Endothelial cells play a crucial role in regulating vascular tone, and injury of these cells is the initial step that leads to irreversible structural abnormalities [28]. In our current study, HRECs were used to imitate the pathogenesis of DR under the condition of high glucose. Similar to previous reports, HRECs in the current study were cultured in either physiologic (5 mM) or high glucose (30 mM) media [28,29]. In line with other studies, our results suggested that high glucose could promote the migration and tube formation of HRECs. However, there have been conflicting reports of high glucose on the proliferation of HRECs. Yuan et al. found the growth of HRECs in 30 mM glucose after 48 h was significantly higher than in 5 mM glucose [28]. Premanad et al. observed that although HREC treated with 30 mM glucose for 72 h showed no significant increase in proliferation when compared with cells treated with 5 mM glucose, there was an increasing trend in the high-glucose condition [29]. In our study, compared to the normal glucose condition, treatment with high glucose for 24 h failed to accelerate the proliferation of HRECs. One reason for this discrepancy may be that endothelial cells used in our study are the cell line while in the previous study, they used the primary cultured endothelial cells from the eyes of donors. Another explanation could be the difference of the culture time in the 30 mM glucose condition.
ADM, a 52-amino acid polypeptide, is well known as an expander of blood vessels and a diuretic active peptide [2]. Accumulating evidence suggests that ADM is a pro-angiogenesis factor that promotes the proliferation, migration, and tube formation of endothelial cells [15-17,19]. Additionally, several lines of proof have suggested that inhibiting the ADM pathway with antibodies or antagonists directed against ADM or ADM receptors can reduce angiogenesis and tumor cell proliferation in mouse cancer models [30]. Zakareia and colleagues [25] reported that ADM was specifically enhanced in the plasma and vitreous humor of DR patients. Recently, it has been shown that ADM promoted the size of laser-induced CNV and ADM antagonist significantly inhibited the CNV [18]. Based on these studies, we assumed that ADM is involved in the process of ocular angiogenesis and adrenomedullin22–52 might be a useful target for ocular angiogenesis intervention.
The angiogenic cascade is a complex and multistep process. Vascular endothelial cell migration and proliferation make up the initial step in angiogenesis, and then endothelial cells differentiate into a capillary-like network [31]. This process is tightly regulated by a series of pro- and anti-angiogenic molecules in normal physiology; when this equilibrium is broken, serious consequences may arise [22]. VEGF is considered to be a major factor of the abnormal angiogenesis that occurs in response to high glucose. At present, most researchers believe that high glucose could stimulate VEGF secretion, which is similar to our findings. However, the relationship between ADM and VEGF is still unclear. Some researchers found adrenomedullin could enhance VEGF-induced EC capillary formation and Akt pathway activation [32]. Other studies have indicated that blocking antibodies to VEGF cannot significantly inhibit AM-induced human umbilical vein endothelial cell (HUVEC) capillary tube formation [15]. Maki and colleagues [23] reported adrenomedullin induced capillary tube formation, which can be suppressed by VEGF neutralization. Our results demonstrated 5 μg/ml adrenomedullin22–52 could inhibit high-glucose- induced HRECs VEGF expression. These seemingly contradictory findings may be due to the differences of cell types, culture media, treatment times, dose of agents, and so on.
As antiangiogenic molecules, TSP-1 and ADAMTS-1, which are also constitutively present in Ecs, can preclude vasculature development [33,34]. TSP-1 levels in Bruch’s membrane and choroidal vessels were decreased in age-related macular degeneration (AMD), and a reduction in TSP-1 may permit the formation of choroidal neovascularization [35]. TSP-1 is processed to release antiangiogenic polypeptides by the action of ADAMTS-1 [36]. At high concentration, ADAMTS-1 can also inhibit endothelial migration induced by the combined treatment with VEGF and basic fibroblast growth factor [37]. Macrophages are the source of TSP-1, ADAMTS-1, and VEGF [38-40]. It was published that adrenomedullin22–52 could suppress the proliferation of Ecs, which was potentiated by TNF-α-treated macrophage cell lines [18]. Thus, macrophages may be associated with the anti-angiogenic effect of adrenomedullin22–52.
