

Abstract
Topical antigen application is a focus of current vaccine research. This immunization route mimics natural antigen exposure across a barrier tissue and generates T cells imprinted for skin-selective homing. Soluble antigens introduced through this route require cross-presentation by DC to generate CD8 T cell responses. Here we have explored the relative contribution of various skin-derived DC subsets to cross-priming and skin-selective imprinting. In our model, DC acquire soluble Ag in vivo from immunized murine skin for cross-presentation to naïve CD8 T cells ex vivo. We find CD11b+ migratory DC to be the relevant cross-priming DC in this model. Both Langerin+ and Langerin- CD11b+ migratory DC can cross-present antigen in our system, but only the Langerin+ subset can induce expression of the skin-selective addressin E-selectin ligand. Thus, the CD11b+ Langerin+ migratory DC population, comprised primarily of Langerhans cells, both cross-primes naïve CD8 T cells and imprints them with skin-homing capabilities.
Introduction
Dendritic cells (DC) are the primary antigen (Ag)-presenting cells that initiate T cell responses. DC were first recognized for presenting exogenous peptides to CD4 T cells via MHCII, and endogenous peptides to CD8 T cells via MHCI (reviewed in [1]). Cross-presentation, a more recently recognized DC function, is the presentation of exogenous peptides to CD8 T cells via MHCI (reviewed in [2]). Cross-presentation is required to initiate responses against tumor cells and intracellular pathogens that do not directly infect DC.
DC subsets can be distinguished by surface markers and by their positioning within tissues. DC within the skin-draining lymph nodes (sdLN) include plasmacytoid DC, LN-resident (classical) DC and migratory DC, which migrate from skin to sdLN. Classical DC are MHCIIintCD11chi and contain CD8α+CD11b- and CD8α-CD11b+ subsets [3]. Migratory DC can transport Ag from skin to sdLN via afferent lymphatics [4], [5], are MHCIIhiCD11cint, and require functional CCR7 expression to enter sdLN. This population contains CD11b-CD103+ and CD11b+CD103- subsets. Langerin (CD207) is expressed by all epidermal migratory DC (i.e. Langerhans cells, LC) and some dermal migratory DC [6], [7]. Despite common Langerin expression, LC and Lang+ dermal DC are functionally and developmentally distinct subsets [8].
CD8α+ classical DC are considered the primary subset for cross-priming naïve CD8 T cells [2], [9] and may possess specialized intracellular machinery for processing and presenting exogenous Ag on MHCI [10]. Recent studies suggest that CD103+ migratory DC also cross-present Ag [11]. However, some of these studies used viruses that may directly infect some DC [12], so these findings may be attributable to classical MHCI presentation of endogenous Ag.
DC can imprint naïve T cells to express homing molecules that direct the primed T cells to preferentially enter certain barrier tissues, as reviewed in [13], [14]. For example, T cells in peripheral blood use the carbohydrate ligand of E-selectin (E-lig, or CLA in humans) to enter skin and integrin α4β7 to enter intestinal tissues [13]. Prior work showed that peptide-pulsed DC from sdLN or Peyer's patches can imprint CD8 T cells to express E-lig or α4β7, respectively [15].
We set out to more clearly define the in vivo DC subsets that cross-present cutaneous soluble Ag and/or imprint naïve CD8 T cells with skin-homing profiles. We used a murine in vivo system in which DC acquire Ag from inflamed skin. We isolated these “Ag-charged” DC from the sdLN of immunized mice and tested their ability to cross-prime Ag-specific naïve CD8 T cells ex vivo. This allowed us to test the contribution of various DC subsets to cross-priming and tissue-selective imprinting.
Materials and Methods
Mice
C57Bl/6 CD45.2 mice were purchased from Charles River Labs (Wilmington, MA). Lang-DTREGFP (Lang-DTR) and Lang-EGFP mice were a generous gift from Bernard Malissen, Centre d'immunologie de Marseille Luminy [16]. CCR7-/- [17] and CD45.1 OT-I [18] mice were from our colony, the founders obtained from Jackson Labs. (TCRα-/- OT-I T cells were used for some experiments, but no differences in proliferation were seen with respect to TCRα+ OT-I T cells.)
Topical Skin Immunization
Topical immunization of ear skin was performed as described in [19], [20]. Briefly, the stratum corneum on each side of each ear was gently stripped with ten applications of adhesive tape (Scotch matte finish magic tape, 3 M), taking care not to break the skin or cause bleeding. To remove cutaneous lipids that would repel Ag in aqueous solution, 25 µl of acetone was spread over each ear. After evaporation of the acetone, 25 µl of an aqueous mixture containing 1 mg/ml cholera toxin (CT) adjuvant (List Biological Labs, Campbell, CA) was applied to each ear and uniformly spread with a small paint brush. Control mice received only the CT adjuvant, while experimental mice also received 50 µl of an aqueous mixture containing 100 mg/ml ovalbumin (OVA) Ag (Sigma-Aldrich product #A5503) on each ear.
Treatment of Lang-DTR mice with Diphtheria Toxin
For some cocultures (as indicated), WT or Lang-DTR mice were treated with diphtheria toxin (DT) (List Biological Labs, Campbell, CA). Mice were injected intraperitoneally with 1 µg DT in 100 µl PBS. Mice were treated one day preceding immunization and one day after immunization.