Several signaling pathways are involved in the process of neovascularization. The PI3K/Akt signaling pathway can stimulate EC migration, proliferation, and tube formation [41,42], and PI3K is a downstream mediator of VEGF [43]. Maki and colleagues [23] found that ADM could upregulate the expression of VEGF via the PI3K pathway. Furthermore, pretreatment with 10−10 mol/l adrenomedullin22-52 slightly decreased the phosphorylation of Akt, and treatment with 10−6mol/l partially suppressed the phosphorylation of Akt [19]. We found that high glucose could stimulate the phosphorylation of PI3K, which can be suppressed by 5 μg/ml adrenomedullin22–52. These results indicate that adrenomedullin22–52 can inhibit the phosphorylation of PI3K.
In conclusion, all of our results suggest that adrenomedullin22–52 could inhibit HREC migration, proliferation, and tube formation induced by high glucose by decreasing the expression of VEGF and the phosphorylation of the PI3K pathway. Although our findings provide evidence that ADM may be a potential drug to control the incidence and progression of DR in the future, further in vivo studies are still needed to evaluate its safety and efficacy.
Acknowledgments
This work was supported by the National Natural Science Foundation in China (NSFC30771978, NSFC30972712), Jiangsu Province's Key Medical Talents Program (RC2011104) and the Soochow Scholar Project of Soochow University (to Peirong Lu).
References
- 1.Gao R, Zhu BH, Tang SB, Wang JF, Ren J. Scutellarein inhibits hypoxia- and moderately-high glucose-induced proliferation and VEGF expression in human retinal endothelial cells. Acta Pharmacol Sin. 2008;29:707–12. doi: 10.1111/j.1745-7254.2008.00797.x. [DOI] [PubMed] [Google Scholar]
- 2.Kitamura K, Kangawa K, Kawamoto M, Ichiki Y, Nakamura S, Matsuo H, Eto T. Adrenomedullin: a novel hypotensive peptide isolated from human pheochromocytoma. Biochem Biophys Res Commun. 2012;425:548–55. doi: 10.1016/j.bbrc.2012.08.022. [DOI] [PubMed] [Google Scholar]
- 3.Wimalawansa SJ. Amylin, calcitonin gene-related peptide, calcitonin, and adrenomedullin: a peptide superfamily. Crit Rev Neurobiol. 1997;11:167–239. doi: 10.1615/critrevneurobiol.v11.i2-3.40. [DOI] [PubMed] [Google Scholar]
- 4.McLatchie LM, Fraser NJ, Main MJ, Wise A, Brown J, Thompson N, Solari R, Lee MG, Foord SM. RAMPs regulate the transport and ligand specificity of the calcitonin-receptor-like receptor. Nature. 1998;393:333–9. doi: 10.1038/30666. [DOI] [PubMed] [Google Scholar]
- 5.Ribatti D, Nico B, Spinazzi R, Vacca A, Nussdorfer GG. The role of adrenomedullin in angiogenesis. Peptides. 2005;26:1670–5. doi: 10.1016/j.peptides.2005.02.017. [DOI] [PubMed] [Google Scholar]
- 6.Sugo S, Minamino N, Kangawa K, Miyamoto K, Kitamura K, Sakata J, Eto T, Matsuo H. Endothelial cells actively synthesize and secrete adrenomedullin. Biochem Biophys Res Commun. 1994;201:1160–6. doi: 10.1006/bbrc.1994.1827. [DOI] [PubMed] [Google Scholar]
- 7.Sugo S, Minamino N, Shoji H, Kangawa K, Kitamura K, Eto T, Matsuo H. Production and secretion of adrenomedullin from vascular smooth muscle cells: augmented production by tumor necrosis factor-alpha. Biochem Biophys Res Commun. 1994;203:719–26. doi: 10.1006/bbrc.1994.2241. [DOI] [PubMed] [Google Scholar]
- 8.Blom J, Giove TJ, Pong WW, Blute TA, Eldred WD. Evidence for a functional adrenomedullin signaling pathway in the mouse retina. Mol Vis. 2012;18:1339–53. [PMC free article] [PubMed] [Google Scholar]
- 9.Nagaya N, Mori H, Murakami S, Kangawa K, Kitamura S. Adrenomedullin: angiogenesis and gene therapy. Am J Physiol Regul Integr Comp Physiol. 2005;288:R1432–7. doi: 10.1152/ajpregu.00662.2004. [DOI] [PubMed] [Google Scholar]
- 10.Er H, Doğanay S, Ozerol E, Yürekli M. Adrenomedullin and leptin levels in diabetic retinopathy and retinal diseases. Ophthalmologica. 2005;219:107–11. doi: 10.1159/000083270. [DOI] [PubMed] [Google Scholar]
- 11.Shaw SG, Boden JP, Biecker E, Reichen J, Rothen B. Endothelin antagonism prevents diabetic retinopathy in NOD mice: a potential role of the angiogenic factor adrenomedullin. Exp Biol Med. 2006;231:1101–5. [PubMed] [Google Scholar]
- 12.Evereklioglu C, Doganay S, Er H, Yurekli M. Aqueous humor adrenomedullin levels differ in patients with different types of glaucoma. Jpn J Ophthalmol. 2002;46:203–8. doi: 10.1016/s0021-5155(01)00501-9. [DOI] [PubMed] [Google Scholar]
- 13.Udono T, Takahashi K, Abe T, Shibahara S, Tamai M. Elevated immunoreactive-adrenomedullin levels in the aqueous humor of patients with uveitis and vitreoretinal disorders. Peptides. 2002;23:1865–8. doi: 10.1016/s0196-9781(02)00145-6. [DOI] [PubMed] [Google Scholar]
- 14.Vingolo EM, Lupo S, Domanico D, Cotesta D, Petramala L, Grenga R, Letizia C. Adrenomedullin plasma concentrations in patients with retinitis pigmentosa. Clin Biochem. 2005;38:735–8. doi: 10.1016/j.clinbiochem.2005.04.007. [DOI] [PubMed] [Google Scholar]
- 15.Fernandez-Sauze S, Delfino C, Mabrouk K, Dussert C, Chinot O, Martin PM, Grisoli F, Ouafik L, Boudouresque F. Effect of adrenomedullin on endothelial cells in the multistep process of angiogenesis: involvement of CRLR/RAMP2 and CRLR/RAMP3 receptors. Int J Cancer. 2004;108:797–804. doi: 10.1002/ijc.11663. [DOI] [PubMed] [Google Scholar]
- 16.Hermansen SE, Lund T, Kalstad T, Ytrehus K, Myrmel T. Adrenomedullin augments the angiogenic potential of late outgrowth endothelial progenitor cells. Am J Physiol Cell Physiol. 2011;300:C783–91. doi: 10.1152/ajpcell.00044.2010. [DOI] [PubMed] [Google Scholar]
- 17.Miyashita K, Itoh H, Sawada N, Fukunaga Y, Sone M, Yamahara K, Yurugi T, Nakao K. Adrenomedullin promotes proliferation and migration of cultured endothelial cells. Hypertens Res. 2003;26(Suppl):S93–8. doi: 10.1291/hypres.26.s93. [DOI] [PubMed] [Google Scholar]
- 18.Sakimoto S, Kidoya H, Kamei M, Naito H, Yamakawa D, Sakaguchi H, Wakabayashi T, Nishida K, Takakura N. An angiogenic role for adrenomedullin in choroidal neovascularization. PLoS ONE. 2013;8:e58096. doi: 10.1371/journal.pone.0058096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kim W, Moon SO, Sung MJ, Kim SH, Lee S, So JN, Park SK. Angiogenic role of adrenomedullin through activation of Akt, mitogen-activated protein kinase, and focal adhesion kinase in endothelial cells. FASEB J. 2003;17:1937–9. doi: 10.1096/fj.02-1209fje. [DOI] [PubMed] [Google Scholar]