Dendritic Cell Isolation
SdLN were harvested from immunized mice, then disrupted between frosted microscope slides and filtered through 80 µm mesh. Remaining solid stroma was incubated for 30 min at 37 C with 1 mg/ml collagenase (Sigma). Digested product was filtered through mesh and added to the rest of the LN prep prior to washing and counting.
Dendritic Cell Sorting
DC were enriched from the sdLN prep after exclusion of T and B cells with anti-B220 (clone RA3.3A1/6.1, ATCC hybridoma supt) and mThy-1.2 Ab (BioXCell, West Lebanon, NH) followed by incubation with mαr Igκ microbeads (Miltenyi Biotec, Auburn, CA) and sorting on an AutoMACS Separator. CD8α+ DC selection was done using the CD8+ dendritic cell isolation kit, mouse (Miltenyi). Isolation of individual DC subsets was performed on a BD FACSARIA (Becton Dickinson, San Jose, CA).
T Cell Preparation
Spleens were harvested from CD45.1 OT-I (or OT-I TCRα-/-) mice for single cell suspensions. RBC were lysed and remaining cells washed and loaded with CFSE. CD8+ T cell selection was performed using “CD8+ T cell isolation kit II, mouse” (Miltenyi).
Coculture
After sorting, isolated populations were resuspended in 5 ml RPMI+10% FBS and a sample taken to identify DC or CD8 T cells by FACS analysis. A known number of 5 µm beads was added for accurate counting. DC and T cells were plated for co-culture in 96-well round-bottom plates. A 1∶1 DC:T cell ratio was found to provide the most reproducible T cell proliferation and was maintained for all co-cultures. This high ratio was likely required due to the relative rarity of DC carrying in vivo-acquired Ag. Co-cultures were incubated for 6 days and cells stained for FACS analysis.
In vivo Transfers
CD45.1 OT-I spleen and pLN were harvested and single cell suspensions prepared. Red blood cells were lysed and remaining cells were washed and loaded with CFSE. After counting, approximately 1.5×107 T cells were retro-orbitally injected into anesthetized mice. Mice were immunized on ear skin (as described above) and LNs were harvested and analyzed for T cell proliferation five days later. WT and Lang-DTR mice were used as recipients. DT-treated mice were injected with DT one day before and one day after T cell transfer. Timeline: day -2, first DTX treatment; day -1, OT-I cells transferred IV to recipients; day 0, ear skin immunized and second DTX treatment given; day 5, skin-draining LN harvested.
Flow Cytometry
Directly conjugated mAbs were purchased from eBioscience (La Jolla, CA) or BD Pharmingen (San Jose, CA). Flow cytometry was performed on a BD FACS Canto (Becton Dickinson) and analyzed by FlowJo software version 8.8.6 (Treestar, Inc., Stanford, CA).
Statistics
All statistics were performed using one-tailed Mann-Whitney U-tests using Prism software version 5.0a (GraphPad, Inc., La Jolla, CA).
Ethics Statement
This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Institutional Animal Care and Use Committee of Harvard Medical School (Animal Welfare Assurance of Compliance number: A3431-01). All immunizations were performed under ketamine and xylazine anesthesia and all efforts were made to minimize suffering.
Results
Measuring functionality of individual DC subsets from skin
We set out to examine CD8 T cell cross-priming after the cutaneous introduction of soluble Ag. Our approach involves two stages, the first in vivo and the second ex vivo (Fig S1). C57Bl/6 wildtype (WT) mice (or genetically-modified mice on the C57Bl/6 background) were immunized with OVA protein on ear skin along with cholera toxin (CT) adjuvant. CT was chosen because its properties as an adjuvant suggest that it is a promising candidate for topical vaccination of human patients [21]. After immunization, “Ag-charged” DC were isolated from cervical LN, which are a primary sdLN downstream of the ear skin. At the same time, splenic CD8 T cells were isolated from naive OT-I mice, which express a transgenic TCR specific for the H2-Kb-restricted peptide OVA257-264. The DC-enriched sdLN cells and OT-I T cells were then co-cultured ex vivo.
The sole source of Ag in our experimental co-cultures is the OVA protein that was topically applied several days earlier to the DC donor mice. We also established control co-cultures in which exogenous OVA was added directly to culture wells, to confirm DC viability and functionality. Fig 1 shows that DC from sdLN isolated four days post-immunization yielded the maximal OT-I proliferative response.
Figure 1. Time course for peak day of sdLN DC harvest after topical immunization for OT-I proliferation.
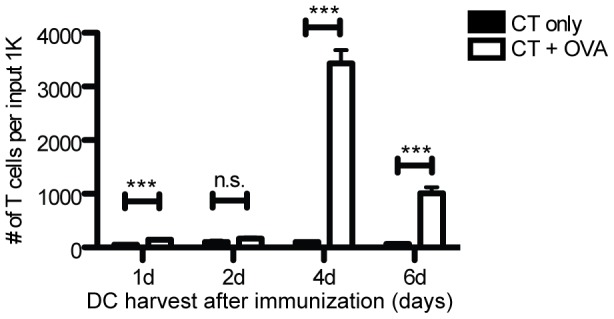
DCs harvested from sdLN various days after topical ear skin immunization and co-cultured with CFSE-labeled naïve OT-I cells. Black bars: CT (adjuvant) only. White bars: CT + OVA protein. CD45.1+ CD3+ CD8+ cells were gated for assessment of CFSE loss. sdLN from 3-4 mice per condition were pooled before DC isolation. Results are from two (1d, 6d) or three (2d, 4d) independent experiments. Proliferation measured at d6 of ex vivo culture. One-tailed Mann-Whitney p values shown. *p<.05; ***p<.0001; n.s. = not significant.
The migratory DC subset is essential for CD8 T cell proliferation
To determine whether Lang-expressing DC are necessary for cross-presentation, we used Lang-DTR-EGFP (Lang-DTR) mice. Diphtheria toxin (DT) receptor is knocked into the Langerin locus, and treatment with DT leads to selective loss of Lang+ DC, including LC, within 24 hours [7]. WT or Lang-DTR mice were treated with DT and immunized on ear skin. We found Lang-depleted DC to be only ∼50% as efficient as WT DC at stimulating OT-I proliferation in our 1∶1 DC/T cell co-cultures (Fig 2A, left). The defect was restricted to Ag acquired in vivo, as both populations were able to cross-present exogenous Ag added to the culture wells ex vivo (Fig 2A, right).
Figure 2. Migratory DCs are essential for CD8+ T cell proliferation after topical immunization.

A: WT and Lang-DTR mice were topically immunized with CT + OVA protein or CT alone. Mice were injected with diphtheria toxin (DT) five and three days prior to cell isolation. On day 4 post-immunization, sdLN cells were isolated, enriched for DCs, and co-cultured with CFSE-labeled OT-I T cells. sdLN from 4–5 mice per condition were pooled before DC isolation. N = 4 experiments. B: CD8α+ DCs were isolated from immunized WT mice using an AutoMACS kit. Unsorted DCs were treated as in A. DCs were cultured with CFSE-labeled OT-I T cells. sdLN from 4–5 mice per condition were pooled before DC isolation. N = 3 experiments. C: WT and CCR7-/- mice were immunized and DC harvested on d4 post-immunization and cultured with CFSE-labeled OT-I cells. sdLN from 4–5 mice per condition pooled before DC isolation. N = 3 experiments. For all experiments shown, T cell proliferation was analyzed on day 6 of ex vivo culture. Proliferated T cells per 1000 input T cells is depicted. Flow cytometry plots were gated on CD45.1+ CD3+ CD8+ cells. Adj = adjuvant. Ag = antigen. Tx = treatment. Left panels: The only Ag present in co-culture wells was that carried by DC from immunized mice. Right panels: Exogenous OVA protein was directly added to “positive control” wells. One-tailed Mann-Whitney p values shown. *p<.01; ***p<.0001; n.s. = not significant.
We next focused on the CD8α+ DC subset. Interestingly, the capability to cross-present in vivo-acquired Ag did not reside within this population (Fig 2B, left). CD8α+ DCs could, however, cross-present exogenous OVA (Fig 2B, right).
We next assessed the cross-presentation ability of classical DC in the absence of migratory DC. CCR7-/- mice lack migratory DC in sdLN, as CCR7 is required for DC migration from skin [17]. SdLN DC from CCR7-/- mice were unable to cross-prime OT-I cells (Fig 2C, left). CCR7-/- sdLN DC could cross-present exogenous OVA (Fig 2C, right).
These data, if considered by themselves, suggest that migratory DC are required in the LN for direct Ag presentation to T cells or to “license” CD8α+ DC [2]. To distinguish between these possibilities, we asked whether migratory DC from sdLN could independently cross-present Ag acquired in vivo.
The ability to cross-prime naïve CD8 T cells with in vivo-acquired Ag resides within the migratory DC subset
We used fluorescent cell sorting to separate the MHCIIhiCD11cint migratory DCs into CD11b+ CD103- and CD11b- CD103+ subsets (Fig S2A). We also isolated MHCIIintCD11chi classical DC (which contain the CD8α+ DC population) for comparison with migratory DC. Surprisingly, we found that CD11b+ migratory DC were the only subset to induce appreciable T cell proliferation in co-culture; CD103+ DC and classical DC stimulated negligible proliferation (Fig 3A). All DC populations could cross-present exogenous OVA (Fig 3B), proving them to remain viable and functional after sorting.
Figure 3. Cross-priming ability resides within the CD11b+ MHCIIhi subset.

A: Sorted DCs from immunized mice co-cultured with CFSE-labeled OT-I T cells. The only Ag present in co-culture wells was that carried by DC from immunized mice. Mice were immunized four days before lymph node harvest. N = 3 experiments. B: “Positive control” for 3A - exogenous OVA protein added to wells to confirm DC viability and functionality post-sorting. N = 3 experiments. C: Mice were immunized on ear skin four days before lymph node harvest as previously described, except LPS was used as adjuvant instead of cholera toxin. DCs were sorted and cultured as in 3A. N = 3 experiments. D: Sorted DCs from immunized mice co-cultured with CFSE-labeled OT-I T cells. Mice were immunized two days before lymph node harvest, instead of 4 days. N = 2 experiments. E: CD11b+ migratory DCs were subdivided into Lang+ and Lang- subsets. Sorted DCs from mice immunized four days before lymph node harvest were co-cultured with CFSE-labeled OT-I T cells. N = 4 experiments. F: “Positive control” for 3E. N = 4 experiments. For all experiments shown, sdLN from 10 immunized mice were pooled before DC sorting. T cell proliferation was analyzed on d6 of ex vivo culture with sdLN DC isolated on d4 after immunization (except for 3D). The number of proliferated T cells per 1000 input T cells is depicted. Flow cytometry plots were gated on CD45.1+CD3+CD8+ cells. One-tailed Mann-Whitney p values shown. *p<.05; **p<.001; ***p<.0001; n.s. = not significant.