- 20.Miyashita K, Itoh H, Sawada N, Fukunaga Y, Sone M, Yamahara K, Yurugi-Kobayashi T, Park K, Nakao K. Adrenomedullin provokes endothelial Akt activation and promotes vascular regeneration both in vitro and in vivo. FEBS Lett. 2003;544:86–92. doi: 10.1016/s0014-5793(03)00484-8. [DOI] [PubMed] [Google Scholar]
- 21.Schwarz N, Renshaw D, Kapas S, Hinson JP. Adrenomedullin increases the expression of calcitonin-like receptor and receptor activity modifying protein 2 mRNA in human microvascular endothelial cells. J Endocrinol. 2006;190:505–14. doi: 10.1677/joe.1.06806. [DOI] [PubMed] [Google Scholar]
- 22.Distler JH, Hirth A, Kurowska-Stolarska M, Gay RE, Gay S, Distler O. Angiogenic and angiostatic factors in the molecular control of angiogenesis. Q J Nucl Med. 2003;47:149–61. [PubMed] [Google Scholar]
- 23.Maki T, Ihara M, Fujita Y, Nambu T, Harada H, Ito H, Nakao K, Tomimoto H, Takahashi R. Angiogenic roles of adrenomedullin through vascular endothelial growth factor induction. Neuroreport. 2011;22:442–7. doi: 10.1097/WNR.0b013e32834757e4. [DOI] [PubMed] [Google Scholar]
- 24.Guidolin D, Albertin G, Spinazzi R, Sorato E, Mascarin A, Cavallo D, Antonello M, Ribatti D. Adrenomedullin stimulates angiogenic response in cultured human vascular endothelial cells: involvement of the vascular endothelial growth factor receptor 2. Peptides. 2013;29:213–23. doi: 10.1016/j.peptides.2008.07.009. [DOI] [PubMed] [Google Scholar]
- 25.Zakareia FA, Alderees AA, Al Regaiy KA, Alrouq FA. Correlation of electroretinography b-wave absolute latency, plasma levels of human basic fibroblast growth factor, vascular endothelial growth factor, soluble fatty acid synthase, and adrenomedullin in diabetic retinopathy. J Diabetes Complications. 2010;24:179–85. doi: 10.1016/j.jdiacomp.2008.12.007. [DOI] [PubMed] [Google Scholar]
- 26.Stitt AW, McGoldrick C, Rice-McCaldin A, McCance DR, Glenn JV, Hsu DK, Liu FT, Thorpe SR, Gardiner TA. Impaired retinal angiogenesis in diabetes: role of advanced glycation end products and galectin-3. Diabetes. 2005;54:785–94. doi: 10.2337/diabetes.54.3.785. [DOI] [PubMed] [Google Scholar]
- 27.Dagher Z, Park YS, Asnaghi V, Hoehn T, Gerhardinger C, Lorenzi M. Studies of rat and human retinas predict a role for the polyol pathway in human diabetic retinopathy. Diabetes. 2004;53:2404–11. doi: 10.2337/diabetes.53.9.2404. [DOI] [PubMed] [Google Scholar]
- 28.Yuan L, Hu J, Luo Y, Liu Q, Li T, Parish CR, Freeman C, Zhu X, Ma W, Hu X, Yu H, Tang S. Upregulation of heparanase in high-glucose-treated endothelial cells promotes endothelial cell migration and proliferation and correlates with Akt and extracellular-signal-regulated kinase phosphorylation. Mol Vis. 2012;18:1684–95. [PMC free article] [PubMed] [Google Scholar]
- 29.Premanand C, Rema M, Sameer MZ, Sujatha M, Balasubramanyam M. Effect of curcumin on proliferation of human retinal endothelial cells under in vitro conditions. Invest Ophthalmol Vis Sci. 2006;47:2179–84. doi: 10.1167/iovs.05-0580. [DOI] [PubMed] [Google Scholar]