This unexpected finding was not specific to the CT adjuvant, as CD11b+ migratory DC remained the sole cross-presenting population when LPS was used as an alternative adjuvant (Fig 3C). Furthermore, this finding was not specific to the time point chosen for harvesting DC after immunization (day 4); we performed identical experiments at day 2 and found that CD11b+ migratory DC remained the only subset to induce appreciable T cell proliferation in co-culture (albeit the proliferation was somewhat less at this earlier time point) (Fig 3D).
We next used Lang-EGFP mice [16] as the DC source to allow subdivision of the CD11b+ migratory DC population into Lang+ and Lang- subsets by fluorescent sorting (Fig S2B). Both populations stimulated significant T cell proliferation (Fig 3E). Again, all DC populations could cross-present exogenous OVA added to the culture wells (Fig 3F), proving them to remain viable and functional after sorting.
CD11b+ Langerin+ DC are responsible for the majority of E-lig imprinting on T cells
DC are capable of imprinting primed T cells to express tissue-selective homing markers [13], [14], [22]. E-lig is required for entry of T cells into skin from blood [13]. Early work showed that peptide-pulsed DC from sdLN could stimulate E-lig expression on CD8 T cells [15], but direct exposure of DC to peptide Ag in vitro does not maintain key components of in vivo Ag transport and processing.
Unsorted DC induced E-lig expression on proliferating CD8 T cells, and sorted CD11b+ migratory DC retained this function (Fig 4A, B). However, after sorting the Lang+ and Lang- subsets, we found that the ability to induce E-lig expression resided largely within the Lang+ population (Fig 4C, D). This is informative, as previous work suggested that activated CD8 T cells express E-lig by default unless they receive gut-specific imprinting signals [15], [23].
Figure 4. CD11b+Langerin+MHCIIhi DCs induce E-lig.

A: Representative example of E-lig expression on CD8 T cells after co-culture with unsorted DCs or sorted CD11b+ MHCIIhi DCs. Control was stained in the presence of EDTA (which disrupts Ca++-dependent E-selectin binding) to determine non-specific binding of the E-selectin-Ig chimera. Flow cytometry plots are gated on CFSE-low CD8+ T cells. Vertical axis: side scatter. Plots representative of 3 different experiments of 5–8 wells each (depending on DC yield). 3433 cells shown per panel. B: E-lig expression depicted as the percentage of proliferated T cells expressing high levels of E-lig. E-lighi expression was observed only on proliferated T cells, so undivided T cells are not included in the calculation. Data from co-cultures with unsorted DCs or sorted CD11b+ MHCIIhi DCs are compared within a given experiment. sdLN from 10 immunized mice were pooled before DC sorting. N = 3 experiments. One-tailed Mann-Whitney p values shown. n.s. = not significant. C: Representative example of E-lig expression on CD8 T cells after co-culture with CD11b+ Lang+ or CD11b+ Lang- MHCIIhi DCs. Control stained as in A. Flow cytometry plots are gated on CFSE-low CD8+ T cells. Vertical axis: side scatter. Plots are representative of samples from 4 different experiments of 3–8 wells each (depending on DC yield). 1623 cells shown per panel. D: E-lig expression depicted as the percentage of proliferated OT-I T cells that are E-lighi as in B, Data from co-cultures with unsorted DCs and sorted Lang+ or Lang- CD11b+ MHCIIhi DCs compared within a given experiment. sdLN from 10 immunized mice were pooled before DC sorting. N = 4 experiments. One-tailed Mann-Whitney p values shown. *p<.01; ***p<.0001.
In vivo verification of ex vivo findings
Our in vivo/ex vivo assay suggested that CD11b+Lang+ DC are the key subset mediating E-lig induction on naïve CD8 T cells after topical immunization. This result implied that a mouse lacking Lang+ DC would be impaired in its ability to produce E-lig+ T cells after topical immunization. To test this hypothesis, naïve OT-I cells were transferred directly into DTX-treated Lang-DTR mice for comparison to other recipient mice bearing normal numbers of Lang+ DC. In the absence of Lang+ DC, we found the induction of E-lig expression on OT-I cells to be reduced dramatically (Fig 5).
Figure 5. Decreased proliferation and E-lig induction for OT-I cells adoptively transferred into Lang+ DC-depleted mice.
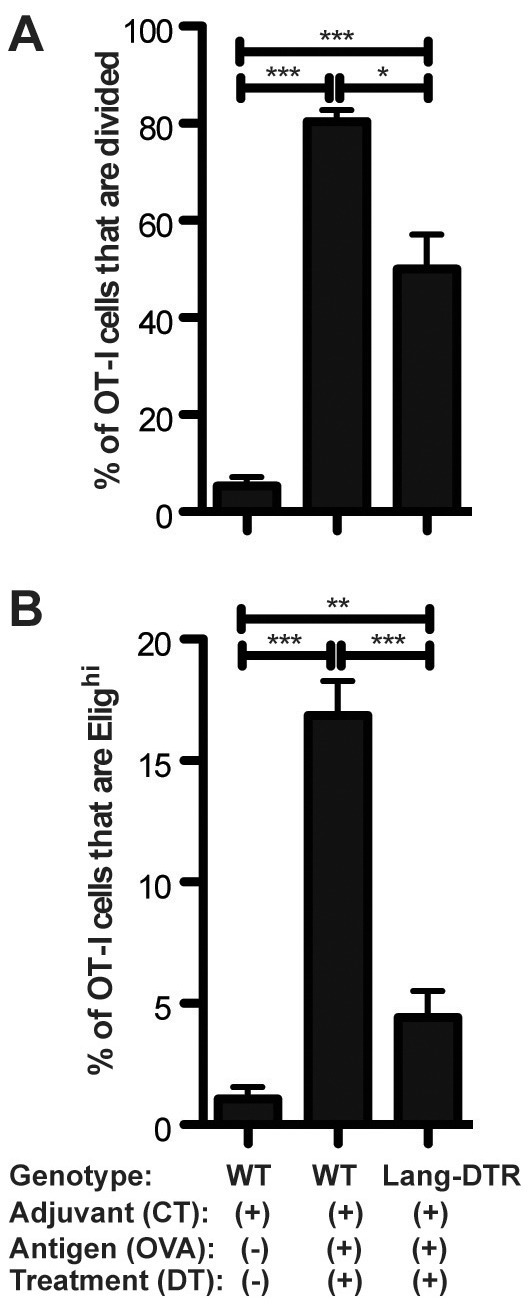
CFSE-labeled splenocytes from CD45.1+ OT-I donor mice were injected iv into recipient WT and Lang-DTR mice. On the following day, recipients were topically immunized with CT+OVA protein or CT alone on the ear skin, using the same immunization techniques as in our in vivo/ex vivo assays. All mice were treated with twice diphtheria toxin. Timeline: day -2, first DTX treatment; day -1, OT-I cells transferred IV to recipients; day 0, ear skin immunized and second DTX treatment given; day 5, skin-draining LN harvested. A: Proliferation depicted as the percentage of total OT-I T cells that are CFSE low. B: E-lig expression depicted as the percentage of total. N = 3 experiments of 4–5 mice per group. For all experiments shown, sdLN cells were isolated and gated on CD45.1+CD3+CD8+ cells. One-tailed Mann-Whitney p values shown. *p<.05; **p<.001; ***p<.0001; n.s. = not significant.
Discussion
There are at least three general mechanisms through which DC can obtain cutaneous Ag from skin: 1) migratory DC transport Ag directly to the sdLN [24]; 2) LN-resident classical DC obtain soluble Ag within the sdLN from afferent lymph [25]; or 3) LN-resident classical DC obtain Ag within the sdLN by transfer from migratory DC [4]. The third mechanism is currently believed to generate effective cross-presentation in vivo (reviewed in [2]).
It is important to understand the mechanism by which immunization occurs through injured skin as investigated in this study. One of the most successful immunization procedures in history is Edward Jenner's 18th century inoculation of patients with non-virulent vaccinia virus through skin scarification to protect against smallpox. It has been proven today that the skin scarification method of immunization is many times more effective for this particular virus than more modern inoculation methods such as subcutaneous injection [26]. This suggests that antigen delivery through skin can induce a type of immune reaction different from those induced through other routes.
We have established a model system to study cross-presentation by loading DC with Ag through a tissue-specific immunization route in vivo, and used these “Ag-charged” DC to elicit proliferation and activation of Ag-specific naïve T cells ex vivo. Our approach goes beyond the abilities of many existing models, which remove any physical or mechanistic barriers that might normally prevent a given DC subtype from accessing Ag in vivo. Our approach preserves the loading of DC with Ag within the complex three-dimensional developmental niches of the intact skin and sdLN, which would be difficult if not impossible to accurately reproduce in vitro with current technology. Thus, our approach combines the robustness of a reductionist ex vivo model with the crucial in vivo components of Ag-acquisition by DC and subsequent transport of Ag to the LN. This approach has provided us with significant advantages towards understanding the cells, molecules and processes required for Ag cross-presentation, and the potential for such immunization routes to generate tissue-specific cellular immune responses in human patients.
The question of which skin-resident DC subset is responsible for the cross-presentation of soluble Ag is quite well studied, but far from resolved. Our results demonstrate that CD11b+ migratory DC cross-present soluble protein Ag applied to skin. We found all other DC subsets essentially incapable of cross-priming CD8 T cells with in vivo-acquired Ag. This is surprising, given the association of CD8α+ classical DCs and CD103+ migratory DCs with cross-presentation in other contexts, especially as these DCs are thought to be developmentally related [2], [9], [11]. However, under our co-culture conditions, neither of these subsets was able to cross-prime CD8 T cells. We found that CD11b+ migratory DC cross-prime CD8 T cells directly, not by licensing other DC populations. By using an approach that requires DC to acquire Ag in vivo, we have preserved the physical barriers that may prevent some DC from accessing Ag in vivo. As all DC subsets tested were capable of cross-presenting Ag added directly to culture wells, this finding emphasizes the importance of Ag access, which can only be observed in vivo.
Most interestingly, we also found that not all DC subsets that can cross-prime CD8 T cells are also capable of imprinting them for skin-selective homing (Fig 4). Our data demonstrate that the entire population of CD11b+ migratory DC can cross-present in vivo-acquired Ag, but only the Lang+ subset could effectively imprint the activated CD8 T cells to express E-lig. This finding was confirmed in vivo by demonstrating that adoptively transferred OT-I populations within the skin-draining nodes of hosts lacking Lang+ DC contained dramatically fewer E-lig+ cells.