- 30.Nikitenko LL, Fox SB, Kehoe S, Rees MC, Bicknell R. Adrenomedullin and tumour angiogenesis. Br J Cancer. 2006;94:1–7. doi: 10.1038/sj.bjc.6602832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Griffioen AW, Molema G. Angiogenesis: potentials for pharmacologic intervention in the treatment of cancer, cardiovascular diseases, and chronic inflammation. Pharmacol Rev. 2000;52:237–68. [PubMed] [Google Scholar]
- 32.Iimuro S, Shindo T, Moriyama N, Amaki T, Niu P, Takeda N, Iwata H, Zhang Y, Ebihara A, Nagai R. Angiogenic effects of adrenomedullin in ischemia and tumor growth. Circ Res. 2004;95:415–23. doi: 10.1161/01.RES.0000138018.61065.d1. [DOI] [PubMed] [Google Scholar]
- 33.DiPietro LA, Nebgen DR, Polverini PJ. Downregulation of endothelial cell thrombospondin 1 enhances in vitro angiogenesis. J Vasc Res. 1994;31:178–85. doi: 10.1159/000319585. [DOI] [PubMed] [Google Scholar]
- 34.Porter S, Clark IM, Kevorkian L, Edwards DR. The ADAMTS metalloproteinases. Biochem J. 2005;386:15–27. doi: 10.1042/BJ20040424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Uno K, Bhutto IA, McLeod DS, Merges C, Lutty GA. Impaired expression of thrombospondin-1 in eyes with age related macular degeneration. Br J Ophthalmol. 2006;90:48–54. doi: 10.1136/bjo.2005.074005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lee NV, Sato M, Annis DS, Loo JA, Wu L, Mosher DF, Iruela-Arispe ML. ADAMTS1 mediates the release of antiangiogenic polypeptides from TSP1 and 2. EMBO J. 2006;25:5270–83. doi: 10.1038/sj.emboj.7601400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Krampert M, Kuenzle S, Thai SN, Lee N, Iruela-Arispe ML, Werner S. ADAMTS1 proteinase is up-regulated in wounded skin and regulates migration of fibroblasts and endothelial cells. J Biol Chem. 2005;280:23844–52. doi: 10.1074/jbc.M412212200. [DOI] [PubMed] [Google Scholar]
- 38.Sunderkötter C, Steinbrink K, Goebeler M, Bhardwaj R, Sorg C. Macrophages and angiogenesis. J Leukoc Biol. 1994;55:410–22. doi: 10.1002/jlb.55.3.410. [DOI] [PubMed] [Google Scholar]
- 39.Lu P, Li L, Kuno K, Wu Y, Baba T, Li YY, Zhang X, Mukaida N. Protective roles of the fractalkine/CX3CL1–CX3CR1 interactions in alkali-induced corneal neovascularization through enhanced antiangiogenic factor expression. J Immunol. 2008;180:4283–91. doi: 10.4049/jimmunol.180.6.4283. [DOI] [PubMed] [Google Scholar]
- 40.Cursiefen C, Masli S, Ng TF, Dana MR, Bornstein P, Lawler J, Streilein JW. Roles of thrombospondin-1 and −2 in regulating corneal and iris angiogenesis. Invest Ophthalmol Vis Sci. 2004;45:1117–24. doi: 10.1167/iovs.03-0940. [DOI] [PubMed] [Google Scholar]
- 41.Graupera M, Potente M. Regulation of angiogenesis by PI3K signaling networks. Exp Cell Res. 2013;319:1348–55. doi: 10.1016/j.yexcr.2013.02.021. [DOI] [PubMed] [Google Scholar]
- 42.Dimmeler S, Zeiher AM. Akt takes center stage in angiogenesis signaling. Circ Res. 2000;86:4–5. doi: 10.1161/01.res.86.1.4. [DOI] [PubMed] [Google Scholar]
- 43.Abid MR, Guo S, Minami T, Spokes KC, Ueki K, Skurk C, Walsh K, Aird WC. Vascular endothelial growth factor activates PI3K/Akt/forkhead signaling in endothelial cells. Arterioscler Thromb Vasc Biol. 2004;24:294–300. doi: 10.1161/01.ATV.0000110502.10593.06. [DOI] [PubMed] [Google Scholar]