Some have suggested that imprinting T cells to express homing markers is independent of activation site or tissue of origin [27]. Other work has focused on stromal or LN microenvironment factors [28], [29]. The variation in E-lig imprinting between Lang+ and Lang- CD11b+ migratory DC strongly suggests that skin-selective imprinting requires signals distinct from those that stimulate proliferation. It also implies that imprinting is an instructive process orchestrated by DC, independent of contemporaneous exposure to afferent lymph or LN-derived stroma cells (albeit such factors may influence DC imprinting capabilities prior to ex vivo culture).
The immunophenotype of the population we found capable of both cross-presentation and imprinting is consistent with that of LC (i.e. Lang+ CD11b+ CD103- DC within the MHCIIhi CD11cint migratory DC population). The cross-presentation capability of LC has been controversial, with some showing LC to be unnecessary for CD8 T cell responses [30], [31], and others showing them to be important [21], [32]. Work with LC ablation models has offered mixed results with regard to the role of LC in contact hypersensitivity (reviewed in [33]). The finding that LC transfer HSV Ag to CD8α+ DC that then prime CD8 T cells supports the idea that LC do not directly cross-prime T cells [4]. However, other work has found LC pulsed ex vivo with Ag to be capable of cross-presentation [34], [35]. The present study demonstrates that each DC subset we isolated from sdLN is capable of cross-presentation when pulsed with OVA ex vivo (Figs 3B, 3F). Importantly, however, we have demonstrated that when OVA is applied physiologically, requiring passage of Ag through the skin barrier tissue, only the CD11b+ migratory DCs are able to cross-present. While the LCs are not the only DC subset to cross-present (the Lang- CD11b+ migratory DCs do so as well), they are the only DC subset able to imprint E-lig.
The lack of consensus on LC cross-presentation ability may stem from the use of different Ag under varying immunization conditions. DC subsets that do not typically cross-present Ag may do so when competing DCs are removed. Using LC depletion models to examine the role of LC could thus obscure their true role in vivo, especially if the depletion is chronic. However, acute depletion models are useful for narrowing down DC subsets for further investigation, as we have done in the current study. In addition, it is possible that LC directly present some Ag (e.g. soluble protein Ag), but transfer others (e.g. HSV Ag) to LN-resident DC. Further studies are required to build a complete picture for the role of LC in cross-presentation, but we have demonstrated the ability of LCs to both cross-present and imprint skin-homing markers on CD8 T cells.
Interestingly, LC are often considered to be tolerogenic, as LC internalize self-Ag in the steady state without causing autoimmune responses [36]. LC also arrive late in the sdLN and inefficiently induce CD80 and CD86 [24]. However, it should be noted that peak T cell proliferation occurred in our system when DC were harvested at day 4 after in vivo immunization (Fig 1), the peak day for LC arrival in the sdLN [24]. It is possible that LC tolerogenicity depends on maturation state (reviewed in [37]), as immature LC can migrate to draining LN [38]. Also, as LC were not distinguished from Lang+ dermal DC in many earlier studies, Lang+ dermal DC may cause the tolerogenic effects currently ascribed to LC [11], [39], reviewed in [37].
We found that the ability to cross-present skin-derived exogenous protein Ag under our experimental conditions resided within the CD11b+ subset of CCR7+ migratory DC. Within this subset, only the Lang+ DCs efficiently induced E-lig expression. This is a novel biological finding, supporting the notion that tissue-selective imprinting is an instructive process. It would be enlightening to further explore how different environmental conditions affect the DC's ability to cross-present; this would be key for DC vaccine design, as targeting a vaccine to a specific DC subset would only be useful if that subset cross-presented Ag under the conditions used for vaccine application.
Supporting Information
Description of cell isolation and co-culture setup. DCs: cells were harvested from skin-draining lymph nodes (sdLN) post-topical skin immunization on ears and pooled within each group. Pooled cells were depleted of T and B cells and counted. T cells: spleen from OT-I mouse was harvested and red blood cells were lysed. Cells were labeled with CFSE and enriched for CD8+ T cells with an AutoMACS kit. DCs and T cells were cultured in a 1∶1 ratio. After 6 days of culture, cells were harvested and stained for flow cytometry analysis.
(TIF)
Depiction of dendritic cell subset sorting strategies. A: To isolate DCs for Fig 3A , sdLN cells were first sorted on MHCII vs CD11c expression. CD11chi MHCIIint (“LN-resident”) dendritic cells were sorted as one population. CD11cint MHCIIhi migratory DCs were sorted into CD11b+ and CD103+ sub-populations. B: To isolate DCs for Fig 3D , sdLN were first sorted based on MHCII vs CD11c expression. CD11chi MHCIIint (“LN-resident”) dendritic cells were sorted as one population. CD11cint MHCIIhi migratory DCs were sorted into CD11b+ and CD103+ sub-populations; CD11b+ DCs were further divided based on Langerin-EGFP expression.
(TIF)
Acknowledgments
We thank Drs. Niro Anandasabapathy and Noah Tubo for critical reading of the manuscript and thoughtful scientific advice.
Funding Statement
This work was supported by NIH and R21AI092388 & R21AI097468 (to J.J.C.). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Burgdorf S, Kurts C (2008) Endocytosis mechanisms and the cell biology of antigen presentation. Current Opinion in Immunology 20: 89–95. [DOI] [PubMed] [Google Scholar]
- 2. Kurts C, Robinson BWS, Knolle PA (2010) Cross-priming in health and disease. Nat Rev Immunol 10: 403–414. [DOI] [PubMed] [Google Scholar]
- 3. Shortman K, Shortman K, Liu Y-J, Liu Y-J (2002) Mouse and human dendritic cell subtypes. Nature Reviews Immunology 2: 151. [DOI] [PubMed] [Google Scholar]
- 4. Allan R, Waithman J, Bedoui S, Jones C, Villadangos J, et al. (2006) Migratory Dendritic Cells Transfer Antigen to a Lymph Node-Resident Dendritic Cell Population for Efficient CTL Priming. Immunity 25: 153–162. [DOI] [PubMed] [Google Scholar]
- 5. Lee HK, Zamora M, Linehan MM, Iijima N, Gonzalez D, et al. (2009) Differential roles of migratory and resident DCs in T cell priming after mucosal or skin HSV-1 infection. The Journal of Experimental Medicine 206: 359–370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Bursch LS, Wang L, Igyarto B, Kissenpfennig A, Malissen B, et al. (2007) Identification of a novel population of Langerin+ dendritic cells. Journal of Experimental Medicine 204: 3147–3156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Poulin LF, Henri S, de Bovis B, Devilard E, Kissenpfennig A, et al. (2007) The dermis contains langerin+ dendritic cells that develop and function independently of epidermal Langerhans cells. The Journal of Experimental Medicine 204: 3119–3131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. del Rio M-L, Bernhardt G, Rodriguez-Barbosa J-I, Förster R (2010) Development and functional specialization of CD103+ dendritic cells. Immunol Rev 234: 268–281. [DOI] [PubMed] [Google Scholar]
- 9. Den Haan JMM, Lehar SM, Bevan MJ (2000) Cd8+ but Not Cd8- Dendritic Cells Cross-Prime Cytotoxic T Cells in Vivo. Journal of Experimental Medicine 192: 1685–1696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Dudziak D, Kamphorst AO, Heidkamp GF, Buchholz VR, Trumpfheller C, et al. (2007) Differential Antigen Processing by Dendritic Cell Subsets in Vivo. Science 315: 107–111. [DOI] [PubMed] [Google Scholar]
- 11. Bedoui S, Whitney PG, Waithman J, Eidsmo L, Wakim L, et al. (2009) Cross-presentation of viral and self antigens by skin-derived CD103+ dendritic cells. Nature Immunology 10: 488–495. [DOI] [PubMed] [Google Scholar]
- 12. Goldwich A, Prechtel AT, Mühl-Zürbes P, Pangratz NM, Stössel H, et al. (2011) Herpes simplex virus type I (HSV-1) replicates in mature dendritic cells but can only be transferred in a cell-cell contact-dependent manner. Journal of Leukocyte Biology 89: 973–979. [DOI] [PubMed] [Google Scholar]
- 13. Butcher EC, Williams M, Youngman K, Rott L, Briskin M (1999) Lymphocyte trafficking and regional immunity. Adv Immunol 72: 209–253. [DOI] [PubMed] [Google Scholar]
- 14. Campbell JJ, Butcher EC (2000) Chemokines in tissue-specific and microenvironment-specific lymphocyte homing. Current Opinion in Immunology 12: 336–341. [DOI] [PubMed] [Google Scholar]
- 15. Mora JR (2005) Reciprocal and dynamic control of CD8 T cell homing by dendritic cells from skin- and gut-associated lymphoid tissues. Journal of Experimental Medicine 201: 303–316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Kissenpfennig A, Henri S, Dubois B, Laplace-Builhé C, Perrin P, et al. (2005) Dynamics and function of Langerhans cells in vivo: dermal dendritic cells colonize lymph node areas distinct from slower migrating Langerhans cells. Immunity 22: 643–654. [DOI] [PubMed] [Google Scholar]
- 17. Förster R, Schubel A, Breitfeld D, Kremmer E, Renner-Müller I, et al. (1999) CCR7 coordinates the primary immune response by establishing functional microenvironments in secondary lymphoid organs. Cell 99: 23–33. [DOI] [PubMed] [Google Scholar]
- 18. Hogquist KA, Jameson SC, Heath WR, Howard JL, Bevan MJ, et al. (1994) T cell receptor antagonist peptides induce positive selection. Cell 76: 17–27. [DOI] [PubMed] [Google Scholar]
- 19. Baekkevold ES (2005) A role for CCR4 in development of mature circulating cutaneous T helper memory cell populations. Journal of Experimental Medicine 201: 1045–1051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Campbell JJ, O'Connell DJ, Wurbel M-A (2007) Cutting Edge: Chemokine receptor CCR4 is necessary for antigen-driven cutaneous accumulation of CD4 T cells under physiological conditions. J Immunol 178: 3358–3362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Olvera-Gomez I, Hamilton SE, Xiao Z, Guimaraes CP, Ploegh HL, et al. (2012) Cholera toxin activates nonconventional adjuvant pathways that induce protective CD8 T-cell responses after epicutaneous vaccination. Proc Natl Acad Sci USA 109: 2072–2077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Dudda JC, Simon JC, Martin S (2004) Dendritic cell immunization route determines CD8+ T cell trafficking to inflamed skin: role for tissue microenvironment and dendritic cells in establishment of T cell-homing subsets. J Immunol 172: 857–863. [DOI] [PubMed] [Google Scholar]
- 23. Iwata M, Hirakiyama A, Eshima Y, Kagechika H, Kato C, et al. (2004) Retinoic acid imprints gut-homing specificity on T cells. Immunity 21: 527–538. [DOI] [PubMed] [Google Scholar]
- 24. Shklovskaya E, Roediger B, Fazekas de St Groth B (2008) Epidermal and dermal dendritic cells display differential activation and migratory behavior while sharing the ability to stimulate CD4+ T cell proliferation in vivo. J Immunol 181: 418–430. [DOI] [PubMed] [Google Scholar]
- 25. Anderson AO, Shaw S (2005) Conduit for privileged communications in the lymph node. Immunity 22: 3–5. [DOI] [PubMed] [Google Scholar]
- 26. Liu L, Zhong Q, Tian T, Dubin K, Athale SK, et al. (2010) Epidermal injury and infection during poxvirus immunization is crucial for the generation of highly protective T cell-mediated immunity. Nature Medicine 16: 224–227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Masopust D, Vezys V, Usherwood EJ, Cauley LS, Olson S, et al. (2004) Activated primary and memory CD8 T cells migrate to nonlymphoid tissues regardless of site of activation or tissue of origin. Journal of immunology (Baltimore, Md: 1950) 172: 4875–4882. [DOI] [PubMed] [Google Scholar]
- 28. Hammerschmidt SI, Ahrendt M, Bode U, Wahl B, Kremmer E, et al. (2008) Stromal mesenteric lymph node cells are essential for the generation of gut-homing T cells in vivo. Journal of Experimental Medicine 205: 2483–2490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Edele F, Molenaar R, Gütle D, Dudda JC, Jakob T, et al. (2008) Cutting edge: instructive role of peripheral tissue cells in the imprinting of T cell homing receptor patterns. J Immunol 181: 3745–3749. [DOI] [PubMed] [Google Scholar]
- 30. Bursch LS, Rich BE, Hogquist KA (2009) Langerhans cells are not required for the CD8 T cell response to epidermal self-antigens. Journal of immunology (Baltimore, Md: 1950) 182: 4657–4664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Igyártó BZ, Haley K, Ortner D, Bobr A, Gerami-Nejad M, et al. (2011) Skin-resident murine dendritic cell subsets promote distinct and opposing antigen-specific T helper cell responses. Immunity 35: 260–272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Wang X, Simeoni L, Lindquist JA, Saez-Rodriguez J, Ambach A, et al. (2008) Dynamics of proximal signaling events after TCR/CD8-mediated induction of proliferation or apoptosis in mature CD8+ T cells. J Immunol 180: 6703–6712. [DOI] [PubMed] [Google Scholar]
- 33. Kaplan DH, Kissenpfennig A, Clausen BE (2008) Insights into Langerhans cell function from Langerhans cell ablation models. Eur J Immunol 38: 2369–2376. [DOI] [PubMed] [Google Scholar]
- 34. Stoitzner P, Tripp CH, Eberhart A, Price KM, Jung JY, et al. (2006) Langerhans cells cross-present antigen derived from skin. Proceedings of the National Academy of Sciences of the United States of America 103: 7783–7788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Cao T, Ueno H, Glaser C, Fay JW, Palucka AK, et al. (2007) Both Langerhans cells and interstitial DC cross-present melanoma antigens and efficiently activate antigen-specific CTL. Eur J Immunol 37: 2657–2667. [DOI] [PubMed] [Google Scholar]
- 36. Morelli AE, Hackstein H, Thomson AW (2001) Potential of tolerogenic dendritic cells for transplantation. Seminars in Immunology 13: 323–335. [DOI] [PubMed] [Google Scholar]
- 37. Lutz MB, Döhler A, Azukizawa H (2010) Revisiting the tolerogenicity of epidermal Langerhans cells. Immunology and Cell Biology 88: 381. [DOI] [PubMed] [Google Scholar]
- 38. Geissmann F, Dieu-Nosjean MC, Dezutter C, Valladeau J, Kayal S, et al. (2002) Accumulation of immature Langerhans cells in human lymph nodes draining chronically inflamed skin. The Journal of Experimental Medicine 196: 417–430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Ginhoux F, Collin MP, Bogunovic M, Abel M, Leboeuf M, et al. (2007) Blood-derived dermal langerin+ dendritic cells survey the skin in the steady state. The Journal of Experimental Medicine 204: 3133–3146. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Description of cell isolation and co-culture setup. DCs: cells were harvested from skin-draining lymph nodes (sdLN) post-topical skin immunization on ears and pooled within each group. Pooled cells were depleted of T and B cells and counted. T cells: spleen from OT-I mouse was harvested and red blood cells were lysed. Cells were labeled with CFSE and enriched for CD8+ T cells with an AutoMACS kit. DCs and T cells were cultured in a 1∶1 ratio. After 6 days of culture, cells were harvested and stained for flow cytometry analysis.
(TIF)
Depiction of dendritic cell subset sorting strategies. A: To isolate DCs for Fig 3A , sdLN cells were first sorted on MHCII vs CD11c expression. CD11chi MHCIIint (“LN-resident”) dendritic cells were sorted as one population. CD11cint MHCIIhi migratory DCs were sorted into CD11b+ and CD103+ sub-populations. B: To isolate DCs for Fig 3D , sdLN were first sorted based on MHCII vs CD11c expression. CD11chi MHCIIint (“LN-resident”) dendritic cells were sorted as one population. CD11cint MHCIIhi migratory DCs were sorted into CD11b+ and CD103+ sub-populations; CD11b+ DCs were further divided based on Langerin-EGFP expression.
(TIF)